
Estradiol Inhibits Human Brain Vascular Pericyte Migration Activity: A Functional and Transcriptomic Analysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Count
2.3. Migration Studies
2.4. Western Blot Analysis
2.5. Transendothelial Electric Resistance (TEER)
2.6. Microarray
2.7. Statistical Analysis
3. Results
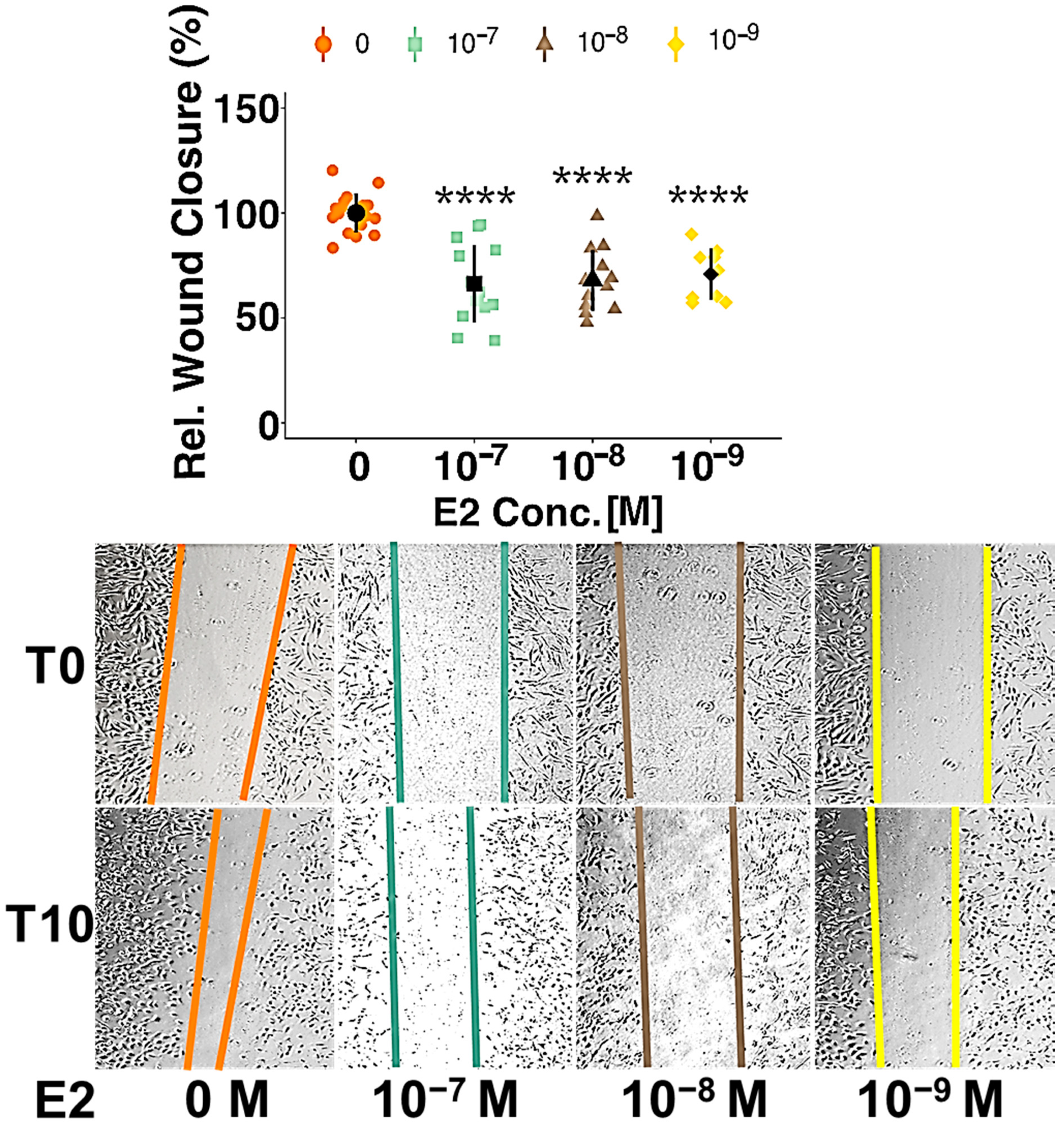
3.1. Estradiol Inhibits Migration of Pericytes but Has No Impact on Proliferation
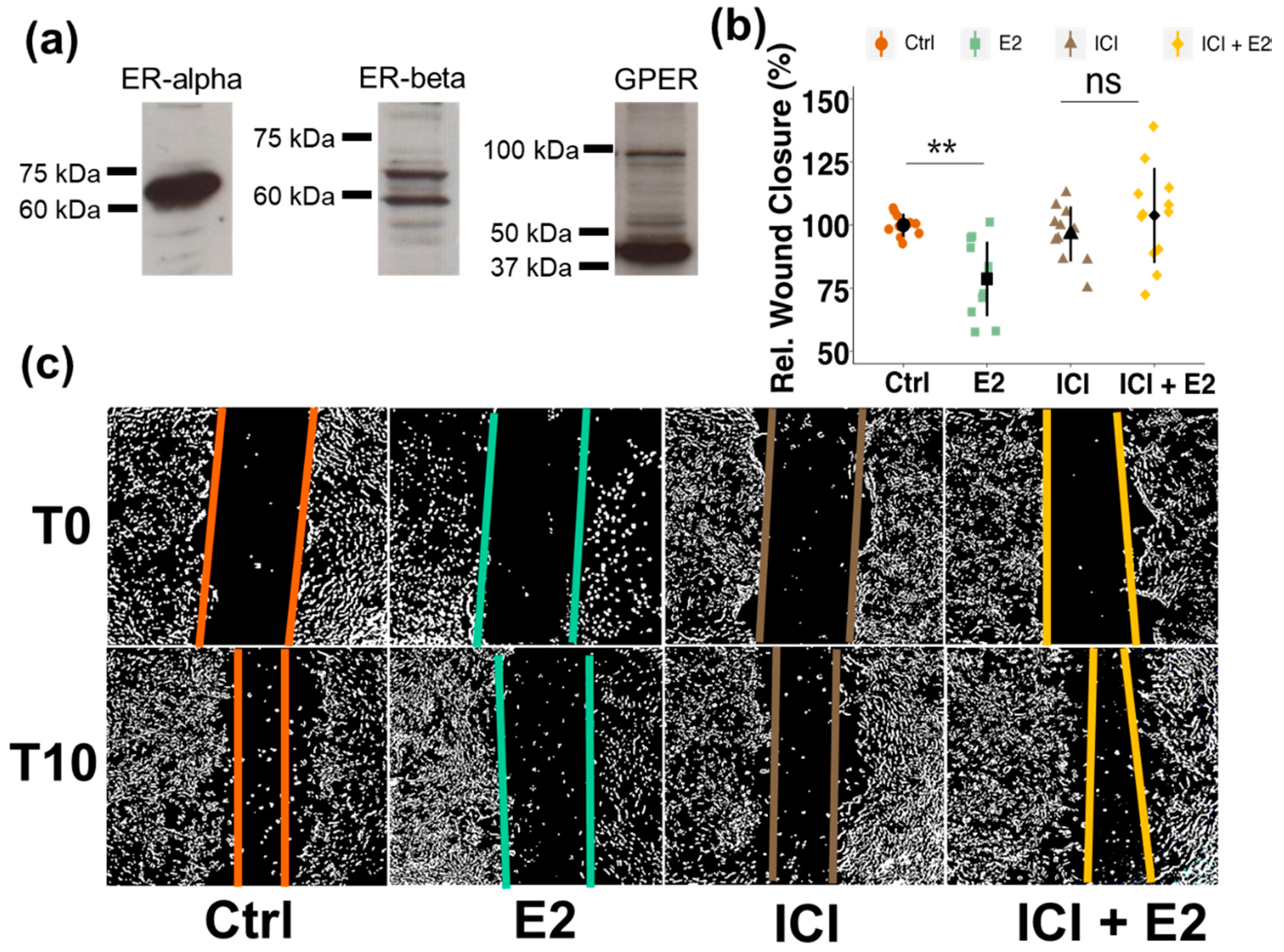
3.2. Brain Pericytes Express All Three Estrogen Receptors
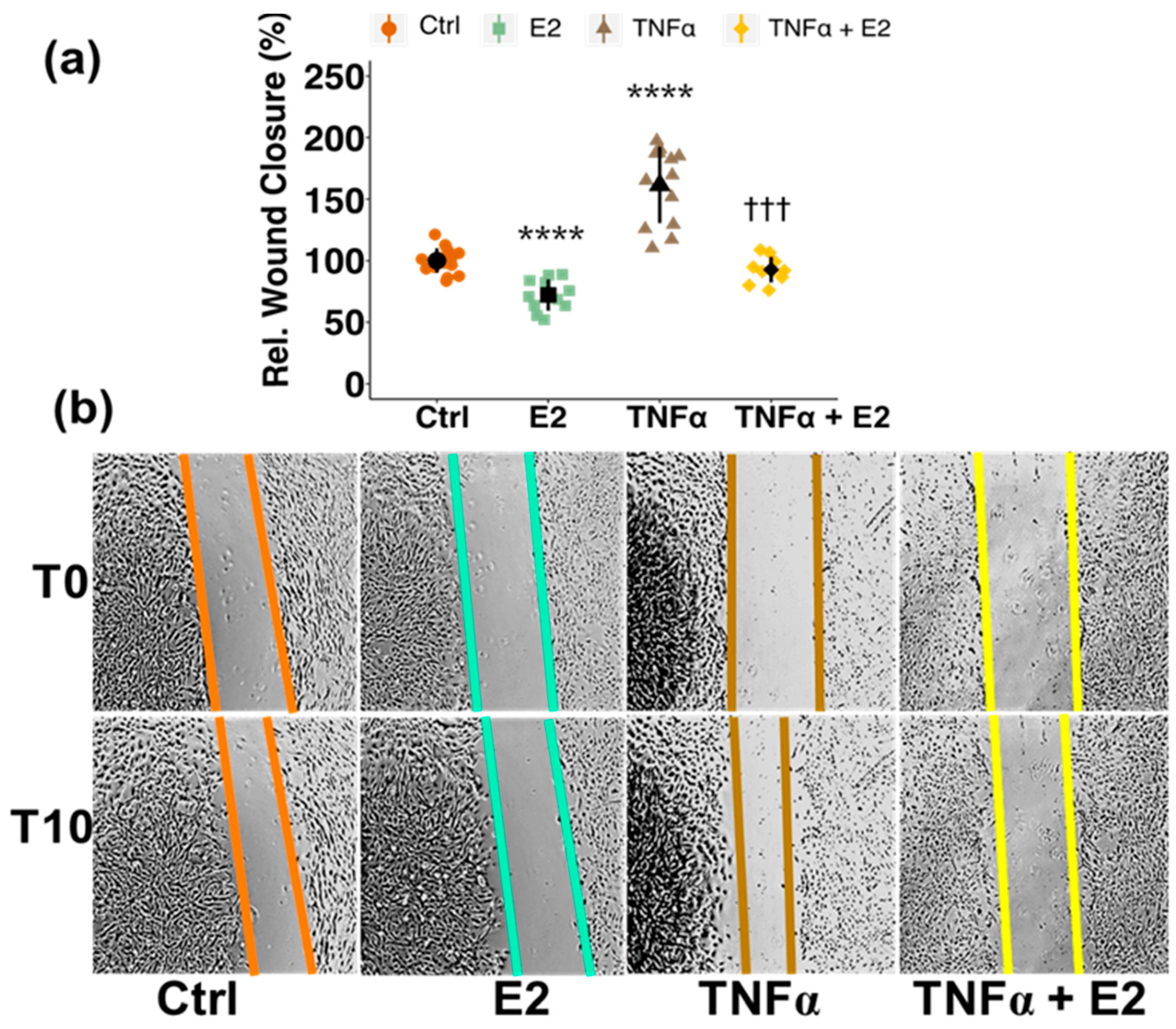
3.3. Estradiol Attenuates Tumor Necrosis Factor Alpha (TNFα)-Induced Migration of PCs
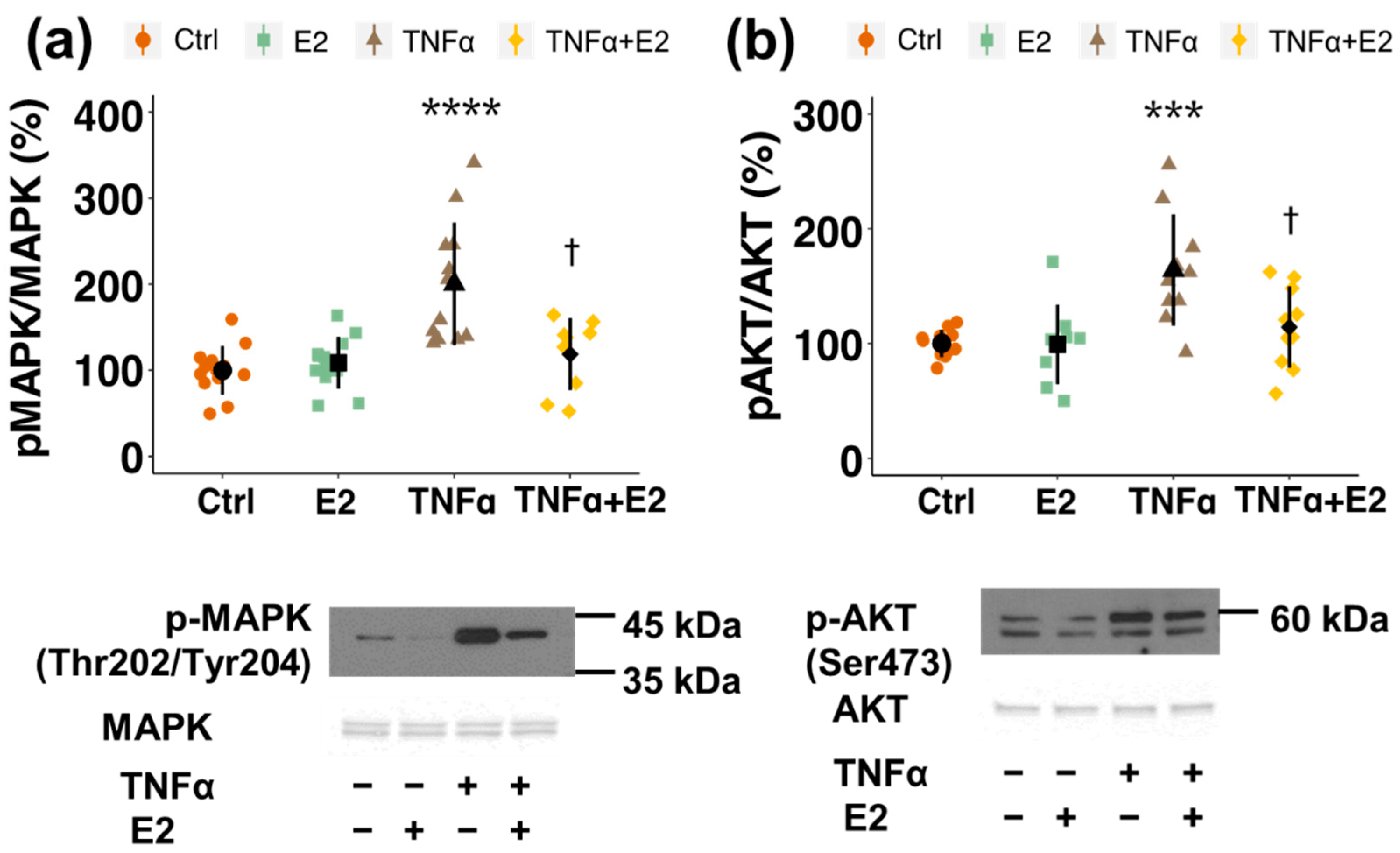
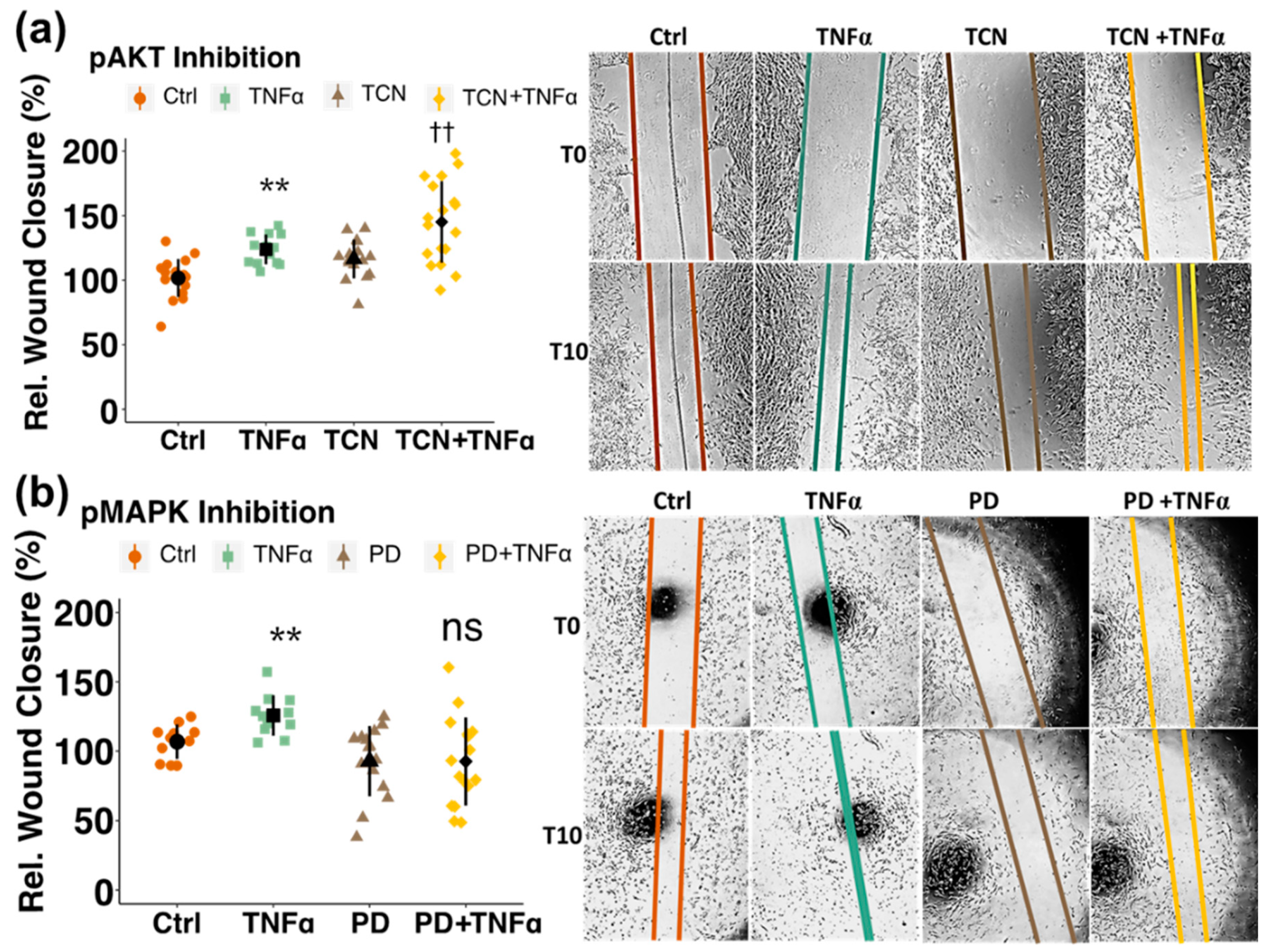
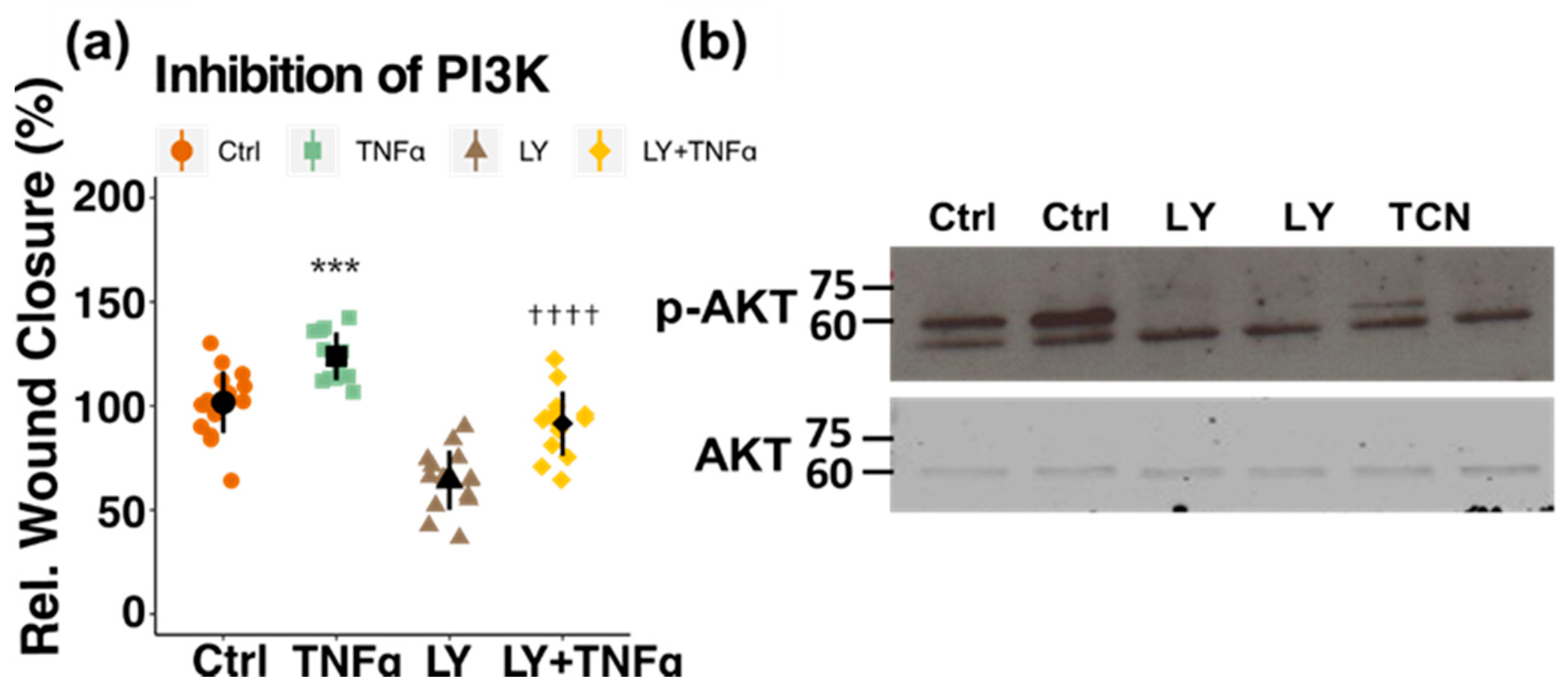
3.4. Involvement of Kinase Cascades
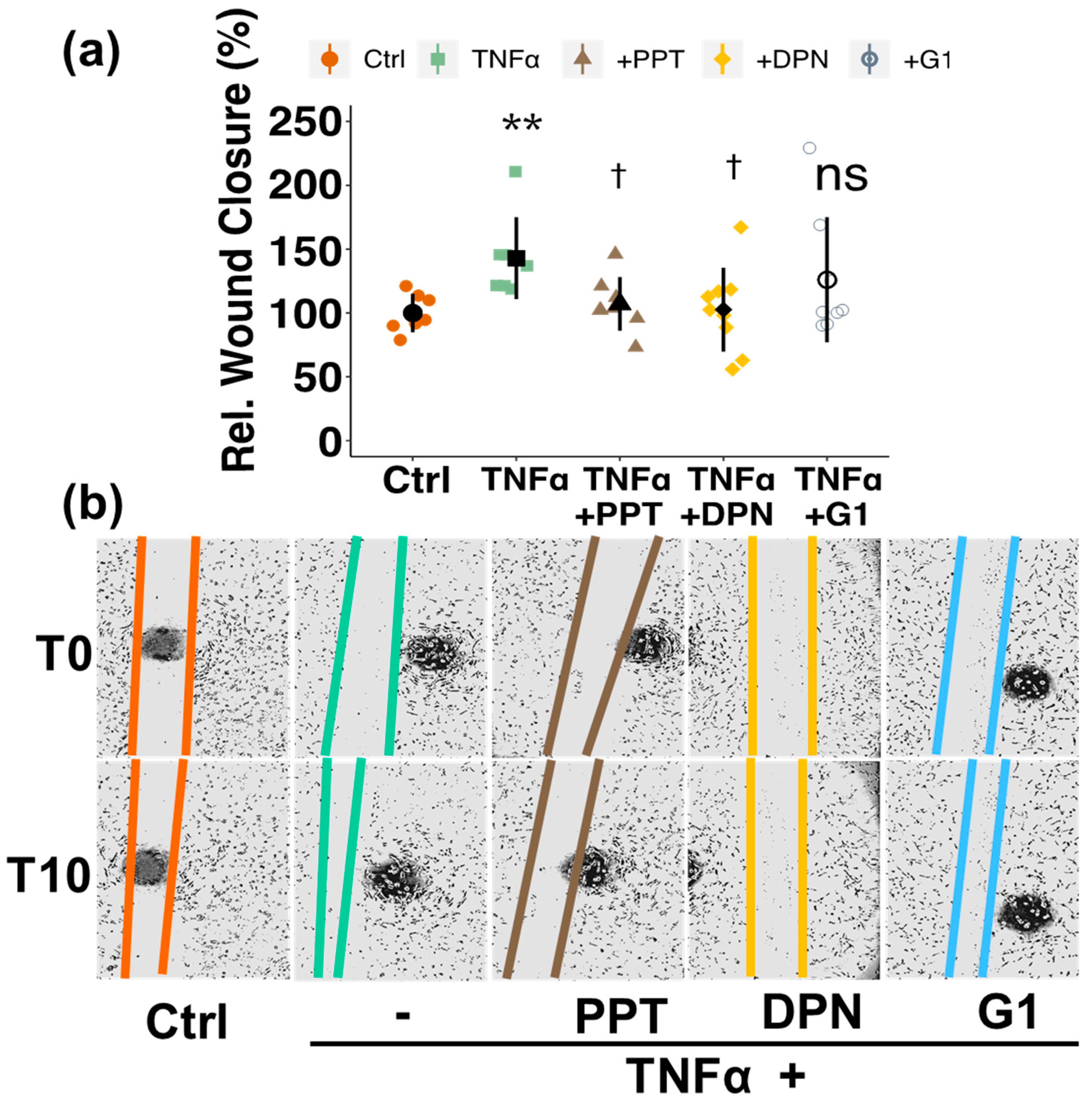
3.5. Involvement of Estrogen Receptors (ERs)
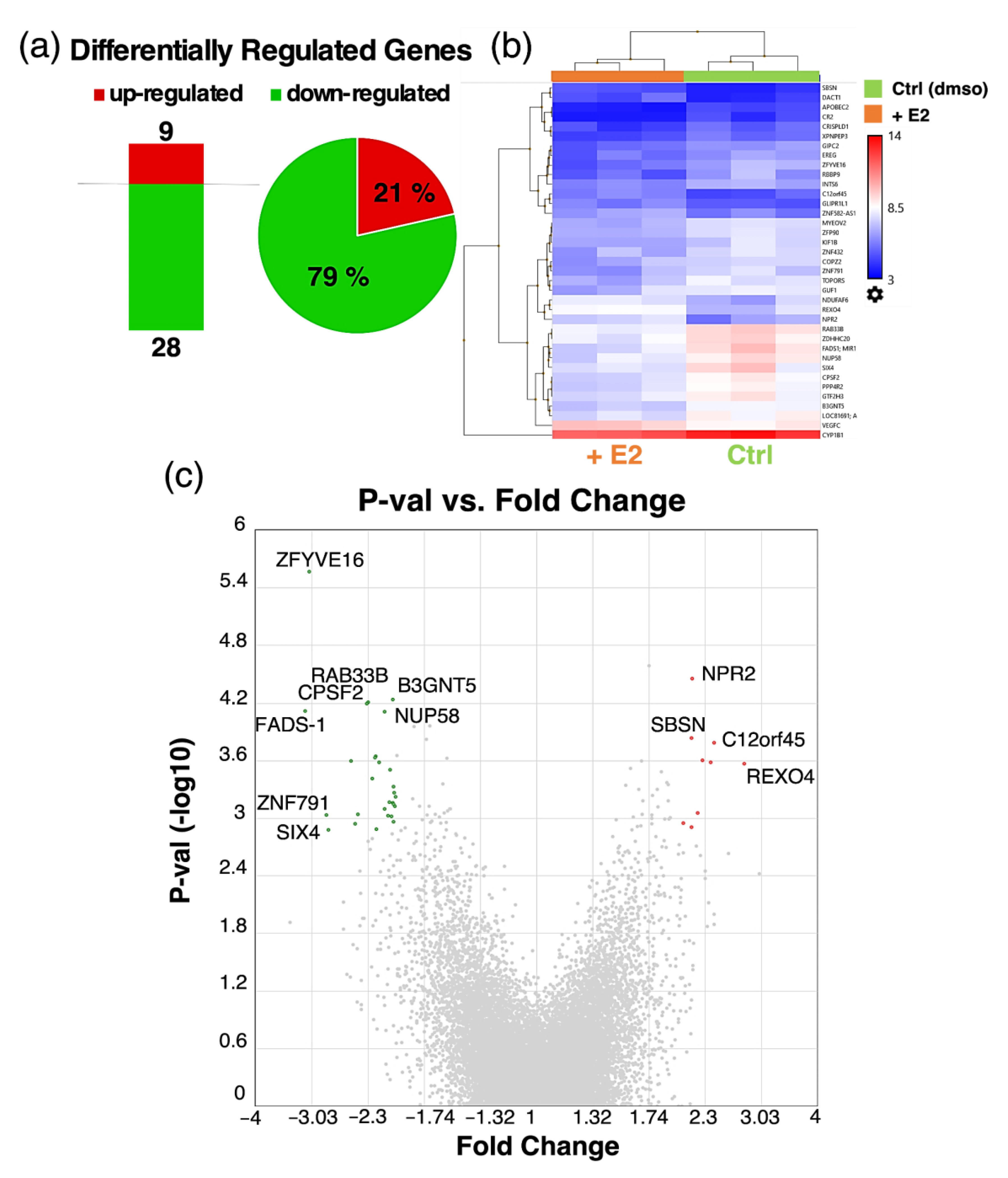
3.6. Microarray Analysis of Estradiol Treated Pericytes
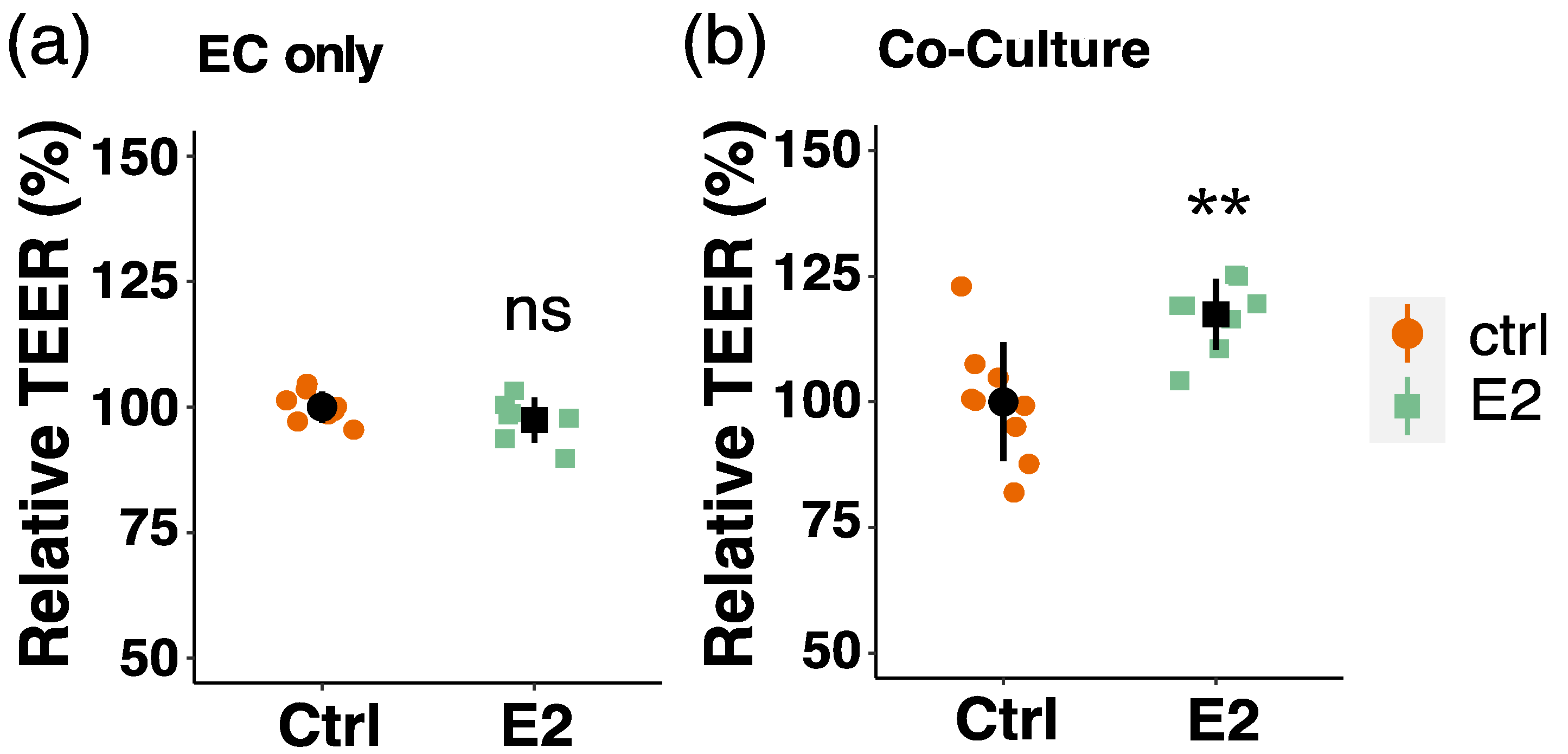
3.7. Effect of Estradiol on Endothelial Barrier Function
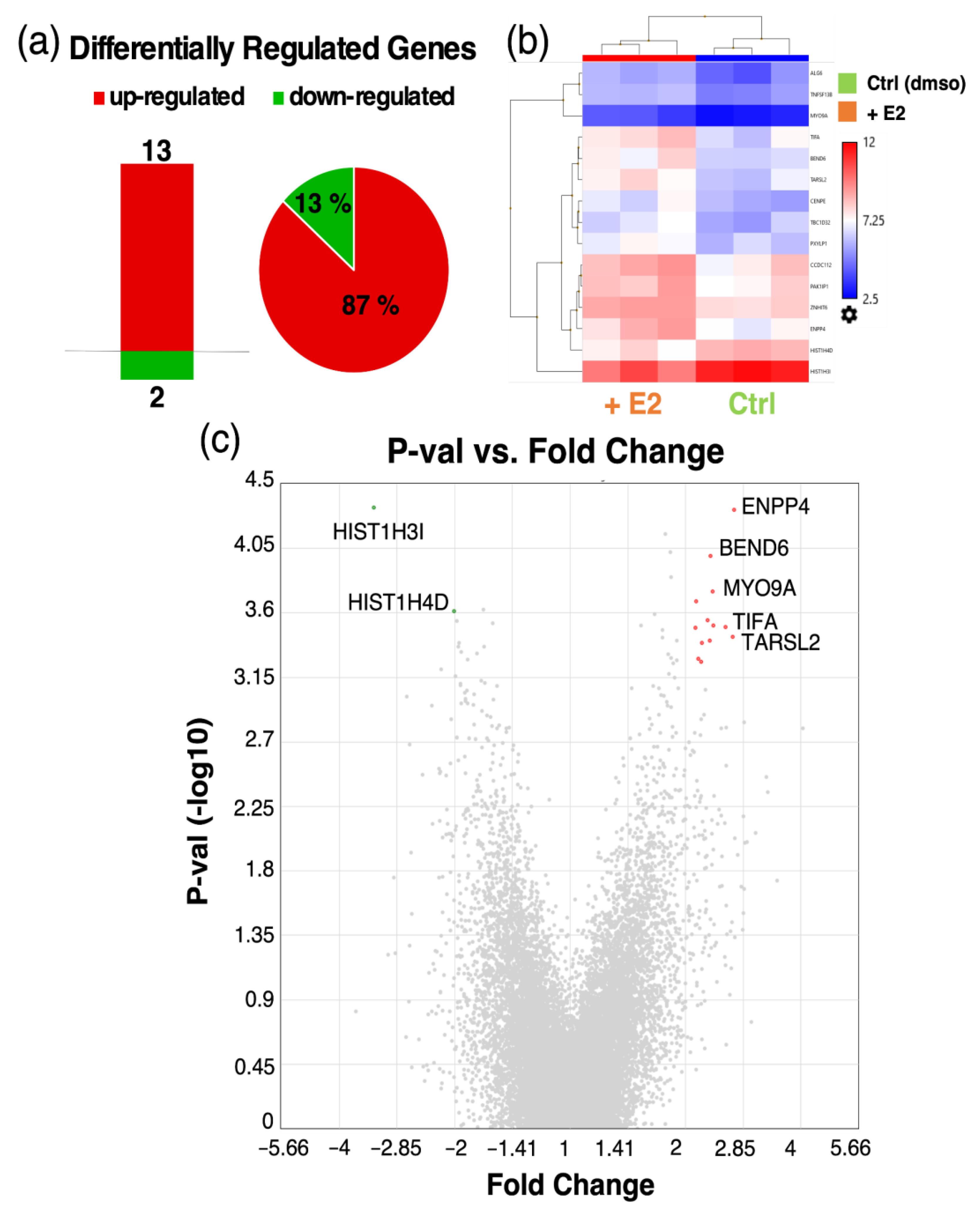
3.8. Microarray Analysis of Co-Cultured Pericytes Treated with Estradiol
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Demel, S.L.; Kittner, S.; Ley, S.H.; McDermott, M.; Rexrode, K.M. Stroke Risk Factors Unique to Women. Stroke 2018, 49, 518–523. [Google Scholar] [CrossRef] [Green Version]
- Koellhoffer, E.C.; McCullough, L.D. The Effects of Estrogen in Ischemic Stroke. Transl. Stroke Res. 2013, 4, 390–401. [Google Scholar] [CrossRef] [Green Version]
- Iorga, A.; Cunningham, C.M.; Moazeni, S.; Ruffenach, G.; Umar, S.; Eghbali, M. The protective role of estrogen and estrogen receptors in cardiovascular disease and the controversial use of estrogen therapy. Biol. Sex Differ. 2017, 8, 33. [Google Scholar] [CrossRef]
- Xing, D.; Nozell, S.; Chen, Y.-F.; Hage, F.; Oparil, S. Estrogen and Mechanisms of Vascular Protection. Arter. Thromb. Vasc. Biol. 2009, 29, 289–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadareishvili, Z.; Simpkins, A.N.; Hitomi, E.; Reyes, D.; Leigh, R. Post-Stroke Blood-Brain Barrier Disruption and Poor Functional Outcome in Patients Receiving Thrombolytic Therapy. Cerebrovasc. Dis. 2019, 47, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Yang, S.; Jeong, S.; Park, H.; Choi, Y.-H.; Park, E.-M. Activation of estrogen receptor β reduces blood–brain barrier breakdown following ischemic injury. Neuroscience 2013, 235, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Burek, M.; Arias-Loza, P.A.; Roewer, N.; Forster, C.Y. Claudin-5 as a Novel Estrogen Target in Vascular Endothelium. Arter. Thromb. Vasc. Biol. 2010, 30, 298–304. [Google Scholar] [CrossRef] [Green Version]
- Nadkarni, S.; McArthur, S. Oestrogen and immunomodulation: New mechanisms that impact on peripheral and central immunity. Curr. Opin. Pharmacol. 2013, 13, 576–581. [Google Scholar] [CrossRef]
