
Statistical Approach to Incorporating Experimental Variability into a Mathematical Model of the Voltage-Gated Na+ Channel and Human Atrial Action Potential



Abstract
:1. Introduction
2. Materials and Methods
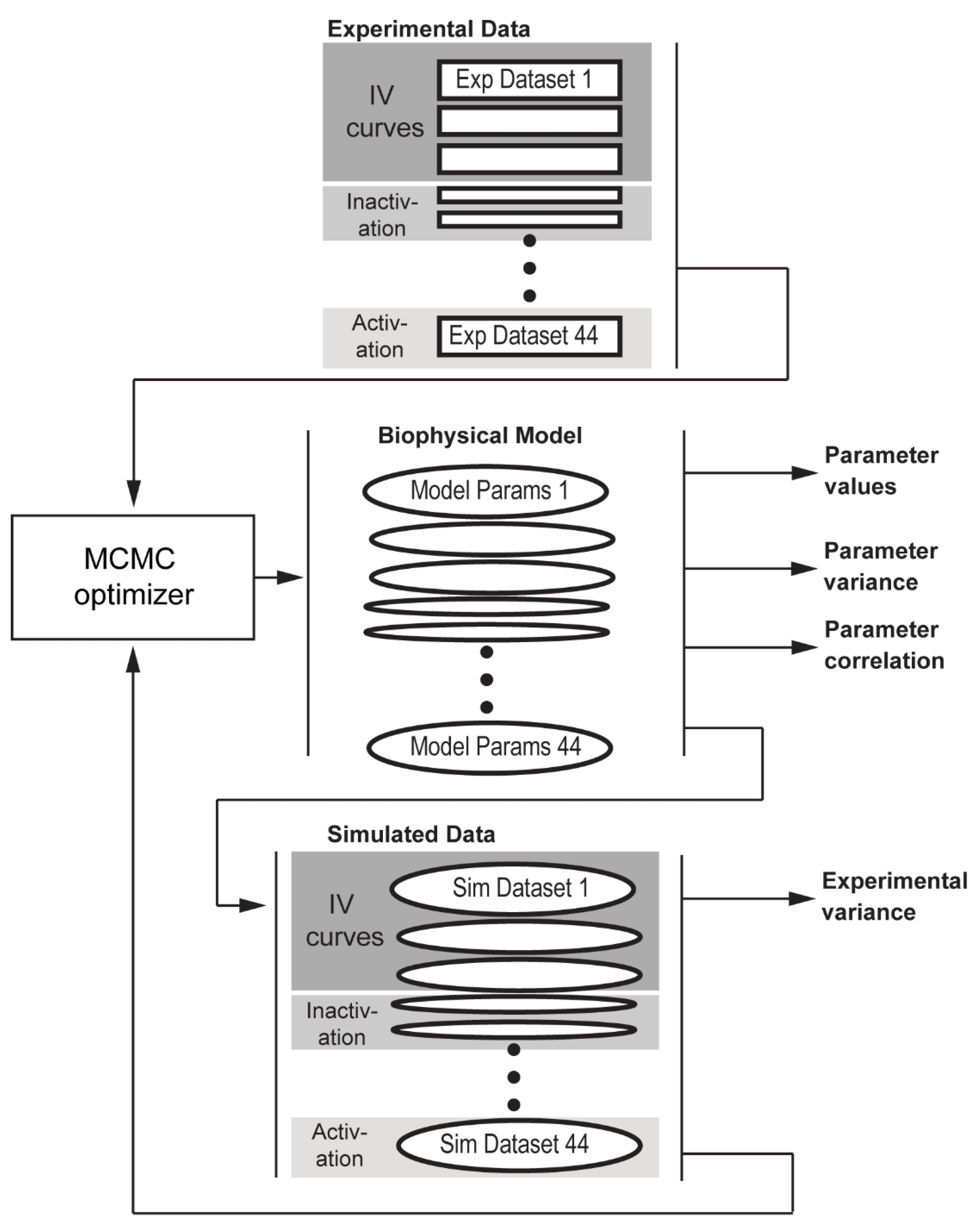
2.1. Statistical Approach to Parameter Fitting for a Biophysical Model of the Voltage-Gated Na+ Channel Using Multiple Datasets
2.2. Curation of Experimental Data from the Literature
2.3. Mathematical Model of the Voltage-Gated Sodium Channel Nav1.5
2.4. Data Normalization
2.5. Bayesian Statistical Model Parameters
2.6. Parameter Estimation
2.7. Code
2.8. Simulations
3. Results
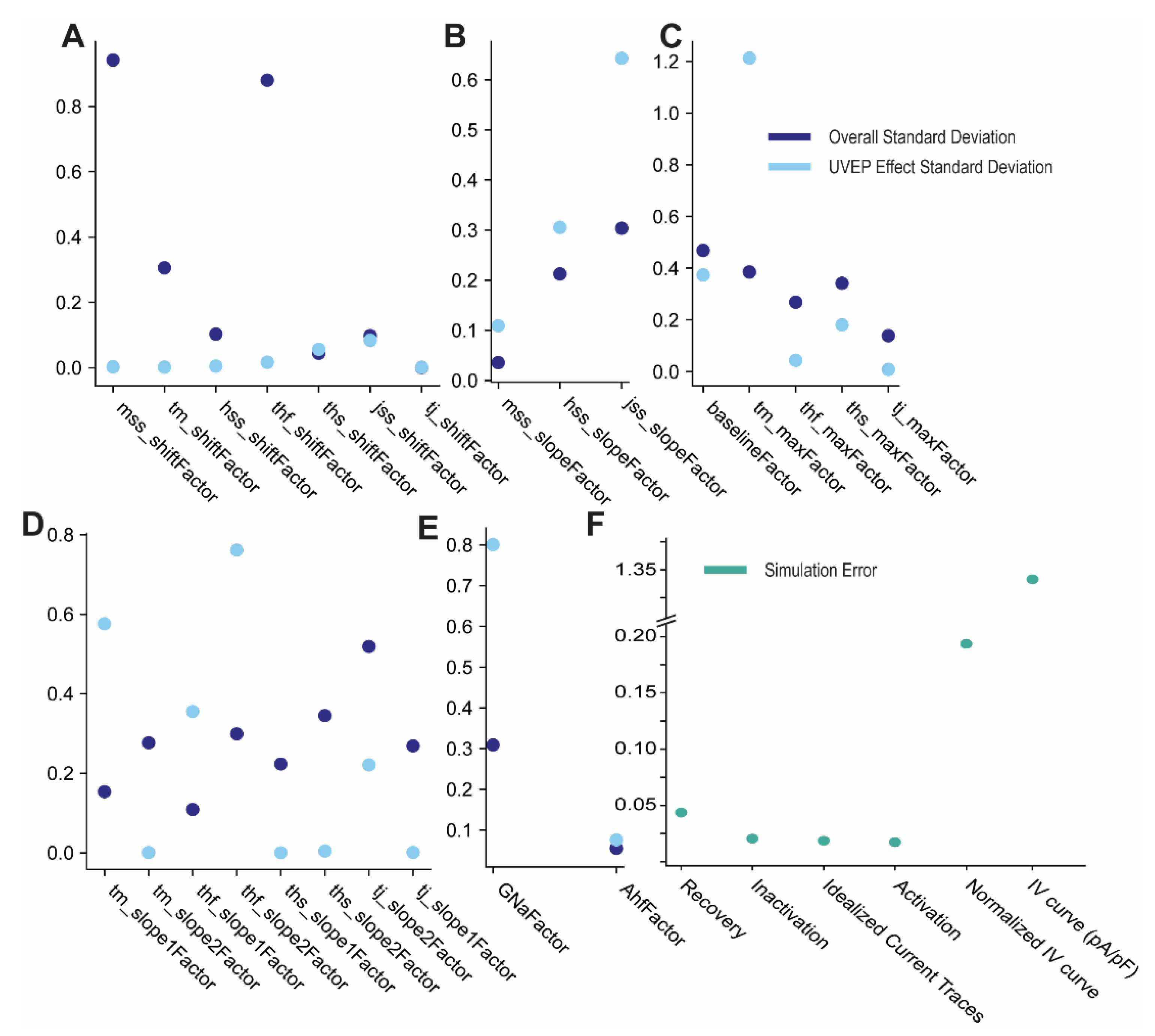
3.1. Estimates of Nav Model Parameters: Simultaneous Fitting of Values Corresponding to Individual Experiments and across the Population
3.2. Generation of a Population of Nav Models Based on Overall Parameter Fits
3.3. Incorporation of Nav Model into a Comprehensive Model of the Human Atrial Action Potential
4. Discussion
4.1. Bayesian Modeling Provides a Natural Way to Incorporate Different Data into One Model
4.2. Variability Is of the Next Frontier for Electrophysiological Modeling
4.3. Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Full Statistical Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Transformation | Model Parameter Factors |
|---|---|---|
| 0 | ||
| 1 | ||
| 2 | ||
| 3 | ||
| 4 | ||
| 5 | ||
| 6 | ||
| 7 | ||
| 8 | ||
| 9 | ||
| 10 | ||
| 11 | ||
| 12 | ||
| 13 | ||
| 14 | ||
| 15 | ||
| 16 | ||
| 17 | ||
| 18 | ||
| 19 | ||
| 20 | ||
| 21 | ||
| 22 | ||
| 23 | ||
| 24 |
Appendix B. Equation Reparameterization
Appendix C. Markov Model
References
- Ang, Y.-S.; Rajamani, S.; Haldar, S.M.; Hüser, J. A New Therapeutic Framework for Atrial Fibrillation Drug Development. Circ. Res. 2020, 127, 184–201. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.D.; Zhu, Z.I.; Yang, P.-C.; Bankston, J.R.; Jeng, M.-T.; Kang, C.; Wang, L.; Bayer, J.D.; Christini, D.J.; Trayanova, N.A.; et al. A Computational Model to Predict the Effects of Class I Anti-Arrhythmic Drugs on Ventricular Rhythms. Sci. Transl. Med. 2011, 3, 98ra83. [Google Scholar] [CrossRef] [Green Version]
- Burashnikov, A.; Antzelevitch, C. Atrial-selective sodium channel block for the treatment of atrial fibrillation. Expert Opin. Emerg. Drugs 2009, 14, 233–249. [Google Scholar] [CrossRef] [PubMed]
