
Dietary Fatty Acids at the Crossroad between Obesity and Colorectal Cancer: Fine Regulators of Adipose Tissue Homeostasis and Immune Response

 , , and
, , and
Abstract
:1. Introduction
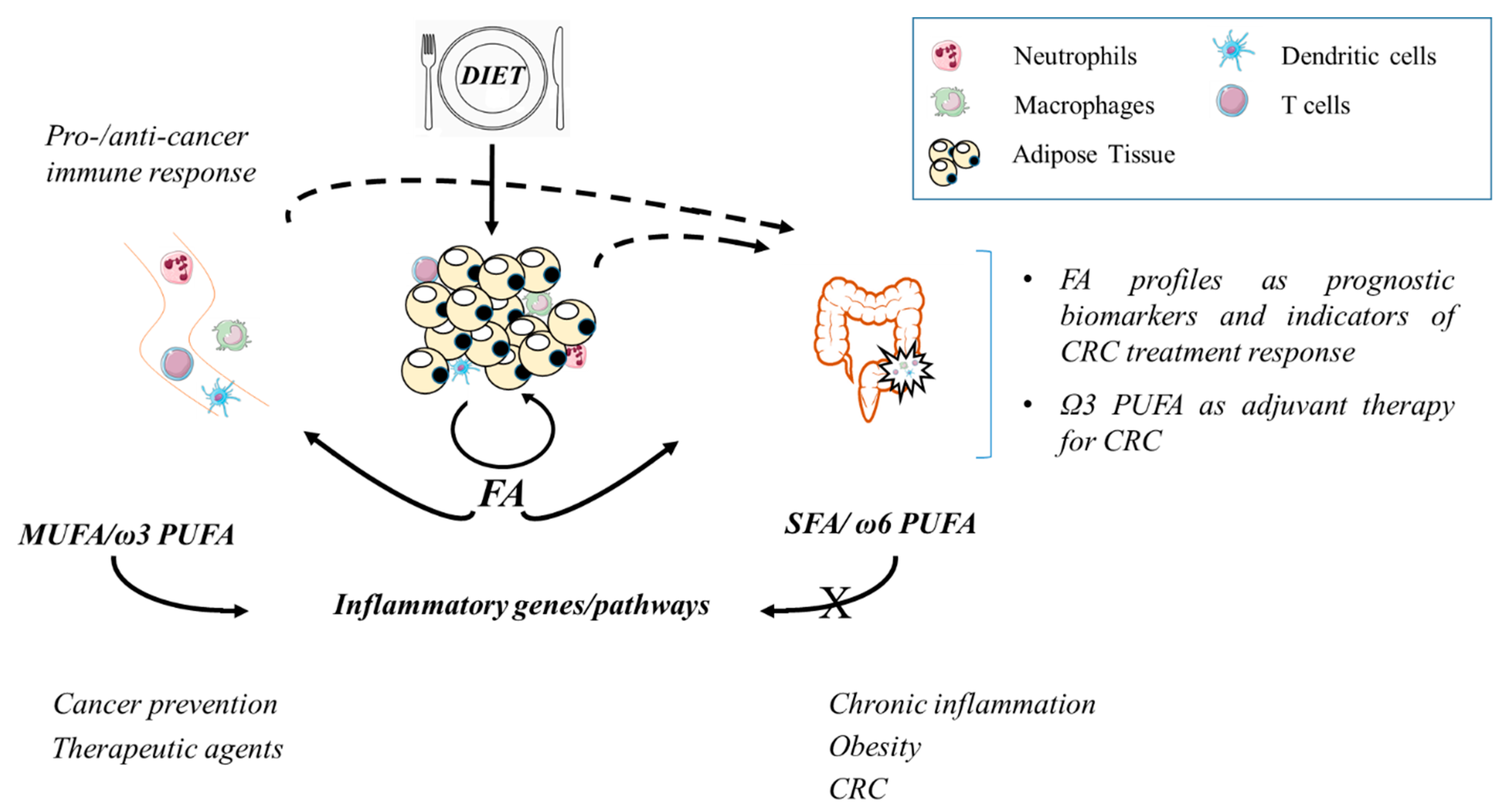
2. Fatty Acid Profiles in Obesity and Colorectal Cancer and Their Relationship with Dietary Intake
3. Fatty Acids and Adipose Tissue Homeostasis
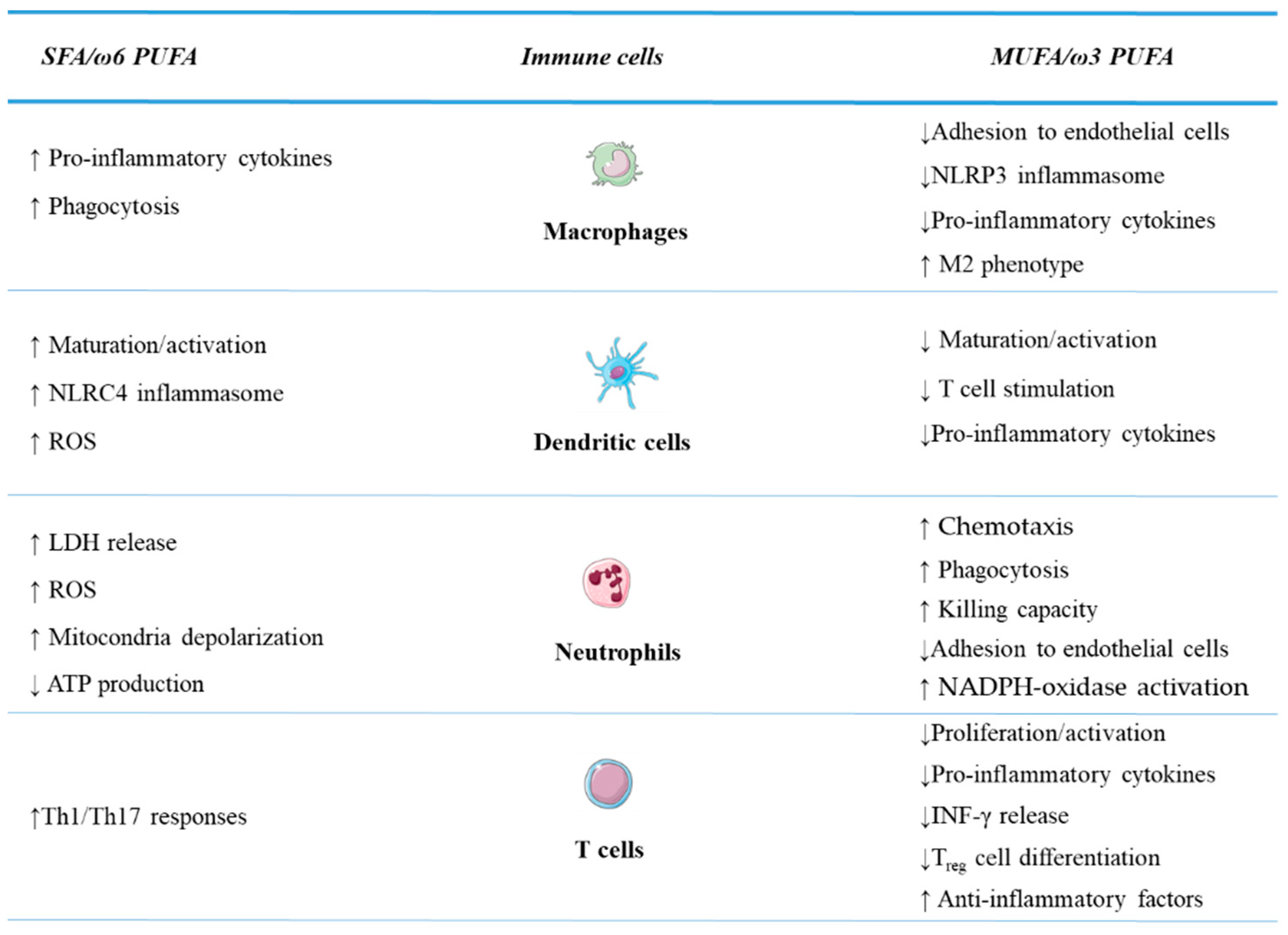
4. Fatty Acids and Immune Cells: Regulation of Transcription Factors, Inflammatory Pathways, and Effector Functions
4.1. Effects of Dietary Fatty Acid Supplementation on Blood Immune Cell Gene Expression Profiles
4.2. Effects of In Vitro Fatty Acid Exposure on Immune Cell Functions
4.2.1. Innate Immunity Cells
4.2.2. Adaptive Immunity Cells
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collaboration, N.C.D.R.F. Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar] [CrossRef] [Green Version]
- James, W.P.T.; McPherson, K. The costs of overweight. Lancet. Public Health 2017, 2, e203–e204. [Google Scholar] [CrossRef] [Green Version]
- Malik, V.S.; Willett, W.C.; Hu, F.B. Global obesity: Trends, risk factors and policy implications. Nat. Rev. Endocrinol. 2013, 9, 13–27. [Google Scholar] [CrossRef]
- Cozzo, A.J.; Fuller, A.M.; Makowski, L. Contribution of adipose tissue to development of cancer. Compr. Physiol. 2017, 8, 237–282. [Google Scholar] [CrossRef]
- Thanikachalam, K.; Khan, G. Colorectal cancer and nutrition. Nutrients 2019, 11, 164. [Google Scholar] [CrossRef] [Green Version]
- Keum, N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef]
- Bardou, M.; Barkun, A.N.; Martel, M. Obesity and colorectal cancer. Gut 2013, 62, 933–947. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Morley, T.S.; Kim, M.; Clegg, D.J.; Scherer, P.E. Obesity and cancer--mechanisms underlying tumour progression and recurrence. Nat. Rev. Endocrinol. 2014, 10, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Useros, J.; Garcia-Foncillas, J. Obesity and colorectal cancer: Molecular features of adipose tissue. J. Transl. Med. 2016, 14, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alwarawrah, Y.; Kiernan, K.; MacIver, N.J. Changes in nutritional status impact immune cell metabolism and function. Front. Immunol. 2018, 9, 1055. [Google Scholar] [CrossRef] [Green Version]
- Matafome, P.; Seica, R. Function and dysfunction of adipose tissue. Adv. Neurobiol. 2017, 19, 3–31. [Google Scholar] [CrossRef]
- Huh, J.Y.; Park, Y.J.; Ham, M.; Kim, J.B. Crosstalk between adipocytes and immune cells in adipose tissue inflammation and metabolic dysregulation in obesity. Mol. Cells 2014, 37, 365–371. [Google Scholar] [CrossRef]
- Kershaw, E.E.; Flier, J.S. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef]
- Meiliana, A.; Dewi, N.M.; Wijaya, A. Adipose tissue, inlammation (meta-inlammation) and obesity management. Indones Biomed. J. 2015, 7, 129–146. [Google Scholar] [CrossRef]
- Song, Y.C.; Lee, S.E.; Jin, Y.; Park, H.W.; Chun, K.H.; Lee, H.W. Classifying the linkage between adipose tissue inflammation and tumor growth through cancer-associated adipocytes. Mol. Cells 2020, 43, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Masoodi, M.; Kuda, O.; Rossmeisl, M.; Flachs, P.; Kopecky, J. Lipid signaling in adipose tissue: Connecting inflammation & metabolism. Biochim. Biophys. Acta 2015, 1851, 503–518. [Google Scholar] [CrossRef] [PubMed]
- Rocha, D.M.; Bressan, J.; Hermsdorff, H.H. The role of dietary fatty acid intake in inflammatory gene expression: A critical review. Sao Paulo Med. J. Rev. Paul. Med. 2017, 135, 157–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes: From molecules to man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Beek, C.M.; Dejong, C.H.C.; Troost, F.J.; Masclee, A.A.M.; Lenaerts, K. Role of short-chain fatty acids in colonic inflammation, carcinogenesis, and mucosal protection and healing. Nutr. Rev. 2017, 75, 286–305. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C.; Yaqoob, P. Omega-3 polyunsaturated fatty acids and human health outcomes. BioFactors 2009, 35, 266–272. [Google Scholar] [CrossRef]
- Ralston, J.C.; Lyons, C.L.; Kennedy, E.B.; Kirwan, A.M.; Roche, H.M. Fatty Acids and NLRP3 inflammasome-mediated inflammation in metabolic tissues. Annu. Rev. Nutr. 2017, 37, 77–102. [Google Scholar] [CrossRef]
- Wajchenberg, B.L. Subcutaneous and visceral adipose tissue: Their relation to the metabolic syndrome. Endocr. Rev. 2000, 21, 697–738. [Google Scholar] [CrossRef]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose tissue dysfunction as determinant of obesity-associated metabolic complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [Green Version]
- Vaittinen, M.; Mannisto, V.; Kakela, P.; Agren, J.; Tiainen, M.; Schwab, U.; Pihlajamaki, J. Interorgan cross talk between fatty acid metabolism, tissue inflammation, and FADS2 genotype in humans with obesity. Obesity 2017, 25, 545–552. [Google Scholar] [CrossRef] [Green Version]
- Garaulet, M.; Perez-Llamas, F.; Perez-Ayala, M.; Martinez, P.; de Medina, F.S.; Tebar, F.J.; Zamora, S. Site-specific differences in the fatty acid composition of abdominal adipose tissue in an obese population from a Mediterranean area: Relation with dietary fatty acids, plasma lipid profile, serum insulin, and central obesity. Am. J. Clin. Nutr. 2001, 74, 585–591. [Google Scholar] [CrossRef] [Green Version]
- D’Archivio, M.; Scazzocchio, B.; Giammarioli, S.; Fiani, M.L.; Vari, R.; Santangelo, C.; Veneziani, A.; Iacovelli, A.; Giovannini, C.; Gessani, S.; et al. omega3-PUFAs exert anti-inflammatory activity in visceral adipocytes from colorectal cancer patients. PLoS ONE 2013, 8, e77432. [Google Scholar] [CrossRef] [Green Version]
