
Adipose-Derived Stem Cell Features and MCF-7

 , , , , , ,
, , , , , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Adipose-Derived Stem Cell Isolation and Culturing
2.2. MCF-7 Cell Culturing
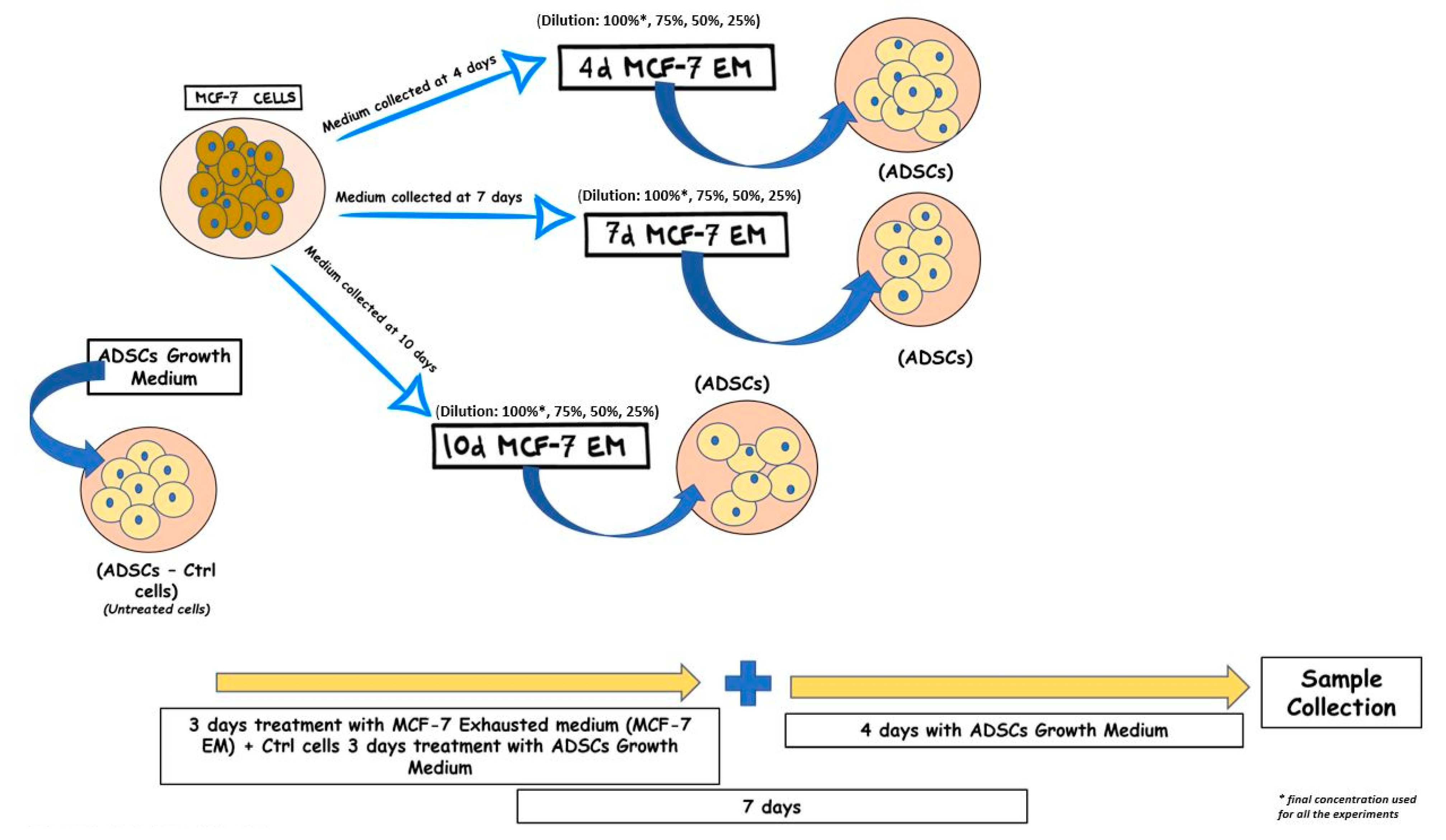
2.3. ADSC Treatment
2.4. BrdU Cell Proliferation Assay
2.5. MTT Viability Assay
2.6. RNA Extraction and Real-Time Quantitative Polymerase Chain
2.7. Autophagosome Detection Assays
2.8. Polyamine Analysis
2.8.1. Chemicals
2.8.2. LC-HRMS
2.8.3. Sample Preparation
2.9. Statistical Analysis
3. Results
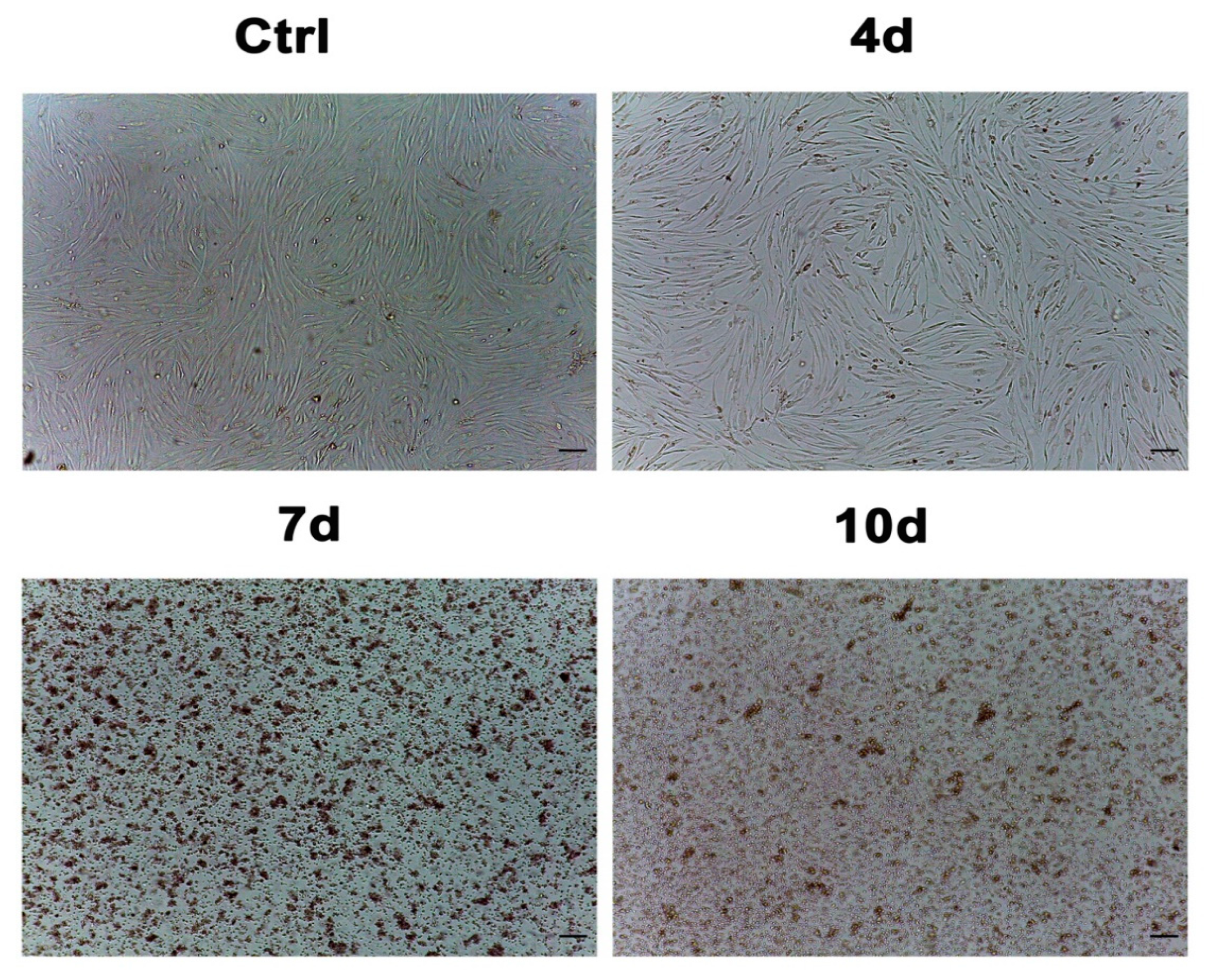
3.1. Morphological Analysis of ADSCs Exposed to MCF7-Exhausted Medium
3.2. hADSCs Exposed to Exhausted Medium Undergo Cell-Cycle Progression through G1/S Phase
3.3. Exhausted Medium Does Not Affect Cell Viability
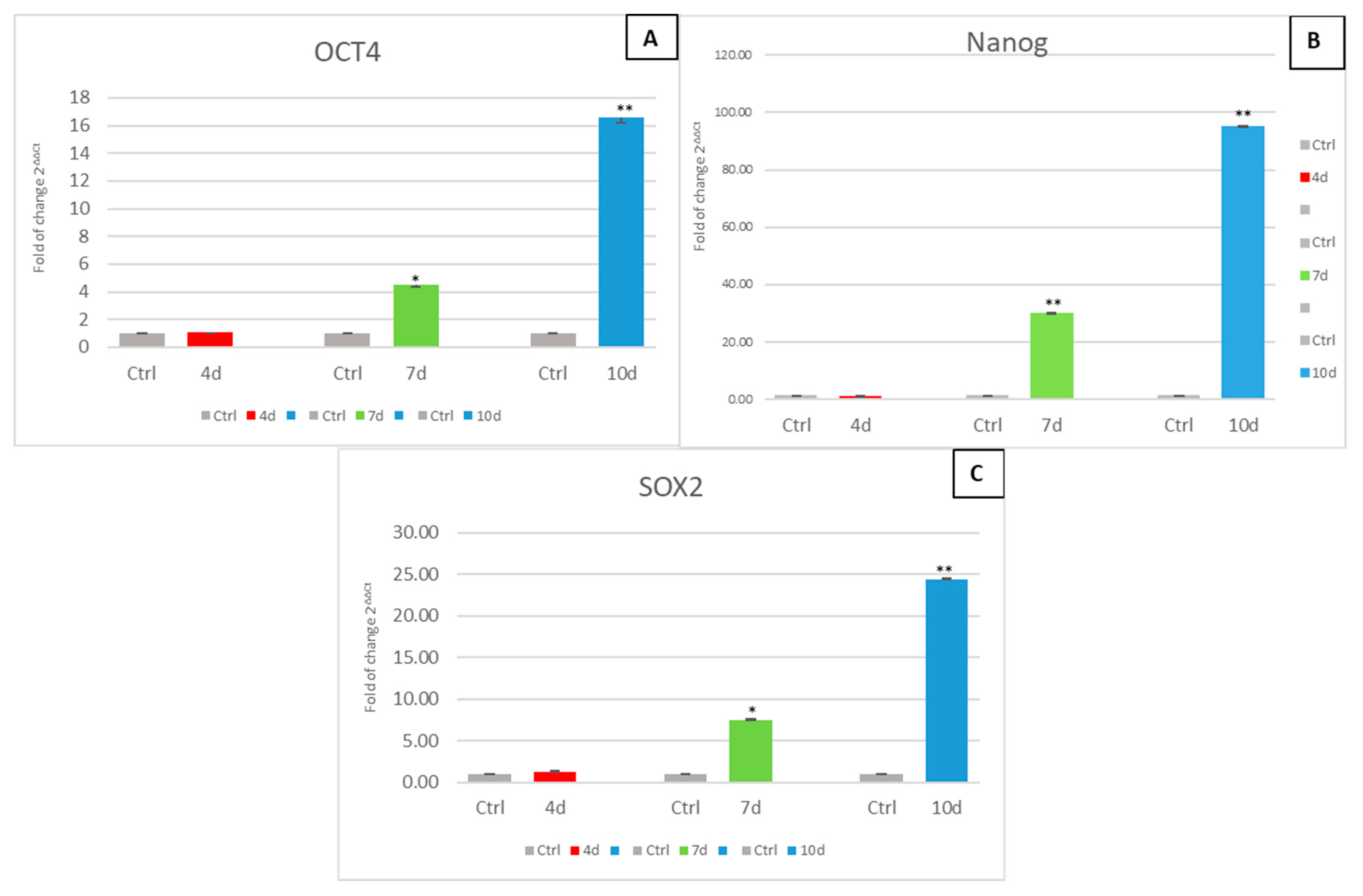
3.4. Exposure of hADSCs to Exhausted MCF- 7 Medium Increases Stem Cell Potency
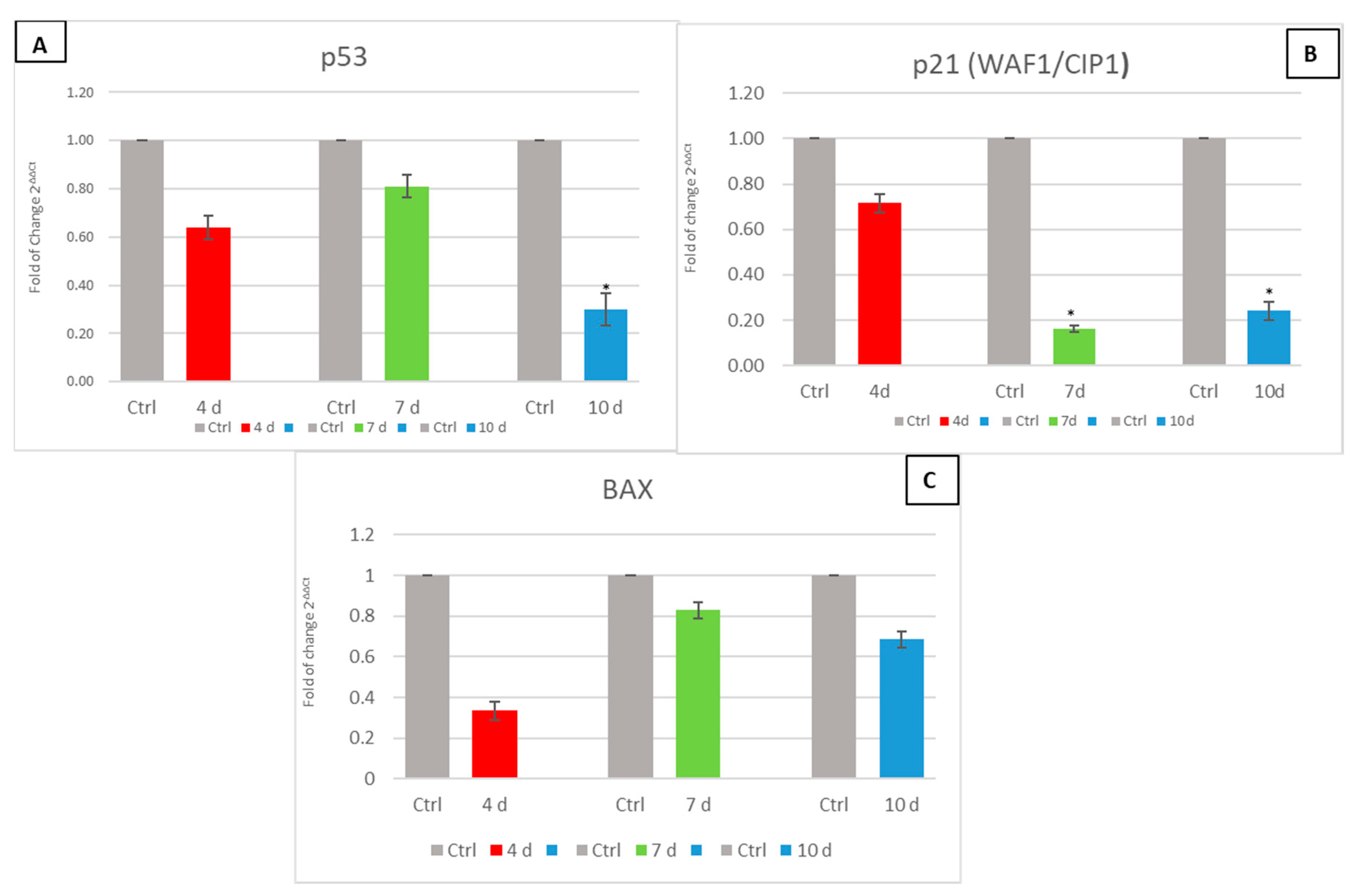
3.5. Exhausted MCF- 7 Medium Influence on p53, p21 (WAF1/CIP1) and BAX
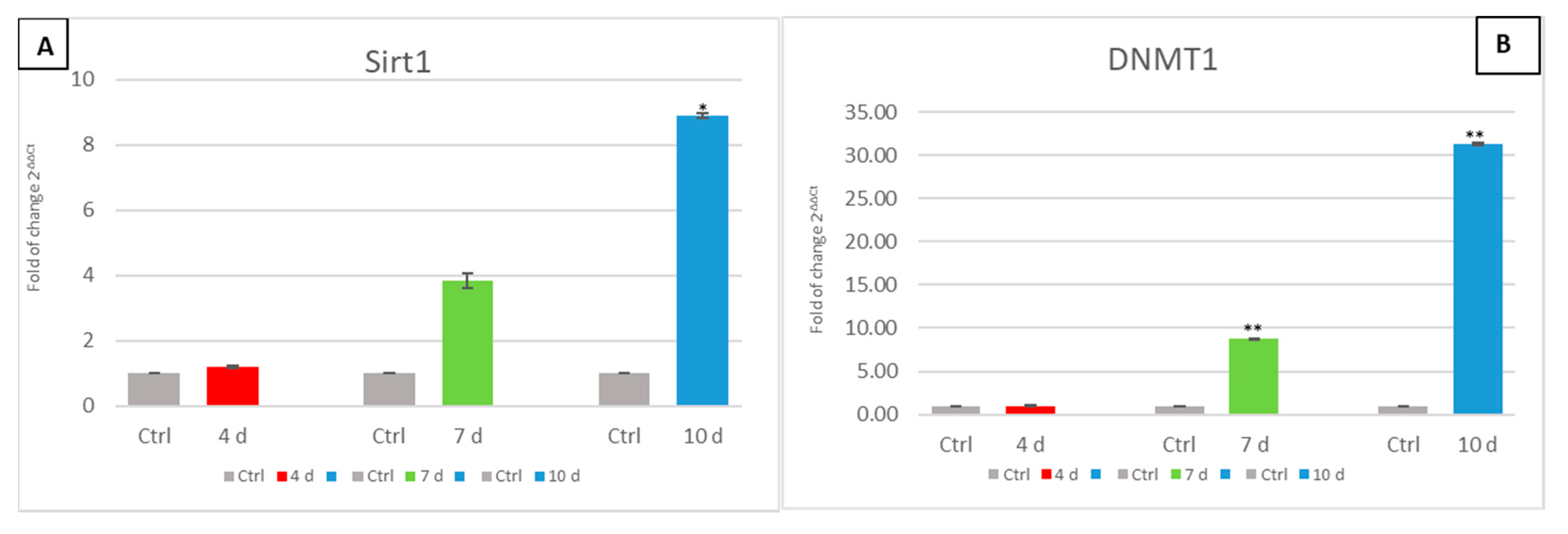
3.6. Exhausted MCF- 7 Medium Affects Epigenetic Modulating Genes in hADSCs
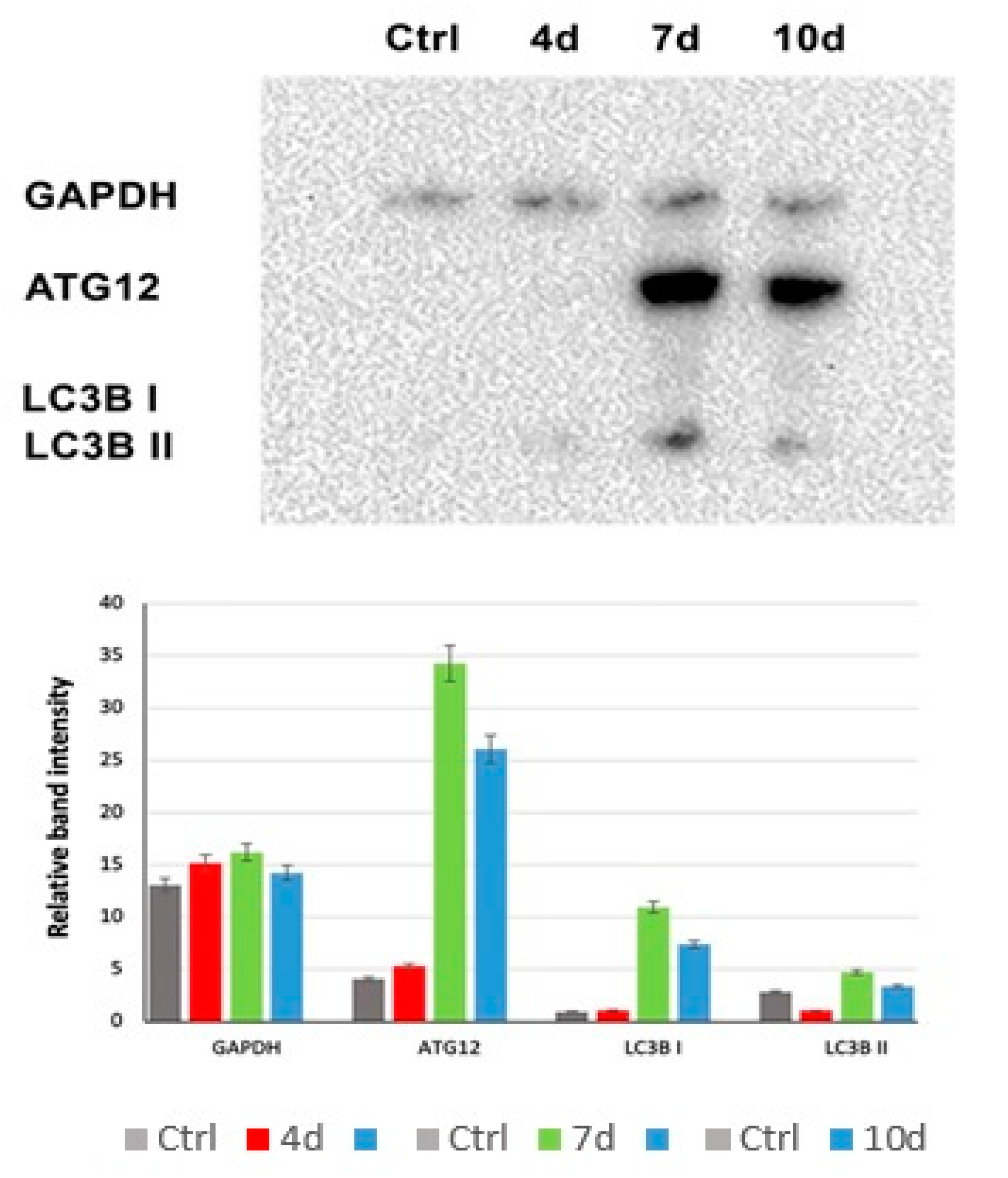
3.7. MCF7-Exhausted Media Induce Autophagy in hADSCS
3.8. Polyamine Detection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Naderi, N.; Combellack, E.J.; Griffin, M.; Sedaghati, T.; Javed, M.; Findlay, M.W.; Wallace, C.G.; Mosahebi, A.; Butler, P.E.; Seifalian, A.M.; et al. The regenerative role of adipose-derived stem cells (ADSC) in plastic and reconstructive surgery. Int. Wound J. 2017, 14, 112–124. [Google Scholar] [CrossRef]
- Wankhade, U.D.; Shen, M.; Kolhe, R.; Fulzele, S. Advances in adipose-derived stem cells isolation, characterization, and application in regenerative tissue engineering. Stem Cells Int. 2016, 2016, 3206807. [Google Scholar] [CrossRef] [Green Version]
- Leto Barone, A.A.; Khalifian, S.; Lee, W.P.; Brandacher, G. Immunomodulatory effects of adipose-derived stem cells: Fact or fiction? Biomed Res. Int. 2013, 2013, 383685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrow, K.L.; Hoyland, J.A.; Richardson, S.M. Human adipose-derived stem cells exhibit enhanced proliferative capacity and retain multipotency longer than donor-matched bone marrow mesenchymal stem cells during expansion In Vitro. Stem Cells Int. 2017, 2017, 2541275. [Google Scholar] [CrossRef]
