
Improving In Vitro Culture of Human Male Fetal Germ Cells

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics for Human Material
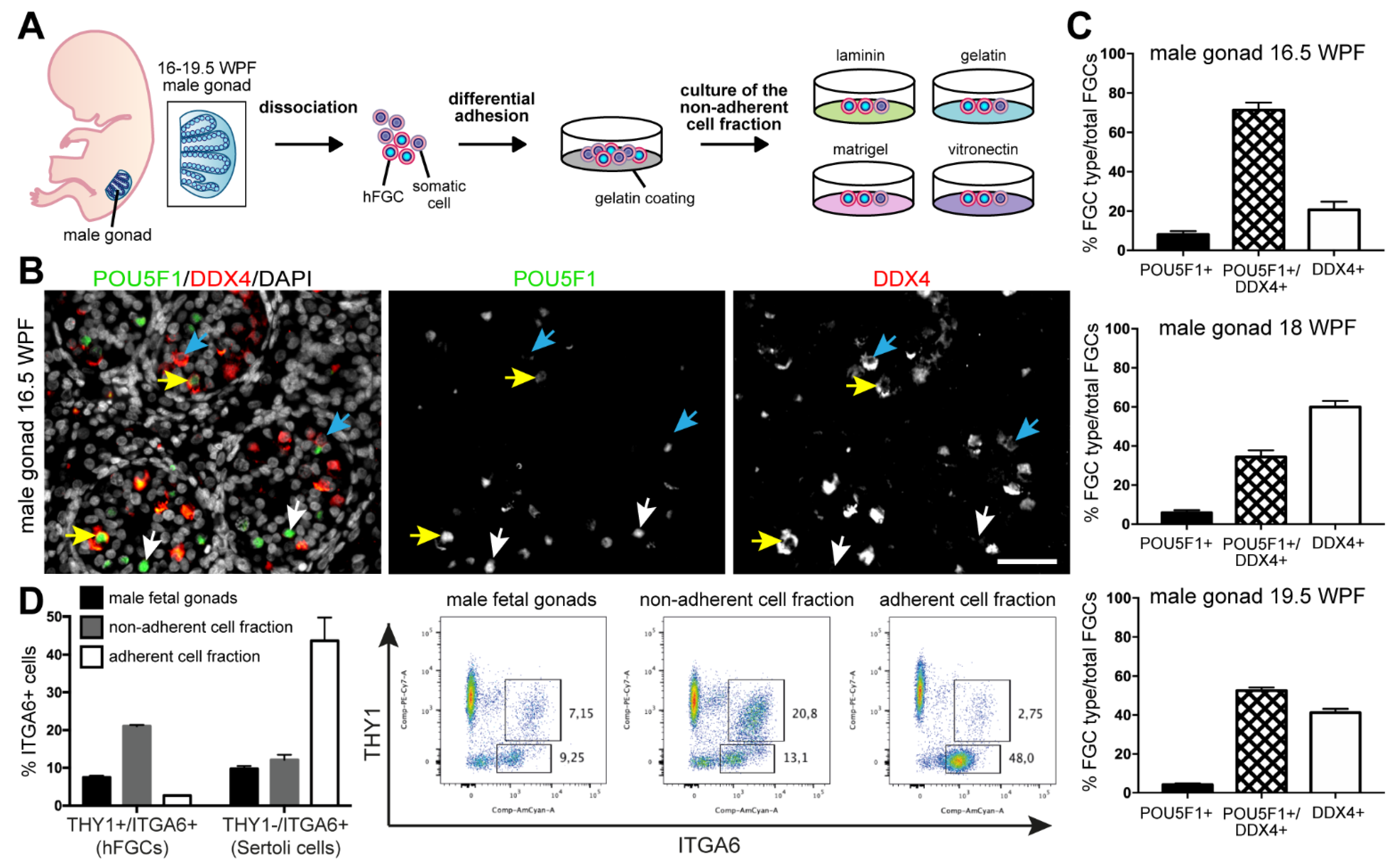
2.2. Isolation of Male hFGCs
2.3. FACS Analysis of Male hFGCs
2.4. Culture of Male hFGCs
2.5. Immunofluorescence and Histology
2.6. Imaging, Quantification and Statistical Analysis
2.7. Analysis of Single-Cell Transcriptomics Data
3. Results
3.1. Second Trimester Male Fetal Gonads Contained Populations of hFGCs Differing in the Expression of POU5F1 and DDX4
3.2. The Substrates Gelatin and Vitronectin Sustained In Vitro Culture of hFGCs
3.3. Gene Expression Single Cell RNA Sequencing Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saitou, M.; Miyauchi, H. Gametogenesis from Pluripotent Stem Cells. Cell Stem Cell 2016, 18, 721–735. [Google Scholar] [CrossRef]
- Li, L.; Li, L.; Li, Q.; Liu, X.; Ma, X.; Yong, J.; Gao, S.; Wu, X.; Wei, Y.; Wang, X.; et al. Dissecting the epigenomic dynamics of human fetal germ cell development at single-cell resolution. Cell Res. 2021, 31, 463–477. [Google Scholar] [CrossRef]
- Li, L.; Dong, J.; Yan, L.; Yong, J.; Liu, X.; Hu, Y.; Fan, X.; Wu, X.; Guo, H.; Wang, X.; et al. Single-Cell RNA-Seq Analysis Maps Development of Human Germline Cells and Gonadal Niche Interactions. Cell Stem Cell 2017, 20, 858–873.e4. [Google Scholar] [CrossRef] [Green Version]
- Overeem, A.W.; Chang, Y.W.; Spruit, J.; Roelse, C.M.; Chuva De Sousa Lopes, S.M. Ligand-Receptor Interactions Elucidate Sex-Specific Pathways in the Trajectory From Primordial Germ Cells to Gonia During Human Development. Front. Cell Dev. Biol. 2021, 9, 661243. [Google Scholar] [CrossRef] [PubMed]
- Culty, M. Gonocytes, the forgotten cells of the germ cell lineage. Birth Defects Res. Part C Embryo Today Rev. 2009, 87, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Hermann, B.P.; Sukhwani, M.; Winkler, F.; Pascarella, J.N.; Peters, K.A.; Sheng, Y.; Valli, H.; Rodriguez, M.; Ezzelarab, M.; Dargo, G.; et al. Spermatogonial Stem Cell Transplantation into Rhesus Testes Regenerates Spermatogenesis Producing Functional Sperm. Cell Stem Cell 2012, 11, 715–726. [Google Scholar] [CrossRef] [Green Version]
- Kanatsu-Shinohara, M.; Shinohara, T. Spermatogonial Stem Cell Self-Renewal and Development. Annu. Rev. Cell Dev. Biol. 2013, 29, 163–187. [Google Scholar] [CrossRef]
- Kobayashi, T.; Surani, M.A. On the origin of the human germline. Development 2018, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, T.; Zhang, H.; Tang, W.W.C.; Irie, N.; Withey, S.; Klisch, D.; Sybirna, A.; Dietmann, S.; Contreras, D.A.; Webb, R.; et al. Principles of early human development and germ cell program from conserved model systems. Nature 2017, 546, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.T.; Cowan, G.; Morris, K.D.; Anderson, R.A.; Fraser, H.M.; Mckenzie, K.J.; Wallace, W.H.B.; Kelnar, C.J.H.; Saunders, P.T.K.; Sharpe, R.M. Germ cell differentiation in the marmoset (Callithrix jacchus) during fetal and neonatal life closely parallels that in the human. Hum. Reprod. 2008, 23, 2755–2765. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.M.; Nakagawa, T.; Ichikawa, R.; Yoshida, S.; Simons, B.D. Mouse Germ Line Stem Cells Undergo Rapid and Stochastic Turnover. Cell Stem Cell 2010, 7, 214–224. [Google Scholar] [CrossRef] [Green Version]