- Del Corno, M.; D’Archivio, M.; Conti, L.; Scazzocchio, B.; Vari, R.; Donninelli, G.; Varano, B.; Giammarioli, S.; De Meo, S.; Silecchia, G.; et al. Visceral fat adipocytes from obese and colorectal cancer subjects exhibit distinct secretory and omega6 polyunsaturated fatty acid profiles and deliver immunosuppressive signals to innate immunity cells. Oncotarget 2016, 7, 63093–63105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, J.; Campos, H.; McGarvey, S.; Wu, Z.; Goldberg, R.; Baylin, A. Genetic variation in stearoyl-CoA desaturase 1 is associated with metabolic syndrome prevalence in Costa Rican adults. J. Nutr. 2011, 141, 2211–2218. [Google Scholar] [CrossRef] [Green Version]
- Scazzocchio, B.; Vari, R.; Silenzi, A.; Giammarioli, S.; Masotti, A.; Baldassarre, A.; Santangelo, C.; D’Archivio, M.; Giovannini, C.; Del Corno, M.; et al. Dietary habits affect fatty acid composition of visceral adipose tissue in subjects with colorectal cancer or obesity. Eur. J. Nutr. 2020, 59, 1463–1472. [Google Scholar] [CrossRef] [PubMed]
- Boden, G. Obesity and free fatty acids. Endocrinol. Metab. Clin. N. Am. 2008, 37, 635–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arner, P.; Ryden, M. Fatty acids, obesity and insulin resistance. Obes. Facts 2015, 8, 147–155. [Google Scholar] [CrossRef]
- Hodson, L.; McQuaid, S.E.; Karpe, F.; Frayn, K.N.; Fielding, B.A. Differences in partitioning of meal fatty acids into blood lipid fractions: A comparison of linoleate, oleate, and palmitate. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E64–E71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iggman, D.; Arnlov, J.; Cederholm, T.; Riserus, U. Association of adipose tissue fatty acids with cardiovascular and all-cause mortality in elderly men. JAMA Cardiol. 2016, 1, 745–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Z.; Cao, C.; Li, Q.; Xu, Y.J.; Liu, Y. Different dietary lipid consumption affects the serum lipid profiles, colonic short chain fatty acid composition and the gut health of Sprague Dawley rats. Food Res. Int. 2020, 132, 109117. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, S.J.; Feskens, E.J.; Bos, M.B.; Hoelen, D.W.; Heijligenberg, R.; Bromhaar, M.G.; de Groot, L.C.; de Vries, J.H.; Muller, M.; Afman, L.A. A saturated fatty acid-rich diet induces an obesity-linked proinflammatory gene expression profile in adipose tissue of subjects at risk of metabolic syndrome. Am. J. Clin. Nutr. 2009, 90, 1656–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May-Wilson, S.; Sud, A.; Law, P.J.; Palin, K.; Tuupanen, S.; Gylfe, A.; Hanninen, U.A.; Cajuso, T.; Tanskanen, T.; Kondelin, J.; et al. Pro-inflammatory fatty acid profile and colorectal cancer risk: A Mendelian randomisation analysis. Eur. J. Cancer 2017, 84, 228–238. [Google Scholar] [CrossRef] [Green Version]
- Carroll, R.G.; Zaslona, Z.; Galvan-Pena, S.; Koppe, E.L.; Sevin, D.C.; Angiari, S.; Triantafilou, M.; Triantafilou, K.; Modis, L.K.; O’Neill, L.A. An unexpected link between fatty acid synthase and cholesterol synthesis in proinflammatory macrophage activation. J. Biol. Chem. 2018, 293, 5509–5521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pakiet, A.; Kobiela, J.; Stepnowski, P.; Sledzinski, T.; Mika, A. Changes in lipids composition and metabolism in colorectal cancer: A review. Lipids Health Dis. 2019, 18, 29. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.A.; Xing, X.; Harris, J.W.; Zaytseva, Y.Y.; Mitov, M.I.; Napier, D.L.; Weiss, H.L.; Mark Evers, B.; Gao, T. Adipocytes activate mitochondrial fatty acid oxidation and autophagy to promote tumor growth in colon cancer. Cell Death Dis. 2017, 8, e2593. [Google Scholar] [CrossRef]
- Okuno, M.; Hamazaki, K.; Ogura, T.; Kitade, H.; Matsuura, T.; Yoshida, R.; Hijikawa, T.; Kwon, M.; Arita, S.; Itomura, M.; et al. Abnormalities in fatty acids in plasma, erythrocytes and adipose tissue in Japanese patients with colorectal cancer. In Vivo 2013, 27, 203–210. [Google Scholar]
- Giuliani, A.; Ferrara, F.; Scimo, M.; Angelico, F.; Olivieri, L.; Basso, L. Adipose tissue fatty acid composition and colon cancer: A case-control study. Eur. J. Nutr. 2014, 53, 1029–1037. [Google Scholar] [CrossRef]
- Cottet, V.; Vaysse, C.; Scherrer, M.L.; Ortega-Deballon, P.; Lakkis, Z.; Delhorme, J.B.; Deguelte-Lardiere, S.; Combe, N.; Bonithon-Kopp, C. Fatty acid composition of adipose tissue and colorectal cancer: A case-control study. Am. J. Clin. Nutr. 2015, 101, 192–201. [Google Scholar] [CrossRef] [Green Version]
- Berry, E.M.; Zimmerman, J.; Peser, M.; Ligumsky, M. Dietary fat, adipose tissue composition, and the development of carcinoma of the colon. J. Natl. Cancer Inst. 1986, 77, 93–97. [Google Scholar]
- Neoptolemos, J.P.; Clayton, H.; Heagerty, A.M.; Nicholson, M.J.; Johnson, B.; Mason, J.; Manson, K.; James, R.F.; Bell, P.R. Dietary fat in relation to fatty acid composition of red cells and adipose tissue in colorectal cancer. Br. J. Cancer 1988, 58, 575–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosconi, E.; Minicozzi, A.; Marzola, P.; Cordiano, C.; Sbarbati, A. (1) H-MR spectroscopy characterization of the adipose tissue associated with colorectal tumor. J. Magn. Reson. Imaging JMRI 2014, 39, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Bakker, N.; Van’t Veer, P.; Zock, P.L. Adipose fatty acids and cancers of the breast, prostate and colon: An ecological study. EURAMIC Study Group. Int. J. Cancer 1997, 72, 587–591. [Google Scholar] [CrossRef]
- Zhang, L.; Han, L.; He, J.; Lv, J.; Pan, R.; Lv, T. A high serum-free fatty acid level is associated with cancer. J. Cancer Res. Clin. Oncol. 2020, 146, 705–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aglago, E.K.; Huybrechts, I.; Murphy, N.; Casagrande, C.; Nicolas, G.; Pischon, T.; Fedirko, V.; Severi, G.; Boutron-Ruault, M.C.; Fournier, A.; et al. Consumption of fish and long-chain n-3 polyunsaturated fatty acids is associated with reduced risk of colorectal cancer in a Large European Cohort. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2020, 18, 654–666.e656. [Google Scholar] [CrossRef] [PubMed]
