
Fisher vs. the Worms: Extraordinary Sex Ratios in Nematodes and the Mechanisms that Produce Them


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Nematoda as a Model Phylum for the Evolution of Sex Ratios
2.1. Fisherian Nematodes
2.2. Non-Fisherian Nematodes
3. Mechanisms Producing Non-Fisherian Sex Ratios
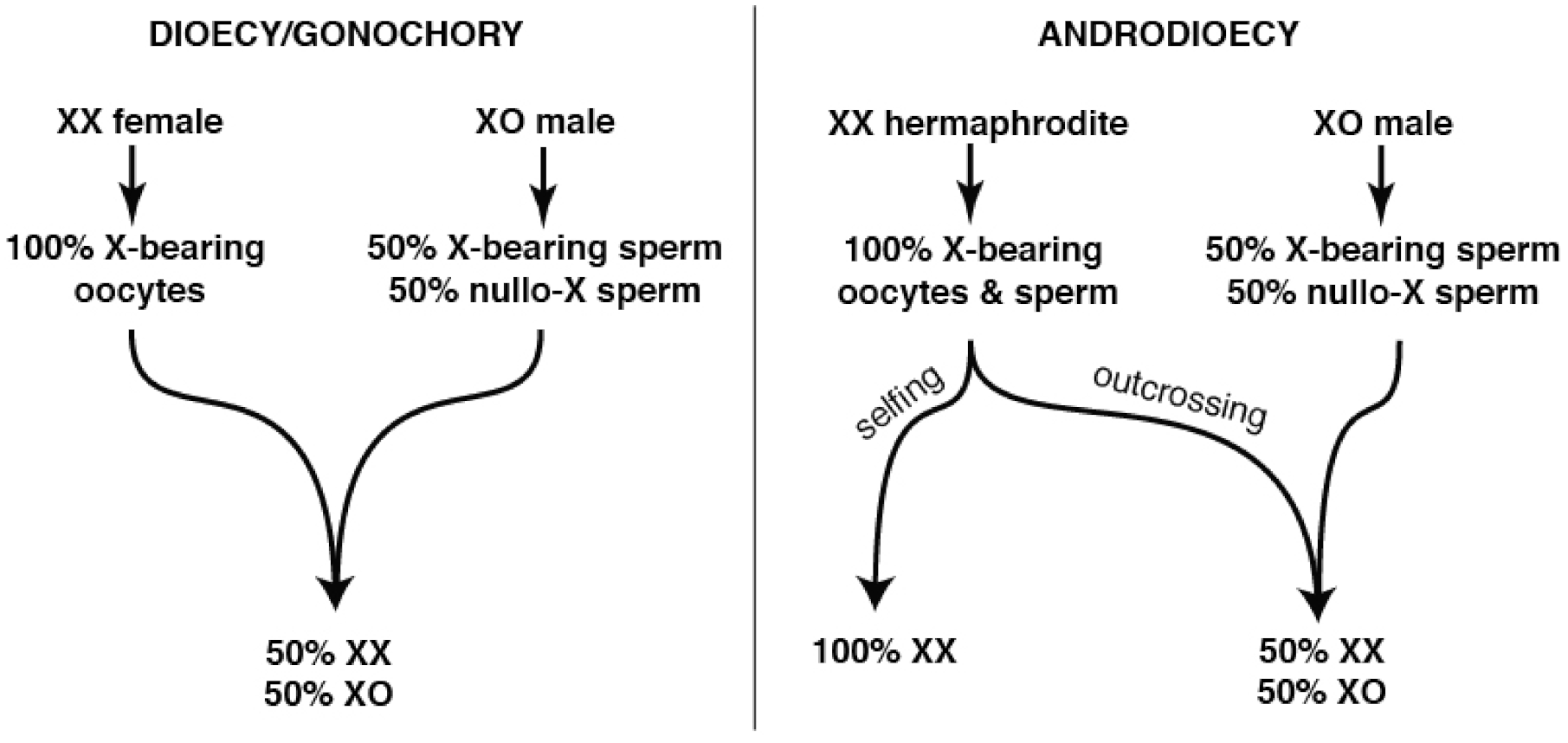
3.1. Path 1: Maintenance of GSD, but with Self-Sperm (Androdioecy)
3.2. Path 2: Maintain GSD, but Enrich for Females with Facultative Parthenogenesis
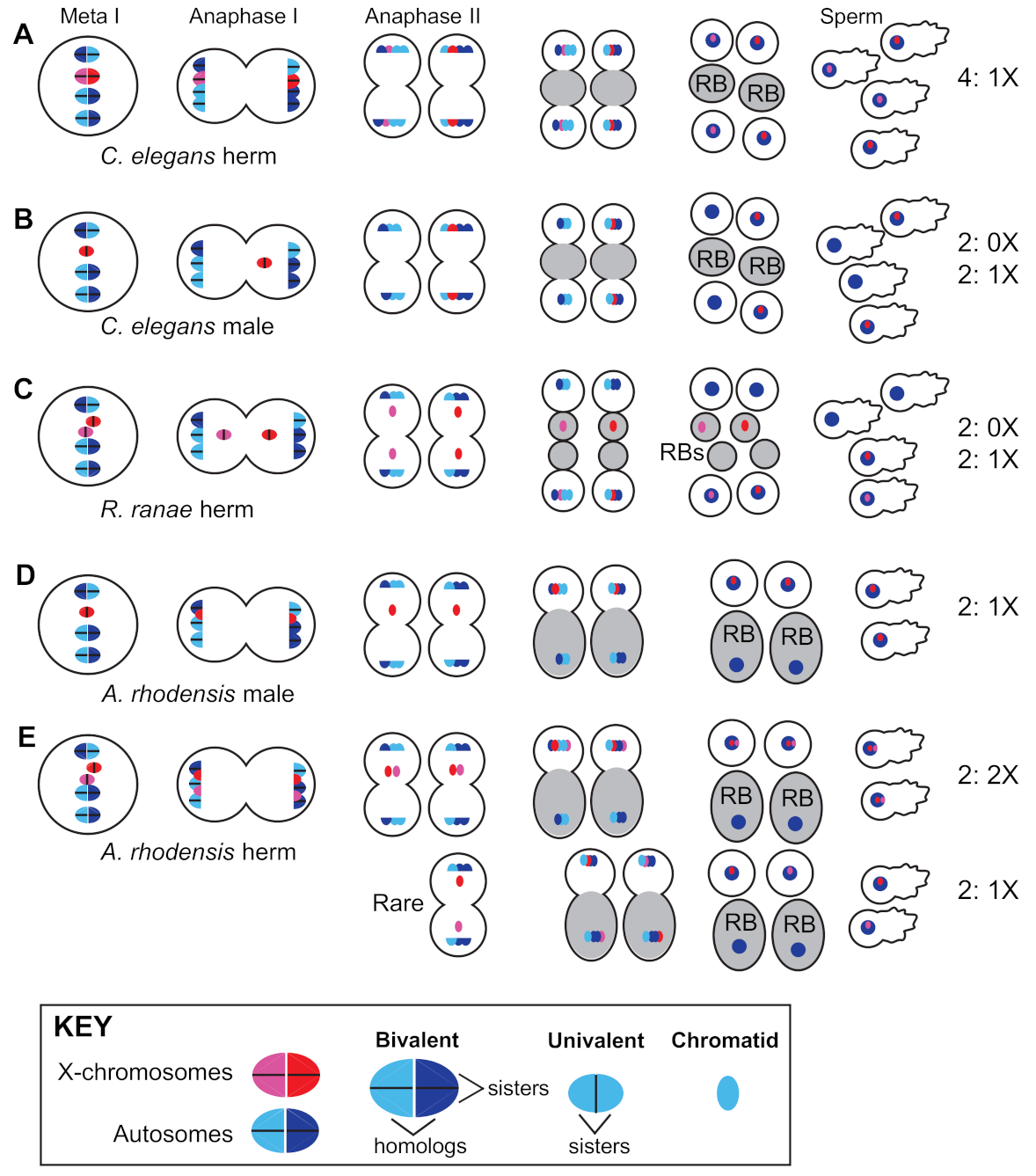
3.3. Path 3: Combine GSD with Non-Mendelian Sex Chromosome Segregation
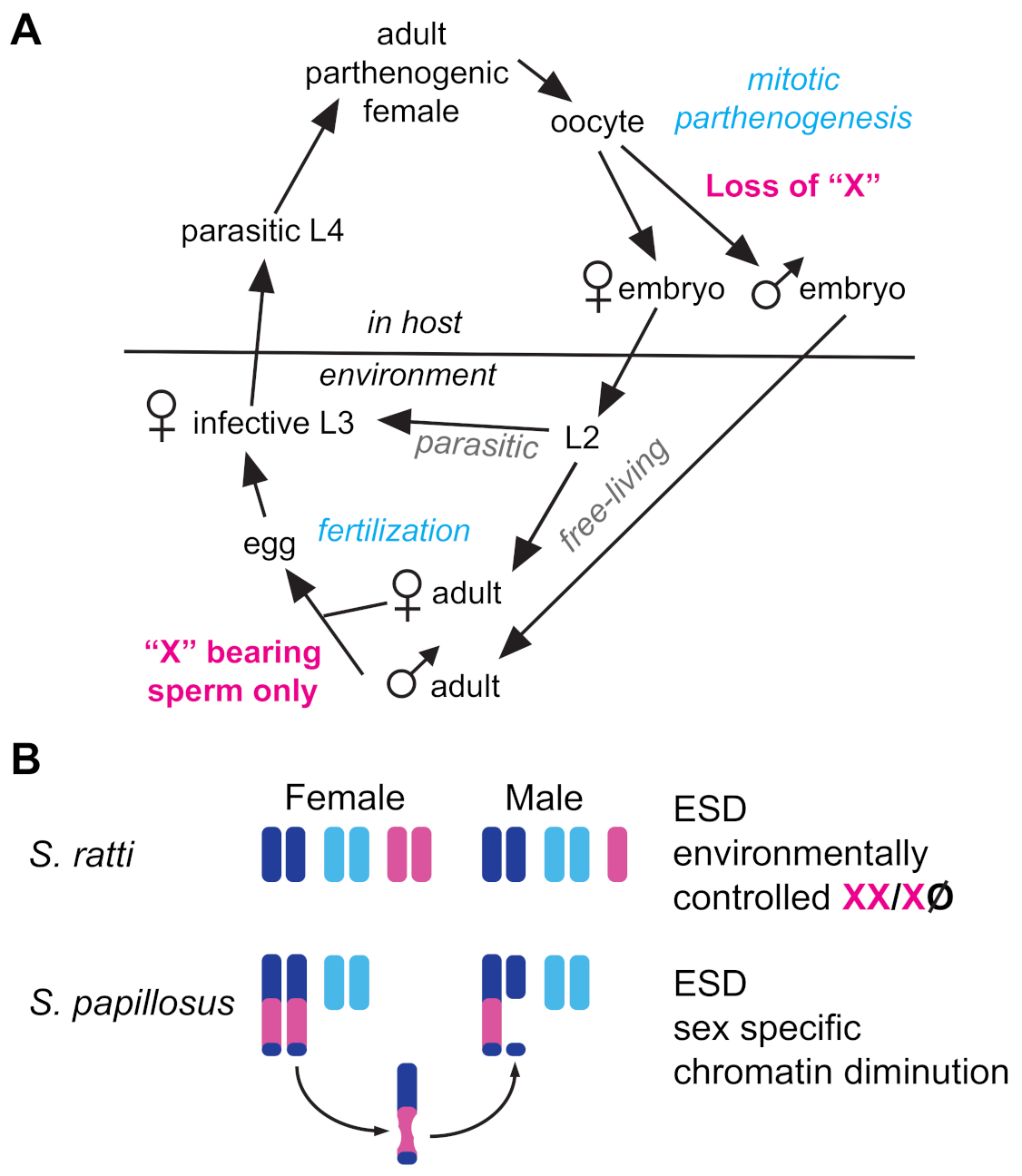
3.4. Path 4: Abandon GSD and Use Environmental Sex Determination
3.5. Intersectionality—Auanema and Beyond
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobzhansky, T. Nothing in Biology Makes Sense Except in the Light of Evolution. Am. Biol. Teach. 1973, 35, 125–129. [Google Scholar] [CrossRef]
- Parker, G.A.; Baker, R.R.; Smith, V.G.F. The origin and evolution of gamete dimorphism and the male-female phenomenon. J. Theor. Biol. 1972, 36, 529–553. [Google Scholar] [CrossRef]
- Kodric-Brown, A.; Brown, J.H. Anisogamy, sexual selection, and the evolution and maintenance of sex. Evol. Ecol. 1987, 1, 95–105. [Google Scholar] [CrossRef]
- Schärer, L.; Rowe, L.; Arnqvist, G. Anisogamy, chance and the evolution of sex roles. Trends Ecol. Evol. 2012, 27, 260–264. [Google Scholar] [CrossRef]
- Lehtonen, J.; Parker, G.A.; Schärer, L. Why anisogamy drives ancestral sex roles. Evolution 2016, 70, 1129–1135. [Google Scholar] [CrossRef]
- Darwin, C. The Descent of Man and Selection in Relation to Sex, 1st ed.; John Murray: London, UK, 1871. [Google Scholar]
- Edwards, A.W.F. Carl Düsing (1884) on The Regulation of the Sex-Ratio. Theor. Popul. Biol. 2000, 58, 255–257. [Google Scholar] [CrossRef]
- Fisher, R.A. The Genetical Theory of Natural Selection; Oxford University Press: Oxford, UK, 1930. [Google Scholar]
- Trivers, R.L.; Willard, D.E. Natural selection of parental ability to vary the sex ratio of offspring. Science 1973, 179, 90–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orzack, S.H.; Stubblefield, J.W.; Akmaev, V.R.; Colls, P.; Munné, S.; Scholl, T.; Steinsaltz, D.; Zuckerman, J.E. The human sex ratio from conception to birth. Proc. Natl. Acad. Sci. USA 2015, 112, E2102–E2111. [Google Scholar] [CrossRef] [Green Version]
- Wright, S. Evolution in Mendelian Populations. Genetics 1931, 16, 97–159. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.A. Sperm competition and its evolutionary consequences in the insects. Biol. Rev. 1970, 45, 525–567. [Google Scholar] [CrossRef]
- Wedell, N.; Gage, M.J.G.; Parker, G.A. Sperm competition, male prudence and sperm-limited females. Trends Ecol. Evol. 2002, 17, 313–320. [Google Scholar] [CrossRef]
- Parker, G.A.; Pizzari, T. Sperm competition and ejaculate economics. Biol. Rev. 2010. [Google Scholar] [CrossRef] [PubMed]
- Haldane, J.B.S. Sex ratio and unisexual sterility in hybrid animals. J. Genet. 1922, 12, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.M. The Evolution of Sex; Cambridge University Press: Cambridge, UK, 1978. [Google Scholar]
- Darwin, C. The Descent of Man and Selection in Relation to Sex, 2nd ed.; John Murray: London, UK, 1882. [Google Scholar]
- Hamilton, W.D. Extraordinary Sex Ratios. Science 1967, 156, 477–488. [Google Scholar] [CrossRef]
- Bull, J. Evolution of Sex Determining Mechanisms; Benjamin Cummings Publishing Company: San Francisco, CA, USA, 1983. [Google Scholar]
- Charnov, E. The Theory of Sex Allocation; Princeton University Press: Princeton, NJ, USA, 1982. [Google Scholar]
- Taylor, P.D.; Bulmer, M.G. Local mate competition and the sex ratio. J. Theor. Biol. 1980, 86, 409–419. [Google Scholar] [CrossRef]
- West, S. Sex Allocation; Princeton University Press: Princeton, NJ, USA, 2009. [Google Scholar]
- Colwell, R.K. Group selection is implicated in the evolution of female-biased sex ratios. Nature 1981, 290, 401–404. [Google Scholar] [CrossRef]
- Wilson, D.S.; Colwell, R.K. Evolution of Sex Ratio in Structured Demes. Evolution 1981, 35, 882–897. [Google Scholar] [CrossRef]
- Lynch, M. The genetic interpretation of inbreeding depression and outbreeding depression. Evolution 1991, 45, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Edmands, S.; Timmerman, C.C. Modeling Factors Affecting the Severity of Outbreeding Depression. Conserv. Biol. 2003, 17, 883–892. [Google Scholar] [CrossRef] [Green Version]
- Green, R.F.; Gordh, G.; Hawkins, B.A. Precise Sex Ratios in Highly Inbred Parasitic Wasps. Am. Nat. 1982, 120, 653–665. [Google Scholar] [CrossRef]
- Hardy, I.C.W.; Cook, J.M. Brood sex ratio variance, developmental mortality and virginity in a gregarious parasitoid wasp. Oecologia 1995, 103, 162–169. [Google Scholar] [CrossRef]
- Verhulst, E.C.; Beukeboom, L.W.; Van De Zande, L. Maternal Control of Haplodiploid Sex Determination in the Wasp Nasonia. Science 2010, 328, 620–623. [Google Scholar] [CrossRef] [Green Version]
- Macke, E.; Magalhães, S.; Bach, F.; Olivieri, I. Experimental evolution of reduced sex ratio adjustment under local mate competition. Science 2011, 334, 1127–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herre, E.A. Sex ratio adjustment in fig wasps. Science 1985, 228, 896–898. [Google Scholar] [CrossRef] [PubMed]