- Czlonkowska, A.; Ciesielska, A.; Gromadzka, G.; Kurkowska-Jastrzebska, I. Estrogen and Cytokines Production - The Possible Cause of Gender Differences in Neurological Diseases. Curr. Pharm. Des. 2005, 11, 1017–1030. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, Z.; Sun, W.; Koehler, R.C.; Huang, J. 17β-estradiol attenuates breakdown of blood–brain barrier and hemorrhagic transformation induced by tissue plasminogen activator in cerebral ischemia. Neurobiol. Dis. 2011, 44, 277–283. [Google Scholar] [CrossRef] [Green Version]
- McCullough, L.D.; Hurn, P.D. Estrogen and ischemic neuroprotection: An integrated view. Trends Endocrinol. Metab. 2003, 14, 228–235. [Google Scholar] [CrossRef]
- Gorodeski, G.I. Estrogen Decrease in Tight Junctional Resistance Involves Matrix-Metalloproteinase-7-Mediated Remodeling of Occludin. Endocrinology 2007, 148, 218–231. [Google Scholar] [CrossRef] [Green Version]
- Groten, T.; Pierce, A.A.; Huen, A.C.; Schnaper, H.W. 17 β-estradiol transiently disrupts adherens junctions in endothelial cells. FASEB J. 2005, 19, 1368–1370. [Google Scholar] [CrossRef]
- Straub, R.H. The Complex Role of Estrogens in Inflammation. Endocr. Rev. 2007, 28, 521–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuruca, S.E.; Karadenizli, S.; Akgun-Dar, K.; Kapucu, A.; Kaptan, Z.; Uzum, G. The effects of 17β-estradiol on blood brain barrier integrity in the absence of the estrogen receptor alpha; an in-vitro model. Acta Histochem. 2017, 119, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Gastfriend, B.D.; Palecek, S.P.; Shusta, E.V. Modeling the blood–brain barrier: Beyond the endothelial cells. Curr. Opin. Biomed. Eng. 2018, 5, 6–12. [Google Scholar] [CrossRef]
- Armulik, A.; Genové, G.; Mäe, M.; Nisancioglu, M.H.; Wallgard, E.; Niaudet, C.; He, L.; Norlin, J.; Lindblom, P.; Strittmatter, K.; et al. Pericytes regulate the blood-brain barrier. Nature 2010, 468, 557–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daneman, R.; Zhou, L.; Kebede, A.A.; Barres, B.A. Pericytes are required for blood–brain barrier integrity during embryogenesis. Nat. Cell Biol. 2010, 468, 562–566. [Google Scholar] [CrossRef] [Green Version]
- Zlokovic, B.V. The Blood-Brain Barrier in Health and Chronic Neurodegenerative Disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michinaga, S.; Koyama, Y. Dual Roles of Astrocyte-Derived Factors in Regulation of Blood-Brain Barrier Function after Brain Damage. Int. J. Mol. Sci. 2019, 20, 571. [Google Scholar] [CrossRef] [Green Version]
- Armulik, A.; Genové, G.; Betsholtz, C. Pericytes: Developmental, Physiological, and Pathological Perspectives, Problems, and Promises. Dev. Cell 2011, 21, 193–215. [Google Scholar] [CrossRef] [Green Version]
- Trost, A.; Lange, S.; Schroedl, F.; Bruckner, D.; Motloch, K.A.; Bogner, B.; Kaser-Eichberger, A.; Strohmaier, C.; Runge, C.; Aigner, L.; et al. Brain and Retinal Pericytes: Origin, Function and Role. Front. Cell. Neurosci. 2016, 10, 20. [Google Scholar] [CrossRef] [Green Version]