- Suganami, T.; Tanimoto-Koyama, K.; Nishida, J.; Itoh, M.; Yuan, X.; Mizuarai, S.; Kotani, H.; Yamaoka, S.; Miyake, K.; Aoe, S.; et al. Role of the Toll-like receptor 4/NF-kappaB pathway in saturated fatty acid-induced inflammatory changes in the interaction between adipocytes and macrophages. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzik, T.J.; Skiba, D.S.; Touyz, R.M.; Harrison, D.G. The role of infiltrating immune cells in dysfunctional adipose tissue. Cardiovasc. Res. 2017, 113, 1009–1023. [Google Scholar] [CrossRef] [Green Version]
- Del Corno, M.; Conti, L.; Gessani, S. Innate lymphocytes in adipose tissue homeostasis and their alterations in obesity and colorectal cancer. Front. Immunol. 2018, 9, 2556. [Google Scholar] [CrossRef] [Green Version]
- Castellano-Castillo, D.; Morcillo, S.; Clemente-Postigo, M.; Crujeiras, A.B.; Fernandez-Garcia, J.C.; Torres, E.; Tinahones, F.J.; Macias-Gonzalez, M. Adipose tissue inflammation and VDR expression and methylation in colorectal cancer. Clin. Epigenetics 2018, 10, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, C.; Eter, L.; Lanzetta, N.; Abrishami, S.; Varghese, M.; McKernan, K.; Muir, L.; Lane, J.; Lumeng, C.N.; Singer, K. TLR4, TRIF, and MyD88 are essential for myelopoiesis and CD11c(+) adipose tissue macrophage production in obese mice. J. Biol. Chem. 2018, 293, 8775–8786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luna-Vital, D.; Luzardo-Ocampo, I.; Cuellar-Nunez, M.L.; Loarca-Pina, G.; Gonzalez de Mejia, E. Maize extract rich in ferulic acid and anthocyanins prevents high-fat-induced obesity in mice by modulating SIRT1, AMPK and IL-6 associated metabolic and inflammatory pathways. J. Nutr. Biochem. 2020, 79, 108343. [Google Scholar] [CrossRef] [PubMed]
- Boughanem, H.; Cabrera-Mulero, A.; Hernandez-Alonso, P.; Bandera-Merchan, B.; Tinahones, A.; Tinahones, F.J.; Morcillo, S.; Macias-Gonzalez, M. The expression/methylation profile of adipogenic and inflammatory transcription factors in adipose tissue are linked to obesity-related colorectal cancer. Cancers 2019, 11, 1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennacer, A.F.; Haffaf, E.; Kacimi, G.; Oudjit, B.; Koceir, E.A. Association of polyunsaturated/saturated fatty acids to metabolic syndrome cardiovascular risk factors and lipoprotein (a) in hypertensive type 2 diabetic patients. Ann. Biol. Clin. 2017, 75, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Yacoubian, S.; Yang, R. Anti-inflammatory and proresolving lipid mediators. Annu. Rev. Pathol. 2008, 3, 279–312. [Google Scholar] [CrossRef] [Green Version]
- Neacsu, O.; Cleveland, K.; Xu, H.; Tchkonia, T.T.; Kirkland, J.L.; Boney, C.M. IGF-I attenuates FFA-induced activation of JNK1 phosphorylation and TNFalpha expression in human subcutaneous preadipocytes. Obesity 2013, 21, 1843–1849. [Google Scholar] [CrossRef] [Green Version]
- Youssef-Elabd, E.M.; McGee, K.C.; Tripathi, G.; Aldaghri, N.; Abdalla, M.S.; Sharada, H.M.; Ashour, E.; Amin, A.I.; Ceriello, A.; O’Hare, J.P.; et al. Acute and chronic saturated fatty acid treatment as a key instigator of the TLR-mediated inflammatory response in human adipose tissue, in vitro. J. Nutr. Biochem. 2012, 23, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Murumalla, R.K.; Gunasekaran, M.K.; Padhan, J.K.; Bencharif, K.; Gence, L.; Festy, F.; Cesari, M.; Roche, R.; Hoareau, L. Fatty acids do not pay the toll: Effect of SFA and PUFA on human adipose tissue and mature adipocytes inflammation. Lipids Health Dis. 2012, 11, 175. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, J.F.; Roberts-Lee, K.; Borcea, C.; Smith, H.M.; Midgette, Y.; Shah, R. Omega-3 polyunsaturated fatty acids attenuate inflammatory activation and alter differentiation in human adipocytes. J. Nutr. Biochem. 2019, 64, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.R.; Midgette, Y.; Shah, R. Fish Oil Derived Omega 3 Fatty Acids Suppress Adipose NLRP3 Inflammasome Signaling in Human Obesity. J. Endocr. Soc. 2019, 3, 504–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donninelli, G.; Del Corno, M.; Pierdominici, M.; Scazzocchio, B.; Vari, R.; Varano, B.; Pacella, I.; Piconese, S.; Barnaba, V.; D’Archivio, M.; et al. Distinct blood and visceral adipose tissue regulatory t cell and innate lymphocyte profiles characterize obesity and colorectal cancer. Front. Immunol. 2017, 8, 643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Corno, M.; Baldassarre, A.; Calura, E.; Conti, L.; Martini, P.; Romualdi, C.; Vari, R.; Scazzocchio, B.; D’Archivio, M.; Masotti, A.; et al. Transcriptome profiles of human visceral adipocytes in obesity and colorectal cancer unravel the effects of body mass index and polyunsaturated fatty acids on genes and biological processes related to tumorigenesis. Front. Immunol. 2019, 10, 265. [Google Scholar] [CrossRef] [Green Version]
- Ralston, J.C.; Matravadia, S.; Gaudio, N.; Holloway, G.P.; Mutch, D.M. Polyunsaturated fatty acid regulation of adipocyte FADS1 and FADS2 expression and function. Obesity 2015, 23, 725–728. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Geng, T.; Huang, T.; Zhao, Q. Fish oil supplementation and insulin sensitivity: A systematic review and meta-analysis. Lipids Health Dis. 2017, 16, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, D.R.; Pieri, B.; Comim, V.H.; Marques, S.O.; Luciano, T.F.; Rodrigues, M.S.; De Souza, C.T. Fish oil reduces subclinical inflammation, insulin resistance, and atherogenic factors in overweight/obese type 2 diabetes mellitus patients: A pre-post pilot study. J. Diabetes Complicat. 2020, 34, 107553. [Google Scholar] [CrossRef] [PubMed]