- Lord, T.; Oatley, J.M. Regulation of spermatogonial stem cell maintenance and self-Renewal. In The Biology of Mammalian Spermatogonia; Springer: New York, NY, USA, 2017; pp. 91–129. ISBN 9781493975051. [Google Scholar]
- Lukassen, S.; Bosch, E.; Ekici, A.B.; Winterpacht, A. Characterization of germ cell differentiation in the male mouse through single-cell RNA sequencing. Sci. Rep. 2018, 8, 6521. [Google Scholar] [CrossRef]
- Brinster, R.L. Male germline stem cells: From mice to men. Science 2007, 316, 404–405. [Google Scholar] [CrossRef] [Green Version]
- Clermont, Y. Two classes of spermatogonial stem cells in the monkey (Cercopithecus aethiops). Am. J. Anat. 1969, 126, 57–71. [Google Scholar] [CrossRef]
- Hermann, B.P.; Sukhwani, M.; Hansel, M.C.; Orwig, K.E. Spermatogonial stem cells in higher primates: Are there differences from those in rodents? Reproduction 2010, 139, 479–493. [Google Scholar] [CrossRef] [Green Version]
- Sohni, A.; Tan, K.; Song, H.-W.; Burow, D.; de Rooij, D.G.; Laurent, L.; Hsieh, T.-C.; Rabah, R.; Hammoud, S.S.; Vicini, E.; et al. The Neonatal and Adult Human Testis Defined at the Single-Cell Level. Cell Rep. 2019, 26, 1501–1517.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, Y. Spermatogenesis in Man: A Study of the Spermatogonial Population. Fertil. Steril. 1966, 17, 705–721. [Google Scholar] [CrossRef]
- Neuhaus, N.; Yoon, J.; Terwort, N.; Kliesch, S.; Seggewiss, J.; Huge, A.; Voss, R.; Schlatt, S.; Grindberg, R.V.; Schöler, H.R. Single-cell gene expression analysis reveals diversity among human spermatogonia. Mol. Hum. Reprod. 2017, 23, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Wistuba, J.; Pock, T.; Schlatt, S.; Neuhaus, N. Spermatogonial stem cells: Updates from specification to clinical relevance. Hum. Reprod. Update 2019, 25, 275–297. [Google Scholar] [CrossRef]
- Guo, J.; Sosa, E.; Chitiashvili, T.; Nie, X.; Rojas, E.J.; Oliver, E.; Plath, K.; Hotaling, J.M.; Stukenborg, J.-B.; Clark, A.T.; et al. Single-cell analysis of the developing human testis reveals somatic niche cell specification and fetal germline stem cell establishment. Cell Stem Cell 2021, 28, 764–778.e4. [Google Scholar] [CrossRef]
- Avarbock, M.R.; Brinster, C.J.; Brinster, R.L. Reconstitution of spermatogenesis from frozen spermatogonial stem cells. Nat. Med. 1996, 2, 693–696. [Google Scholar] [CrossRef]
- Brinster, R.L.; Zimmermann, J.W. Spermatogenesis following male germ-cell transplantation. Proc. Natl. Acad. Sci. USA 1994, 91, 11298–11302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langenstroth, D.; Kossack, N.; Westernströer, B.; Wistuba, J.; Behr, R.; Gromoll, J.; Schlatt, S. Separation of somatic and germ cells is required to establish primate spermatogonial cultures. Hum. Reprod. 2014, 29, 2018–2031. [Google Scholar] [CrossRef] [Green Version]