- Lendahl, U.; Nilsson, P.; Betsholtz, C. Emerging links between cerebrovascular and neurodegenerative diseases—a special role for pericytes. EMBO Rep. 2019, 20, e48070. [Google Scholar] [CrossRef]
- Hall, C.; Reynell, C.; Gesslein, B.; Hamilton-Whitaker, N.; Mishra, A.; Sutherland, B.; O’Farrell, F.; Buchan, A.M.; Lauritzen, M.; Attwell, D. Capillary pericytes regulate cerebral blood flow in health and disease. Nat. Cell Biol. 2014, 508, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, M.D.; Sagare, A.P.; Zlokovic, B.V. Blood–brain barrier breakdown in Alzheimer disease and other neurodegenerative disorders. Nat. Rev. Neurol. 2018, 14, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Laredo, F.; Plebanski, J.; Tedeschi, A. Pericytes: Problems and Promises for CNS Repair. Front. Cell. Neurosci. 2019, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Ferland-McCollough, D.; Slater, S.; Richard, J.; Reni, C.; Mangialardi, G. Pericytes, an overlooked player in vascular pathobiology. Pharmacol. Ther. 2017, 171, 30–42. [Google Scholar] [CrossRef]
- Nishioku, T.; Dohgu, S.; Takata, F.; Eto, T.; Ishikawa, N.; Kodama, K.; Nakagawa, S.; Yamauchi, A.; Kataoka, Y. Detachment of Brain Pericytes from the Basal Lamina is Involved in Disruption of the Blood–Brain Barrier Caused by Lipopolysaccharide-Induced Sepsis in Mice. Cell. Mol. Neurobiol. 2008, 29, 309–316. [Google Scholar] [CrossRef]
- Eshaq, R.S.; Aldalati, A.M.; Alexander, J.S.; Harris, N.R. Diabetic retinopathy: Breaking the barrier. Pathophysiology 2017, 24, 229–241. [Google Scholar] [CrossRef]
- Zehendner, C.M.; Sebastiani, A.; Hugonnet, A.; Bischoff, F.; Luhmann, H.J.; Thal, S.C. Traumatic brain injury results in rapid pericyte loss followed by reactive pericytosis in the cerebral cortex. Sci. Rep. 2015, 5, 13497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geranmayeh, M.H.; Rahbarghazi, R.; Farhoudi, M. Targeting pericytes for neurovascular regeneration. Cell Commun. Signal. 2019, 17, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Tigges, U.; Boroujerdi, A.; Welser-Alves, J.V.; Milner, R. TNF-α promotes cerebral pericyte remodeling in vitro, via a switch from α1 to α2 integrins. J. Neuroinflamm. 2013, 10, 33. [Google Scholar] [CrossRef] [Green Version]
- Takata, F.; Dohgu, S.; Matsumoto, J.; Takahashi, H.; Machida, T.; Wakigawa, T.; Harada, E.; Miyaji, H.; Koga, M.; Nishioku, T.; et al. Brain pericytes among cells constituting the blood-brain barrier are highly sensitive to tumor necrosis factor-α, releasing matrix metalloproteinase-9 and migrating in vitro. J. Neuroinflamm. 2011, 8, 106. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Kastin, A.J. Tumor necrosis factor and stroke: Role of the blood–brain barrier. Prog. Neurobiol. 2007, 83, 363–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clausen, B.H.; Wirenfeldt, M.; Høgedal, S.S.; Frich, L.H.; Nielsen, H.H.; Schrøder, H.D.; Østergaard, K.; Finsen, B.; Kristensen, B.W.; Lambertsen, K.L. Characterization of the TNF and IL-1 systems in human brain and blood after ischemic stroke. Acta Neuropathol. Commun. 2020, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mendelsohn, M.E.; Karas, R.H. The Protective Effects of Estrogen on the Cardiovascular System. N. Engl. J. Med. 1999, 340, 1801–1811. [Google Scholar] [CrossRef]