- De Mello, V.D.; Dahlman, I.; Lankinen, M.; Kurl, S.; Pitkanen, L.; Laaksonen, D.E.; Schwab, U.S.; Erkkila, A.T. The effect of different sources of fish and camelina sativa oil on immune cell and adipose tissue mRNA expression in subjects with abnormal fasting glucose metabolism: A randomized controlled trial. Nutr. Diabetes 2019, 9, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, R.D.; Xue, C.; Zhang, H.; Tuteja, S.; Li, M.; Reilly, M.P.; Ferguson, J.F. Expression of Calgranulin Genes S100A8, S100A9 and S100A12 Is Modulated by n-3 PUFA during inflammation in adipose tissue and mononuclear cells. PLoS ONE 2017, 12, e0169614. [Google Scholar] [CrossRef] [PubMed]
- Huerta, A.E.; Prieto-Hontoria, P.L.; Fernandez-Galilea, M.; Escote, X.; Martinez, J.A.; Moreno-Aliaga, M.J. Effects of dietary supplementation with EPA and/or alpha-lipoic acid on adipose tissue transcriptomic profile of healthy overweight/obese women following a hypocaloric diet. BioFactors 2017, 43, 117–131. [Google Scholar] [CrossRef]
- Holt, P.R.; Aleman, J.O.; Walker, J.M.; Jiang, C.S.; Liang, Y.; de Rosa, J.C.; Giri, D.D.; Iyengar, N.M.; Milne, G.L.; Hudis, C.A.; et al. Docosahexaenoic acid supplementation is not anti-inflammatory in adipose tissue of healthy obese postmenopausal women. Int. J. Nutr. 2017, 1, 31–49. [Google Scholar] [CrossRef] [Green Version]
- Klingel, S.L.; Valsesia, A.; Astrup, A.; Kunesova, M.; Saris, W.H.M.; Langin, D.; Viguerie, N.; Mutch, D.M. FADS1 genotype is distinguished by human subcutaneous adipose tissue fatty acids, but not inflammatory gene expression. Int. J. Obes. 2019, 43, 1539–1548. [Google Scholar] [CrossRef]
- Yessoufou, A.; Ple, A.; Moutairou, K.; Hichami, A.; Khan, N.A. Docosahexaenoic acid reduces suppressive and migratory functions of CD4+CD25+ regulatory T-cells. J. Lipid Res. 2009, 50, 2377–2388. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Khan, N.A.; McMurray, D.N.; Prior, I.A.; Wang, N.; Chapkin, R.S. Regulatory activity of polyunsaturated fatty acids in T-cell signaling. Prog. Lipid Res. 2010, 49, 250–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Urso, C.J.; Jadeja, V. Saturated fatty acids in obesity-associated inflammation. J. Inflamm. Res. 2020, 13, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radzikowska, U.; Rinaldi, A.O.; Celebi Sozener, Z.; Karaguzel, D.; Wojcik, M.; Cypryk, K.; Akdis, M.; Akdis, C.A.; Sokolowska, M. The influence of dietary fatty acids on immune responses. Nutrients 2019, 11, 2990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulven, S.M.; Holven, K.B.; Rundblad, A.; Myhrstad, M.C.W.; Leder, L.; Dahlman, I.; Mello, V.D.; Schwab, U.; Carlberg, C.; Pihlajamaki, J.; et al. An Isocaloric Nordic Diet Modulates RELA and TNFRSF1A Gene Expression in Peripheral Blood Mononuclear Cells in Individuals with Metabolic Syndrome-A SYSDIET Sub-Study. Nutrients 2019, 11, 2932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouwens, M.; Afman, L.A.; Muller, M. Fasting induces changes in peripheral blood mononuclear cell gene expression profiles related to increases in fatty acid beta-oxidation: Functional role of peroxisome proliferator activated receptor alpha in human peripheral blood mononuclear cells. Am. J. Clin. Nutr. 2007, 86, 1515–1523. [Google Scholar] [CrossRef]
- O’Grada, C.M.; Morine, M.J.; Morris, C.; Ryan, M.; Dillon, E.T.; Walsh, M.; Gibney, E.R.; Brennan, L.; Gibney, M.J.; Roche, H.M. PBMCs reflect the immune component of the WAT transcriptome--implications as biomarkers of metabolic health in the postprandial state. Mol. Nutr. Food Res. 2014, 58, 808–820. [Google Scholar] [CrossRef]
- Ulven, S.M.; Christensen, J.J.; Nygard, O.; Svardal, A.; Leder, L.; Ottestad, I.; Lysne, V.; Laupsa-Borge, J.; Ueland, P.M.; Midttun, O.; et al. Using metabolic profiling and gene expression analyses to explore molecular effects of replacing saturated fat with polyunsaturated fat-a randomized controlled dietary intervention study. Am. J. Clin. Nutr. 2019, 109, 1239–1250. [Google Scholar] [CrossRef]
- Rundblad, A.; Larsen, S.V.; Myhrstad, M.C.; Ottestad, I.; Thoresen, M.; Holven, K.B.; Ulven, S.M. Differences in peripheral blood mononuclear cell gene expression and triglyceride composition in lipoprotein subclasses in plasma triglyceride responders and non-responders to omega-3 supplementation. Genes Nutr. 2019, 14, 10. [Google Scholar] [CrossRef] [Green Version]
- Rudkowska, I.; Paradis, A.M.; Thifault, E.; Julien, P.; Tchernof, A.; Couture, P.; Lemieux, S.; Barbier, O.; Vohl, M.C. Transcriptomic and metabolomic signatures of an n-3 polyunsaturated fatty acids supplementation in a normolipidemic/normocholesterolemic Caucasian population. J. Nutr. Biochem. 2013, 24, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Rudkowska, I.; Ponton, A.; Jacques, H.; Lavigne, C.; Holub, B.J.; Marette, A.; Vohl, M.C. Effects of a supplementation of n-3 polyunsaturated fatty acids with or without fish gelatin on gene expression in peripheral blood mononuclear cells in obese, insulin-resistant subjects. J. Nutr. Nutr. 2011, 4, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Polus, A.; Zapala, B.; Razny, U.; Gielicz, A.; Kiec-Wilk, B.; Malczewska-Malec, M.; Sanak, M.; Childs, C.E.; Calder, P.C.; Dembinska-Kiec, A. Omega-3 fatty acid supplementation influences the whole blood transcriptome in women with obesity, associated with pro-resolving lipid mediator production. Biochim. Biophys. Acta 2016, 1861, 1746–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vedin, I.; Cederholm, T.; Freund-Levi, Y.; Basun, H.; Garlind, A.; Irving, G.F.; Eriksdotter-Jonhagen, M.; Wahlund, L.O.; Dahlman, I.; Palmblad, J. Effects of DHA-rich n-3 fatty acid supplementation on gene expression in blood mononuclear leukocytes: The OmegAD study. PLoS ONE 2012, 7, e35425. [Google Scholar] [CrossRef] [Green Version]