- Herre, E.A. Optimality, plasticity and selective regime in fig wasp sex ratios. Nature 1987, 329, 627–629. [Google Scholar] [CrossRef]
- West, S.A.; Herre, E.A. Stabilizing selection and variance in fig wasp sex ratios. Evolution 1998, 52, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.A. Hierarchical selection theory and sex ratios. II. on applying the theory, and a test with fig wasps. Evolution 1985, 39, 949–964. [Google Scholar] [CrossRef]
- Ghiselin, M.T. The Evolution of Hermaphroditism Among Animals. Q. Rev. Biol. 1969, 44, 189–208. [Google Scholar] [CrossRef]
- Heath, D.J. Brooding and the evolution of hermaphroditism. J. Theor. Biol. 1979, 81, 151–155. [Google Scholar] [CrossRef]
- Charnov, E.L. Simultaneous hermaphroditism and sexual selection. Proc. Natl. Acad. Sci. USA 1979, 76, 2480–2484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Kooi, C.J.; Matthey-Doret, C.; Schwander, T. Evolution and comparative ecology of parthenogenesis in haplodiploid arthropods. Evol. Lett. 2017, 1, 304–316. [Google Scholar] [CrossRef] [Green Version]
- Glémin, S.; Galtier, N. Genome Evolution in Outcrossing Versus Selfing Versus Asexual Species; Humana Press: Totowa, NJ, USA, 2012; pp. 311–335. [Google Scholar]
- Barrett, S.C. Evolution of Mating Systems: Outcrossing Versus Selfing; Princeton University Press: Princeton, NJ, USA, 2014; pp. 356–362. [Google Scholar]
- Thomas, C.G.; Woodruff, G.C.; Haag, E.S. Causes and consequences of the evolution of reproductive mode in Caenorhabditis nematodes. Trends Genet. 2012, 28, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Baker, H.G. Self-Compatibility and establishment after ‘long-distance’ dispersal. Evolution 1955, 9, 347–349. [Google Scholar] [CrossRef]
- Lively, C.M.; Johnson, S.G. Brooding and the evolution of parthenogenesis: Strategy models and evidence from aquatic invertebrates. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1994, 256, 89–95. [Google Scholar] [CrossRef]
- Morran, L.T.; Parmenter, M.D.; Phillips, P.C. Mutation load and rapid adaptation favour outcrossing over self-fertilization. Nature 2009, 462, 350–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haigh, J. The accumulation of deleterious genes in a population—Muller’s Ratchet. Theor. Popul. Biol. 1978, 14, 251–267. [Google Scholar] [CrossRef]
- Nordborg, M. Structured coalescent processes on different time scales. Genetics 1997, 146, 1501–1514. [Google Scholar] [CrossRef] [PubMed]
- Kamran-Disfani, A.; Agrawal, A.F. Selfing, adaptation and background selection in finite populations. J. Evol. Biol. 2014, 27, 1360–1371. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, E.E.; Kohn, J.R.; Lande, R.; Robertson, K.A.; Smith, S.A.; Igić, B. Species Selection Maintains Self-Incompatibility. Science 2010, 330, 493–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, M.; Gabriel, W. Phenotypic Evolution and Parthenogenesis. Am. Nat. 1983, 122, 745–764. [Google Scholar] [CrossRef] [Green Version]
- Charnov, E.L.; Bull, J.J.; Maynard Smith, J. Why be an hermaphrodite? Nature 1976, 263, 125–126. [Google Scholar] [CrossRef]
- Handel, M.A.; Schimenti, J.C. Genetics of mammalian meiosis: Regulation, dynamics and impact on fertility. Nat. Rev. Genet. 2010, 11, 124–136. [Google Scholar] [CrossRef]
- O’Neill, M.J.; O’Neill, R.J. Sex chromosome repeats tip the balance towards speciation. Mol. Ecol. 2018, 27, 3783–3798. [Google Scholar] [CrossRef]
- Murphy, R.W.; Fu, J.; Macculloch, R.D.; Darevsky, I.S.; Kupriyanova, L.A. A fine line between sex and unisexuality: The phylogenetic constraints on parthenogenesis in lacertid lizards. Zool. J. Linn. Soc. 2000, 130, 527–549. [Google Scholar] [CrossRef]
- Hillis, D.M.; Green, D.M. Evolutionary changes of heterogametic sex in the phylogenetic history of amphibians. J. Evol. Biol. 1990, 3, 49–64. [Google Scholar] [CrossRef]
- Eckelbarger, K.J.; Watling, L. Role of Phylogenetic Constraints in Determining Reproductive Patterns in Deep-Sea Invertebrates. Invertebr. Biol. 1995, 114, 256. [Google Scholar] [CrossRef]
- Charnov, E.L.; Bull, J. When is sex environmentally determined? Nature 1977, 266, 828–830. [Google Scholar] [CrossRef]
- Bull, J.J. Evolution of environmental sex determination from genotypic sex determination. Heredity 1981, 47, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Janzen, F.J.; Paukstis, G.L. Environmental Sex Determination in Reptiles: Ecology, Evolution, and Experimental Design. Q. Rev. Biol. 1991, 66, 149–179. [Google Scholar] [CrossRef]
- Rovatsos, M.; Vukić, J.; Mrugała, A.; Suwala, G.; Lymberakis, P.; Kratochvíl, L. Little evidence for switches to environmental sex determination and turnover of sex chromosomes in lacertid lizards. Sci. Rep. 2019, 9, 7832. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, S.; Literman, R.; Neuwald, J.L.; Valenzuela, N. Thermal Response of Epigenetic Genes Informs Turtle Sex Determination with and without Sex Chromosomes. Sex. Dev. 2018, 12, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Ciofi, C.; Swingland, I.R. Environmental sex determination in reptiles. Appl. Anim. Behav. Sci. 1997, 51, 251–265. [Google Scholar] [CrossRef]
- Janzen, F.J.; Phillips, P.C. Exploring the evolution of environmental sex determination, especially in reptiles. J. Evol. Biol. 2006, 19, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
