
In Vitro Study of Human Immune Responses to Hyaluronic Acid Hydrogels, Recombinant Spidroins and Human Neural Progenitor Cells of Relevance to Spinal Cord Injury Repair

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Spinal Cord Injury
1.2. Biomaterials in CNS Repair
1.3. Spider Silk
1.4. Hyaluronic Acid Hydrogel
- Taken together, previous and present research indicate that both HA hydrogels and recombinant spidroins are interesting components that may serve as potent base substrates in human SCI repair. In order to develop treatment strategies and apply these biomaterials as part of composite approaches for SCI, it is of value to investigate their biocompatibility and immunogenicity in clinically relevant human cell cultures. Herein, we present an in vitro study observing how human peripheral blood mononuclear cells, hPBMCs, reacted to recombinant miniature spidroins (either the NT2RepCT or VN–NT2RepCT protein, in the form of film and soluble protein) or HA hydrogels (either in its bulk or granular form). We also co-cultured hPBMCs with allogeneic human spinal cord derived NPCs (hNPCs) in the presence of these biomaterials, aiming to mimic an allogeneic host–donor interaction when applying biomaterial as part of a composite experimental intervention. We monitored the human immune cell viability, proliferative response and evaluated different immune cell phenotypes after co-culture using NucleoCounter and flow cytometry.
2. Materials and Methods
2.1. Ethical Considerations
2.2. Isolation and Culture of Human Spinal Cord Derived Neural Progenitor Cells
2.3. Artificial Spidroins
2.4. Bulk and Granular HA Hydrogel Fabrication
2.5. hNPC Encapsulation in Bulk and Granular HA Hydrogels
2.6. hPBMC Preparation
2.7. Viability Test and Cell Count
2.8. hPBMC Response to Biomaterials and hNPCs
2.8.1. Short-Term (15–18 h) Co-Culture and Subpopulation Activation Assay
2.8.2. Long-Term (5 d) Co-Culture and Proliferation Assay
2.9. Flow Cytometry, Gating Strategies and Data Analysis
2.10. Limolus Amebocyte Lyste Test
2.11. Statistics
3. Results
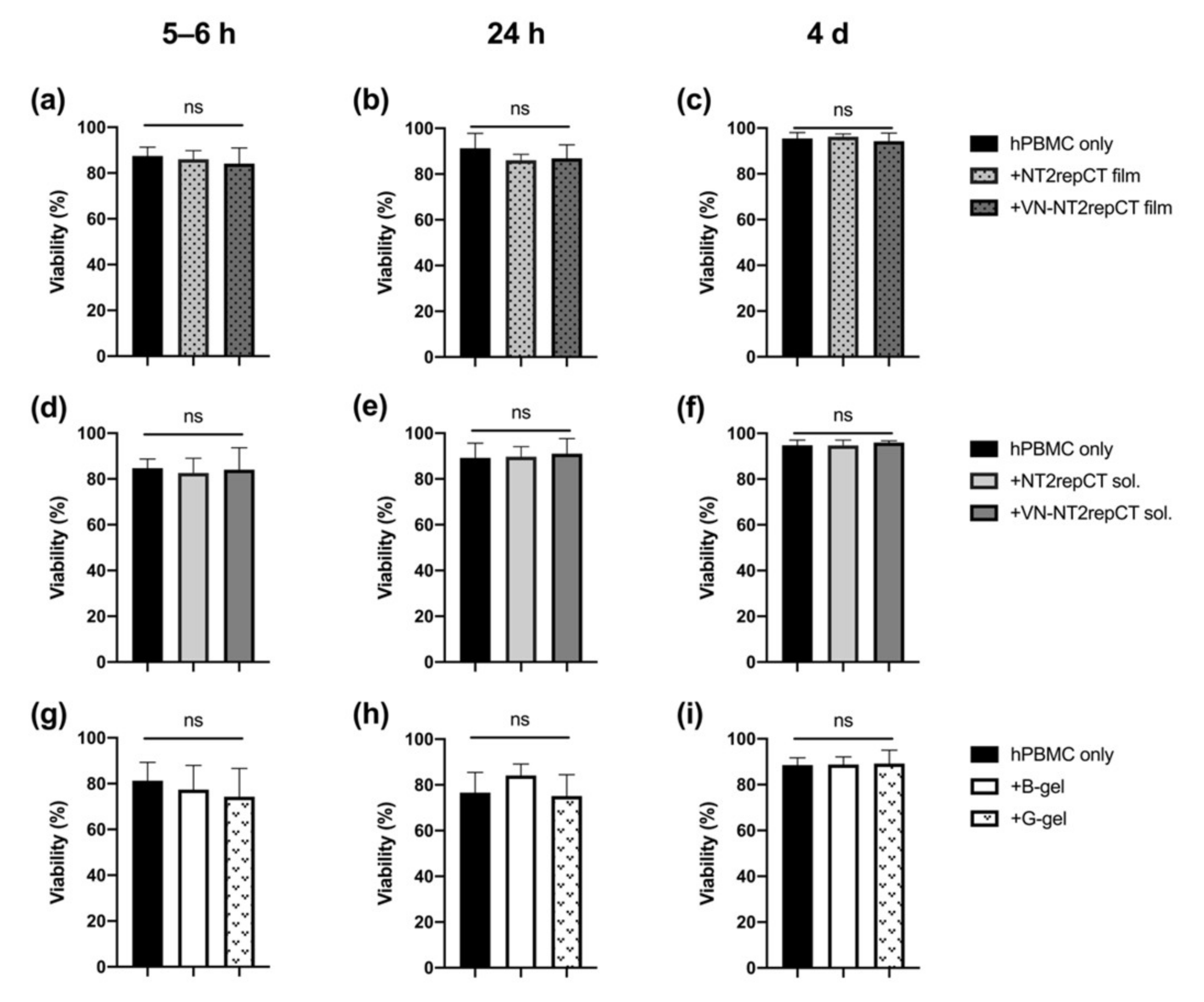
3.1. Spidroins or HA Hydrogel Did Not Compromise the Viability of hPBMCs
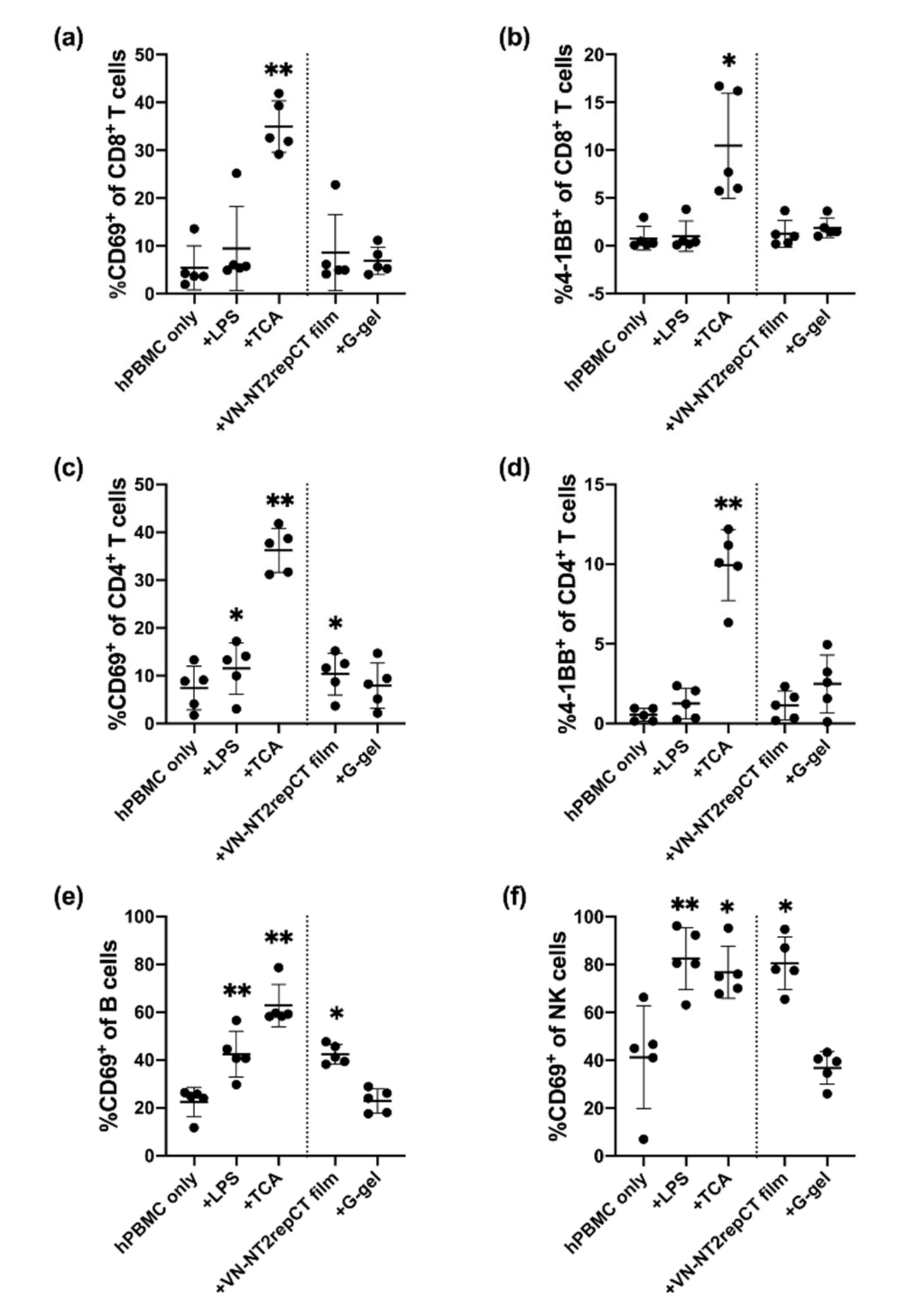
3.2. Spidroin, but Not HA Hydrogel, Activated Immune Cells In Vitro
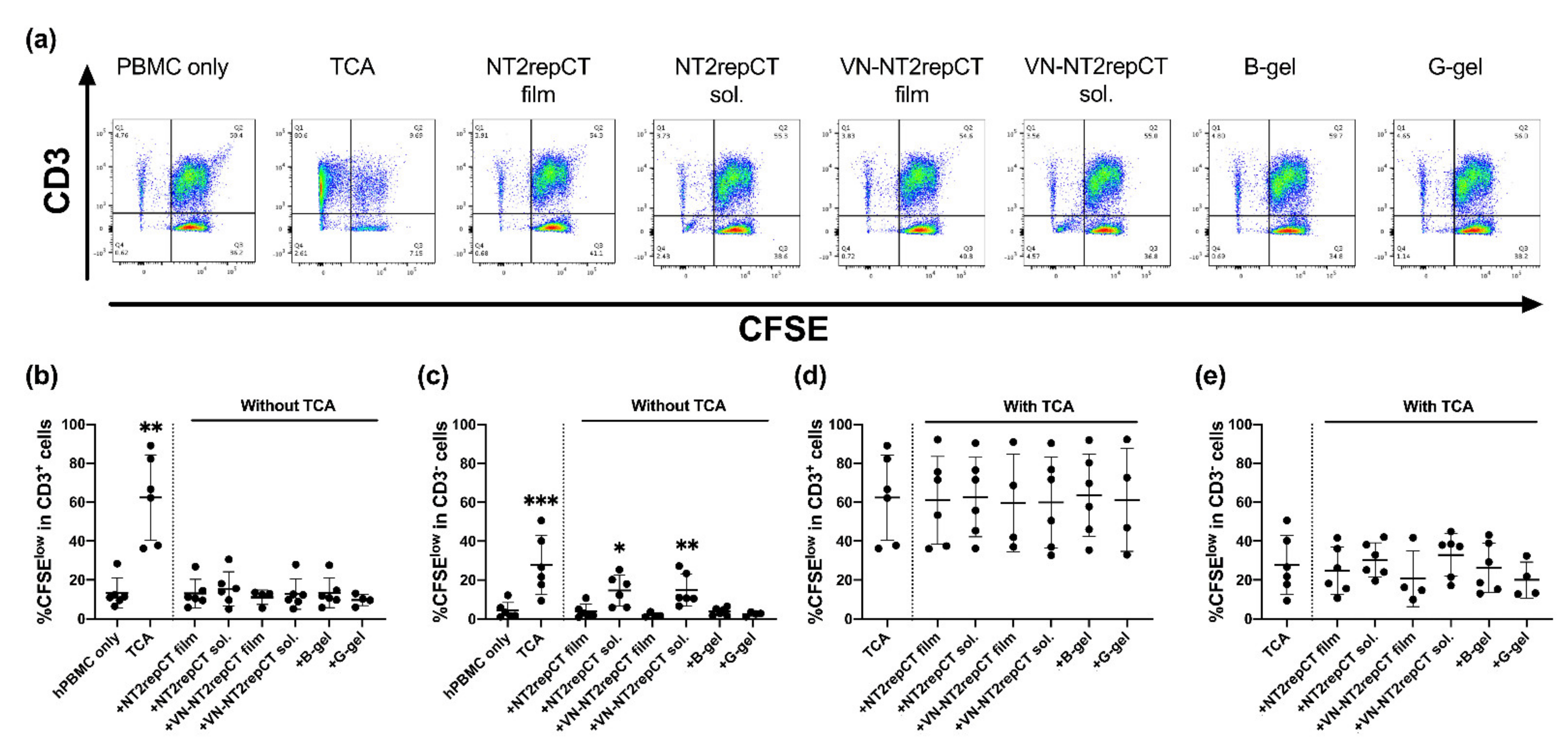
3.3. Soluble Spidroins Stimulated the Proliferation of hPBMCs In Vitro
3.4. No Observed Biomaterial Immunomodulatory Properties
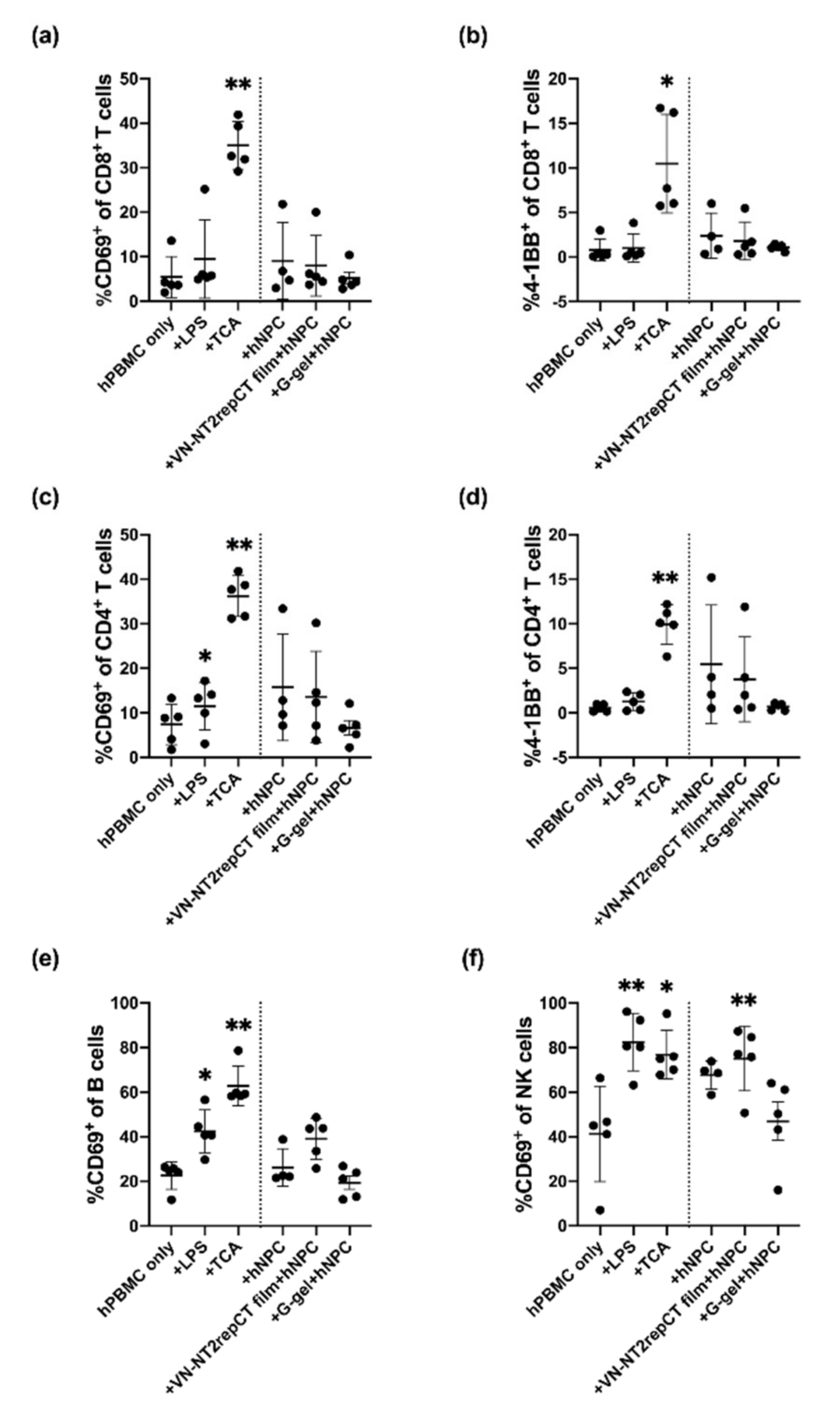
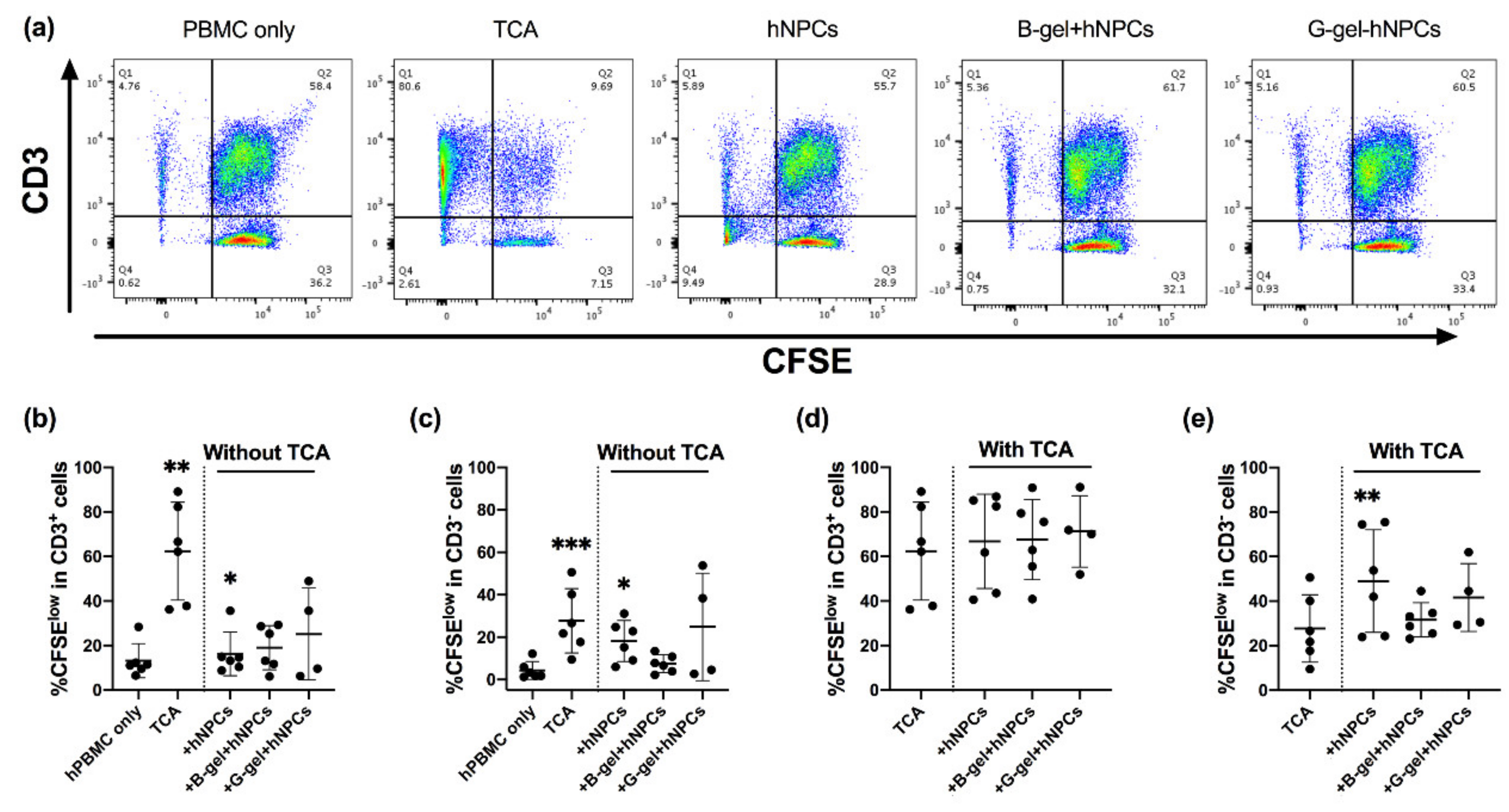
3.5. Evaluation of hPBMC Activation in Co-Culture with hNPCs, with or without Presence of Biomaterials
3.6. Evaluation of hPBMC Proliferation in Co-Culture with Allogeneic NPCs with or without the Presence of Two Different Forms of Hydrogel
3.7. hNPCs in Cell Suspension or Encapsulated in Bulk or Granular HA Hydrogel with TCA
