
Unveiling the Roles of Low-Density Lipoprotein Receptor-Related Protein 6 in Intestinal Homeostasis, Regeneration and Oncogenesis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies and Reagents
2.2. Cell Culture
2.3. Mice
2.4. Colitis Induction with DSS and Clinical Evaluation
2.5. Macroadenoma Count, Histological Staining, Immunofluorescence and Immunohistochemistry
2.6. Enteroids
2.7. Western Blot Analysis
2.8. RNA Extraction and qRT–PCR
2.9. LRP6 Silencing
2.10. Soft Agar Assay
2.11. Clonogenic Assay
2.12. Statistical Analyses
3. Results
3.1. Lrp6 Deletion Does Not Alter Intestinal Architecture and Homeostasis but Impairs Wnt Target Gene Expression
3.2. LRP6 Expression Is Required for Ex Vivo Intestinal Crypt Regeneration
3.3. Lrp6IEC-KO Mice Are More Sensitive to DSS-Induced Epithelial Damage and Colitis
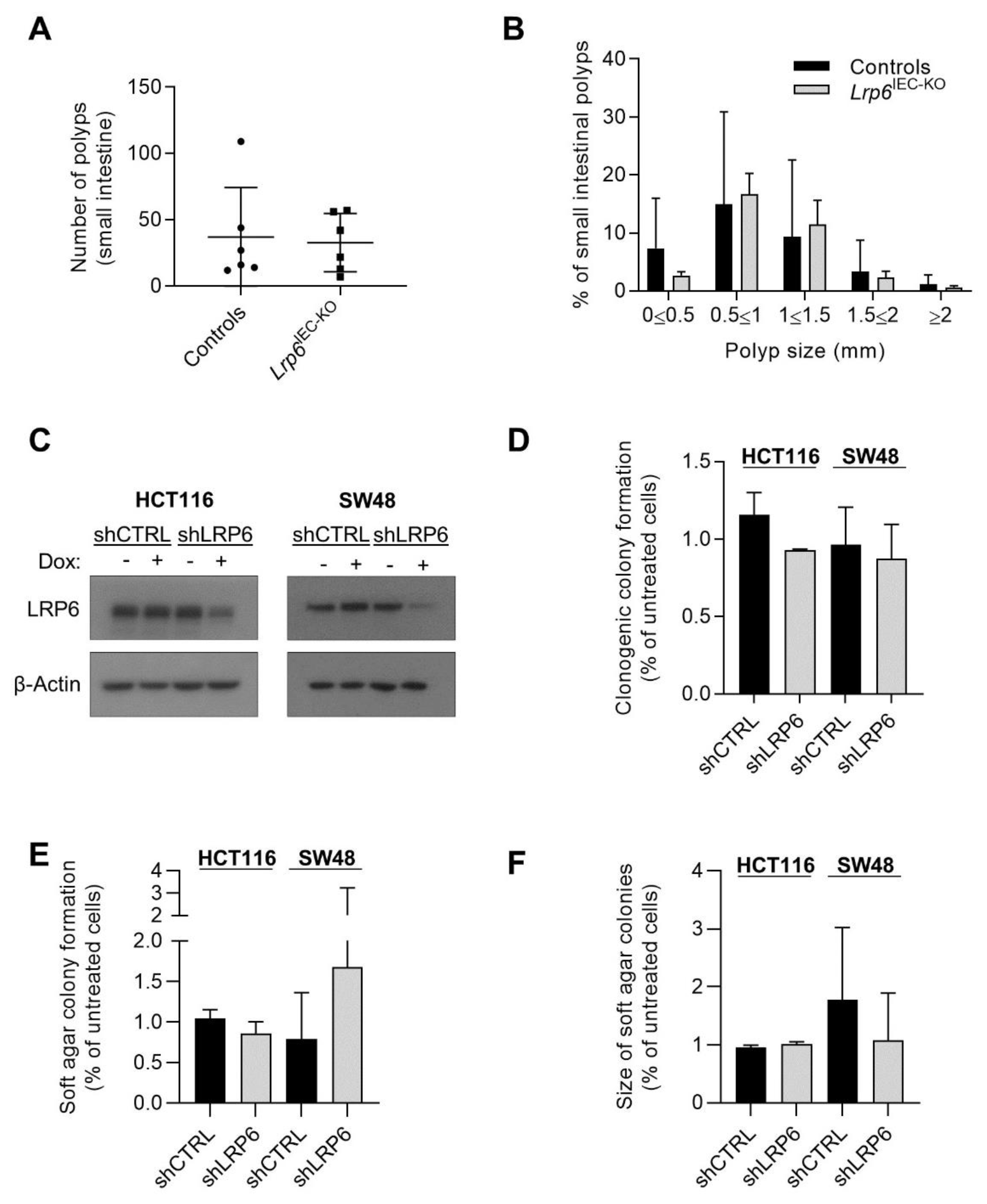
3.4. LRP6 Is Dispensable for Adenoma Formation in ApcMin/+ Mice and for Growth of Human Colorectal Cancer Cells
3.5. ERK-Dependent Phosphorylation of LRP6 in KRAS- and BRAF-Mutated Colorectal Cancer Cells—A Role for LRP6 in BRAF-Induced Intestinal Oncogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Der Flier, L.G.; Clevers, H. Stem Cells, Self-Renewal, and Differentiation in the Intestinal Epithelium. Annu. Rev. Physiol. 2009, 71, 241–260. [Google Scholar] [CrossRef]
- Takahashi, T.; Shiraishi, A. Stem Cell Signaling Pathways in the Small Intestine. Int. J. Mol. Sci. 2020, 21, 2032. [Google Scholar] [CrossRef] [Green Version]
- Pinson, K.I.; Brennan, J.; Monkley, S.; Avery, B.J.; Skarnes, W.C. An LDL-receptor-related protein mediates Wnt signalling in mice. Nat. Cell Biol. 2000, 407, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.J.; Yamagami, T.; Zhao, T.; Song, L.; Wang, K.; Wang, Y.-Z. Generation of Lrp6 conditional gene-targeting mouse line for modeling and dissecting multiple birth defects/congenital anomalies. Dev. Dyn. 2009, 239, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Baker, J.J.; Zylstra-Diegel, C.R.; Williams, B.O. Lrp5 and Lrp6 play compensatory roles in mouse intestinal development. J. Cell. Biochem. 2012, 113, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Fearon, E.R.; Vogelstein, B. A genetic model for colorectal tumorigenesis. Cell 1990, 61, 759–767. [Google Scholar] [CrossRef]
- Lemieux, E.; Cagnol, S.; Beaudry, K.; Carrier, J.; Rivard, N. Oncogenic KRAS signalling promotes the Wnt/β-catenin pathway through LRP6 in colorectal cancer. Oncogene 2014, 34, 4914–4927. [Google Scholar] [CrossRef]