- Glynn, P.; Unudurthi, S.D.; Hund, T.J. Mathematical modeling of physiological systems: An essential tool for discovery. Life Sci. 2014, 111, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudy, Y.; Silva, J.R. Computational biology in the study of cardiac ion channels and cell electrophysiology. Q. Rev. Biophys. 2006, 39, 57–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sager, P.T.; Gintant, G.; Turner, J.R.; Pettit, S.; Stockbridge, N. Rechanneling the cardiac proarrhythmia safety paradigm: A meeting report from the Cardiac Safety Research Consortium. Am. Heart J. 2014, 167, 292–300. [Google Scholar] [CrossRef]
- Onal, B.; Gratz, D.; Hund, T.J. Ca 2+/calmodulin kinase II-dependent regulation of atrial myocyte late Na+ current, Ca2+ cycling and excitability: A mathematical modeling study. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H1227–H1239. [Google Scholar] [CrossRef] [Green Version]
- Grandi, E.; Pandit, S.V.; Voigt, N.; Workman, A.J.; Dobrev, D.; Jalife, J.; Bers, D.M. Human Atrial Action Potential and Ca2+ Model: Sinus Rhythm and Chronic Atrial Fibrillation. Circ. Res. 2011, 109, 1055–1066. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, C.; Wiedmann, F.; Voigt, N.; Zhou, X.-B.; Heijman, J.; Lang, S.; Albert, V.; Kallenberger, S.; Ruhparwar, A.; Szabó, G.; et al. Upregulation of K 2P 3.1 K + Current Causes Action Potential Shortening in Patients With Chronic Atrial Fibrillation. Circulation 2015, 132, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Koivumäki, J.T.; Korhonen, T.; Tavi, P. Impact of Sarcoplasmic Reticulum Calcium Release on Calcium Dynamics and Action Potential Morphology in Human Atrial Myocytes: A Computational Study. PLoS Comput. Biol. 2011, 7, e1001067. [Google Scholar] [CrossRef]
- Gong, J.Q.X.; Sobie, E.A. Population-based mechanistic modeling allows for quantitative predictions of drug responses across cell types. npj Syst. Biol. Appl. 2018, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Ni, H.; Morotti, S.; Grandi, E. A Heart for Diversity: Simulating Variability in Cardiac Arrhythmia Research. Front. Physiol. 2018, 9, 958. [Google Scholar] [CrossRef] [PubMed]