3.8. Spidroin Preparations Still Contained Endotoxins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Badhiwala, J.H.; Ahuja, C.S.; Fehlings, M.G. Time is spine: A review of translational advances in spinal cord injury. J. Neurosurg. Spine 2018, 30, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biering-Sorensen, F.; Bickenbach, J.E.; El Masry, W.S.; Officer, A.; von Groote, P.M. ISCoS-WHO collaboration. International Perspectives of Spinal Cord Injury (IPSCI) report. Spinal. Cord 2011, 49, 679–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alizadeh, A.; Dyck, S.M.; Karimi-Abdolrezaee, S. Traumatic Spinal Cord Injury: An Overview of Pathophysiology, Models and Acute Injury Mechanisms. Front. Neurol. 2019, 10, 25. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Xie, Y.Y.; Wang, B. Role and prospects of regenerative biomaterials in the repair of spinal cord injury. Neural Regen. Res. 2019, 14, 1352–1363. [Google Scholar] [CrossRef]
- Thompson, R.E.; Pardieck, J.; Smith, L.; Kenny, P.; Crawford, L.; Shoichet, M.; Sakiyama-Elbert, S. Effect of hyaluronic acid hydrogels containing astrocyte-derived extracellular matrix and/or V2a interneurons on histologic outcomes following spinal cord injury. Biomaterials 2018, 162, 208–223. [Google Scholar] [CrossRef]
- Li, L.-M.; Han, M.; Jiang, X.-C.; Yin, X.-Z.; Chen, F.; Zhang, T.-Y.; Ren, H.; Zhang, J.-W.; Hou, T.-J.; Chen, Z.; et al. Peptide-Tethered Hydrogel Scaffold Promotes Recovery from Spinal Cord Transection via Synergism with Mesenchymal Stem Cells. ACS Appl. Mater. Interfaces 2017, 9, 3330–3342. [Google Scholar] [CrossRef] [PubMed]
- Zaviskova, K.; Tukmachev, D.; Dubisova, J.; Vackova, I.; Hejcl, A.; Bystronova, J.; Pravda, M.; Scigalkova, I.; Sulakova, R.; Velebny, V.; et al. Injectable hydroxyphenyl derivative of hyaluronic acid hydrogel modified with RGD as scaffold for spinal cord injury repair. J. Biomed. Mater. Res. Part A 2018, 106, 1129–1140. [Google Scholar] [CrossRef]
- Han, S.; Wang, B.; Jin, W.; Xiao, Z.; Li, X.; Ding, W.; Kapur, M.; Chen, B.; Yuan, B.; Zhu, T.; et al. The linear-ordered collagen scaffold-BDNF complex significantly promotes functional recovery after completely transected spinal cord injury in canine. Biomaterials 2015, 41, 89–96. [Google Scholar] [CrossRef]
- Li, X.; Zhao, Y.; Cheng, S.; Han, S.; Shu, M.; Chen, B.; Chen, X.; Tang, F.; Wang, N.; Tu, Y.; et al. Cetuximab modified collagen scaffold directs neurogenesis of injury-activated endogenous neural stem cells for acute spinal cord injury repair. Biomaterials 2017, 137, 73–86. [Google Scholar] [CrossRef]
- Koffler, J.; Zhu, W.; Qu, X.; Platoshyn, O.; Dulin, J.N.; Brock, J.; Graham, L.; Lu, P.; Sakamoto, J.; Marsala, M.; et al. Biomimetic 3D-printed scaffolds for spinal cord injury repair. Nat. Med. 2019. [Google Scholar] [CrossRef]
- Rooney, G.E.; Knight, A.M.; Madigan, N.N.; Gross, L.; Chen, B.; Giraldo, C.V.; Seo, S.; Nesbitt, J.J.; Dadsetan, M.; Yaszemski, M.J.; et al. Sustained delivery of dibutyryl cyclic adenosine monophosphate to the transected spinal cord via oligo [(polyethylene glycol) fumarate] hydrogels. Tissue Eng. Part A 2011, 17, 1287–1302. [Google Scholar] [CrossRef] [PubMed]