- Bouwens, M.; van de Rest, O.; Dellschaft, N.; Bromhaar, M.G.; de Groot, L.C.; Geleijnse, J.M.; Muller, M.; Afman, L.A. Fish-oil supplementation induces antiinflammatory gene expression profiles in human blood mononuclear cells. Am. J. Clin. Nutr. 2009, 90, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Rahmani, E.; Jamilian, M.; Dadpour, B.; Nezami, Z.; Vahedpoor, Z.; Mahmoodi, S.; Aghadavod, E.; Taghizadeh, M.; Beiki Hassan, A.; Asemi, Z. The effects of fish oil on gene expression in patients with polycystic ovary syndrome. Eur. J. Clin. Investig. 2018, 48. [Google Scholar] [CrossRef]
- Skulas-Ray, A.C.; Kris-Etherton, P.M.; Harris, W.S.; Vanden Heuvel, J.P.; Wagner, P.R.; West, S.G. Dose-response effects of omega-3 fatty acids on triglycerides, inflammation, and endothelial function in healthy persons with moderate hypertriglyceridemia. Am. J. Clin. Nutr. 2011, 93, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Myhrstad, M.C.; Ulven, S.M.; Gunther, C.C.; Ottestad, I.; Holden, M.; Ryeng, E.; Borge, G.I.; Kohler, A.; Bronner, K.W.; Thoresen, M.; et al. Fish oil supplementation induces expression of genes related to cell cycle, endoplasmic reticulum stress and apoptosis in peripheral blood mononuclear cells: A transcriptomic approach. J. Intern. Med. 2014, 276, 498–511. [Google Scholar] [CrossRef] [Green Version]
- Rundblad, A.; Holven, K.B.; Bruheim, I.; Myhrstad, M.C.; Ulven, S.M. Effects of fish and krill oil on gene expression in peripheral blood mononuclear cells and circulating markers of inflammation: A randomised controlled trial. J. Nutr. Sci. 2018, 7, e10. [Google Scholar] [CrossRef] [Green Version]
- Bouwens, M.; Grootte Bromhaar, M.; Jansen, J.; Muller, M.; Afman, L.A. Postprandial dietary lipid-specific effects on human peripheral blood mononuclear cell gene expression profiles. Am. J. Clin. Nutr. 2010, 91, 208–217. [Google Scholar] [CrossRef] [Green Version]
- Zhao, N.; Wang, L.; Guo, N. alpha-Linolenic acid increases the G0/G1 switch gene 2 mRNA expression in peripheral blood mononuclear cells from obese patients: A pilot study. Lipids Health Dis. 2016, 15, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashemzadeh, A.A.; Nasoohi, N.; Raygan, F.; Aghadavod, E.; Akbari, E.; Taghizadeh, M.; Memarzadeh, M.R.; Asemi, Z. Flaxseed oil supplementation improve gene expression levels of PPAR-gamma, LP(a), IL-1 and TNF-alpha in Type 2 diabetic patients with coronary heart disease. Lipids 2017, 52, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Thies, F.; Nebe-von-Caron, G.; Powell, J.R.; Yaqoob, P.; Newsholme, E.A.; Calder, P.C. Dietary supplementation with eicosapentaenoic acid, but not with other long-chain n-3 or n-6 polyunsaturated fatty acids, decreases natural killer cell activity in healthy subjects aged >55 y. Am. J. Clin. Nutr. 2001, 73, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Thies, F.; Nebe-von-Caron, G.; Powell, J.R.; Yaqoob, P.; Newsholme, E.A.; Calder, P.C. Dietary supplementation with gamma-linolenic acid or fish oil decreases T lymphocyte proliferation in healthy older humans. J. Nutr. 2001, 131, 1918–1927. [Google Scholar] [CrossRef] [Green Version]
- Georgiadi, A.; Kersten, S. Mechanisms of gene regulation by fatty acids. Adv. Nutr. 2012, 3, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Afman, L.A.; Muller, M. Human nutrigenomics of gene regulation by dietary fatty acids. Prog. Lipid Res. 2012, 51, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Haversen, L.; Danielsson, K.N.; Fogelstrand, L.; Wiklund, O. Induction of proinflammatory cytokines by long-chain saturated fatty acids in human macrophages. Atherosclerosis 2009, 202, 382–393. [Google Scholar] [CrossRef]
- Kochumon, S.; Wilson, A.; Chandy, B.; Shenouda, S.; Tuomilehto, J.; Sindhu, S.; Ahmad, R. Palmitate Activates CCL4 Expression in Human Monocytic Cells via TLR4/MyD88 Dependent Activation of NF-kappaB/MAPK/ PI3K Signaling Systems. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 46, 953–964. [Google Scholar] [CrossRef]
- Samblas, M.; Carraro, J.C.; Martinez, J.A.; Milagro, F.I. The regulation of inflammation-related genes after palmitic acid and DHA treatments is not mediated by DNA methylation. J. Physiol. Biochem. 2019, 75, 341–349. [Google Scholar] [CrossRef]
- Huang, S.; Rutkowsky, J.M.; Snodgrass, R.G.; Ono-Moore, K.D.; Schneider, D.A.; Newman, J.W.; Adams, S.H.; Hwang, D.H. Saturated fatty acids activate TLR-mediated proinflammatory signaling pathways. J. Lipid Res. 2012, 53, 2002–2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, R.; Al-Roub, A.; Kochumon, S.; Akther, N.; Thomas, R.; Kumari, M.; Koshy, M.S.; Tiss, A.; Hannun, Y.A.; Tuomilehto, J.; et al. The Synergy between Palmitate and TNF-alpha for CCL2 production is dependent on the TRIF/IRF3 Pathway: Implications for metabolic inflammation. J. Immunol. 2018, 200, 3599–3611. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.; Kochumon, S.; Al-Ozairi, E.; Tuomilehto, J.; Ahmad, R. Association between adipose tissue interleukin-33 and immunometabolic markers in individuals with varying degrees of glycemia. Dis. Markers 2019, 2019, 7901062. [Google Scholar] [CrossRef] [Green Version]
- Sindhu, S.; Kochumon, S.; Shenouda, S.; Wilson, A.; Al-Mulla, F.; Ahmad, R. The Cooperative Induction of CCL4 in Human Monocytic Cells by TNF-alpha and Palmitate Requires MyD88 and Involves MAPK/NF-kappaB Signaling Pathways. Int. J. Mol. Sci. 2019, 20, 4658. [Google Scholar] [CrossRef] [Green Version]