- Rismani, E.; Fazeli, M.S.; Mahmoodzadeh, H.; Movassagh, A.; Azami, S.; Karimipoor, M.; Teimoori-Toolabi, L. Pattern of LRP6 gene expression in tumoral tissues of colorectal cancer. Cancer Biomark. 2017, 19, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; An, Y.; Hou, W.; Cao, Y.-N.; Yao, M.-F.; Ma, N.-N.; Hou, L.; Zhang, H.; Liu, H.-J.; Zhang, B. LRP6 promotes invasion and metastasis of colorectal cancer through cytoskeleton dynamics. Oncotarget 2017, 8, 109632–109645. [Google Scholar] [CrossRef] [Green Version]
- Perlman, R.L. Mouse Models of Human Disease: An Evolutionary Perspective. Evol. Med. Public Health 2016, 2016, 170–176. [Google Scholar] [CrossRef] [Green Version]
- Chinwalla, A.T.; Cook, L.L.; Delehaunty, K.D.; Fewell, G.A.; Fulton, L.A.; Fulton, R.S.; Graves, T.A.; Hillier, L.W.; Mardis, E.R.; McPherson, J.D.; et al. Initial sequencing and comparative analysis of the mouse genome. Nature 2002, 420, 520–562. [Google Scholar] [CrossRef]
- Bergeron, S.; Lemieux, E.; Durand, V.; Cagnol, S.; Carrier, J.C.; Lussier, J.G.; Boucher, M.-J.; Rivard, N. The serine protease inhibitor serpinE2 is a novel target of ERK signaling involved in human colorectal tumorigenesis. Mol. Cancer 2010, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Investig. 1993, 69, 238–249. [Google Scholar] [PubMed]
- Leblanc, C.; Langlois, M.; Coulombe, G.; Vaillancourt-Lavigueur, V.; Jones, C.; Carrier, J.C.; Boudreau, F.; Rivard, N. Epithelial Src homology region 2 domain–containing phosphatase-1 restrains intestinal growth, secretory cell differentiation, and tumorigenesis. FASEB J. 2017, 31, 3512–3526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonneaud, A.; Jones, C.; Turgeon, N.; Lévesque, D.; Asselin, C.; Boudreau, F.; Boisvert, F.-M. A SILAC-Based Method for Quantitative Proteomic Analysis of Intestinal Organoids. Sci. Rep. 2016, 6, 38195 . [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiederschain, D.; Susan, W.; Chen, L.; Loo, A.; Yang, G.; Huang, A.; Chen, Y.; Caponigro, G.; Yao, Y.-M.; Lengauer, C.; et al. Single-vector inducible lentiviral RNAi system for oncology target validation. Cell Cycle 2009, 8, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Babeu, J.-P.; Jones, C.; Geha, S.; Carrier, J.C.; Boudreau, F. P1 promoter-driven HNF4α isoforms are specifically repressed by β-catenin signaling in colorectal cancer cells. J. Cell Sci. 2018, 131, jcs.214734. [Google Scholar] [CrossRef] [Green Version]
- Bian, B.; Mongrain, S.; Cagnol, S.; Langlois, M.-J.; Boulanger, J.; Bernatchez, G.; Carrier, J.C.; Boudreau, F.; Rivard, N. Cathepsin B promotes colorectal tumorigenesis, cell invasion, and metastasis. Mol. Carcinog. 2015, 55, 671–687. [Google Scholar] [CrossRef]