- Siekmann, I.; Sneyd, J.; Crampin, E.J. MCMC can detect nonidentifiable models. Biophys. J. 2012, 103, 2275–2286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hines, K.E.; Middendorf, T.R.; Aldrich, R.W. Determination of parameter identifiability in nonlinear biophysical models: A bayesian approach. J. Gen. Physiol. 2014, 143, 401–416. [Google Scholar] [CrossRef] [Green Version]
- Ball, F.G.; Cai, Y.; Kadane, J.B.; O’Hagan, A. Bayesian inference for ion–channel gating mechanisms directly from single–channel recordings, using Markov chain Monte Carlo. Proc. R. Soc. Lond. Ser. A Math. Phys. Eng. Sci. 1999, 455, 2879–2932. [Google Scholar] [CrossRef]
- Siekmann, I.; Wagner, L.E.; Yule, D.; Fox, C.; Bryant, D.; Crampin, E.J.; Sneyd, J. MCMC Estimation of Markov Models for Ion Channels. Biophys. J. 2011, 100, 1919–1929. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, R.H.; Chang, E.T.Y.; Bardenet, R.; de Boer, T.P.; Gavaghan, D.J.; Pathmanathan, P.; Clayton, R.H.; Mirams, G.R. Uncertainty and variability in models of the cardiac action potential: Can we build trustworthy models? J. Mol. Cell. Cardiol. 2016, 96, 49–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, C.L.; Clerx, M.; Beattie, K.A.; Melgari, D.; Hancox, J.C.; Gavaghan, D.J.; Polonchuk, L.; Wang, K.; Mirams, G.R. Rapid Characterization of hERG Channel Kinetics II: Temperature Dependence. Biophys. J. 2019, 117, 2455–2470. [Google Scholar] [CrossRef] [Green Version]
- Lei, C.L.; Clerx, M.; Gavaghan, D.J.; Polonchuk, L.; Mirams, G.R.; Wang, K. Rapid Characterization of hERG Channel Kinetics I: Using an Automated High-Throughput System. Biophys. J. 2019, 117, 2438–2454. [Google Scholar] [CrossRef] [Green Version]
- O’hara, T.; Szló Virá, G.L.; Varró, A.S.; Rudy, Y. Simulation of the Undiseased Human Cardiac Ventricular Action Potential: Model Formulation and Experimental Validation. PLoS Comput. Biol. 2011, 7, e1002061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koval, O.M.; Snyder, J.S.; Wolf, R.M.; Pavlovicz, R.E.; Glynn, P.; Curran, J.; Leymaster, N.D.; Dun, W.; Wright, P.J.; Cardona, N.; et al. Ca2+/calmodulin-dependent protein kinase ii-based regulation of voltage-gated na+ channel in cardiac disease. Circulation 2012, 126, 2084–2094. [Google Scholar] [CrossRef] [Green Version]