- Snodgrass, R.G.; Huang, S.; Choi, I.W.; Rutledge, J.C.; Hwang, D.H. Inflammasome-mediated secretion of IL-1beta in human monocytes through TLR2 activation; modulation by dietary fatty acids. J. Immunol. 2013, 191, 4337–4347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholas, D.A.; Zhang, K.; Hung, C.; Glasgow, S.; Aruni, A.W.; Unternaehrer, J.; Payne, K.J.; Langridge, W.H.R.; De Leon, M. Palmitic acid is a toll-like receptor 4 ligand that induces human dendritic cell secretion of IL-1beta. PLoS ONE 2017, 12, e0176793. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hao, Q.; Li, Q.R.; Yan, X.W.; Ye, S.; Li, Y.S.; Li, N.; Li, J.S. Omega-3 polyunsaturated fatty acids affect lipopolysaccharide-induced maturation of dendritic cells through mitogen-activated protein kinases p38. Nutrition 2007, 23, 474–482. [Google Scholar] [CrossRef]
- Zeyda, M.; Saemann, M.D.; Stuhlmeier, K.M.; Mascher, D.G.; Nowotny, P.N.; Zlabinger, G.J.; Waldhausl, W.; Stulnig, T.M. Polyunsaturated fatty acids block dendritic cell activation and function independently of NF-kappaB activation. J. Biol. Chem. 2005, 280, 14293–14301. [Google Scholar] [CrossRef] [Green Version]
- Hatanaka, E.; Levada-Pires, A.C.; Pithon-Curi, T.C.; Curi, R. Systematic study on ROS production induced by oleic, linoleic, and gamma-linolenic acids in human and rat neutrophils. Free Radic. Biol. Med. 2006, 41, 1124–1132. [Google Scholar] [CrossRef]
- McCambridge, G.; Agrawal, M.; Keady, A.; Kern, P.A.; Hasturk, H.; Nikolajczyk, B.S.; Bharath, L.P. Saturated fatty acid activates t cell inflammation through a nicotinamide nucleotide transhydrogenase (NNT)-dependent mechanism. Biomolecules 2019, 9, 79. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Wang, G.; Lyu, Y.; Wang, L.; Zuo, S.; Zou, J.; Sun, L.; Zhao, W.; Shu, C.; Yang, Y.G.; et al. Upregulation of SLAMF3 on human T cells is induced by palmitic acid through the STAT5-PI3K/Akt pathway and features the chronic inflammatory profiles of type 2 diabetes. Cell Death Dis. 2019, 10, 559. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, C.; Del Mar Cavia, M.; Roelofs, H.; Wanten, G.; Alonso-Torre, S.R. Activation of human neutrophils by oleic acid involves the production of reactive oxygen species and a rise in cytosolic calcium Conc.: A comparison with N-6 polyunsaturated fatty acids. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2011, 28, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Passos, M.E.; Alves, H.H.; Momesso, C.M.; Faria, F.G.; Murata, G.; Cury-Boaventura, M.F.; Hatanaka, E.; Massao-Hirabara, S.; Gorjao, R. Differential effects of palmitoleic acid on human lymphocyte proliferation and function. Lipids Health Dis. 2016, 15, 217. [Google Scholar] [CrossRef] [Green Version]
- Rueda, C.M.; Rodriguez-Perea, A.L.; Moreno-Fernandez, M.; Jackson, C.M.; Melchior, J.T.; Davidson, W.S.; Chougnet, C.A. High density lipoproteins selectively promote the survival of human regulatory T cells. J. Lipid Res. 2017, 58, 1514–1523. [Google Scholar] [CrossRef] [Green Version]
- Moghaddami, N.; Costabile, M.; Grover, P.K.; Jersmann, H.P.; Huang, Z.H.; Hii, C.S.; Ferrante, A. Unique effect of arachidonic acid on human neutrophil TNF receptor expression: Up-regulation involving protein kinase C, extracellular signal-regulated kinase, and phospholipase A2. J. Immunol. 2003, 171, 2616–2624. [Google Scholar] [CrossRef] [PubMed]
- Cury-Boaventura, M.F.; Gorjao, R.; de Lima, T.M.; Newsholme, P.; Curi, R. Comparative toxicity of oleic and linoleic acid on human lymphocytes. Life Sci. 2006, 78, 1448–1456. [Google Scholar] [CrossRef] [PubMed]
- Lux, C.A.; Koschinski, A.; Dersch, K.; Husmann, M.; Bhakdi, S. Hypersusceptibility of neutrophil granulocytes towards lethal action of free fatty acids contained in enzyme-modified atherogenic low density lipoprotein. Atherosclerosis 2009, 207, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Bassal, N.K.; Hughes, B.P.; Costabile, M. Arachidonic acid and its COX1/2 metabolites inhibit interferon-gamma mediated induction of indoleamine-2,3 dioxygenase in THP-1 cells and human monocytes. Prostaglandins Leukot. Essent. Fat. Acids 2012, 87, 119–126. [Google Scholar] [CrossRef]
- Aggarwal, N.; Korenbaum, E.; Mahadeva, R.; Immenschuh, S.; Grau, V.; Dinarello, C.A.; Welte, T.; Janciauskiene, S. alpha-Linoleic acid enhances the capacity of alpha-1 antitrypsin to inhibit lipopolysaccharide induced IL-1beta in human blood neutrophils. Mol. Med. 2016, 22, 680–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Chen, L.Y.; Sokolowska, M.; Eberlein, M.; Alsaaty, S.; Martinez-Anton, A.; Logun, C.; Qi, H.Y.; Shelhamer, J.H. The fish oil ingredient, docosahexaenoic acid, activates cytosolic phospholipase A(2) via GPR120 receptor to produce prostaglandin E(2) and plays an anti-inflammatory role in macrophages. Immunology 2014, 143, 81–95. [Google Scholar] [CrossRef]
- Zhao, Y.; Joshi-Barve, S.; Barve, S.; Chen, L.H. Eicosapentaenoic acid prevents LPS-induced TNF-alpha expression by preventing NF-kappaB activation. J. Am. Coll. Nutr. 2004, 23, 71–78. [Google Scholar] [CrossRef]
- Kawano, A.; Ariyoshi, W.; Yoshioka, Y.; Hikiji, H.; Nishihara, T.; Okinaga, T. Docosahexaenoic acid enhances M2 macrophage polarization via the p38 signaling pathway and autophagy. J. Cell. Biochem. 2019, 120, 12604–12617. [Google Scholar] [CrossRef]