- Conover, D.O.; Van Voorhees, D.A.; Ehtisham, A. Sex ratio selection and the evolution of environmental sex determination in laboratory populations of menidia menidia. Evolution 1992, 46, 1722–1730. [Google Scholar] [CrossRef]
- Pannell, J. Mixed genetic and environmental sex determination in an androdioecious population of Mercurialis annua. Heredity 1997, 78, 50–56. [Google Scholar] [CrossRef]
- Kato, Y.; Kobayashi, K.; Watanabe, H.; Iguchi, T. Environmental Sex Determination in the Branchiopod Crustacean Daphnia magna: Deep Conservation of a Doublesex Gene in the Sex-Determining Pathway. PLoS Genet. 2011, 7, e1001345. [Google Scholar] [CrossRef] [Green Version]
- Cotton, S.; Wedekind, C. Population Consequences of Environmental Sex Reversal. Conserv. Biol. 2009, 23, 196–206. [Google Scholar] [CrossRef]
- Blumenthal, T.; Davis, R.E. Exploring nematode diversity. Nat. Genet. 2004, 36, 1246–1247. [Google Scholar] [CrossRef]
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Ettema, C.H. Soil nematode diversity: Species coexistence and ecosystem function. J. Nematol. 1998, 30, 159–169. [Google Scholar]
- Porazinska, D.L.; Giblin-Davis, R.M.; Faller, L.; Farmerie, W.; Kanzaki, N.; Morris, K.; Powers, T.O.; Tucker, A.E.; Sung, W.; Thomas, W.K. Evaluating high-throughput sequencing as a method for metagenomic analysis of nematode diversity. Mol. Ecol. Resour. 2009, 9, 1439–1450. [Google Scholar] [CrossRef]
- Vanreusel, A.; Fonseca, G.; Danovaro, R.; Da Silva, M.C.; Esteves, A.M.; Ferrero, T.; Gad, G.; Galtsova, V.; Gambi, C.; Da Fonsêca Genevois, V.; et al. The contribution of deep-sea macrohabitat heterogeneity to global nematode diversity. Mar. Ecol. 2010, 31, 6–20. [Google Scholar] [CrossRef] [Green Version]
- Poinar, G.O., Jr. The Natural History of Nematodes; Prentice-Hall, Inc.: Hoboken, NJ, USA, 1983. [Google Scholar]
- Dorris, M.; De Ley, P.; Blaxter, M.L. Molecular Analysis of Nematode Diversity and the Evolution of Parasitism. Parasitol. Today 1999, 15, 188–193. [Google Scholar] [CrossRef]
- Frézal, L.; Félix, M.-A.C. C. elegans outside the Petri dish. eLife 2015, 4, e05849. [Google Scholar] [CrossRef]
- Van Goor, J.; Piatscheck, F.; Houston, D.D.; Nason, J.D. Figs, pollinators, and parasites: A longitudinal study of the effects of nematode infection on fig wasp fitness. Acta Oecologica 2018, 90, 140–150. [Google Scholar] [CrossRef]
- Bergstrom, B.J.; Rose, R.K.; Bellows, A.S. Stomach nematodes of cotton rats: Parasites, commensals, or mutualists? J. Mammal. 2019, 100, 1831–1836. [Google Scholar] [CrossRef]
- Pryor, G.S.; Bjorndal, K.A. Effects of the nematode Gyrinicola batrachiensis on development, gut morphology, and fermentation in bullfrog tadpoles (Rana catesbeiana): A novel mutualism. J. Exp. Zool. Part A Comp. Exp. Biol. 2005, 303A, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Van Goor, J.; Piatscheck, F.; Houston, D.D.; Nason, J.D. Differential effects of nematode infection on pollinating and non-pollinating fig wasps: Can shared antagonism provide net benefits to a mutualism? J. Anim. Ecol. 2021, 90, 1764–1775. [Google Scholar] [CrossRef]
- Van Voorhies, W.A.; Fuchs, J.; Thomas, S. The longevity of Caenorhabditis elegans in soil. Biol. Lett. 2005, 1, 247–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consortium, C.e.S. Genome sequence of the nematode C. elegans: A platform for investigating biology. Science 1998, 282, 2012–2018. [Google Scholar] [CrossRef]
- Stevens, L.; Félix, M.-A.; Beltran, T.; Braendle, C.; Caurcel, C.; Fausett, S.; Fitch, D.; Frézal, L.; Gosse, C.; Kaur, T.; et al. Comparative genomics of 10 new Caenorhabditis species. Evol. Lett. 2019, 3, 217–236. [Google Scholar] [CrossRef] [Green Version]
- Leroy, S.; Duperray, C.; Morand, S. Flow cytometry for parasite nematode genome size measurement. Mol. Biochem. Parasitol. 2003, 128, 91–93. [Google Scholar] [CrossRef]
- Montarry, J.; Mimee, B.; Danchin, E.G.J.; Koutsovoulos, G.D.; Ste-Croix, D.T.; Grenier, E. Recent Advances in Population Genomics of Plant-Parasitic Nematodes. Phytopathology 2021, 111, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Rödelsperger, C.; Streit, A.; Sommer, R.J. Structure, Function and Evolution of The Nematode Genome. eLS 2013, 1, 1–9. [Google Scholar] [CrossRef]
- Kumar, S.; Koutsovoulos, G.; Kaur, G.; Blaxter, M. Toward 959 nematode genomes. Worm 2012, 1, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Denver, D.; Burnette, K.; Raboin, M. Reproductive mode evolution in nematodes: Insights from molecular phylogenies and recently discovered species. Mol. Phylogenet. Evol. 2011, 61, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Susoy, V.; Herrmann, M.; Kanzaki, N.; Kruger, M.; Nguyen, C.N.; Rödelsperger, C.; Röseler, W.; Weiler, C.; Giblin-Davis, R.M.; Ragsdale, E.J.; et al. Large-scale diversification without genetic isolation in nematode symbionts of figs. Sci. Adv. 2016, 2, e1501031. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, A.; Dyka, A.; Nemetschke, L.; Grant, W.N.; Streit, A. Parastrongyloides trichosuri suggests that XX/XO sex determination is ancestral in Strongyloididae (Nematoda). Parasitology 2013, 140, 1822. [Google Scholar] [CrossRef]
- Nigon, V. Les Modalités de la Reproduction et le Déterminisme du Sexe Chez Quelques Nématodes Libres; Masson: Paris, France, 1949. [Google Scholar]
- Tandonnet, S.; Koutsovoulos, G.D.; Adams, S.; Cloarec, D.; Parihar, M.; Blaxter, M.L.; Pires-Dasilva, A. Chromosome-Wide Evolution and Sex Determination in the Three-Sexed Nematode Auanema rhodensis. G3 2019, 9, 1211–1230. [Google Scholar] [CrossRef] [Green Version]