- Clancy, C.E.; Rudy, Y. Na + Channel Mutation That Causes Both Brugada and Long-QT Syndrome Phenotypes. Circulation 2002, 105, 1208–1213. [Google Scholar] [CrossRef] [Green Version]
- Nagatomo, T.; Fan, Z.; Ye, B.; Tonkovich, G.S.; January, C.T.; Kyle, J.W.; Makielski, J.C. Temperature dependence of early and late currents in human cardiac wild-type and long Q-T ΔKPQ Na + channels. Am. J. Physiol. Circ. Physiol. 1998, 275, H2016–H2024. [Google Scholar] [CrossRef]
- Michaels, D.C.; Matyas, E.P.; Jalife, J. Mechanisms of sinoartrial pacemaker synchronization: A new hypothesis. Circ. Res. 1987, 61, 704–714. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, M.D.; Gelman, A. The No-U-Turn Sampler: Adaptively Setting Path Lengths in Hamiltonian Monte Carlo. arXiv 2011, arXiv:1111.4246. [Google Scholar]
- Andrieu, C.; Thoms, J. A tutorial on adaptive MCMC. Stat. Comput. 2008, 18, 343–373. [Google Scholar] [CrossRef]
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef]
- McKinney, W. Data structures for statistical computing in python. In Proceedings of the 9th Python in Science Conference, Austin, TX, USA, 28 June–3 July 2010; Volume 445, pp. 51–56. [Google Scholar]
- Van Rossum, G.; Drake, F.L. Python 3 Reference Manual; CreateSpace: Charleston, SC, USA, 2009. [Google Scholar]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental algorithms for scientific computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onal, B.; Gratz, D.; Hund, T. LongQt: A cardiac electrophysiology simulation platform. MethodsX 2016, 3, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Gratz, D.; Onal, B.; Dalic, A.; Hund, T.J. Synchronization of Pacemaking in the Sinoatrial Node: A Mathematical Modeling Study. Front. Phys. 2018, 6, 63. [Google Scholar] [CrossRef]
- Feng, J.; Li, G.R.; Fermini, B.; Nattel, S. Properties of sodium and potassium currents of cultured adult human atrial myocytes. Am. J. Physiol. Circ. Physiol. 1996, 270, H1676–H1686. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Mu, X.; Gong, D.; Jiang, S.; Li, J.; Meng, Q.; Bai, Y.; Liu, Y.; Wang, X.; Tan, X.; et al. Difference of Sodium Currents between Pediatric and Adult Human Atrial Myocytes: Evidence for Developmental Changes of Sodium Channels. Int. J. Biol. Sci. 2011, 7, 708–714. [Google Scholar] [CrossRef] [Green Version]