- Allam-Ndoul, B.; Guenard, F.; Barbier, O.; Vohl, M.C. A Study of the Differential Effects of Eicosapentaenoic Acid (EPA) and Docosahexaenoic Acid (DHA) on Gene Expression Profiles of Stimulated Thp-1 Macrophages. Nutrients 2017, 9, 424. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Jiang, W.; Spinetti, T.; Tardivel, A.; Castillo, R.; Bourquin, C.; Guarda, G.; Tian, Z.; Tschopp, J.; Zhou, R. Omega-3 fatty acids prevent inflammation and metabolic disorder through inhibition of NLRP3 inflammasome activation. Immunity 2013, 38, 1154–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marion-Letellier, R.; Butler, M.; Dechelotte, P.; Playford, R.J.; Ghosh, S. Comparison of cytokine modulation by natural peroxisome proliferator-activated receptor gamma ligands with synthetic ligands in intestinal-like Caco-2 cells and human dendritic cells--potential for dietary modulation of peroxisome proliferator-activated receptor gamma in intestinal inflammation. Am. J. Clin. Nutr. 2008, 87, 939–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zapata-Gonzalez, F.; Rueda, F.; Petriz, J.; Domingo, P.; Villarroya, F.; Diaz-Delfin, J.; de Madariaga, M.A.; Domingo, J.C. Human dendritic cell activities are modulated by the omega-3 fatty acid, docosahexaenoic acid, mainly through PPAR(gamma):RXR heterodimers: Comparison with other polyunsaturated fatty acids. J. Leukoc. Biol. 2008, 84, 1172–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajnavolgyi, E.; Laczik, R.; Kun, V.; Szente, L.; Fenyvesi, E. Effects of RAMEA-complexed polyunsaturated fatty acids on the response of human dendritic cells to inflammatory signals. Beilstein J. Org. Chem. 2014, 10, 3152–3160. [Google Scholar] [CrossRef]
- Sethi, S.; Ziouzenkova, O.; Ni, H.; Wagner, D.D.; Plutzky, J.; Mayadas, T.N. Oxidized omega-3 fatty acids in fish oil inhibit leukocyte-endothelial interactions through activation of PPAR alpha. Blood 2002, 100, 1340–1346. [Google Scholar] [CrossRef] [Green Version]
- Sundqvist, M.; Christenson, K.; Holdfeldt, A.; Gabl, M.; Martensson, J.; Bjorkman, L.; Dieckmann, R.; Dahlgren, C.; Forsman, H. Similarities and differences between the responses induced in human phagocytes through activation of the medium chain fatty acid receptor GPR84 and the short chain fatty acid receptor FFA2R. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 695–708. [Google Scholar] [CrossRef]
- Sureda, A.; Martorell, M.; Bibiloni, M.D.M.; Bouzas, C.; Gallardo-Alfaro, L.; Mateos, D.; Capo, X.; Tur, J.A.; Pons, A. Effect of free fatty acids on inflammatory gene expression and hydrogen peroxide production by ex vivo blood mononuclear cells. Nutrients 2020, 12, 146. [Google Scholar] [CrossRef] [Green Version]
- Jaudszus, A.; Gruen, M.; Watzl, B.; Ness, C.; Roth, A.; Lochner, A.; Barz, D.; Gabriel, H.; Rothe, M.; Jahreis, G. Evaluation of suppressive and pro-resolving effects of EPA and DHA in human primary monocytes and T-helper cells. J. Lipid Res. 2013, 54, 923–935. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Bates, E.J. Eicosanoids, fatty acids and neutrophils: Their relevance to the pathophysiology of disease. Prostaglandins Leukot. Essent. Fat. Acids 1995, 53, 75–86. [Google Scholar] [CrossRef]
- Mauro, C.; Smith, J.; Cucchi, D.; Coe, D.; Fu, H.; Bonacina, F.; Baragetti, A.; Cermenati, G.; Caruso, D.; Mitro, N.; et al. Obesity-induced metabolic stress leads to biased effector memory CD4(+) T Cell Differentiation via PI3K p110delta-akt-mediated signals. Cell Metab. 2017, 25, 593–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurtado, C.G.; Wan, F.; Housseau, F.; Sears, C.L. Roles for Interleukin 17 and adaptive immunity in pathogenesis of colorectal cancer. Gastroenterology 2018, 155, 1706–1715. [Google Scholar] [CrossRef]
- Conti, L.; Del Corno, M.; Gessani, S. Revisiting the impact of lifestyle on colorectal cancer risk in a gender perspective. Crit. Rev. Oncol. Hematol. 2020, 145, 102834. [Google Scholar] [CrossRef]
- Currie, E.; Schulze, A.; Zechner, R.; Walther, T.C.; Farese, R.V., Jr. Cellular fatty acid metabolism and cancer. Cell Metab. 2013, 18, 153–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balaban, S.; Shearer, R.F.; Lee, L.S.; van Geldermalsen, M.; Schreuder, M.; Shtein, H.C.; Cairns, R.; Thomas, K.C.; Fazakerley, D.J.; Grewal, T.; et al. Adipocyte lipolysis links obesity to breast cancer growth: Adipocyte-derived fatty acids drive breast cancer cell proliferation and migration. Cancer Metab. 2017, 5, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellino, G.; Gallo, G.; Pallante, P.; Capasso, R.; De Stefano, A.; Maretto, I.; Malapelle, U.; Qiu, S.; Nikolaou, S.; Barina, A.; et al. Noninvasive biomarkers of colorectal cancer: Role in diagnosis and personalised treatment perspectives. Gastroenterol. Res. Pract. 2018, 2018, 2397863. [Google Scholar] [CrossRef] [PubMed]
- Volpato, M.; Hull, M.A. Omega-3 polyunsaturated fatty acids as adjuvant therapy of colorectal cancer. Cancer Metastasis Rev. 2018, 37, 545–555. [Google Scholar] [CrossRef] [Green Version]
- Notarnicola, M.; Lorusso, D.; Tutino, V.; De Nunzio, V.; De Leonardis, G.; Marangelli, G.; Guerra, V.; Veronese, N.; Caruso, M.G.; Giannelli, G. Differential tissue fatty acids profiling between colorectal cancer patients with and without synchronous metastasis. Int. J. Mol. Sci. 2018, 19, 962. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Dietary Compound/FA | Subjects | Treatment Duration and Doses | Genes/Pathways Targeted | TFs Involved | Reference |
|---|---|---|---|---|---|
| ω3 PUFA | Metabolically healthy overweight and obese subjects | 6 weeks 3 g/day | ↓Oxidative stress response | NRF2, PPAR-α, HIF, NF-κB | [82] |
| ω3 PUFA | Insulin-resistant obese subjects | 8 weeks 1.8 g/day | NRF2, PPAR-α, HIF, NF-κB | [83] | |
| ω3 PUFA | Obese women | 3 months | ↑Lipid metabolism ↑Antioxidant enzymes | ↑PPAR-α, NRF2 | [84] |
| FO | Alzheimer disease subjects | 6 months 1.7 g/day DHA, 0.6 g/day EPA | ↓Neuro-inflammation | [85] | |
| FO | Elderly subjects | 26 weeks 0.4–1.8 g/day EPA + DHA | ↓Eicosanoid synthesis ↓TLR/Interleukin/MAPK signalling ↓Oxidative stress ↓Cell adhesion ↓Hypoxia signalling | ↓NF-κB ↑PPAR/LXR/RXR | [86] |
| HOSO | 26 weeks | 4.0 g/day ↓Eicosanoid synthesis ↓Interleukin/MAPK signalling | |||
| FO | PCOS women | 12 weeks 2.0 g/day | ↓IL-1, CXCL8 | ↑PPAR-γ | [87] |