- Foster, J.M.; Grote, A.; Mattick, J.; Tracey, A.; Tsai, Y.-C.; Chung, M.; Cotton, J.A.; Clark, T.A.; Geber, A.; Holroyd, N.; et al. Sex chromosome evolution in parasitic nematodes of humans. Nat. Commun. 2020, 11, 1964. [Google Scholar] [CrossRef]
- Post, R. The chromosomes of the Filariae. Filaria J. 2005, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Hodgkin, J. Exploring the Envelope: Systematic Alteration in the Sex-Determination System of the Nematode Caenorhabditis elegans. Genetics 2002, 162, 767–780. [Google Scholar] [CrossRef]
- Haag, E.; Helder, J.; Mooijman, P.; Yin, D.; Hu, S. The Evolution of Uniparental Reproduction in Rhabditina Nematodes: Phylogenetic Patterns, Developmental Causes, and Surprising Consequences. In Transitions between Sexual Systems; Springer: Cham, Switzerland, 2018; pp. 99–122. [Google Scholar] [CrossRef]
- Cutter, A.D.; Morran, L.T.; Phillips, P.C. Males, Outcrossing, and Sexual Selection in Caenorhabditis Nematodes. Genetics 2019, 213, 27–57. [Google Scholar] [CrossRef] [PubMed]
- Cutter, A.D.; Baird, S.E.; Charlesworth, D. High nucleotide polymorphism and rapid decay of linkage disequilibrium in wild populations of Caenorhabditis remanei. Genetics 2006, 174, 901–913. [Google Scholar] [CrossRef] [Green Version]
- Haag, E.; Ackerman, A. Intraspecific variation in fem-3 and tra-2, two rapidly coevolving nematode sex-determining genes. Gene 2005, 349, 35–42. [Google Scholar] [CrossRef]
- Derycke, S.; Backeljau, T.; Moens, T. Dispersal and gene flow in free-living marine nematodes. Front. Zool. 2013, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Gorton, M.J.; Kasl, E.L.; Detwiler, J.T.; Criscione, C.D. Testing local-scale panmixia provides insights into the cryptic ecology, evolution, and epidemiology of metazoan animal parasites. Parasitology 2012, 139, 981. [Google Scholar] [CrossRef]
- Renahan, T.; Lo, W.-S.; Werner, M.S.; Rochat, J.; Herrmann, M.; Sommer, R.J. Nematode biphasic ‘boom and bust’ dynamics are dependent on host bacterial load while linking dauer and mouth-form polyphenisms. Environ. Microbiol. 2021. [Google Scholar] [CrossRef]
- Riddle, D.L.; Swanson, M.M.; Albert, P.S. Interacting genes in nematode dauer larva formation. Nature 1981, 290, 668–671. [Google Scholar] [CrossRef]
- Lee, H.; Choi, M.-K.; Lee, D.; Kim, H.-S.; Hwang, H.; Kim, H.; Park, S.; Paik, Y.-K.; Lee, J. Nictation, a dispersal behavior of the nematode Caenorhabditis elegans, is regulated by IL2 neurons. Nat. Neurosci. 2012, 15, 107. [Google Scholar] [CrossRef]
- Haag, E.S.; Lo, T.W. How to Make a Billion Parasites. Dev. Cell. 2018, 45, 147–148. [Google Scholar] [CrossRef]
- Adamson, M.L. Evolutionary Biology of the Oxyurida (Nematoda): Biofacies of a Haplodiploid Taxon. In Advances in Parasitology; Baker, J.R., Muller, R., Eds.; Academic Press: Cambridge, MA, USA, 1989; Volume 28, pp. 175–228. [Google Scholar]
- Lewis, E.E.; Gaugler, R. Entomopathogenic nematode (Rhabdita: Steinernematidae) sex ratio relates to foraging strategy. J. Invertebr. Pathol. 1994, 64, 238–242. [Google Scholar] [CrossRef]
- Zenner, A.N.R.L.; O’Callaghan, K.M.; Griffin, C.T. Lethal Fighting in Nematodes Is Dependent on Developmental Pathway: Male-Male Fighting in the Entomopathogenic Nematode Steinernema longicaudum. PLoS ONE 2014, 9, e89385. [Google Scholar] [CrossRef] [Green Version]
- Alsaiyah, M.A.M.; Ebssa, L.; Zenner, A.; O’Callaghan, K.M.; Griffin, C.T. Sex ratios and sex-biased infection behaviour in the Entomopathogenic nematode genus Steinernema. Int. J. Parasitol. 2009, 39, 725–734. [Google Scholar] [CrossRef] [Green Version]
- Poulin, R. Population abundance and sex ratio in dioecious helminth parasites. Oecologia 1997, 111, 375–380. [Google Scholar] [CrossRef]
- Haukisalmi, V.; Henttonen, H.; Vikman, P. Variability of sex ratio, mating probability and egg production in an intestinal nematode in its fluctuating host population. Int. J. Parasitol. 1996, 26, 755–763. [Google Scholar] [CrossRef]
- Eyualem, A.; Blaxter, M. Comparison of biological, molecular, and morphological methods of species identification in a set of cultured Panagrolaimus isolates. J. Nematol. 2003, 35, 119. [Google Scholar]
- Lewis, S.C.; Dyal, L.A.; Hilburn, C.F.; Weitz, S.; Liau, W.-S.; Lamunyon, C.W.; Denver, D.R. Molecular evolution in Panagrolaimus nematodes: Origins of parthenogenesis, hermaphroditism and the Antarctic species P. davidi. BMC Evol. Biol. 2009, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Baldi, C.; Cho, S.; Ellis, R.E. Mutations in Two Independent Pathways Are Sufficient to Create Hermaphroditic Nematodes. Science 2009, 326, 1002–1005. [Google Scholar] [CrossRef]
- Click, A.; Savaliya, C.H.; Kienle, S.; Herrmann, M.; Pires-Dasilva, A. Natural variation of outcrossing in the hermaphroditic nematode Pristionchus pacificus. BMC Evol. Biol. 2009, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Thorne, M.A.S.; Kagoshima, H.; Clark, M.S.; Marshall, C.J.; Wharton, D.A. Molecular Analysis of the Cold Tolerant Antarctic Nematode, Panagrolaimus davidi. PLoS ONE 2014, 9, e104526. [Google Scholar] [CrossRef] [Green Version]
- Garcia, L.R.; LeBoeuf, B.; Koo, P. Diversity in mating behavior of hermaphroditic and male-female Caenorhabditis nematodes. Genetics 2007, 175, 1761–1771. [Google Scholar] [CrossRef] [Green Version]