- Lalevée, N.; Barrère-lemaire, S.; Gautier, P.; Nargeot, J.; Richard, S. Effects of Amiodarone and Dronedarone on Voltage-Dependent Sodium Current in Human Cardiomyocytes. J. Cardiovasc. Electrophysiol. 2003, 14, 885–890. [Google Scholar] [CrossRef]
- Wettwer, E.; Christ, T.; Endig, S.; Rozmaritsa, N.; Matschke, K.; Lynch, J.J.; Pourrier, M.; Gibson, J.K.; Fedida, D.; Knaut, M.; et al. The new antiarrhythmic drug vernakalant: Ex vivo study of human atrial tissue from sinus rhythm and chronic atrial fibrillation. Cardiovasc. Res. 2013, 98, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, Y.; Wasserstrom, J.A.; Furukawa, T.; Jia, H.; Arentzen, C.E.; Hartz, R.S.; Singer, D.H. Characterization of the sodium current in single human atrial myocytes. Circ. Res. 1992, 71, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.; Proebstle, T.; Hombach, V.; Hannekum, A.; Rüdel, R. Characterization of the sodium currents in isolated human cardiocytes. Pflügers Arch. Eur. J. Physiol. 1994, 428, 84–90. [Google Scholar] [CrossRef]
- Nygren, A.; Fiset, C.; Firek, L.; Clark, J.W.; Lindblad, D.S.; Clark, R.B.; Giles, W.R. Mathematical Model of an Adult Human Atrial Cell. Circ. Res. 1998, 82, 63–81. [Google Scholar] [CrossRef] [Green Version]
- Molina, C.E.; Abu-Taha, I.H.; Wang, Q.; Roselló-Díez, E.; Kamler, M.; Nattel, S.; Ravens, U.; Wehrens, X.H.T.; Hove-Madsen, L.; Heijman, J.; et al. Profibrotic, Electrical, and Calcium-Handling Remodeling of the Atria in Heart Failure Patients with and Without Atrial Fibrillation. Front. Physiol. 2018, 9, 1383. [Google Scholar] [CrossRef] [Green Version]
- Poulet, C.; Wettwer, E.; Grunnet, M.; Jespersen, T.; Fabritz, L.; Matschke, K.; Knaut, M.; Ravens, U. Late Sodium Current in Human Atrial Cardiomyocytes from Patients in Sinus Rhythm and Atrial Fibrillation. PLoS ONE 2015, 10, e0131432. [Google Scholar] [CrossRef] [Green Version]
- Le Grand, B.; Le Heuzey, J.Y.; Perier, P.; Peronneau, P.; Lavergne, T.; Hatem, S.; Guize, L. Cellular electrophysiological effects of flecainide on human atrial fibres. Cardiovasc. Res. 1990, 24, 232–238. [Google Scholar] [CrossRef]