- Basoli, V.; Santaniello, S.; Cruciani, S.; Ginesu, G.C.; Cossu, M.L.; Delitala, A.P.; Serra, P.A.; Ventura, C.; Maioli, M. Melatonin and vitamin D interfere with the adipogenic fate of adipose-derived stem cells. Int. J. Mol. Sci. 2017, 18, 981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santaniello, S.; Cruciani, S.; Basoli, V.; Balzano, F.; Bellu, E.; Garroni, G.; Ginesu, G.C.; Cossu, M.L.; Facchin, F.; Delitala, A.P.; et al. Melatonin and vitamin D orchestrate adipose derived stem cell fate by modulating epigenetic regulatory genes. Int. J. Med. Sci. 2018, 15, 1631–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruciani, S.; Garroni, G.; Balzano, F.; Pala, R.; Bellu, E.; Cossu, M.L.; Ginesu, G.C.; Ventura, C.; Maioli, M. Tuning adipogenic differentiation in ADSCs by metformin and vitamin D: Involvement of miRNAs. Int. J. Mol. Sci. 2020, 21, 6181. [Google Scholar] [CrossRef]
- Guilak, F.; Cohen, D.M.; Estes, B.T.; Gimble, J.M.; Liedtke, W.; Chen, C.S. Control of stem cell fate by physical interactions with the extracellular matrix. Cell Stem Cell. 2009, 5, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Maioli, M.; Rinaldi, S.; Pigliaru, G.; Santaniello, S.; Basoli, V.; Castagna, A.; Fontani, V.; Ventura, C. REAC technology and hyaluron synthase 2, an interesting network to slow down stem cell senescence. Sci. Rep. 2016, 6, 28682. [Google Scholar] [CrossRef] [Green Version]
- Maioli, M.; Santaniello, S.; Montella, A.; Bandiera, P.; Cantoni, S.; Cavallini, C.; Bianchi, F.; Lionetti, V.; Rizzolio, F.; Marchesi, I.; et al. Hyaluronan esters drive Smad gene expression and signaling enhancing cardiogenesis in mouse embryonic and human mesenchymal stem cells. PLoS ONE 2010, 5, e15151. [Google Scholar] [CrossRef]
- Chambers, I.; Tomlinson, S.R. The transcriptional foundation of pluripotency. Development 2009, 136, 2311–2322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, G.; Jin, Y. Role of Oct4 in maintaining and regaining stem cell pluripotency. Stem Cell Res. Ther. 2010, 1, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.M.; Han, S.H.; Coh, Y.R.; Jang, G.; Ra, J.C.; Kang, S.K.; Lee, H.W.; Youn, H.Y. Enhanced proliferation and differentiation of Oct4- and Sox2-overexpressing human adipose tissue mesenchymal stem cells. Exp. Mol. Med. 2014, 46, e101. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, K.; Lin, B.; Zhao, M.; Yang, X.; Chen, M.; Gao, A.; Liu, F.; Que, J.; Lan, X. The multiple roles for Sox2 in stem cell maintenance and tumorigenesis. Cell. Signal. 2013, 25, 1264–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, A.X.; Liu, C.J.; Sun, Z.Q.; Wei, Z. NANOG: A promising target for digestive malignant tumors. World J. Gastroenterol. 2014, 20, 13071–13078. [Google Scholar] [CrossRef]
- Wang, M.L.; Chiou, S.H.; Wu, C.W. Targeting cancer stem cells: emerging role of Nanog transcription factor. Onco. Targets Ther. 2013, 6, 1207–1220. [Google Scholar] [CrossRef] [Green Version]
- Seymour, T.; Twigger, A.-J.; Kakulas, F. Pluripotency genes and their functions in the normal and aberrant breast and brain. Int. J. Mol. Sci. 2015, 16, 27288–27301. [Google Scholar] [CrossRef] [Green Version]
- Ling, G.Q.; Chen, D.B.; Wang, B.Q.; Zhang, L.S. Expression of the pluripotency markers Oct3/4, Nanog and Sox2 in human breast cancer cell lines. Oncol. Lett. 2012, 4, 1264–1268. [Google Scholar] [CrossRef] [Green Version]
- You, L.; Guo, X.; Huang, Y. Correlation of cancer stem-cell markers OCT4, SOX2, and NANOG with clinicopathological features and prognosis in operative patients with rectal cancer. Yonsei Med. J. 2018, 59, 35–42. [Google Scholar] [CrossRef]
- Sharma, S.; Kelly, T.K.; Jones, P.A. Epigenetics in cancer. Carcinogenesis 2010, 31, 27–36. [Google Scholar] [CrossRef]
- Montalbán-Loro, R.; Domingo-Muelas, A.; Bizy, A.; Ferrón, S.R. Epigenetic regulation of stemness maintenance in the neurogenic niches. World J. Stem Cells 2015, 7, 700–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Kim, J.; Yuan, X.; Braun, T. Epigenetic modifications of stem cells: A paradigm for the control of cardiac progenitor cells. Circ. Res. 2011, 109, 1067–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, R.M.; Fernandez, A.F.; Fraga, M.F. Role of sirtuins in stem cell differentiation. Genes Cancer 2013, 4, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Costello, J. DNA methylation: An