- Kornfeld, T.; Nessler, J.; Helmer, C.; Hannemann, R.; Waldmann, K.H.; Peck, C.T.; Hoffmann, P.; Brandes, G.; Vogt, P.M.; Radtke, C. Spider silk nerve graft promotes axonal regeneration on long distance nerve defect in a sheep model. Biomaterials 2021, 271, 120692. [Google Scholar] [CrossRef]
- Radtke, C.; Allmeling, C.; Waldmann, K.-H.; Reimers, K.; Thies, K.; Schenk, H.C.; Hillmer, A.; Guggenheim, M.; Brandes, G.; Vogt, P.M. Spider Silk Constructs Enhance Axonal Regeneration and Remyelination in Long Nerve Defects in Sheep. PLoS ONE 2011, 6, e16990. [Google Scholar] [CrossRef] [Green Version]
- Hunt, J.A.; Chen, R.; van Veen, T.; Bryan, N. Hydrogels for tissue engineering and regenerative medicine. J. Mater. Chem. B 2014, 2, 5319–5338. [Google Scholar] [CrossRef]
- Xue, K.; Wang, X.; Yong, P.W.; Young, D.J.; Wu, Y.-L.; Li, Z.; Loh, X.J. Hydrogels as Emerging Materials for Translational Biomedicine. Adv. Ther. 2019, 2, 1800088. [Google Scholar] [CrossRef] [Green Version]
- Gosline, J.M.; Guerette, P.A.; Ortlepp, C.S.; Savage, K.N. The mechanical design of spider silks: From fibroin sequence to mechanical function. J. Exp. Biol. 1999, 202, 3295–3303. [Google Scholar] [CrossRef] [PubMed]
- Salehi, S.; Koeck, K.; Scheibel, T. Spider Silk for Tissue Engineering Applications. Molecules 2020, 25, 737. [Google Scholar] [CrossRef] [Green Version]
- Vollrath, F.; Barth, P.; Basedow, A.; Engstrom, W.; List, H. Local tolerance to spider silks and protein polymers in vivo. In Vivo 2002, 16, 229–234. [Google Scholar] [PubMed]
- Varone, A.; Knight, D.; Lesage, S.; Vollrath, F.; Rajnicek, A.M.; Huang, W. The potential of Antheraea pernyi silk for spinal cord repair. Sci. Rep. 2017, 7, 13790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.; Kim, T.Y.; Lee, S.Y. Recent advances in production of recombinant spider silk proteins. Curr. Opin. Biotechnol. 2012, 23, 957–964. [Google Scholar] [CrossRef]
- Rising, A.; Johansson, J. Toward spinning artificial spider silk. Nat. Chem. Biol. 2015, 11, 309–315. [Google Scholar] [CrossRef]
- Andersson, M.; Jia, Q.; Abella, A.; Lee, X.-Y.; Landreh, M.; Purhonen, P.; Hebert, H.; Tenje, M.; Robinson, C.V.; Meng, Q.; et al. Biomimetic spinning of artificial spider silk from a chimeric minispidroin. Nat. Chem. Biol. 2017, 13, 262–264. [Google Scholar] [CrossRef] [PubMed]
- Lewicka, M.; Hermanson, O.; Rising, A.U. Recombinant spider silk matrices for neural stem cell cultures. Biomaterials 2012, 33, 7712–7717. [Google Scholar] [CrossRef] [Green Version]
- Widhe, M.; Bysell, H.; Nystedt, S.; Schenning, I.; Malmsten, M.; Johansson, J.; Rising, A.; Hedhammar, M. Recombinant spider silk as matrices for cell culture. Biomaterials 2010, 31, 9575–9585. [Google Scholar] [CrossRef]
- Wu, S.; Johansson, J.; Hovatta, O.; Rising, A. Efficient passage of human pluripotent stem cells on spider silk matrices under xeno-free conditions. Cell. Mol. Life Sci. 2015. [Google Scholar] [CrossRef]