- Hansson, A.; Holm, M.; Blomström, P.; Johansson, R.; Lührs, C.; Brandt, J.; Olsson, S.B. Right atrial free wall conduction velocity and degree of anisotropy in patients with stable sinus rhythm studied during open heart surgery. Eur. Heart J. 1998, 19, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Skibsbye, L.; Jespersen, T.; Christ, T.; Maleckar, M.M.; van den Brink, J.; Tavi, P.; Koivumäki, J.T. Refractoriness in human atria: Time and voltage dependence of sodium channel availability. J. Mol. Cell. Cardiol. 2016, 101, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Lei, C.L.; Clerx, M.; Whittaker, D.G.; Gavaghan, D.J.; de Boer, T.P.; Mirams, G.R. Accounting for variability in ion current recordings using a mathematical model of artefacts in voltage-clamp experiments. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2020, 378, 20190348. [Google Scholar] [CrossRef]
- Golowasch, J.; Goldman, M.S.; Abbott, L.F.; Marder, E. Failure of Averaging in the Construction of a Conductance-Based Neuron Model. J. Neurophysiol. 2002, 87, 1129–1131. [Google Scholar] [CrossRef] [PubMed]
- Gilliam, F.R.; Starmer, C.F.; Grant, A.O. Blockade of rabbit atrial sodium channels by lidocaine. Characterization of continuous and frequency-dependent blocking. Circ. Res. 1989, 65, 723–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gratz, D.; Winkle, A.J.; Weinberg, S.H.; Hund, T.J. Statistical Approach to Incorporating Experimental Variability into a Mathematical Model of the Voltage-Gated Na+ Channel and Human Atrial Action Potential. Cells 2021, 10, 1516. https://doi.org/10.3390/cells10061516
Gratz D, Winkle AJ, Weinberg SH, Hund TJ. Statistical Approach to Incorporating Experimental Variability into a Mathematical Model of the Voltage-Gated Na+ Channel and Human Atrial Action Potential. Cells. 2021; 10(6):1516. https://doi.org/10.3390/cells10061516
Chicago/Turabian StyleGratz, Daniel, Alexander J Winkle, Seth H Weinberg, and Thomas J Hund. 2021. "Statistical Approach to Incorporating Experimental Variability into a Mathematical Model of the Voltage-Gated Na+ Channel and Human Atrial Action Potential" Cells 10, no. 6: 1516. https://doi.org/10.3390/cells10061516
APA StyleGratz, D., Winkle, A. J., Weinberg, S. H., & Hund, T. J. (2021). Statistical Approach to Incorporating Experimental Variability into a Mathematical Model of the Voltage-Gated Na+ Channel and Human Atrial Action Potential. Cells, 10(6), 1516. https://doi.org/10.3390/cells10061516