- Fierst, J.L.; Willis, J.H.; Thomas, C.G.; Wang, W.; Reynolds, R.M.; Ahearne, T.E.; Cutter, A.D.; Phillips, P.C. Reproductive mode and the evolution of genome size and structure in Caenorhabditis nematodes. PLoS Genet. 2015, 11, e1005323. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.G.; Li, R.; Smith, H.E.; Woodruff, G.C.; Oliver, B.; Haag, E.S. Simplification and desexualization of gene expression in self-fertile nematodes. Curr. Biol. 2012, 22, 2167–2172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, D.; Schwarz, E.M.; Thomas, C.G.; Felde, R.L.; Korf, I.F.; Cutter, A.D.; Schartner, C.M.; Ralston, E.J.; Meyer, B.J.; Haag, E.S. Rapid genome shrinkage in a self-fertile nematode reveals sperm competition proteins. Science 2018, 359, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, T.S.; Yang, F.-J.; Lo, Y.-H.; Chang, T.C.; Hsu, J.-C.; Kao, C.-Y.; Wang, J. Non-Mendelian assortment of homologous autosomes of different sizes in males is the ancestral state in the Caenorhabditis lineage. Sci. Rep. 2017, 7, 12819. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, P.J.; Wang, G.J.; Keller, L. Chromosome Size Differences May Affect Meiosis and Genome Size. Science 2010, 329, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimond, C.; Jovelin, R.; Han, S.; Ferrari, C.; Cutter, A.D.; Braendle, C. Outbreeding depression with low genetic variation in selfing Caenorhabditis nematodes. Evolution 2013, 67, 3087–3101. [Google Scholar] [CrossRef]
- Hodgkin, J. Male Phenotypes and Mating Efficiency in Caenorhabditis elegans. Genetics 1983, 103, 43–64. [Google Scholar] [CrossRef]
- Nishimura, H.; L’Hernault, S.W. Spermatogenesis-defective (spe) mutants of the nematode Caenorhabditis elegans provide clues to solve the puzzle of male germline functions during reproduction. Dev. Dyn. 2010, 239, 1502–1514. [Google Scholar] [CrossRef] [Green Version]
- Yin, D.; Haag, E.S. Evolution of sex ratio through gene loss. Proc. Natl. Acad. Sci. USA 2019, 116, 12919–12924. [Google Scholar] [CrossRef] [Green Version]
- Katju, V.; LaBeau, E.M.; Lipinski, K.J.; Bergthorsson, U. Sex change by gene conversion in a Caenorhabditis elegans fog-2 mutant. Genetics 2008, 180, 669–672. [Google Scholar] [CrossRef] [Green Version]
- Félix, M.-A.; Duveau, F. Population dynamics and habitat sharing of natural populations of Caenorhabditis elegans and C. briggsae. BMC Biol. 2012, 10, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinya, R.; Hasegawa, K.; Chen, A.; Kanzaki, N.; Sternberg, P.W. Evidence of Hermaphroditism and Sex Ratio Distortion in the Fungal Feeding Nematode Bursaphelenchus okinawaensis. G3 2014, 4, 1907–1917. [Google Scholar] [CrossRef] [Green Version]
- L’Hernault, S.W.; Shakes, D.C.; Ward, S. Developmental genetics of chromosome I spermatogenesis-defective mutants in the nematode Caenorhabditis elegans. Genetics 1988, 120, 435–452. [Google Scholar] [CrossRef]
- L’Hernault, S.W.; Roberts, T.M. Chapter 12 Cell Biology of Nematode Sperm. In Methods in Cell Biology; Epstein, H.F., Shakes, D.C., Eds.; Academic Press: Cambridge, MA, USA, 1995; Volume 48, pp. 273–301. [Google Scholar]
- Singson, A. Every Sperm Is Sacred: Fertilization in Caenorhabditis elegans. Dev. Biol. 2001, 230, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.; Nguyen, V.; Lee, M.; Kosinski, M.; Schedl, T.; Caprioli, R.; Greenstein, D. A Sperm Cytoskeletal Protein That Signals Oocyte Meiotic Maturation and Ovulation. Science 2001, 291, 2144–2147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leidel, S.; Gönczy, P. Centrosome Duplication and Nematodes: Recent Insights from an Old Relationship. Dev. Cell 2005, 9, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Bornens, M. The Centrosome in Cells and Organisms. Science 2012, 335, 422–426. [Google Scholar] [CrossRef]
- Browning, H.; Strome, S. A sperm-supplied factor required for embryogenesis in C. elegans. Development 1996, 122, 391–404. [Google Scholar] [CrossRef]
- Johnston, W.L.; Krizus, A.; Dennis, J.W. Eggshell chitin and chitin-interacting proteins prevent polyspermy in C. elegans. Curr. Biol. 2010, 20, 1932–1937. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.R.; Stanfield, G.M. TRY-5 Is a Sperm-Activating Protease in Caenorhabditis elegans Seminal Fluid. PLoS Genet. 2011, 7, e1002375. [Google Scholar] [CrossRef]
- Rudel, D.; Riebesell, M.; Sommer, R.J. Gonadogenesis in Pristionchus pacificus and organ evolution: Development, adult morphology and cell–cell interactions in the hermaphrodite gonad. Dev. Biol. 2005, 277, 200–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haag, E.S. Chapter 3 Caenorhabditis nematodes as a model for the adaptive evolution of germ cells. Curr. Top. Dev. Biol. 2009, 86, 43–66. [Google Scholar]
- Wei, Q.; Zhao, Y.; Guo, Y.; Stomel, J.; Stires, R.; Ellis, R.E. Co-option of alternate sperm activation programs in the evolution of self-fertile nematodes. Nat. Commun. 2014, 5, 5888. [Google Scholar] [CrossRef] [Green Version]
- Haag, E.S.; Fitch, D.H.A.; Delattre, M. From “the Worm” to “the Worms” and Back Again: The Evolutionary Developmental Biology of Nematodes. Genetics 2018, 210, 397–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clifford, R.; Lee, M.H.; Nayak, S.; Ohmachi, M.; Giorgini, F.; Schedl, T. FOG-2, a novel F-box containing protein, associates with the GLD-1 RNA binding protein and directs male sex determination in the C. elegans hermaphrodite germline. Development 2000, 127, 5265–5276. [Google Scholar] [CrossRef]