- Wu, S.; Johansson, J.; Damdimopoulou, P.; Shahsavani, M.; Falk, A.; Hovatta, O.; Rising, A. Spider silk for xeno-free long-term self-renewal and differentiation of human pluripotent stem cells. Biomaterials 2014, 35, 8496–8502. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson, C.; Hedhammar, M.; Feinstein, R.; Nordling, K.; Kratz, G.; Johansson, J.; Huss, F.; Rising, A. Tissue Response to Subcutaneously Implanted Recombinant Spider Silk: An in Vivo Study. Materials 2009, 2, 1908. [Google Scholar] [CrossRef] [Green Version]
- Vecino, E.; Kwok, J.C.F. The Extracellular Matrix in the Nervous System: The Good and the Bad Aspects. In Composition and Function of the Extracellular Matrix in the Human Body; InTechOpen: London, UK, 2016. [Google Scholar]
- Khaing, Z.Z.; Milman, B.D.; Vanscoy, J.E.; Seidlits, S.K.; Grill, R.J.; Schmidt, C.E. High molecular weight hyaluronic acid limits astrocyte activation and scar formation after spinal cord injury. J. Neural Eng. 2011, 8, 046033. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, E.M. Hydrogel: Preparation, characterization, and applications: A review. J. Adv. Res. 2015, 6, 105–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peppas, N.A.; Bures, P.; Leobandung, W.; Ichikawa, H. Hydrogels in pharmaceutical formulations. Eur. J. Pharm. Biopharm. 2000, 50, 27–46. [Google Scholar] [CrossRef]
- Assuncao-Silva, R.C.; Gomes, E.D.; Sousa, N.; Silva, N.A.; Salgado, A.J. Hydrogels and Cell Based Therapies in Spinal Cord Injury Regeneration. Stem Cells Int. 2015, 2015, 24. [Google Scholar] [CrossRef] [PubMed]
- Schizas, N.; Rojas, R.; Kootala, S.; Andersson, B.; Pettersson, J.; Hilborn, J.; Hailer, N.P. Hyaluronic acid-based hydrogel enhances neuronal survival in spinal cord slice cultures from postnatal mice. J. Biomater. Appl. 2014, 28, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Madl, C.M.; LeSavage, B.L.; Dewi, R.E.; Lampe, K.J.; Heilshorn, S.C. Matrix Remodeling Enhances the Differentiation Capacity of Neural Progenitor Cells in 3D Hydrogels. Adv. Sci. 2019, 6, 1801716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madl, C.M.; LeSavage, B.L.; Dewi, R.E.; Dinh, C.B.; Stowers, R.S.; Khariton, M.; Lampe, K.J.; Nguyen, D.; Chaudhuri, O.; Enejder, A.; et al. Maintenance of neural progenitor cell stemness in 3D hydrogels requires matrix remodelling. Nat. Mater. 2017, 16, 1233. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-C.; George, J.H.; Waller, S.; Besnard, C.; Nagel, D.; Hill, E.; Coleman, M.D.; Korsunsky, A.M.; Cui, Z.; Ye, H. Increased Connectivity of hiPSC-derived Neural Networks in Multiphase of Granular Hydrogel Scaffolds. Bioact. Mater. Under revision.
- Riley, L.; Schirmer, L.; Segura, T. Granular hydrogels: Emergent properties of jammed hydrogel microparticles and their applications in tissue repair and regeneration. Curr. Opin. Biotechnol. 2018, 60, 1–8. [Google Scholar] [CrossRef]
- George, J.; Hsu, C.C.; Nguyen, L.T.B.; Ye, H.; Cui, Z. Neural tissue engineering with structured hydrogels in CNS models and therapies. Biotechnol. Adv. 2019. [Google Scholar] [CrossRef]