- Nagano, M.; McCarrey, J.R.; Brinster, R.L. Primate Spermatogonial Stem Cells Colonize Mouse Testes1. Biol. Reprod. 2001, 64, 1409–1416. [Google Scholar] [CrossRef] [Green Version]
- Giudice, M.G.; de Michele, F.; Poels, J.; Vermeulen, M.; Wyns, C. Update on fertility restoration from prepubertal spermatogonial stem cells: How far are we from clinical practice? Stem Cell Res. 2017, 21, 171–177. [Google Scholar] [CrossRef]
- Reis, M.M.; Tsai, M.C.; Schlegel, P.N.; Feliciano, M.; Raffaelli, R.; Rosenwaks, Z.; Palermo, G.D. Xenogeneic transplantation of human spermatogonia. Zygote 2000, 8, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Wyns, C.; Curaba, M.; Vanabelle, B.; Van Langendonckt, A.; Donnez, J. Options for fertility preservation in prepubertal boys. Hum. Reprod. Update 2010, 16, 312–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlatt, S.; Kim, S.S.; Gosden, R. Spermatogenesis and steroidogenesis in mouse, hamster and monkey testicular tissue after cryopreservation and heterotopic grafting to castrated hosts. Reproduction 2002, 124, 339–346. [Google Scholar] [CrossRef]
- Guan, K.; Wolf, F.; Becker, A.; Engel, W.; Nayernia, K.; Hasenfuss, G. Isolation and cultivation of stem cells from adult mouse testes. Nat. Protoc. 2009, 4, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Hamra, F.K.; Chapman, K.M.; Nguyen, D.M.; Williams-Stephens, A.A.; Hammer, R.E.; Garbers, D.L. Self renewal, expansion, and transfection of rat spermatogonial stem cells in culture. Proc. Natl. Acad. Sci. USA 2005, 102, 17430–17435. [Google Scholar] [CrossRef] [Green Version]
- Kanatsu-Shinohara, M.; Shinohara, T. Culture and genetic modification of mouse germline stem cells. In Proceedings of the Annals of the New York Academy of Sciences; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2007; Volume 1120, pp. 59–71. [Google Scholar]
- Sato, T.; Katagiri, K.; Yokonishi, T.; Kubota, Y.; Inoue, K.; Ogonuki, N.; Matoba, S.; Ogura, A.; Ogawa, T. In vitro production of fertile sperm from murine spermatogonial stem cell lines. Nat. Commun. 2011, 2, 472. [Google Scholar] [CrossRef] [Green Version]
- Baazm, M.; Abolhassani, F.; Abbasi, M.; Habibi Roudkenar, M.; Amidi, F.; Beyer, C. An Improved Protocol for Isolation and Culturing of Mouse Spermatogonial Stem Cells. Cell. Reprogram. 2013, 15, 329–336. [Google Scholar] [CrossRef]
- Aponte, P.M.; Soda, T.; Teerds, K.J.; Mizrak, S.C.; van de Kant, H.J.G.; de Rooij, D.G. Propagation of bovine spermatogonial stem cells in vitro. Reproduction 2008, 136, 543–557. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Chen, X.; Zheng, Y.; Zhu, J.; Qin, Y.; Lv, Y.; Zeng, W. Long-Term Propagation of Porcine Undifferentiated Spermatogonia. Stem Cells Dev. 2017, 26, 1121–1131. [Google Scholar] [CrossRef] [Green Version]