- Madison, B.B.; Dunbar, L.; Qiao, X.T.; Braunstein, K.; Braunstein, E.; Gumucio, D.L. cis Elements of the Villin Gene Control Expression in Restricted Domains of the Vertical (Crypt) and Horizontal (Duodenum, Cecum) Axes of the Intestine. J. Biol. Chem. 2002, 277, 33275–33283. [Google Scholar] [CrossRef] [Green Version]
- Moser, A.R.; Pitot, H.C.; Dove, W.F. A dominant mutation that predisposes to multiple intestinal neoplasia in the mouse. Science 1990, 247, 322–324. [Google Scholar] [CrossRef]
- Fevr, T.; Robine, S.; Louvard, D.; Huelsken, J. Wnt/β-Catenin Is Essential for Intestinal Homeostasis and Maintenance of Intestinal Stem Cells. Mol. Cell. Biol. 2007, 27, 7551–7559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, K.S.; Kuo, C.J. Ascl2 reinforces intestinal stem cell identity. Cell Stem Cell 2015, 16, 105–106. [Google Scholar] [CrossRef] [Green Version]
- Brodrick, B.; Vidrich, A.; Porter, E.; Bradley, L.; Buzan, J.M.; Cohn, S.M. Fibroblast Growth Factor Receptor-3 (FGFR-3) Regulates Expression of Paneth Cell Lineage-specific Genes in Intestinal Epithelial Cells through both TCF4/β-Catenin-dependent and -independent Signaling Pathways. J. Biol. Chem. 2011, 286, 18515–18525. [Google Scholar] [CrossRef] [Green Version]
- Dubé, P.E.; Rowland, K.J.; Brubaker, P.L. Glucagon-Like Peptide-2 Activates β-Catenin Signaling in the Mouse Intestinal Crypt: Role of Insulin-Like Growth Factor-I. Endocrinology 2008, 149, 291–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-L.; Yang, J.; James, I.O.; Zhang, H.-Y.; Besner, G.E. Heparin-binding epidermal growth factor-like growth factor restores Wnt/β-catenin signaling in intestinal stem cells exposed to ischemia/reperfusion injury. Surgery 2014, 155, 1069–1080. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.X.; Zhu, F.; Li, J.Z.; Li, Y.L.; Ding, X.Q.; Yin, J.; Xiong, X.; Yang, H.S. Epidermal growth factor promotes intestinal secretory cell differentiation in weaning piglets via Wnt/β-catenin signalling. Animal 2020, 14, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Durand, A.; Donahue, B.; Peignon, G.; Letourneur, F.; Cagnard, N.; Slomianny, C.; Perret, C.; Shroyer, N.; Romagnolo, B. Functional intestinal stem cells after Paneth cell ablation induced by the loss of transcription factor Math1 (Atoh1). Proc. Natl. Acad. Sci. USA 2012, 109, 8965–8970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farin, H.F.; Van Es, J.H.; Clevers, H. Redundant Sources of Wnt Regulate Intestinal Stem Cells and Promote Formation of Paneth Cells. Gastroenterology 2012, 143, 1518–1529.e7. [Google Scholar] [CrossRef]
- Aoki, R.; Carmel, M.S.; Gao, N.; Shin, S.; May, C.L.; Golson, M.L.; Zahm, A.M.; Ray, M.; Wiser, C.L.; Wright, C.V.; et al. Foxl1-Expressing Mesenchymal Cells Constitute the Intestinal Stem Cell Niche. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 175–188. [Google Scholar] [CrossRef] [Green Version]