- Gerrits, P.O.; de Weerd, H.; van der Want, J.; Kortekaas, R.; Luiten, P.G.; Veening, J.G. Microvascular changes in estrogen-α sensitive brainstem structures of aging female hamsters. Neurosci. Res. 2010, 67, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurmann, L.; Okoniewski, M.; Dubey, R. Transcryptomic Analysis of Human Brain -Microvascular Endothelial Cell Driven Changes in -Vascular Pericytes. Cells 2021, 10, 1784. [Google Scholar] [CrossRef]
- Weksler, B.B.; Subileau, E.A.; Perrière, N.; Charneau, P.; Holloway, C.J.; Leveque, M.; Tricoire-Leignel, H.; Nicotra, A.; Bourdoulous, S.; Turowski, P.; et al. Blood-brain barrier-specific properties of a human adult brain endothelial cell line. FASEB J. 2005, 19, 1872–1874. [Google Scholar] [CrossRef]
- Kurmann, L.; Okoniewski, M.; Ogunshola, O.O.; Leeners, B.; Imthurn, B.; Dubey, R.K. Transcryptomic Analysis of Human Brain-Microvascular Endothelial Response to -Pericytes: Cell Orientation Defines Barrier Function. Cells 2021, 10, 963. [Google Scholar] [CrossRef]
- Melgar, M.A.; Rafols, J.; Gloss, D.; Diaz, F.G. Postischemic Reperfusion: Ultrastructural Blood-Brain Barrier and Hemodynamic Correlative Changes in an Awake Model of Transient Forebrain Ischemia. Neurosurgery 2005, 56, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Dore-Duffy, P.; Owen, C.; Balabanov, R.; Murphy, S.; Beaumont, T.; Rafols, J.A. Pericyte Migration from the Vascular Wall in Response to Traumatic Brain Injury. Microvasc. Res. 2000, 60, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.L.; Merchant, R.E. Blood-brain barrier changes following intracerebral injection of human recombinant tumor necrosis factor-? in the rat. J. Neuro-Oncol. 1994, 20, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Tian, L.; Zhao, B.; Sun, Y.; Cao, J.; Chen, K.; Li, F.; Li, M.; Shang, D.; Liu, M. FADS1 promotes the progression of laryngeal squamous cell carcinoma through activating AKT/mTOR signaling. Cell Death Dis. 2020, 11, 272. [Google Scholar] [CrossRef]
- Xia, X.; Li, Y.; Wang, W.; Tang, F.; Tan, J.; Sun, L.; Li, Q.; Sun, L.; Tang, B.; He, S. MicroRNA-1908 functions as a glioblastoma oncogene by suppressing PTEN tumor suppressor pathway. Mol. Cancer 2015, 14, 154. [Google Scholar] [CrossRef] [Green Version]
- Paek, A.R.; You, H.J.; Lee, C.-H. A role of zinc-finger protein 143 for cancer cell migration and invasion through ZEB1 and E-cadherin in colon cancer cells. Mol. Carcinog. 2013, 53, E161–E168. [Google Scholar] [CrossRef]
- Indukuri, R.; Jafferali, M.H.; Song, D.; Damdimopoulos, A.; Hases, L.; Zhao, C.; Archer, A.; Williams, C. Genome-wide estrogen receptor β chromatin binding in human colon cancer cells reveals its tumor suppressor activity. Int. J. Cancer 2021, 149, 692–706. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Ma, J.; Chen, Q.; Hou, Z.; Luo, X.; Wang, G.; Wang, J.; Hu, J.; Cao, Z. SIX4 promotes metastasis through STAT3 activation in breast cancer. Am. J. Cancer Res. 2020, 10, 224–236. [Google Scholar] [PubMed]