- Bergman, K.; Engstrand, T.; Hilborn, J.; Ossipov, D.; Piskounova, S.; Bowden, T. Injectable cell-free template for bone-tissue formation. J. Biomed. Mater. Res. Part A 2009, 91A, 1111–1118. [Google Scholar] [CrossRef]
- Siegelman, M.H.; DeGrendele, H.C.; Estess, P. Activation and interaction of CD44 and hyaluronan in immunological systems. J. Leukoc. Biol. 1999, 66, 315–321. [Google Scholar] [CrossRef]
- Kajahn, J.; Franz, S.; Rueckert, E.; Forstreuter, I.; Hintze, V.; Moeller, S.; Simon, J.C. Artificial extracellular matrices composed of collagen I and high sulfated hyaluronan modulate monocyte to macrophage differentiation under conditions of sterile inflammation. Biomatter 2012, 2, 226–273. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Yokohama, S.; Yoneda, M.; Okamoto, S.; Tamaki, Y.; Ito, T.; Okada, M.; Aso, K.; Makino, I. High, but not low, molecular weight hyaluronan prevents T-cell-mediated liver injury by reducing proinflammatory cytokines in mice. J. Gastroenterol. 2004, 39, 346–354. [Google Scholar] [CrossRef]
- Kwon, M.Y.; Wang, C.; Galarraga, J.H.; Pure, E.; Han, L.; Burdick, J.A. Influence of hyaluronic acid modification on CD44 binding towards the design of hydrogel biomaterials. Biomaterials 2019, 222, 9. [Google Scholar] [CrossRef]
- Luttikhuizen, D.T.; Harmsen, M.C.; Van Luyn, M.J. Cellular and molecular dynamics in the foreign body reaction. Tissue Eng. 2006, 12, 1955–1970. [Google Scholar] [CrossRef]
- Jones, J.A.; Chang, D.T.; Meyerson, H.; Colton, E.; Kwon, I.K.; Matsuda, T.; Anderson, J.M. Proteomic analysis and quantification of cytokines and chemokines from biomaterial surface-adherent macrophages and foreign body giant cells. J. Biomed. Mater. Res. Part A 2007, 83A, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Andorko, J.I.; Jewell, C.M. Designing biomaterials with immunomodulatory properties for tissue engineering and regenerative medicine. Bioeng. Transl. Med. 2017, 2, 139–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Shea, T.M.; Wollenberg, A.L.; Kim, J.H.; Ao, Y.; Deming, T.J.; Sofroniew, M.V. Foreign body responses in mouse central nervous system mimic natural wound responses and alter biomaterial functions. Nat. Commun. 2020, 11, 6203. [Google Scholar] [CrossRef]
- Odeberg, J.; Piao, J.H.; Samuelsson, E.B.; Falci, S.; Akesson, E. Low immunogenicity of in vitro-expanded human neural cells despite high MHC expression. J. Neuroimmunol. 2005, 161, 1–11. [Google Scholar] [CrossRef]
- Piao, J.H.; Odeberg, J.; Samuelsson, E.B.; Kjaeldgaard, A.; Falci, S.; Seiger, A.; Sundstrom, E.; Akesson, E. Cellular composition of long-term human spinal cord- and forebrain-derived neurosphere cultures. J. Neurosci. Res. 2006, 84, 471–482. [Google Scholar] [CrossRef]