- Degirmenci, B.; Valenta, T.; Dimitrieva, S.; Hausmann, G.; Basler, K. GLI1-expressing mesenchymal cells form the essential Wnt-secreting niche for colon stem cells. Nature 2018, 558, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Greicius, G.; Kabiri, Z.; Sigmundsson, K.; Liang, C.; Bunte, R.; Singh, M.; Virshup, D.M. PDGFRα+ pericryptal stromal cells are the critical source of Wnts and RSPO3 for murine intestinal stem cells in vivo. Proc. Natl. Acad. Sci. USA 2018, 115, E3173–E3181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabiri, Z.; Greicius, G.; Madan, B.; Biechele, S.; Zhong, Z.; Zaribafzadeh, H.; Edison; Aliyev, J.; Wu, Y.; Bunte, R.; et al. Stroma provides an intestinal stem cell niche in the absence of epithelial Wnts. Development 2014, 141, 2206–2215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoshkes-Carmel, M.; Wang, Y.J.; Wangensteen, K.J.; Tóth, B.; Kondo, A.; Massassa, E.E.; Itzkovitz, S.; Kaestner, K.H. Subepithelial telocytes are an important source of Wnts that supports intestinal crypts. Nature 2018, 557, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Holmen, S.L.; Salic, A.; Zylstra, C.R.; Kirschner, M.W.; Williams, B. A Novel Set of Wnt-Frizzled Fusion Proteins Identifies Receptor Components that Activate β-Catenin-dependent Signaling. J. Biol. Chem. 2002, 277, 34727–34735. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Semenov, M.; Tamai, K.; Zeng, X. LDL receptor-related proteins 5 and 6 in Wnt/β-catenin signaling:Arrows point the way. Development 2004, 131, 1663–1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koslowski, M.J.; Teltschik, Z.; Beißner, J.; Schaeffeler, E.; Wang, G.; Kübler, I.; Gersemann, M.; Cooney, R.; Jewell, D.; Reinisch, W.; et al. Association of a Functional Variant in the Wnt Co-Receptor LRP6 with Early Onset Ileal Crohn’s Disease. PLoS Genet. 2012, 8, e1002523. [Google Scholar] [CrossRef] [Green Version]
- Nava, P.; Koch, S.; Laukoetter, M.G.; Lee, W.Y.; Kolegraff, K.; Capaldo, C.T.; Beeman, N.; Addis, C.; Gerner-Smidt, K.; Neumaier, I.; et al. Interferon-γ Regulates Intestinal Epithelial Homeostasis through Converging β-Catenin Signaling Pathways. Immunity 2010, 32, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Eastwood, G.L. Gastrointestinal Epithelial Renewal. Gastroenterology 1977, 72, 962–975. [Google Scholar] [CrossRef]
- Chen, M.; He, X. APC Deficiency Leads to β-Catenin Stabilization and Signaling Independent of LRP5/6. Dev. Cell 2019, 49, 825–826. [Google Scholar] [CrossRef]