epigenetic mark of cellular memory. Exp. Mol. Med. 2017, 49, e322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathania, R.; Ramachandran, S.; Elangovan, S.; Padia, R.; Yang, P.; Cinghu, S.; Veeranan-Karmegam, R.; Arjunan, P.; Gnana-Prakasam, J.P.; Sadanand, F.; et al. DNMT1 is essential for mammary and cancer stem cell maintenance and tumorigenesis. Nat. Commun. 2015, 6, 6910. [Google Scholar] [CrossRef]
- Jin, X.; Wei, Y.; Xu, F.; Zhao, M.; Dai, K.; Shen, R.; Yang, S.; Zhang, N. SIRT1 promotes formation of breast cancer through modulating Akt activity. J. Cancer 2018, 9, 2012–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Halloran, N.; Khan, S.; Gilligan, K.; Dwyer, R.; Kerin, M.; Lowery, A. Oncological risk in autologous stem cell donation for novel tissue-engineering approaches to postmastectomy breast regeneration. breast cancer (Auckl). Breast Cancer Basic Clin. Res. 2019, 13, 1178223419864896. [Google Scholar] [CrossRef]
- Sistigu, A.; Musella, M.; Galassi, C.; Vitale, I.; De Maria, R. Tuning cancer fate: Tumor microenvironment’s role in cancer stem cell quiescence and reawakening. Front. Immunol. 2020, 11, 2166. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, J.; Zhang, L.; Wei, F.; Lian, Y.; Wu, Y.; Gong, Z.; Zhang, S.; Zhou, J.; Cao, K.; et al. Role of tumor microenvironment in tumorigenesis. J. Cancer 2017, 8, 761–773. [Google Scholar] [CrossRef]
- Cozzo, A.J.; Fuller, A.M.; Makowski, L. Contribution of adipose tissue to development of cancer. Compr. Physiol. 2017, 8, 237–282. [Google Scholar] [CrossRef]
- Dominiak, A.; Chełstowska, B.; Olejarz, W.; Nowicka, G. Communication in the cancer microenvironment as a target for therapeutic interventions. Cancers 2020, 12, 1232. [Google Scholar] [CrossRef] [PubMed]
- Wallace, H.M.; Huang, Y.; Marton, L.J.; Woster, P.M.; Casero, R.A. Polyamine analogues targeting epigenetic gene regulation. Essays Biochem. 2009, 46, 95–110. [Google Scholar] [CrossRef]