- Liu, J.; Gotherstrom, C.; Forsberg, M.; Samuelsson, E.B.; Wu, J.; Calzarossa, C.; Hovatta, O.; Sundstrom, E.; Akesson, E. Human neural stem/progenitor cells derived from embryonic stem cells and fetal nervous system present differences in immunogenicity and immunomodulatory potentials in vitro. Stem Cell Res. 2013, 10, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-C.; George, J.H.; Waller, S.; Besnard, C.; Korsunsky, A.M.; Ye, H.; Cui, Z. Culture of Human Induced Pluripotent Stem Cell-Derived Neurons in Multi-phase Granular Hydrogels. In Proceedings of the Biomaterials for Cell and Drug Delivery 2019, Manchester, UK, 17–19 September 2019. [Google Scholar]
- Sancho, D.; Gómez, M.; Sánchez-Madrid, F. CD69 is an immunoregulatory molecule induced following activation. Trends Immunol. 2005, 26, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Vinay, D.S.; Kwon, B.S. Role of 4-1BB in immune responses. Semin. Immunol. 1998, 10, 481–489. [Google Scholar] [CrossRef]
- Aleksander-Konert, E.; Paduszyński, P.; Zajdel, A.; Dzierżewicz, Z.; Wilczok, A. In vitro chondrogenesis of Wharton’s jelly mesenchymal stem cells in hyaluronic acid-based hydrogels. Cell. Mol. Biol. Lett. 2016, 21, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groell, F.; Kalia, Y.N.; Jordan, O.; Borchard, G. Hydrogels in three-dimensional dendritic cell (MUTZ-3) culture as a scaffold to mimic human immuno competent subcutaneous tissue. Int. J. Pharm. 2018, 544, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Mamat, U.; Wilke, K.; Bramhill, D.; Schromm, A.B.; Lindner, B.; Kohl, T.A.; Corchero, J.L.; Villaverde, A.; Schaffer, L.; Head, S.R.; et al. Detoxifying Escherichia coli for endotoxin-free production of recombinant proteins. Microb. Cell Factories 2015, 14, 57. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, A.R.; Tanase, C.E.; Awuah, D.; Vasanthi Bathrinarayanan, P.; Hall, L.; Nikkhah, M.; Khademhosseini, A.; Rose, F.; Alexander, C.; Ghaemmaghami, A.M. Photocrosslinkable Gelatin Hydrogels Modulate the Production of the Major Pro-inflammatory Cytokine, TNF-α, by Human Mononuclear Cells. Front. Bioeng. Biotechnol. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Cibrian, D.; Sanchez-Madrid, F. CD69: From activation marker to metabolic gatekeeper. Eur. J. Immunol. 2017, 47, 946–953. [Google Scholar] [CrossRef]
- Adusei, K.M.; Ngo, T.B.; Sadtler, K. T Lymphocytes as Critical Mediators in Tissue Regeneration, Fibrosis, and the Foreign Body Response. Acta Biomater. 2021. [Google Scholar] [CrossRef]
- Nadol, J.B., Jr.; O’Malley, J.T.; Burgess, B.J.; Galler, D. Cellular immunologic responses to cochlear implantation in the human. Hear. Res. 2014, 318, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Chaudhary, N.; Gemmete, J.J.; Thompson, B.G.; Xi, G.; Pandey, A.S. Reactive tissue proliferation and damage of elastic lamina caused by hydrogel coated coils in experimental rat aneurysms. J. Neurointerv. Surg. 2014, 6, 480–486. [Google Scholar] [CrossRef]
- Baeva, L.F.; Lyle, D.B.; Rios, M.; Langone, J.J.; Lightfoote, M.M. Different molecular weight hyaluronic acid effects on human macrophage interleukin 1β production. J. Biomed. Mater. Res. Part A 2014, 102, 305–314. [Google Scholar] [CrossRef]