- Stukenborg, J.-B.; Kjartansdóttir, K.R.; Reda, A.; Colon, E.; Albersmeier, J.P.; Söder, O. Male Germ Cell Development in Humans. Horm. Res. Paediatr. 2014, 81, 2–12. [Google Scholar] [CrossRef]
- Sadri-Ardekani, H.; Mizrak, S.C.; van Daalen, S.K.M.; Korver, C.M.; Roepers-Gajadien, H.L.; Koruji, M.; Hovingh, S.; de Reijke, T.M.; de la Rosette, J.J.M.C.H.; van der Veen, F.; et al. Propagation of Human Spermatogonial Stem Cells In Vitro. JAMA 2009, 302, 2127–2134. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Hai, Y.; Gong, Y.; Li, Z.; He, Z. Characterization, Isolation, and Culture of Mouse and Human Spermatogonial Stem Cells. J. Cell. Physiol. 2014, 229, 407–413. [Google Scholar] [CrossRef]
- Stukenborg, J.-B.; Wistuba, J.; Luetjens, C.M.; Elhija, M.A.; Huleihel, M.; Lunenfeld, E.; Gromoll, J.; Nieschlag, E.; Schlatt, S. Coculture of Spermatogonia With Somatic Cells in a Novel Three-Dimensional Soft-Agar-Culture-System. J. Androl. 2008, 29, 312–329. [Google Scholar] [CrossRef]
- Dong, L.; Kristensen, S.G.; Hildorf, S.; Gul, M.; Clasen-Linde, E.; Fedder, J.; Hoffmann, E.R.; Cortes, D.; Thorup, J.; Andersen, C.Y. Propagation of Spermatogonial Stem Cell-Like Cells From Infant Boys. Front. Physiol. 2019, 10, 1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Wang, Y.; Li, Y.L. Fibroblast-like cells derived from the gonadal ridges and dorsal mesenteries of human embryos as feeder cells for the culture of human embryonic germ cells. J. Biomed. Sci. 2007, 14, 617–628. [Google Scholar] [CrossRef]
- Shamblott, M.J.; Axelman, J.; Wang, S.; Bugg, E.M.; Littlefield, J.W.; Donovan, P.J.; Blumenthal, P.D.; Huggins, G.R.; Gearhart, J.D. Derivation of pluripotent stem cells from cultured human primordial germ cells. Proc. Natl. Acad. Sci. USA 1998, 95, 13726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnpenny, L.; Brickwood, S.; Spalluto, C.M.; Piper, K.; Cameron, I.T.; Wilson, D.I.; Hanley, N.A. Derivation of Human Embryonic Germ Cells: An Alternative Source of Pluripotent Stem Cells. Stem Cells 2003, 21, 598–609. [Google Scholar] [CrossRef]
- Gell, J.J.; Liu, W.; Sosa, E.; Chialastri, A.; Hancock, G.; Tao, Y.; Wamaitha, S.E.; Bower, G.; Dey, S.S.; Clark, A.T. An Extended Culture System that Supports Human Primordial Germ Cell-like Cell Survival and Initiation of DNA Methylation Erasure. Stem Cell Rep. 2020, 14, 433–446. [Google Scholar] [CrossRef] [Green Version]
- Kanatsu-Shinohara, M.; Ogonuki, N.; Inoue, K.; Miki, H.; Ogura, A.; Toyokuni, S.; Shinohara, T. Long-term proliferation in culture and germline transmission of mouse male germline stem cells. Biol. Reprod. 2003, 69, 612–616. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Wang, M.; Yuan, Y.; Wang, X.; Fu, R.; Wan, H.; Xie, M.; Liu, M.; Guo, X.; Zheng, Y.; et al. Complete Meiosis from Embryonic Stem Cell-Derived Germ Cells in Vitro. Cell Stem Cell 2016, 18, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Ye, S.; Liang, D.; Wang, P.; Fu, J.; Ma, Q.; Kong, R.; Shi, L.; Gong, X.; Chen, W.; et al. In Vitro Modeling of Human Germ Cell Development Using Pluripotent Stem Cells. Stem Cell Rep. 2018, 10, 509–523. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Liu, W.; Lukianchikov, A.; Hancock, G.V.; Zimmerman, J.; Lowe, M.G.; Kim, R.; Galic, Z.; Irie, N.; Surani, M.A.; et al. Germline competency of human embryonic stem cells depends on eomesodermin. Biol. Reprod. 2017, 97, 850–861. [Google Scholar] [CrossRef] [Green Version]
- Hammoud, S.S.; Low, D.H.P.; Yi, C.; Carrell, D.T.; Guccione, E.; Cairns, B.R. Chromatin and Transcription Transitions of Mammalian Adult Germline Stem Cells and Spermatogenesis. Cell Stem Cell 2014, 15, 239–253. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Taelman, J.; Chang, Y.W.; Boel, A.; De Sutter, P.; Heindryckx, B.; Chuva De Sousa Lopes, S.M. Sex-Specific Isolation and Propagation of Human Premeiotic Fetal Germ Cells and Germ Cell-Like Cells. Cells 2021, 10, 1214. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Kanatsu-Shinohara, M.; Ogonuki, N.; Matoba, S.; Morimoto, H.; Ogura, A.; Shinohara, T. Improved Serum- and Feeder-Free Culture of Mouse Germline Stem Cells1. Biol. Reprod. 2014, 91. [Google Scholar] [CrossRef] [Green Version]
- Butler, A.; Hoffman, P.; Smibert, P.; Papalexi, E.; Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 2018, 36, 411–420. [Google Scholar] [CrossRef]
- Huang, X.L.; Khan, M.I.; Wang, J.; Ali, R.; Ali, S.W.; Zahra, Q.A.; Kazmi, A.; Lolai, A.; Huang, Y.L.; Hussain, A.; et al. Role of receptor tyrosine kinases mediated signal transduction pathways in tumor growth and angiogenesis—New insight and futuristic vision. Int. J. Biol. Macromol. 2021, 180, 739–752. [Google Scholar] [CrossRef]
- Nicola, N.A.; Babon, J.J. Leukemia inhibitory factor (LIF). Cytokine Growth Factor Rev. 2015, 26, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Pawlak, J.B.; Blobe, G.C. TGF-β superfamily co-receptors in cancer. Dev. Dyn. 2021. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, Y.; Li, R.; Chen, G. Transcriptional factors mediating retinoic acid signals in the control of energy metabolism. Int. J. Mol. Sci. 2015, 16, 14210–14244. [Google Scholar] [CrossRef] [Green Version]
- Nickkholgh, B.; Mizrak, S.C.; Korver, C.M.; Van Daalen, S.K.M.; Meissner, A.; Repping, S.; Van Pelt, A.M.M. Enrichment of spermatogonial stem cells from long-term cultured human testicular cells. Fertil. Steril. 2014, 102. [Google Scholar] [CrossRef]
- Mizrak, S.C.; Chikhovskaya, J.V.; Sadri-Ardekani, H.; Van Daalen, S.; Korver, C.M.; Hovingh, S.E.; Roepers-Gajadien, H.L.; Raya, A.; Fluiter, K.; De Reijke, T.M.; et al. Embryonic stem cell-like cells derived from adult human testis. Hum. Reprod. 2010, 25, 158–167. [Google Scholar] [CrossRef] [Green Version]
- Zanganeh, B.M.; Rastegar, T.; Roudkenar, M.H.; Kashani, I.R.; Amidi, F.; Abolhasani, F.; Barbarestani, M. Co-Culture of Spermatogonial Stem Cells with Sertoli Cells in the Presence of Testosterone and FSH Improved Differentiation via Up-Regulation of Post Meiotic Genes. Acta Med. Iran. 1970, 51, 1–11. [Google Scholar]