- Shields, D.J.; Niessen, S.; Murphy, E.A.; Mielgo, A.; Desgrosellier, J.S.; Lau, S.K.M.; Barnes, L.A.; Lesperance, J.; Bouvet, M.; Tarin, D.; et al. RBBP9: A tumor-associated serine hydrolase activity required for pancreatic neoplasia. Proc. Natl. Acad. Sci. USA 2010, 107, 2189–2194. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-L.; Zhu, Z.-X.; Yang, X.; Liu, L.-L.; He, Y.-F.; Yang, M.-M.; Guan, X.-Y.; Wang, X.; Yun, J.-P. Cleavage and Polyadenylation Specific Factor 1 Promotes Tumor Progression via Alternative Polyadenylation and Splicing in Hepatocellular Carcinoma. Front. Cell Dev. Biol. 2021, 9, 616835. [Google Scholar] [CrossRef]
- Mansour, M.A. SP3 is associated with migration, invasion, and Akt/PKB signalling in MDA-MB-231 breast cancer cells. J. Biochem. Mol. Toxicol. 2021, 35, e22657. [Google Scholar] [CrossRef]
- Clayton, N.S.; Ridley, A.J. Targeting Rho GTPase Signaling Networks in Cancer. Front. Cell Dev. Biol. 2020, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Huo, X.; Zhang, Y.; Yan, Z.; Wang, Q.; Wen, B. TOPORS, a tumor suppressor protein, contributes to the maintenance of higher-order chromatin architecture. Biochim. Biophys. Acta (BBA) Bioenerg. 2020, 1863, 194518. [Google Scholar] [CrossRef]
- Fu, Y.; Katsaros, D.; Biglia, N.; Wang, Z.; Pagano, I.; Tius, M.; Tiirikainen, M.; Rosser, C.; Yang, H.; Yu, H. Vitamin D receptor upregulates lncRNA TOPORS-AS1 which inhibits the Wnt/β-catenin pathway and associates with favorable prognosis of ovarian cancer. Sci. Rep. 2021, 11, 7484. [Google Scholar] [CrossRef]
- Shi, J.; Li, C.; Wang, H.; Xiao, B.; Qiu, W. NUP58 facilitates metastasis and epithelial-mesenchymal transition of lung adenocarcinoma via the GSK-3 beta/Snail signaling pathway. Am. J. Transl. Res. 2019, 11, 393–405. [Google Scholar]
- Li, X.; Liang, L.; Huang, L.; Ma, X.; Li, D.; Cai, S. High expression of protein phosphatase 4 is associated with the aggressive malignant behavior of colorectal carcinoma. Mol. Cancer 2015, 14, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Wang, L.; Yang, H.; Lin, X.; Li, G.; Han, N.; Du, J.; Fan, Z. Epiregulin promotes the migration and chemotaxis ability of adipose-derived mesenchymal stem cells via mitogen-activated protein kinase signaling pathways. J. Cell. Biochem. 2018, 119, 8450–8459. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Hu, X.; Zhao, X.; Kong, X.; Meng, Y.-M.; Chen, Y.; Su, L.; Jiang, X.; Qiu, X.; Huang, C.; et al. A circular network of coregulated sphingolipids dictates lung cancer growth and progression. EBioMedicine 2021, 66, 103301. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, N.; Ngam, C.R.; Berdis, A.J.; Montano, M.M. The exonuclease activity of hPMC2 is required for transcriptional regulation of the QR gene and repair of estrogen-induced abasic sites. Oncogene 2011, 30, 4731–4739. [Google Scholar] [CrossRef] [Green Version]