- Pegg, A.E. Functions of polyamines in mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borzì, R.M.; Guidotti, S.; Minguzzi, M.; Facchini, A.; Platano, D.; Trisolino, G.; Filardo, G.; Cetrullo, S.; D’Adamo, S.; Stefanelli, C.; et al. Polyamine delivery as a tool to modulate stem cell differentiation in skeletal tissue engineering. Amino Acids 2014, 46, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Coradduzza, D.; Azara, E.; Medici, S.; Arru, C.; Solinas, T.; Madonia, M.; Zinellu, A.; Carru, C. A preliminary study procedure for detection of polyamines in plasma samples as a potential diagnostic tool in prostate cancer. J. Chromatogr. B. 2021, 1162, 122468. [Google Scholar] [CrossRef]
- Balzano, F.; Garroni, G.; Cruciani, S.; Bellu, E.; Dei Giudici, S.; Oggiano, A.; Capobianco, G.; Dessole, S.; Ventura, C.; Maioli, M. Behavioral changes in stem-cell potency by HepG2-exhausted medium. Cells 2020, 9, 1890. [Google Scholar] [CrossRef] [PubMed]
- Glick, D.; Barth, S.; Macleod, K.F. Autophagy: Cellular and molecular mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Debnath, J. Autophagy and tumorigenesis. FEBS Lett. 2010, 584, 1427–1435. [Google Scholar] [CrossRef] [Green Version]
- Connors, S.K.; Goodman, M.S.; Myckatyn, T.; Margenthaler, J.; Gehlert, S. Breast reconstruction after mastectomy at a comprehensive cancer center. SpringerPlus 2016, 5, 955. [Google Scholar] [CrossRef] [Green Version]
- Qin, Q.; Tan, Q.; Lian, B.; Mo, Q.; Huang, Z.; Wei, C. Postoperative outcomes of breast reconstruction after mastectomy: A retrospective study. Medicine 2018, 97, e9766. [Google Scholar] [CrossRef] [PubMed]
- Mehrabani, D.; Mehrabani, G.; Zare, S.; Manafi, A. Adipose-derived stem cells (ADSC) and aesthetic surgery: A mini review. World J. Plast. Surg. 2013, 2, 65–70. [Google Scholar] [PubMed]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef]
- Comşa, Ş.; Cîmpean, A.M.; Raica, M. The story of MCF-7 breast cancer cell line: 40 years of experience in research. Anticancer Res. 2015, 35, 3147–3154. [Google Scholar] [PubMed]
- Chen, K.; Satlof, L.; Stoffels, G.; Kothapalli, U.; Ziluck, N.; Lema, M.; Poretsky, L.; Avtanski, D. Cytokine secretion in breast cancer cells—MILLIPLEX assay data. Data Brief 2019, 28, 104798. [Google Scholar] [CrossRef]
- Tan, A.A.; Phang, W.M.; Gopinath, S.C.; Hashim, O.H.; Kiew, L.V.; Chen, Y. Revealing glycoproteins in the secretome of MCF-7 human breast cancer cells. BioMed Res. Int. 2015, 2015, 453289. [Google Scholar] [CrossRef]