- Lim, J.J.; Kim, H.J.; Kim, K.S.; Hong, J.Y.; Lee, D.R. In vitro culture-induced pluripotency of human spermatogonial stem cells. Biomed Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Kitajima, H.; Niwa, H. Clonal expansion of human pluripotent stem cells on gelatin-coated surface. Biochem. Biophys. Res. Commun. 2010, 396, 933–938. [Google Scholar] [CrossRef]
- Sahare, M.; Kim, S.M.; Otomo, A.; Komatsu, K.; Minami, N.; Yamada, M.; Imai, H. Factors supporting long-term culture of bovine male germ cells. Reprod. Fertil. Dev. 2016, 28, 2039–2050. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.R.; Kim, K.S.; Yang, Y.H.; Oh, H.S.; Lee, S.H.; Chung, T.G.; Cho, J.H.; Kim, H.J.; Yoon, T.K.; Cha, K.Y. Isolation of male germ stem cell-like cells from testicular tissue of non-obstructive azoospermic patients and differentiation into haploid male germ cells in vitro. Hum. Reprod. 2006, 21, 471–476. [Google Scholar] [CrossRef] [Green Version]
- Felding-Habermann, B.; Cheresh, D.A. Vitronectin and its receptors. Curr. Opin. Cell Biol. 1993, 5, 864–868. [Google Scholar] [CrossRef]
- Heng, B.C.; Li, J.; Chen, A.K.L.; Reuveny, S.; Cool, S.M.; Birch, W.R.; Oh, S.K.W. Translating human embryonic stem cells from 2-dimensional to 3-dimensional cultures in a defined medium on laminin- and vitronectin-coated surfaces. Stem Cells Dev. 2012, 21, 1701–1715. [Google Scholar] [CrossRef]
- Higuchi, A.; Hirad, A.H.; Kumar, S.S.; Munusamy, M.A.; Alarfaj, A.A. Thermoresponsive surfaces designed for the proliferation and differentiation of human pluripotent stem cells. Acta Biomater. 2020, 116, 162–173. [Google Scholar] [CrossRef]
- Griswold, M.D. Protein Secretions of Sertoli Cells. Int. Rev. Cytol. 1988, 110, 133–156. [Google Scholar] [CrossRef]
- El Ouali, H.; Leheup, B.P.; Gelly, J.L.; Grignon, G. Laminin ultrastructural immunolocalization in rat testis during ontogenesis. Histochemistry 1991, 95, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Nickkholgh, B.; Mizrak, S.C.; Van Daalen, S.K.M.; Korver, C.M.; Sadri-Ardekani, H.; Repping, S.; Van Pelt, A.M.M. Genetic and epigenetic stability of human spermatogonial stem cells during long-term culture. Fertil. Steril. 2014, 102, 1700–1707.e1. [Google Scholar] [CrossRef]
- Kanatsu-Shinohara, M.; Miki, H.; Inoue, K.; Ogonuki, N.; Toyokuni, S.; Ogura, A.; Shinohara, T. Long-term culture of mouse male germline stem cells under serum- or feeder-free conditions. Biol. Reprod. 2005, 72, 985–991. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Suo, L.J.; Wang, Y.F.; Shang, H.; Li, G.X.; Hu, J.H.; Li, Q.W. Effects of GDNF and LIF on mouse spermatogonial stem cells proliferation in vitro. Cytotechnology 2014, 66, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Sun, Y.C.; Ge, W.; Tan, H.; Cheng, S.F.; Yin, S.; Sun, X.F.; Li, L.; Dyce, P.; Li, J.; et al. The crucial role of activin a on the formation of primordial germ cell-like cells from skin-derived stem cells in vitro. Cell Cycle 2015, 14, 3016–3029. [Google Scholar] [CrossRef] [Green Version]