| EPA + DHA | Healthy subjects with moderate hypertriglyceridemia | 8 weeks 0.85 g/day and 3.4 g/day | No effects on inflammatory and endothelial function | [88] | |
| FO | CHD patients | 8 weeks | No effects on inflammatory and endothelial function | [68] | |
| FO | Healthy lean subjects | 7 weeks 8 g/day | ↑ER stress response ↑Apoptosis ↑Cell cycle regulation ↑Antioxidant response | ↑ 35 TF (i.e., ATF4, MIF1, E2F, TP53, STAT1, FOXO4, SP1, NRF2) | [89] |
| Krill oil | Healthy subjects | 8 weeks 4 g/day | ↓Glucose metabolism ↓Lipid metabolism ↓Inflammation ↓β-oxidation | ↓ SREBF2 ↓LXRα | [90] |
| HOSO | 8 weeks | 4 g/day ↓Lipid metabolism ↓Inflammation | ↓LXRα ↓PPAR-δ | ||
| PUFA (40% DHA) | Healthy males | 6 h | ↓Inflammation ↓LXR signalling ↑Cellular stress response | ↓LXR ↓SREBF1 | [91] |
| SFA | ↑Inflammation | ↑LXR signalling ↓Cellular stress response ↑LXR ↑SREBF1 | |||
| CSO | Impaired fasting glucose subjects | 12 weeks | ↓Inflammation ↓IFNG | ↑PPAR-γ | [68] |
| ALA | Obese subject | 12 weeks 4.0 g/day | ↓Inflammation | ↑PPAR-γ | [92] |
| Flaxseed oil | T2DM subjects with CHD | 12 weeks 400 mg ALA twice a day | ↓Inflammation | ↑PPAR-γ | [93] |
| ω6 PUFA | Healthy subjects | 8 weeks 12.9 g/day | ↓Lipid metabolism (i.e., LDLR, ABCG, SREBF1, and FASN) ↓Inflammation (i.e., IRAK1, TNFSF1, TLR4, GATA3, IL2RG, and CD8A) |
↓PPAR-δ ↓LXRA | [80] |
| Dietary Compound/ FA | Cell Type | Downstream Effects | Pathway(s) Targeted | TFs Involved | Reference |
|---|---|---|---|---|---|
| SFA | |||||
| PA | THP-1 Primary monocytes | ↑IL-1, IL-6, IL-18, TNFα, CCL2, CCL4, CXCL8 | ↑TLR4/MyD88/MAPK | ↑NF-kB, AP-1 | [98,99,100,101] |
| PA (in combination with TNFα) | THP-1 Primary monocytes/macrophages | ↑CCL2, CCL3, CCL4, CXCL8 | ↑TLR4/TRIF | ↑NF-kB, AP-1, IRF3 | [102,103,104] |
| PA | THP-1 | ↑IL-1β | ↑TLR2/TLR1 ↑NLRP3 inflammasome | [105] | |
| PA | MDDC | ↑IL-1β ↑ROS ↑Activation/maturation | ↑TLR4/MD-2 ↑NLRC4 Inflammasome | ↑NF-kB | [106] |
| SFA | MDDC | No effects on inflammation | [107,108] | ||
| OA, LA, GLA | Neutrophils | ↑ROS | [109] | ||
| PA | PBMC | ↓NNT ↑ROS ↑Th17-type cytokines | [110] | ||
| PA | Naïve T lymphocytes | ↑SLAMF3 | ↑PI3K/AKT | ↑STAT5 | [111] |
| MUFA | |||||
| OA | Neutrophils | ↑Phagocytosis and killing ↑ROS ↑IL-1β, CXCL3, VEGF | ↑Intracellular calcium ↑PKC | [109,112] | |
| OA | T lymphocytes |
↓IFNγ ↑IL-4, IL-10 | [113] | ||
| POA | ↓ConA-induced T lymphocyte proliferation | ↓Treg cell differentiation ↓IL-2, IL-6, IFNγ, TNFα, IL-17A ↓CD28 externalization | ↓NFAT, AP-1, NF-κB | ||
| Albumin-bound OA | PBMC-sorted Treg cells | ↑Treg cell survival | [114] | ||
| ω6 PUFA | |||||
| AA | Neutrophils | ↑ROS ↑TNFR1, TNFR2 | ↑Intracellular calcium ↑PKC, ERK1/2, cPLAP2 | [112,115] | |
| ω6 PUFA | Neutrophils | ↓ATP ↑LDH release ↑Mitochondria depolarization | [116,117] | ||
| AA | THP-1 Primary monocytes | ↓IFNγ signalling pathway ↓IDO | ↓STAT1 phosphorylation | [118] | |
| AA | Differentiating MDDC | ↓CD40, CD80, MR ↓LPS-induced CD40, CD80, CD83, CD86 ↓IL-12p40, TNFα ↓T cell proliferation ↓IL-2/IFNγ in co-cultured T cells | NF-κB independent | [108] | |
| A1AT-LA | Neutrophils | Anti-inflammatory | ↓IL-1β | ↑PPAR-γ | [119] |
| ω3 PUFA | |||||
| EPA/DHA | THP-1 | ↓IL-1β, IL-18, TNFα, PAI-1 | ↓NF-κB | [62,100,120,121] | |
| EPA/DHA | U937 | M2 polarization | ↑p38 MAPK | ↑KLF4 | [122] |
| EPA/DHA | THP-1 | ↓LPS-induced cytokine gene expression (i.e., IL6, TNFα, IL1β, MCP1, TNFAIP3, and PTGS2) | [123] | ||
| DHA | MDM | Anti-inflammatory | Via GPR120 ↑cPLAP2 ↑PGE2 signalling | [120] | |
| EPA/DHA | THP-1 | Anti-inflammatory | Via GPR120/GPR40/β-Arrestin-2 ↓NLRP3 inflammasome | [62,124] | |
| EPA/DHA | MDDC | ↓Cell activation and cytokine production ↓MHC-II, CD40, CD80, CD86, CD83 ↓TNFα, IL-10, IL-12 ↓IL-2/IFNγ in co-cultured T cells ↓T cell stimulation | ↓p38 MAPK | ↑PPAR-γ | [107,125] |
| DHA | Differentiating MDDC | ↑MHC-II ↓IL-12p70, IL-6, IL-10 ↓T cell stimulation | PPAR:RXR | [126] | |
| EPA/DHA/ ALA | MDDC | ↓CD1a+ cells ↓IL-6 | ↓GPR120 | [127] | |
| EPA | Differentiating MDDC | ↓CD40, CD80, MR ↓LPS-induced CD40, CD80, CD83, CD86 ↓IL-12p40, TNFα ↓T cell proliferation ↓IL-2/IFNγ in co-cultured T cells | NF-κB independent | [108] | |
| Oxidized EPA | Monocytes Neutrophils | ↓Adhesion to endothelial cells | ↑PPAR-α | [128] | |
| ω3 PUFA | Neutrophils | ↑Chemotaxis, ROS production, NADPH-oxidase activation | ↑GPR84 | [129] | |
| OA/ALA/DHA | PBMC | ↓LPS-induced inflammatory genes (IL-6, TLR2, TNFα, COX2) | ↓NF-κB | [130] | |
| EPA/DHA | PBMC | ↓CD4+ T lymphocyte produced IL-2, TNFα, IL-4 | ↑PPAR-γ | [131] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Cornò, M.; Varì, R.; Scazzocchio, B.; Varano, B.; Masella, R.; Conti, L. Dietary Fatty Acids at the Crossroad between Obesity and Colorectal Cancer: Fine Regulators of Adipose Tissue Homeostasis and Immune Response. Cells 2021, 10, 1738. https://doi.org/10.3390/cells10071738
Del Cornò M, Varì R, Scazzocchio B, Varano B, Masella R, Conti L. Dietary Fatty Acids at the Crossroad between Obesity and Colorectal Cancer: Fine Regulators of Adipose Tissue Homeostasis and Immune Response. Cells. 2021; 10(7):1738. https://doi.org/10.3390/cells10071738
Chicago/Turabian StyleDel Cornò, Manuela, Rosaria Varì, Beatrice Scazzocchio, Barbara Varano, Roberta Masella, and Lucia Conti. 2021. "Dietary Fatty Acids at the Crossroad between Obesity and Colorectal Cancer: Fine Regulators of Adipose Tissue Homeostasis and Immune Response" Cells 10, no. 7: 1738. https://doi.org/10.3390/cells10071738
APA StyleDel Cornò, M., Varì, R., Scazzocchio, B., Varano, B., Masella, R., & Conti, L. (2021). Dietary Fatty Acids at the Crossroad between Obesity and Colorectal Cancer: Fine Regulators of Adipose Tissue Homeostasis and Immune Response. Cells, 10(7), 1738. https://doi.org/10.3390/cells10071738