- Visweswaran, M.; Keane, K.N.; Arfuso, F.; Dilley, R.J.; Newsholme, P.; Dharmarajan, A. The influence of breast tumour-derived factors and Wnt antagonism on the transformation of adipose-derived mesenchymal stem cells into tumour-associated fibroblasts. Cancer Microenviron. 2018, 11, 71–84. [Google Scholar] [CrossRef]
- Cho, J.A.; Park, H.; Lim, E.H.; Lee, K.W. Exosomes from breast cancer cells can convert adipose tissue-derived mesenchymal stem cells into myofibroblast-like cells. Int. J. Oncol. 2012, 40, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Paunescu, V.; Bojin, F.M.; Tatu, C.A.; Gavriliuc, O.I.; Rosca, A.; Gruia, A.T.; Tanasie, G.; Bunu, C.; Crisnic, D.; Gherghiceanu, M.; et al. Tumour-associated fibroblasts and mesenchymal stem cells: More similarities than differences. J. Cell. Mol. Med. 2011, 15, 635–646. [Google Scholar] [CrossRef]
- Whitaker, R.H.; Placzek, W.J. Regulating the BCL2 family to improve sensitivity to microtubule targeting agents. Cells 2019, 8, 346. [Google Scholar] [CrossRef] [Green Version]
- Zhou, N.; Lin, X.; Dong, W.; Huang, W.; Jiang, W.; Lin, L.; Qiu, Q.; Zhang, X.; Shen, J.; Song, Z.; et al. SIRT1 alleviates senescence of degenerative human intervertebral disc cartilage endo-plate cells via the p53/p21 pathway. Sci. Rep. 2016, 6, 22628. [Google Scholar] [CrossRef] [Green Version]
- Fujino, T.; Yokokawa, R.; Oshima, T.; Hayakawa, M. SIRT1 knockdown up-regulates p53 and p21/Cip1 expression in renal adenocarcinoma cells but not in normal renal-derived cells in a deacetylase-independent manner. J. Toxicol. Sci. 2018, 43, 711–715. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Ren, L.; Cai, Q.; Zou, Y.; Fu, Q.; Ma, Y. The role of epigenetic modifications in the osteogenic differentiation of adipose-derived stem cells. Connect Tissue Res. 2019, 60, 507–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.C.; Su, P.F.; Huang, Y.F.; Yew, T.L.; Hung, S.C. Oct4 and Nanog directly regulate Dnmt1 to maintain self-renewal and undifferentiated state in mesenchymal stem cells. Mol. Cell 2012, 47, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Ray, R.M.; Zimmerman, B.J.; McCormack, S.A.; Patel, T.B.; Johnson, L.R. Polyamine depletion arrests cell cycle and induces inhibitors p21(Waf1/Cip1), p27(Kip1), and p53 in IEC-6 cells. Am. J. Physiol. 1999, 276, C684–C691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seiler, N. Functions of polyamine acetylation. Can. J. Physiol. Pharmacol. 1987, 65, 2024–2035. [Google Scholar] [CrossRef] [PubMed]