- Thapa, D.; Huang, S.-B.; Muñoz, A.R.; Yang, X.; Bedolla, R.G.; Hung, C.-N.; Chen, C.-L.; Huang, T.H.-M.; Liss, M.A.; Reddick, R.L.; et al. Attenuation of NAD[P]H:quinone oxidoreductase 1 aggravates prostate cancer and tumor cell plasticity through enhanced TGFβ signaling. Commun. Biol. 2020, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Urra, F.A.; Muñoz, F.; Lovy, A.; Cárdenas, C. The Mitochondrial Complex(I)ty of Cancer. Front. Oncol. 2017, 7, 118. [Google Scholar] [CrossRef]
- Yang, W.; Zhang, K.; Li, L.; Xu, Y.; Ma, K.; Xie, H.; Zhou, J.; Cai, L.; Gong, Y.; Gong, K. Correction to: Downregulation of lncRNA ZNF582-AS1 due to DNA hypermethylation promotes clear cell renal cell carcinoma growth and metastasis by regulating the N(6)-methyladenosine modification of MT-RNR1. J. Exp. Clin. Cancer Res. 2021, 40, 92. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-H.; Lin, L.-K.; Zhang, S. Effects of DACT1 methylation status on invasion and metastasis of nasopharyngeal carcinoma. Biol. Res. 2019, 52, 31. [Google Scholar] [CrossRef]
- Ren, C.; Ren, C.-H.; Li, L.; Goltsov, A.A.; Thompson, T.C. Identification and characterization of RTVP1/GLIPR1-like genes, a novel p53 target gene cluster. Genome 2006, 88, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Nakao, S.; Nakaoke, R.; Nakagawa, S.; Kitagawa, N.; Niwa, M. Effects of hypoxia on endothelial/pericytic co-culture model of the blood–brain barrier. Regul. Pept. 2004, 123, 77–83. [Google Scholar] [CrossRef]
- Hori, S.; Ohtsuki, S.; Hosoya, K.-I.; Nakashima, E.; Terasaki, T. A pericyte-derived angiopoietin-1 multimeric complex induces occludin gene expression in brain capillary endothelial cells through Tie-2 activation in vitro. J. Neurochem. 2004, 89, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Stone, N.L.; England, T.J.; O’Sullivan, S.E. A Novel Transwell Blood Brain Barrier Model Using Primary Human Cells. Front. Cell. Neurosci. 2019, 13, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Jiang, K.; Zhi, T.; Xu, X. miR-720 is a key regulator of glioma migration and invasion by controlling TARSL2 expression. Hum. Cell 2021, 1–13. [Google Scholar] [CrossRef]
- Dai, Q.; Andreu-Agullo, C.; Insolera, R.; Wong, L.C.; Shi, S.-H.; Lai, E.C. BEND6 is a nuclear antagonist of Notch signaling during self-renewal of neural stem cells. Development 2013, 140, 1892–1902. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.W.; Nishiyama, N.C.; Byrum, S.D.; Stanbouly, S.; Jones, T.; Holley, J.; Sridharan, V.; Boerma, M.; Tackett, A.J.; Willey, J.S.; et al. Spaceflight induces oxidative damage to blood-brain barrier integrity in a mouse model. FASEB J. 2020, 34, 15516–15530. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, F.; Li, T.; Xiao, P.; Han, Z.-J.; Shu, L.-F.; Yuan, Z.-Z.; Liu, W.-J.; Long, Y.-Q. Extracellular Histone Promotes Prostate Cancer Migration and Epithelial-Mesenchymal Transition through NF-κB-Mediated Inflammatory Responses. Chemotherapy 2019, 64, 177–186. [Google Scholar] [CrossRef]