- Snetkov, P.; Zakharova, K.; Morozkina, S.; Olekhnovich, R.; Uspenskaya, M. Hyaluronic Acid: The Influence of Molecular Weight on Structural, Physical, Physico-Chemical, and Degradable Properties of Biopolymer. Polymers 2020, 12, 1800. [Google Scholar] [CrossRef] [PubMed]
- Cyphert, J.M.; Trempus, C.S.; Garantziotis, S. Size Matters: Molecular Weight Specificity of Hyaluronan Effects in Cell Biology. Int. J. Cell Biol. 2015, 2015, 563818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedhammar, M.; Bramfeldt, H.; Baris, T.; Widhe, M.; Askarieh, G.; Nordling, K.; Aulock, S.; Johansson, J. Sterilized recombinant spider silk fibers of low pyrogenicity. Biomacromolecules 2010, 11, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Decker, R.E.; Harris, T.I.; Memmott, D.R.; Peterson, C.J.; Lewis, R.V.; Jones, J.A. Method for the Destruction of Endotoxin in Synthetic Spider Silk Proteins. Sci. Rep. 2018, 8, 12166. [Google Scholar] [CrossRef]
- Schacht, K.; Scheibel, T. Controlled Hydrogel Formation of a Recombinant Spider Silk Protein. Biomacromolecules 2011, 12, 2488–2495. [Google Scholar] [CrossRef]
- Osama, I.; Gorenkova, N.; McKittrick, C.M.; Wongpinyochit, T.; Goudie, A.; Seib, F.P.; Carswell, H.V.O. In vitro studies on space-conforming self-assembling silk hydrogels as a mesenchymal stem cell-support matrix suitable for minimally invasive brain application. Sci. Rep. 2018, 8, 13655. [Google Scholar] [CrossRef]
- Hopkins, A.M.; De Laporte, L.; Tortelli, F.; Spedden, E.; Staii, C.; Atherton, T.J.; Hubbell, J.A.; Kaplan, D.L. Silk Hydrogels as Soft Substrates for Neural Tissue Engineering. Adv. Funct. Mater. 2013, 23, 5140–5149. [Google Scholar] [CrossRef]
- Ozaki, M.; Iwanami, A.; Nagoshi, N.; Kohyama, J.; Itakura, G.; Iwai, H.; Nishimura, S.; Nishiyama, Y.; Kawabata, S.; Sugai, K.; et al. Evaluation of the immunogenicity of human iPS cell-derived neural stem/progenitor cells in vitro. Stem Cell Res. 2017, 19, 128–138. [Google Scholar] [CrossRef]
- Akesson, E.; Wolmer-Solberg, N.; Cederarv, M.; Falci, S.; Odeberg, J. Human neural stem cells and astrocytes, but not neurons, suppress an allogeneic lymphocyte response. Stem Cell Res. 2009, 2, 56–67. [Google Scholar] [CrossRef] [Green Version]
- Ubiali, F.; Nava, S.; Nessi, V.; Frigerio, S.; Parati, E.; Bernasconi, P.; Mantegazza, R.; Baggi, F. Allorecognition of human neural stem cells by peripheral blood lymphocytes despite low expression of MHC molecules: Role of TGF-beta in modulating proliferation. Int. Immunol. 2007, 19, 1063–1074. [Google Scholar] [CrossRef] [Green Version]