- Pegg, A.E. Mammalian polyamine metabolism and function. IUBMB Life 2009, 61, 880–894. [Google Scholar] [CrossRef]
- Bekebrede, A.F.; Keijer, J.; Gerrits, W.J.J.; Boer, V.C.J.d. The molecular and physiological effects of protein-derived polyamines in the intestine. Nutrients 2020, 12, 197. [Google Scholar] [CrossRef] [Green Version]
- Sandusky-Beltran, L.A.; Kovalenko, A.; Ma, C.; Calahatian, J.I.T.; Placides, D.S.; Watler, M.D.; Hunt, J.B.; Darling, A.L.; Baker, J.D.; Blair, L.J.; et al. Spermidine/spermine-N1-acetyltransferase ablation impacts tauopathy-induced polyamine stress response. Alzheimers Res. Ther. 2019, 11, 58. [Google Scholar] [CrossRef]
- Mandal, S.; Mandal, A.; Johansson, H.E.; Orjalo, A.V.; Park, M.H. Depletion of cellular polyamines, spermidine and spermine, causes a total arrest in translation and growth in mammalian cells. Proc. Natl. Acad. Sci. USA 2013, 110, 2169–2174. [Google Scholar] [CrossRef] [Green Version]
- Desiderio, M.A.; Bardella, L. Polyamine acetylations in normal and neoplastic growth processes. Amino Acids 1995, 8, 59–68. [Google Scholar] [CrossRef]
- Park, M.H.; Igarashi, K. Polyamines and their metabolites as diagnostic markers of human diseases. Biomol. Ther. 2013, 21, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, T.; Knauer, H.; Schauer, A.; Büttner, S.; Ruckenstuhl, C.; Carmona-Gutierrez, D.; Ring, J.; Schroeder, S.; Magnes, C.; Antonacci, L.; et al. Induction of autophagy by spermidine promotes longevity. Nat. Cell. Biol. 2009, 11, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; He, Y.; Lu, F. Autophagy in stem cell biology: A Perspective on stem cell self-renewal and differentiation. Stem Cells Int. 2018, 2018, 9131397. [Google Scholar] [CrossRef] [PubMed]
- Chang, N.C. Autophagy and stem cells: Self-eating for self-renewal. Front. Cell Dev. Biol. 2020, 8, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mai, S.; Muster, B.; Bereiter-Hahn, J.; Jendrach, M. Autophagy proteins LC3B, ATG5 and ATG12 participate in quality control after mitochondrial damage and influence lifespan. Autophagy 2012, 8, 47–62. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Forward | Reverse |