- Endo, T.; Mikedis, M.M.; Nicholls, P.K.; Page, D.C.; de Rooij, D.G. Retinoic acid and germ cell development in the ovary and testis. Biomolecules 2019, 9, 775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popovic, M.; Bialecka, M.; Gomes Fernandes, M.; Taelman, J.; Van Der Jeught, M.; De Sutter, P.; Heindryckx, B.; De Sousa Lopes, S.M.C. Human blastocyst outgrowths recapitulate primordial germ cell specification events. Mol. Hum. Reprod. 2019, 25, 519–526. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Taelman, J.; Popovic, M.; Tilleman, L.; Duthoo, E.; van der Jeught, M.; Deforce, D.; van Nieuwerburgh, F.; Menten, B.; de Sutter, P.; et al. Activin A-derived human embryonic stem cells show increased competence to differentiate into primordial germ cell-like cells. Stem Cells 2021, 39, 551–563. [Google Scholar] [CrossRef]
- Duggal, G.; Heindryckx, B.; Warrier, S.; Taelman, J.; Van Der Jeught, M.; Deforce, D.; De Sousa Lopes, S.C.; De Sutter, P. Exogenous supplementation of activin a enhances germ cell differentiation of human embryonic stem cells. Mol. Hum. Reprod. 2014, 21, 410–423. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Ohta, H.; Kurimoto, K.; Aramaki, S.; Saitou, M. Reconstitution of the mouse germ cell specification pathway in culture by pluripotent stem cells. Cell 2011, 146, 519–532. [Google Scholar] [CrossRef] [Green Version]
- Tilgner, K.; Atkinson, S.P.; Golebiewska, A.; Stojković, M.; Lako, M.; Armstrong, L. Isolation of Primordial Germ Cells from Differentiating Human Embryonic Stem Cells. Stem Cells 2008, 26, 3075–3085. [Google Scholar] [CrossRef]
- Zheng, Y.; Lei, Q.; Jongejan, A.; Mulder, C.L.; van Daalen, S.K.M.; Mastenbroek, S.; Hwang, G.; Jordan, P.W.; Repping, S.; Hamer, G. The influence of retinoic acid-induced differentiation on the radiation response of male germline stem cells. DNA Repair 2018, 70, 55–66. [Google Scholar] [CrossRef]
- Murase, Y.; Yabuta, Y.; Ohta, H.; Yamashiro, C.; Nakamura, T.; Yamamoto, T.; Saitou, M. Long-term expansion with germline potential of human primordial germ cell-like cells in vitro. EMBO J. 2020, 39, e104929. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n | Mean DAPI+ Cells ± SD | Mean DDX4+ Cells ± SD (% Per DAPI+ ± SD) | Mean DDX4+/POU5F1+ Cells ± SD (% Per DAPI+ ± SD) | Mean POU5F1+ Cells ± SD (% Per DAPI+ ± SD) | Mean SOMA Cells ± SD (% Per DAPI+ ± SD) | |
|---|---|---|---|---|---|---|
| Shinohara-Medium, with EGF, FGF2, GDNF, LIF, Estrogen and Progesterone | ||||||
| D3 | ||||||
| Gelatin | 3 | 792.0 ± 388.8 | 18.3 ± 7.6 (2.3%± 0.7) | 20.0 ± 13.0 (2.5% ± 1.0) | 4.3 ± 3.8 (0.6% ± 0.9) | 749.3 ± 372.1 (94.6% ± 0.8) |
| Laminin | 3 | 1259.3 ± 915.3 | 23.7 ± 13.7 (1.9% ± 1.4) | 42.3 ± 29.7 (3.4% ± 4.0) | 27.3 ± 42.3 (2.2% ± 5.5) | 1166.0 ± 920.8 (92.6%±11.0) |