- Saito-Diaz, K.; Benchabane, H.; Tiwari, A.; Tian, A.; Li, B.; Thompson, J.J.; Hyde, A.S.; Sawyer, L.M.; Jodoin, J.N.; Santos, E.; et al. APC Inhibits Ligand-Independent Wnt Signaling by the Clathrin Endocytic Pathway. Dev. Cell 2018, 44, 566–581.e8. [Google Scholar] [CrossRef] [Green Version]
- Cabel, C.R.; Alizadeh, E.; Robbins, D.J.; Ahmed, Y.; Lee, E.; Thorne, C.A. Single-Cell Analyses Confirm the Critical Role of LRP6 for Wnt Signaling in APC-Deficient Cells. Dev. Cell 2019, 49, 827–828. [Google Scholar] [CrossRef] [PubMed]
- East, J.E.; Atkin, W.S.; Bateman, A.C.; Clark, S.K.; Dolwani, S.; Ket, S.N.; Leedham, S.J.; Phull, P.S.; Rutter, M.D.; Shepherd, N.A.; et al. British Society of Gastroenterology position statement on serrated polyps in the colon and rectum. Gut 2017, 66, 1181–1196. [Google Scholar] [CrossRef] [Green Version]
- Cagnol, S.; Rivard, N. Oncogenic KRAS and BRAF activation of the MEK/ERK signaling pathway promotes expression of dual-specificity phosphatase 4 (DUSP4/MKP2) resulting in nuclear ERK1/2 inhibition. Oncogene 2012, 32, 564–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachs, N.; Clevers, H. Organoid cultures for the analysis of cancer phenotypes. Curr. Opin. Genet. Dev. 2014, 24, 68–73. [Google Scholar] [CrossRef]
- Van De Wetering, M.; Francies, H.E.; Francis, J.M.; Bounova, G.; Iorio, F.; Pronk, A.; Van Houdt, W.; Van Gorp, J.; Taylor-Weiner, A.; Kester, L.; et al. Prospective derivation of a living organoid biobank of colorectal cancer patients. Cell 2015, 161, 933–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raisch, J.; Côté-Biron, A.; Langlois, M.-J.; Leblanc, C.; Rivard, N. Unveiling the Roles of Low-Density Lipoprotein Receptor-Related Protein 6 in Intestinal Homeostasis, Regeneration and Oncogenesis. Cells 2021, 10, 1792. https://doi.org/10.3390/cells10071792
Raisch J, Côté-Biron A, Langlois M-J, Leblanc C, Rivard N. Unveiling the Roles of Low-Density Lipoprotein Receptor-Related Protein 6 in Intestinal Homeostasis, Regeneration and Oncogenesis. Cells. 2021; 10(7):1792. https://doi.org/10.3390/cells10071792
Chicago/Turabian StyleRaisch, Jennifer, Anthony Côté-Biron, Marie-Josée Langlois, Caroline Leblanc, and Nathalie Rivard. 2021. "Unveiling the Roles of Low-Density Lipoprotein Receptor-Related Protein 6 in Intestinal Homeostasis, Regeneration and Oncogenesis" Cells 10, no. 7: 1792. https://doi.org/10.3390/cells10071792
APA StyleRaisch, J., Côté-Biron, A., Langlois, M. -J., Leblanc, C., & Rivard, N. (2021). Unveiling the Roles of Low-Density Lipoprotein Receptor-Related Protein 6 in Intestinal Homeostasis, Regeneration and Oncogenesis. Cells, 10(7), 1792. https://doi.org/10.3390/cells10071792