- Schedl, T.; Kimble, J. fog-2, a germ-line-specific sex determination gene required for hermaphrodite spermatogenesis in Caenorhabditis elegans. Genetics 1988, 119, 43–61. [Google Scholar] [CrossRef]
- Jan, E.; Motzny, C.K.; Graves, L.E.; Goodwin, E.B. The STAR protein, GLD-1, is a translational regulator of sexual identity in Caenorhabditis elegans. EMBO J. 1999, 18, 258–269. [Google Scholar] [CrossRef] [Green Version]
- Nayak, S.; Goree, J.; Schedl, T. fog-2 and the Evolution of Self-Fertile Hermaphroditism in Caenorhabditis. PLoS Biol. 2005, 3, e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Lang, S.; Ellis, R. Independent recruitment of F-box genes to regulate hermaphrodite development during nematode evolution. Curr. Biol. 2009, 19, 1853–1860. [Google Scholar] [CrossRef] [Green Version]
- Hill, R.C.; de Carvalho, C.E.; Salogiannis, J.; Schlager, B.; Pilgrim, D.; Haag, E.S. Genetic flexibility in the convergent evolution of hermaphroditism in Caenorhabditis nematodes. Dev. Cell 2006, 10, 531–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beadell, A.V.; Liu, Q.; Johnson, D.M.; Haag, E.S. Independent recruitments of a translational regulator in the evolution of self-fertile nematodes. Proc. Natl. Acad. Sci. USA 2011, 108, 19672–19677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Stumpf, C.; Wickens, M.; Haag, E.S. Context-dependent function of a conserved translational regulatory module. Development 2012, 139, 1509–1521. [Google Scholar] [CrossRef] [Green Version]
- Ellis, R.E.; Lin, S.Y. The evolutionary origins and consequences of self-fertility in nematodes. F1000Prime Rep. 2014, 6, 62. [Google Scholar] [CrossRef] [Green Version]
- Ward, S.; Argon, Y.; Nelson, G.A. Sperm morphogenesis in wild-type and fertilization-defective mutants of Caenorhabditis elegans. J. Cell. Biol. 1981, 91, 26–44. [Google Scholar] [CrossRef]
- Albertson, D.G.; Thomson, J.N. Segregation of holocentric chromosomes at meiosis in the nematode, Caenorhabditis elegans. Chromosome Res. 1993, 1, 15–26. [Google Scholar] [CrossRef]
- Ting, J.J.; Woodruff, G.C.; Leung, G.; Shin, N.-R.; Cutter, A.D.; Haag, E.S. Intense Sperm-Mediated Sexual Conflict Promotes Reproductive Isolation in Caenorhabditis Nematodes. PLoS Biol. 2014, 12, e1001915. [Google Scholar] [CrossRef] [Green Version]
- Zograf, J. Ultrastructure of spermatogenesis and sperm of the free-living soil nematode Panagrellus redivivus (Rhabditida: Panagrolaimidae). Russ. J. Nematol. 2014, 22, 39–48. [Google Scholar]
- Fisher, H.S.; Hoekstra, H.E. Competition drives cooperation among closely related sperm of deer mice. Nature 2010, 463, 801–803. [Google Scholar] [CrossRef] [PubMed]
- LaMunyon, C.W.; Ward, S. Evolution of sperm size in nematodes: Sperm competition favours larger sperm. Proc. Biol. Sci. 1999, 266, 263–267. [Google Scholar] [CrossRef]
- Palopoli, M.F.; Peden, C.; Woo, C.; Akiha, K.; Ary, M.; Cruze, L.; Anderson, J.L.; Phillips, P.C. Natural and experimental evolution of sexual conflict within Caenorhabditis nematodes. BMC Evol. Biol. 2015, 15, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, J.M.; Chavez, D.R.; Stanfield, G.M. COMP-1 promotes competitive advantage of nematode sperm. eLife 2015, 4, e05423. [Google Scholar] [CrossRef] [PubMed]
- Dolgin, E.S.; Charlesworth, B.; Baird, S.E.; Cutter, A.D. Inbreeding and outbreeding depression in Caenorhabditis nematodes. Evol. Int. J. Org. Evol. 2007, 61, 1339–1352. [Google Scholar] [CrossRef] [PubMed]
- Lahl, V.; Sadler, B.; Schierenberg, E. Egg development in parthenogenetic nematodes: Variations in meiosis and axis formation. Int. J. Dev. Biol. 2006, 50, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Fradin, H.; Kiontke, K.; Zegar, C.; Gutwein, M.; Lucas, J.; Kovtun, M.; Corcoran, D.L.; Baugh, L.R.; Fitch, D.H.A.; Piano, F.; et al. Genome Architecture and Evolution of a Unichromosomal Asexual Nematode. Curr. Biol. 2017, 27, 2928–2939.e6. [Google Scholar] [CrossRef] [Green Version]
- Hiraki, H.; Kagoshima, H.; Kraus, C.; Schiffer, P.H.; Ueta, Y.; Kroiher, M.; Schierenberg, E.; Kohara, Y. Genome analysis of Diploscapter coronatus: Insights into molecular peculiarities of a nematode with parthenogenetic reproduction. BMC Genom. 2017, 18, 478. [Google Scholar] [CrossRef]
- Castagnone-Sereno, P.; Danchin, E.G.J.; Perfus-Barbeoch, L.; Abad, P. Diversity and Evolution of Root-Knot Nematodes, Genus Meloidogyne: New Insights from the Genomic Era. Annu. Rev. Phytopathol. 2013, 51, 203–220. [Google Scholar] [CrossRef]
- Galis, F.; van Alphen, J.J.M. Parthenogenesis and developmental constraints. Evol. Dev. 2020, 22, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Viney, M.E. Developmental switching in the parasitic nematode strongyloides ratti. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1996, 263, 201–208. [Google Scholar] [CrossRef]
- Harvey, S.C.; Gemmill, A.W.; Read, A.F.; Viney, M.E. The control of morph development in the parasitic nematode Strongyloides ratti. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2000, 267, 2057–2063. [Google Scholar] [CrossRef] [Green Version]
- Lamelza, P.; Young, J.M.; Noble, L.M.; Caro, L.; Isakharov, A.; Palanisamy, M.; Rockman, M.V.; Malik, H.S.; Ailion, M. Hybridization promotes asexual reproduction in Caenorhabditis nematodes. PLoS Genet. 2019, 15, e1008520. [Google Scholar] [CrossRef] [Green Version]