| Matrigel | 3 | 2252.3 ± 356.6 | 83.3 ± 50.5 (3.7% ± 3.1) | 54.3 ± 15.0 (2.4% ± 1.1) | 17.0 ± 19.3 (0.8% ± 1.1) | 2097.7 ± 437.9 (93.1% ± 5.3) |
| Vitronectin | 3 | 1092.7 ± 827.7 | 37.7 ± 30.0 (3.4% ± 2.2) | 24.7 ± 28.1 (2.3% ± 1.1) | 6.0 ± 7.9 (0.6% ± 0.4) | 1024.3 ± 766.5 (93.8% ± 1.8) |
| D6 | ||||||
| Gelatin | 3 | 2028.3 ± 767.9 | 82.7 ± 53.8 (4.1% ± 1.1) | 9.,7 ± 11.7 (4.6% ± 2.2) | 19.3 ± 15.9 (1.0% ± 0.7) | 1832.7 ± 709.2 (90.4% ± 0.8) |
| Laminin | 3 | 2931.0 ± 1973.9 | 50.7 ± 43.6 (1.7% ± 2.2) | 43.7 ± 38.4 (1.5% ± 1.9) | 15.0 ± 10.5 (0.5% ± 0.6) | 2821.7 ± 1994.9 (96.3%±4.6) |
| Matrigel | 3 | 1405.0 ± 694.3 | 36.0 ± 28.1 (2.6% ± 1.1) | 36.0 ± 35.1 (2.6% ± 2.7) | 10.3 ± 12.3 (0.7% ± 0.6) | 1322.7 ± 650.2 (94.1% ± 3.4) |
| Vitronectin | 3 | 2331.0 ± 1061.9 | 76.7 ± 92.2 (3.3% ± 2.2) | 20.7 ± 15.9 (0.9% ± 0.9) | 14.0 ± 14.8 (0.6% ± 0.3) | 2219.7 ± 960.4 (95.2% ± 2.1) |
| Zhou-Medium (Adapted), with RA, BMP4 and ActA | ||||||
| D3 | ||||||
| Gelatin | 2 | 1060.5 ± 548.0 | 17.5 ± 6.4 (1.7% ± 1.7) | 6.5 ± 7.8 (0.6% ± 0.5) | 34.5 ± 43.1 (3.3% ± 6.6) | 1002.0 ± 589.7 (94.5% ± 7.8) |
| Laminin | 2 | 928.5 ± 593.3 | 27.0 ± 9.9 (2.9% ± 1.0) | 5.5 ± 2.1 (0.6% ± 0.8) | 0.0 ± 0.0 (0.0% ± 0.0) | 896.0 ± 585.5 (96.5% ± 1.8) |
| Matrigel | 2 | 1148.5 ± 7.8 | 46.5 ± 2.,5 (4.1% ± 1.8) | 16.0 ± 11.3 (1.4% ± 1.0) | 2.5 ± 2.1 (0.2% ± 0.2) | 1083.5 ± 41.7 (94.3% ± 3.0) |
| Vitronectin | 2 | 1387.0 ± 56.6 | 31.0 ± 8.5 (2.2% ± 0.5) | 8.5 ± 6.4 (0.6% ± 0.4) | 2.0 ± 2.8 (0.1% ± 0.2) | 1345.5 ± 38.9 (97.0% ± 1.2) |
| D6 | ||||||
| Gelatin | 2 | 853.0 ± 540.2 | 5.0 ± 2.8 (0.6% ± 0.1) | 2.5 ± 2.1 (0.3% ± 0.1) | 1.0 ± 0.0 (0.1% ± 0.1) | 844.5 ± 535.3 (99.0% ± 0.1) |
| Laminin | 2 | 1833.5 ± 1826.5 | 18.5 ± 20.5 (1.0% ± 0.2) | 6.0 ± 0.0 (0.3% ± 0.7) | 1.0 ± 1.4 (0.1% ± 0.0) | 1808.0 ± 1804.5 (98.6%±0.4) |
| Matrigel | 2 | 1955.5 ± 170.4 | 39.0 ± 39.6 (2.0% ± 1.9) | 6.0 ± 5.7 (0.3% ± 0.3) | 1.5 ± 2.1 (0.1% ± 0.1) | 1909.0 ± 123.0 (97.6% ± 2.2) |
| Vitronectin | 2 | 2819.0 ± 582.7 | 61.0 ± 33.9 (2.2% ± 0.8) | 26.0 ± 2.8 (0.9% ± 0.3) | 6.0 ± 7.1 (0.2% ± 0.2) | 2726.0 ± 544.5 (96.7% ± 0.7) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin-Inaraja, M.; Ferreira, M.; Taelman, J.; Eguizabal, C.; Chuva De Sousa Lopes, S.M. Improving In Vitro Culture of Human Male Fetal Germ Cells. Cells 2021, 10, 2033. https://doi.org/10.3390/cells10082033
Martin-Inaraja M, Ferreira M, Taelman J, Eguizabal C, Chuva De Sousa Lopes SM. Improving In Vitro Culture of Human Male Fetal Germ Cells. Cells. 2021; 10(8):2033. https://doi.org/10.3390/cells10082033
Chicago/Turabian StyleMartin-Inaraja, Myriam, Monica Ferreira, Jasin Taelman, Cristina Eguizabal, and Susana M. Chuva De Sousa Lopes. 2021. "Improving In Vitro Culture of Human Male Fetal Germ Cells" Cells 10, no. 8: 2033. https://doi.org/10.3390/cells10082033
APA StyleMartin-Inaraja, M., Ferreira, M., Taelman, J., Eguizabal, C., & Chuva De Sousa Lopes, S. M. (2021). Improving In Vitro Culture of Human Male Fetal Germ Cells. Cells, 10(8), 2033. https://doi.org/10.3390/cells10082033