- Ilter, D.; Blenis, J.; Gomes, A.P. Histone H3 variants at the root of metastasis. Mol. Cell. Oncol. 2020, 7, 1684128. [Google Scholar] [CrossRef]
- Markouli, M.; Strepkos, D.; Basdra, E.K.; Papavassiliou, A.G.; Piperi, C. Prominent Role of Histone Modifications in the Regulation of Tumor Metastasis. Int. J. Mol. Sci. 2021, 22, 2778. [Google Scholar] [CrossRef]
- Jiang, X.; Andjelkovic, A.V.; Zhu, L.; Yang, T.; Bennett, M.V.L.; Chen, J.; Keep, R.F.; Shi, Y. Blood-brain barrier dysfunction and recovery after ischemic stroke. Prog. Neurobiol. 2018, 163–164, 144–171. [Google Scholar] [CrossRef] [PubMed]
- O’Dowd, B.F.; Nguyen, T.; Marchese, A.; Cheng, R.; Lynch, K.R.; Heng, H.H.; Kolakowski, L.F.; George, S.R. Discovery of Three Novel G-Protein-Coupled Receptor Genes. Genome 1998, 47, 310–313. [Google Scholar] [CrossRef]
- Almey, A.; Milner, T.A.; Brake, W.G. Estrogen receptors in the central nervous system and their implication for dopamine-dependent cognition in females. Horm. Behav. 2015, 74, 125–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Krause, D.N.; Horne, J.; Weiss, J.; Li, X.; Duckles, S.P. Estrogen-Receptor-Mediated Protection of Cerebral Endothelial Cell Viability and Mitochondrial Function after Ischemic Insult in vitro. Br. J. Pharmacol. 2009, 30, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Kuiper, G.G.; Shughrue, P.J.; Merchenthaler, I.; Gustafsson, J.A. The Estrogen Receptor β Subtype: A Novel Mediator of Estrogen Action in Neuroendocrine Systems. Front. Neuroendocr. 1998, 19, 253–286. [Google Scholar] [CrossRef]
- Watson, C.S.; Jeng, Y.; Kochukov, M. Nongenomic actions of estradiol compared with estrone and estriol in pituitary tumor cell signaling and proliferation. FASEB J. 2008, 22, 3328–3336. [Google Scholar] [CrossRef] [Green Version]
- Xiang, D.; Feng, Y.; Wang, J.; Zhang, X.; Shen, J.; Zou, R.; Yuan, Y. Platelet-derived growth factor-BB promotes proliferation and migration of retinal microvascular pericytes by up-regulating the expression of C-X-C chemokine receptor types 4. Exp. Ther. Med. 2019, 18, 4022–4030. [Google Scholar] [CrossRef] [Green Version]
- Milesi, S.; Boussadia, B.; Plaud, C.; Catteau, M.; Rousset, M.-C.; De Bock, F.; Schaeffer, M.; Lerner-Natoli, M.; Rigau, V.; Marchi, N. Redistribution of PDGFRβ cells and NG2DsRed pericytes at the cerebrovasculature after status epilepticus. Neurobiol. Dis. 2014, 71, 151–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Klett, F.; Potas, J.; Hilpert, D.; Blazej, K.; Radke, J.; Huck, J.; Engel, O.; Stenzel, W.; Genové, G.; Priller, J. Early Loss of Pericytes and Perivascular Stromal Cell-Induced Scar Formation after Stroke. Br. J. Pharmacol. 2012, 33, 428–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokhari, F.A.; Shakoori, T.; Butt, A.; Ghafoor, F. TNF-alpha: A risk factor for ischemic stroke. J. Ayub Med Coll. Abbottabad 2015, 26, 111–114. [Google Scholar]