- Ekblad-Nordberg, Å.; Walther-Jallow, L.; Westgren, M.; Götherström, C. Prenatal stem cell therapy for inherited diseases: Past, present, and future treatment strategies. Stem Cells Transl. Med. 2020, 9, 148–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urdzikova, L.M.; Ruzicka, J.; LaBagnara, M.; Karova, K.; Kubinova, S.; Jirakova, K.; Murali, R.; Sykova, E.; Jhanwar-Uniyal, M.; Jendelova, P. Human mesenchymal stem cells modulate inflammatory cytokines after spinal cord injury in rat. Int. J. Mol. Sci. 2014, 15, 11275–11293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirova, M.; Vlierberghe, S.V.; Matyasova, V.; Rossmann, P.; Schacht, E.; Dubruel, P.; Rihova, B. Immunocompatibility evaluation of hydrogel-coated polyimide implants for applications in regenerative medicine. J. Biomed. Mater. Res. Part A 2014, 102, 1982–1990. [Google Scholar] [CrossRef] [PubMed]
- Kłodzińska, S.N.; Pletzer, D.; Rahanjam, N.; Rades, T.; Hancock, R.E.W.; Nielsen, H.M. Hyaluronic acid-based nanogels improve in vivo compatibility of the anti-biofilm peptide DJK-5. Nanomed. Nanotechnol. Biol. Med. 2019, 20, 102022. [Google Scholar] [CrossRef] [PubMed]
- Kampleitner, C.; Obi, J.; Vassilev, N.; Epstein, M.M.; Hoffmann, O. Biological Compatibility Profile on Biomaterials for Bone Regeneration. J. Vis. Exp. 2018. [Google Scholar] [CrossRef] [PubMed]
- Arthe, R.; Arivuoli, D.; Ravi, V. Preparation and characterization of bioactive silk fibroin/paramylon blend films for chronic wound healing. Int. J. Biol. Macromol. 2020, 154, 1324–1331. [Google Scholar] [CrossRef]
- Krüger-Genge, A.; Tondera, C.; Hauser, S.; Braune, S.; Görs, J.; Roch, T.; Klopfleisch, R.; Neffe, A.T.; Lendlein, A.; Pietzsch, J.; et al. Immunocompatibility and non-thrombogenicity of gelatin-based hydrogels. Clin. Hemorheol. Microcirc. 2021, 77, 335–350. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.; Ekblad-Nordberg, Å.; Michaëlsson, J.; Götherström, C.; Hsu, C.-C.; Ye, H.; Johansson, J.; Rising, A.; Sundström, E.; Åkesson, E. In Vitro Study of Human Immune Responses to Hyaluronic Acid Hydrogels, Recombinant Spidroins and Human Neural Progenitor Cells of Relevance to Spinal Cord Injury Repair. Cells 2021, 10, 1713. https://doi.org/10.3390/cells10071713
Lin C, Ekblad-Nordberg Å, Michaëlsson J, Götherström C, Hsu C-C, Ye H, Johansson J, Rising A, Sundström E, Åkesson E. In Vitro Study of Human Immune Responses to Hyaluronic Acid Hydrogels, Recombinant Spidroins and Human Neural Progenitor Cells of Relevance to Spinal Cord Injury Repair. Cells. 2021; 10(7):1713. https://doi.org/10.3390/cells10071713
Chicago/Turabian StyleLin, Chenhong, Åsa Ekblad-Nordberg, Jakob Michaëlsson, Cecilia Götherström, Chia-Chen Hsu, Hua Ye, Jan Johansson, Anna Rising, Erik Sundström, and Elisabet Åkesson. 2021. "In Vitro Study of Human Immune Responses to Hyaluronic Acid Hydrogels, Recombinant Spidroins and Human Neural Progenitor Cells of Relevance to Spinal Cord Injury Repair" Cells 10, no. 7: 1713. https://doi.org/10.3390/cells10071713
APA StyleLin, C., Ekblad-Nordberg, Å., Michaëlsson, J., Götherström, C., Hsu, C. -C., Ye, H., Johansson, J., Rising, A., Sundström, E., & Åkesson, E. (2021). In Vitro Study of Human Immune Responses to Hyaluronic Acid Hydrogels, Recombinant Spidroins and Human Neural Progenitor Cells of Relevance to Spinal Cord Injury Repair. Cells, 10(7), 1713. https://doi.org/10.3390/cells10071713