- Launay, C.; Félix, M.-A.; Dieng, J.; Delattre, M. Diversification and hybrid incompatibility in auto-pseudogamous species of Mesorhabditis nematodes. BMC Evol. Biol. 2020, 20, 105. [Google Scholar] [CrossRef]
- Grosmaire, M.; Launay, C.; Siegwald, M.; Brugière, T.; Estrada-Virrueta, L.; Berger, D.; Burny, C.; Modolo, L.; Blaxter, M.; Meister, P.; et al. Males as somatic investment in a parthenogenetic nematode. Science 2019, 363, 1210–1213. [Google Scholar] [CrossRef] [PubMed]
- Castagnone-Sereno, P. Genetic variability and adaptive evolution in parthenogenetic root-knot nematodes. Heredity 2006, 96, 282–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.L.; Thomas, V.P.; Williamson, V.M. Meiotic parthenogenesis in a root-knot nematode results in rapid genomic homozygosity. Genetics 2007, 176, 1483–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gendron, A.; Marcogliese, D.; Barbeau, S.; Christin, M.S.; Brousseau, P.; Ruby, S.; Cyr, D.; Fournier, M. Exposure of leopard frog to a pesticide mixture affects life history characteristics of the lungworm Rhabdias ranae. Oecologia 2003, 135, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Runey, W.M.; Runey, G.L.; Lauter, F.H. Gametogenesis and fertilization in Rhabdias ranae Walton 1929: I. The parasitic hermaphrodite. J. Parasitol. 1978, 64, 1008–1014. [Google Scholar] [CrossRef] [PubMed]
- Dare, O.K.; Forbes, M.R. Patterns of infection by lungworms, Rhabdias ranae and Haematoloechus spp., in northern leopard frogs: A relationship between sex and parasitism. J. Parasitol. 2009, 95, 275–280. [Google Scholar] [CrossRef]
- Chaudhuri, J.; Bose, N.; Tandonnet, S.; Adams, S.; Zuco, G.; Kache, V.; Parihar, M.; von Reuss, S.H.; Schroeder, F.C.; Pires-daSilva, A. Mating dynamics in a nematode with three sexes and its evolutionary implications. Sci. Rep. 2015, 5, 17676. [Google Scholar] [CrossRef] [Green Version]
- Kanzaki, N.; Kiontke, K.; Tanaka, R.; Hirooka, Y.; Schwarz, A.; Müller-Reichert, T.; Chaudhuri, J.; Pires-daSilva, A. Description of two three-gendered nematode species in the new genus Auanema (Rhabditina) that are models for reproductive mode evolution. Sci. Rep. 2017, 7, 11135. [Google Scholar] [CrossRef] [Green Version]
- McCaig, C.M.; Lin, X.; Farrell, M.; Rehain-Bell, K.; Shakes, D.C. Germ cell cysts and simultaneous sperm and oocyte production in a hermaphroditic nematode. Dev. Biol. 2017, 430, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Shakes, D.C.; Neva, B.J.; Huynh, H.; Chaudhuri, J.; Pires-Dasilva, A. Asymmetric spermatocyte division as a mechanism for controlling sex ratios. Nat. Commun. 2011, 2, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tandonnet, S.; Farrell, M.C.; Koutsovoulos, G.D.; Blaxter, M.L.; Parihar, M.; Sadler, P.L.; Shakes, D.C.; Pires-Dasilva, A. Sex- and Gamete-Specific Patterns of X Chromosome Segregation in a Trioecious Nematode. Curr. Biol. 2018, 28, 93–99.e3. [Google Scholar] [CrossRef]
- Winter, E.S.; Schwarz, A.; Fabig, G.; Feldman, J.L.; Pires-Dasilva, A.; Müller-Reichert, T.; Sadler, P.L.; Shakes, D.C. Cytoskeletal variations in an asymmetric cell division support diversity in nematode sperm size and sex ratios. Development 2017, 144, 3253–3263. [Google Scholar] [CrossRef] [Green Version]
- Hodgkin, J.; Horvitz, H.R.; Brenner, S. Nondisjunction Mutants of the Nematode Caenorhabditis elegans. Genetics 1979, 91, 67–94. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.M.; Wong, C.; Bhalla, N.; Carlton, P.M.; Weiser, P.; Meneely, P.M.; Dernburg, A.F. HIM-8 binds to the X chromosome pairing center and mediates chromosome-specific meiotic synapsis. Cell 2005, 123, 1051–1063. [Google Scholar] [CrossRef] [Green Version]
- Cortes, D.B.; McNally, K.L.; Mains, P.E.; McNally, F.J. The asymmetry of female meiosis reduces the frequency of inheritance of unpaired chromosomes. eLife 2015, 4, e06056. [Google Scholar] [CrossRef]
- Janssen, T.; Karssen, G.; Topalović, O.; Coyne, D.; Bert, W. Integrative taxonomy of root-knot nematodes reveals multiple independent origins of mitotic parthenogenesis. PLoS ONE 2017, 12, e0172190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez Akimori, D.; Dalessandro, E.J.; Nolan, T.J.; Stieha, C.R.; Lok, J.B.; Stoltzfus, J.D.C. Transcriptional profiles in Strongyloides stercoralis males reveal deviations from the Caenorhabditis sex determination model. Sci. Rep. 2021, 11, 8254. [Google Scholar] [CrossRef]
- Streit, A. How to become a parasite without sex chromosomes: A hypothesis for the evolution of Strongyloides spp. and related nematodes. Parasitology 2014, 141, 1244–1254. [Google Scholar] [CrossRef]
- Korpelainen, H. Sex ratios and conditions required for environmental sex determination in animals. Biol. Rev. 1990, 65, 147–184. [Google Scholar] [CrossRef]
- Adams, J.; Greenwood, P.; Naylor, C. Evolutionary Aspects of Environmental Sex Determination. Int. J. Invertebr. Reprod. Dev. 1987, 11, 123–135. [Google Scholar] [CrossRef]