|---|---|---|
| OCT4 | GAGGAGTCCCAGGCAATCAA | CATCGGCCTGTGTATATCCC |
| SOX2 | CCGTTCATGTAGGTCTCGGAGCTG | CAACGGCAGCTACAGCTAGATGC |
| Nanog | CATGAGTGTGGATCCAGCT | CCTGAATAAGCAGATCCAT |
| SIRT1 | CATTTCCATGGCGCTGAGG | TGCTGGTGGAACAATTCCTGT |
| DNMT1 | CGTCCGAGCGTCACACA | GAGCCTTTGCCATTCTTCGC |
| p53 | CAAGCAATGGATGATTTGATGCT | TGGGTCTTCAGTGAACCATTGT |
| p21 (WAF1/CIP) | CAAAGGCCCGCTCTACATCTT | AGGAACCTCCATTCACCCGA |
| BAX | TGCTTCAGGGTTTCATCCAG | GGCGGCAATCATCCTCTG |
| GAPDH | GAGTCAACGGATTTGGTCGT | GACAAGCTTCCCGTTCTCAG |
| Polyamines | Ctrl | 4d-MCF-7-EM | 7d-MCF-7-EM | 10d-MCF-7-EM | p-Value |
|---|---|---|---|---|---|
| Putrescine | 0.79 | 0.68 | 0.63 | 0.60 | 0.0005 *** |
| Spermidine | 0.98 | 1.054 | 1.27 | 1.35 | 0.0009 *** |
| Spermine | 1.29 | 1.22 | 1.09 | 0.86 | 0.001 *** |
| Agmantine | 0.8 | 0.65 | 0.60 | 0.61 | 0.0007 *** |
| N-acetylputrescine | 0.38 | 0.38 | 0.42 | 0.42 | 0.00003 *** |
| N-acetylspermine | 0.411 | 0.420 | 0.510 | 0.515 | 0.0005 *** |
| N-acetylspermidine | 0.322 | 0.284 | 0.268 | 0.270 | 0.0002 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garroni, G.; Balzano, F.; Cruciani, S.; Pala, R.; Coradduzza, D.; Azara, E.; Bellu, E.; Cossu, M.L.; Ginesu, G.C.; Carru, C.; et al. Adipose-Derived Stem Cell Features and MCF-7. Cells 2021, 10, 1754. https://doi.org/10.3390/cells10071754
Garroni G, Balzano F, Cruciani S, Pala R, Coradduzza D, Azara E, Bellu E, Cossu ML, Ginesu GC, Carru C, et al. Adipose-Derived Stem Cell Features and MCF-7. Cells. 2021; 10(7):1754. https://doi.org/10.3390/cells10071754
Chicago/Turabian StyleGarroni, Giuseppe, Francesca Balzano, Sara Cruciani, Renzo Pala, Donatella Coradduzza, Emanuela Azara, Emanuela Bellu, Maria Laura Cossu, Giorgio C. Ginesu, Ciriaco Carru, and et al. 2021. "Adipose-Derived Stem Cell Features and MCF-7" Cells 10, no. 7: 1754. https://doi.org/10.3390/cells10071754
APA StyleGarroni, G., Balzano, F., Cruciani, S., Pala, R., Coradduzza, D., Azara, E., Bellu, E., Cossu, M. L., Ginesu, G. C., Carru, C., Ventura, C., & Maioli, M. (2021). Adipose-Derived Stem Cell Features and MCF-7. Cells, 10(7), 1754. https://doi.org/10.3390/cells10071754