- Hunt, V.L.; Tsai, I.J.; Coghlan, A.; Reid, A.J.; Holroyd, N.; Foth, B.J.; Tracey, A.; Cotton, J.A.; Stanley, E.J.; Beasley, H.; et al. The genomic basis of parasitism in the Strongyloides clade of nematodes. Nat. Genet. 2016, 48, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Nemetschke, L.; Eberhardt, A.G.; Hertzberg, H.; Streit, A. Genetics, chromatin diminution, and sex chromosome evolution in the parasitic nematode genus Strongyloides. Curr. Biol. 2010, 20, 1687–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albertson, D.G.; Nwaorgu, O.C.; Sulston, J.E. Chromatin diminution and a chromosomal mechanism of sexual differentiation in Strongyloides papillosus. Chromosoma 1979, 75, 75–87. [Google Scholar] [CrossRef]
- Kimura, K.D.; Tissenbaum, H.A.; Liu, Y.; Ruvkun, G. daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science 1997, 277, 942–946. [Google Scholar] [CrossRef] [PubMed]
- Ren, P.; Lim, C.S.; Johnsen, R.; Albert, P.S.; Pilgrim, D.; Riddle, D.L. Control of C. elegans larval development by neuronal expression of a TGF-beta homolog. Science 1996, 274, 1389–1391. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, A.; Streit, A.; Antebi, A.; Sommer, R.J. A conserved endocrine mechanism controls the formation of dauer and infective larvae in nematodes. Curr. Biol. 2009, 19, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Moens, T.; Vincx, M. Temperature and salinity constraints on the life cycle of two brackish-water nematode species. J. Exp. Mar. Biol. Ecol. 2000, 243, 115–135. [Google Scholar] [CrossRef]
- Triantaphyllou, A. Environmental sex differentiation of nematodes in relation to pest management. Annu. Rev. Phytopathol. 1973, 11, 441–462. [Google Scholar] [CrossRef]
- Anjam, M.S.; Shah, S.J.; Matera, C.; Różańska, E.; Sobczak, M.; Siddique, S.; Grundler, F.M.W. Host factors influence the sex of nematodes parasitizing roots of Arabidopsis thaliana. Plant Cell Environ. 2020, 43, 1160–1174. [Google Scholar] [CrossRef] [Green Version]
- Christie, J.R. Some observations on sex in the Mermithidae. J. Exp. Zool. 1929, 53, 59–76. [Google Scholar] [CrossRef]
- Blackmore, M.; Charnov, E. Adaptive variation in environmental sex determination in a nematode. Am. Nat. 1989, 134, 817–823. [Google Scholar] [CrossRef]
- Tingley, G.; Anderson, R. Environmental sex determination and density-dependent population regulation in the entomogenous nematode Romanomermis culcivorax. Parasitology 1986, 92, 431–449. [Google Scholar] [CrossRef]
- Van Goor, J. A Worm’s Tale: An Evaluation of the Natural History and Infectious Dynamics of Parasitodiplogaster nematodes. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2018. [Google Scholar]
- Van Goor, J.; Herre, E.A.; Gomez, A.; Nason, J.D. Extraordinarily precise nematode sex ratios: Adaptive responses to vanishingly rare mating options. BioRxiv 2021. [Google Scholar] [CrossRef]
- Anderson, A.G.; Bubrig, L.T.; Fierst, J.L. Environmental stress maintains trioecy in nematode worms. Evolution 2020, 74, 518–527. [Google Scholar] [CrossRef]
- Shih, P.-Y.; Lee, J.S.; Shinya, R.; Kanzaki, N.; Pires-daSilva, A.; Badroos, J.M.; Goetz, E.; Sapir, A.; Sternberg, P.W. Newly Identified Nematodes from Mono Lake Exhibit Extreme Arsenic Resistance. Curr. Biol. 2019, 29, 3339–3344.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhuri, J.; Kache, V.; Pires-daSilva, A. Regulation of sexual plasticity in a nematode that produces males, females, and hermaphrodites. Curr. Biol. 2011, 21, 1548–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Félix, M.A. Alternative morphs and plasticity of vulval development in a rhabditid nematode species. Dev. Genes Evol. 2004, 214, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Robles, P.; Turner, A.; Zuco, G.; Adams, S.; Paganopolou, P.; Winton, M.; Hill, B.; Kache, V.; Bateson, C.; Pires-daSilva, A. Parental energy-sensing pathways control intergenerational offspring sex determination in the nematode Auanema freiburgensis. BMC Biol. 2021, 19, 102. [Google Scholar] [CrossRef]
- Haldane, J.B.S. Disease and evolution. Ric. Sci. 1949, 19, 68–76. [Google Scholar]
- Wharton, D.A. The environmental physiology of Antarctic terrestrial nematodes: A review. J. Comp. Physiol. B 2003, 173, 621–628. [Google Scholar] [CrossRef]
- Zeppilli, D.; Bellec, L.; Cambon-Bonavita, M.-A.; Decraemer, W.; Fontaneto, D.; Fuchs, S.; Gayet, N.; Mandon, P.; Michel, L.N.; Portail, M.; et al. Ecology and trophic role of Oncholaimus dyvae sp. nov. (Nematoda: Oncholaimidae) from the lucky strike hydrothermal vent field (Mid-Atlantic Ridge). BMC Zool. 2019, 4, 6. [Google Scholar] [CrossRef]
- Ahmed, N.H. Cultivation of parasites. Trop. Parasitol. 2014, 4, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.E.; Boone, B.E. Next-Generation Sequencing Strategies. Cold Spring Harb. Perspect Med. 2019, 9, a025791. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Goor, J.; Shakes, D.C.; Haag, E.S. Fisher vs. the Worms: Extraordinary Sex Ratios in Nematodes and the Mechanisms that Produce Them. Cells 2021, 10, 1793. https://doi.org/10.3390/cells10071793
Van Goor J, Shakes DC, Haag ES. Fisher vs. the Worms: Extraordinary Sex Ratios in Nematodes and the Mechanisms that Produce Them. Cells. 2021; 10(7):1793. https://doi.org/10.3390/cells10071793
Chicago/Turabian StyleVan Goor, Justin, Diane C. Shakes, and Eric S. Haag. 2021. "Fisher vs. the Worms: Extraordinary Sex Ratios in Nematodes and the Mechanisms that Produce Them" Cells 10, no. 7: 1793. https://doi.org/10.3390/cells10071793
APA StyleVan Goor, J., Shakes, D. C., & Haag, E. S. (2021). Fisher vs. the Worms: Extraordinary Sex Ratios in Nematodes and the Mechanisms that Produce Them. Cells, 10(7), 1793. https://doi.org/10.3390/cells10071793