- Wilkins, H.M.; Swerdlow, R.H. TNFα in cerebral ischemia: Another stroke against you? J. Neurochem. 2015, 132, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Shen, Y.; Li, R. Targeting TNF: A therapeutic strategy for Alzheimer’s disease. Drug Discov. Today 2014, 19, 1822–1827. [Google Scholar] [CrossRef]
- Smyth, L.C.D.; Rustenhoven, J.; Park, T.I.-H.; Schweder, P.; Jansson, D.; Heppner, P.A.; O’Carroll, S.J.; Mee, E.W.; Faull, R.L.M.; Curtis, M.; et al. Unique and shared inflammatory profiles of human brain endothelia and pericytes. J. Neuroinflamm. 2018, 15, 138. [Google Scholar] [CrossRef]
- Candelario-Jalil, E.; Taheri, S.; Yang, Y.; Sood, R.; Grossetete, M.; Estrada, E.Y.; Fiebich, B.L.; Rosenberg, G.A. Cyclooxygenase Inhibition Limits Blood-Brain Barrier Disruption following Intracerebral Injection of Tumor Necrosis Factor-α in the Rat. J. Pharmacol. Exp. Ther. 2007, 323, 488–498. [Google Scholar] [CrossRef]
- Rochfort, K.; Collins, L.E.; McLoughlin, A.; Cummins, P.M. Tumour necrosis factor-α-mediated disruption of cerebrovascular endothelial barrier integrity in vitro involves the production of proinflammatory interleukin-6. J. Neurochem. 2016, 136, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.Q.; Fang, Z.; Chen, X.L.; Yang, S.; Zhou, Y.F.; Mao, L.; Xia, Y.P.; Jin, H.J.; Li, Y.N.; You, M.F.; et al. Microglia-derived TNF-α mediates endothelial necroptosis aggravating blood brain–barrier disruption after ischemic stroke. Cell Death Dis. 2019, 10, 487. [Google Scholar] [CrossRef]
- Liu, T.; Clark, R.K.; McDonnell, P.C.; Young, P.R.; White, R.F.; Barone, F.C.; Feuerstein, G.Z. Tumor necrosis factor-alpha expression in ischemic neurons. Stroke 1994, 25, 1481–1488. [Google Scholar] [CrossRef] [Green Version]
- Welser-Alves, J.V.; Boroujerdi, A.; Tigges, U.; Milner, R. Analysis of TNF-alpha-Mediated Cerebral Pericyte Remodeling. Adv. Struct. Saf. Stud. 2014, 1155, 81–93. [Google Scholar] [CrossRef]
- Florian, M.; Florianova, L.; Hussain, S.; Magder, S. Interaction of Estrogen and Tumor Necrosis Factor α in Endothelial Cell Migration and Early Stage of Angiogenesis. Endothelium 2008, 15, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Fortini, F.; Sega, F.V.D.; Caliceti, C.; Aquila, G.; Pannella, M.; Pannuti, A.; Miele, L.; Ferrari, R.; Rizzo, P. Estrogen receptor β–dependent Notch1 activation protects vascular endothelium against tumor necrosis factor α (TNFα)-induced apoptosis. J. Biol. Chem. 2017, 292, 18178–18191. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Cheng, Y.; Simoncini, T.; Xu, S. 17β-Estradiol inhibits TNF-α-induced proliferation and migration of vascular smooth muscle cells via suppression of TRAIL. Gynecol. Endocrinol. 2016, 32, 581–586. [Google Scholar] [CrossRef]
- Ito, A.; Bebo, B.F.; Matejuk, A.; Zamora, A.; Silverman, M.; Fyfe-Johnson, A.; Offner, H. Estrogen Treatment Down-Regulates TNF-α Production and Reduces the Severity of Experimental Autoimmune Encephalomyelitis in Cytokine Knockout Mice. J. Immunol. 2001, 167, 542–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.; Ribeiro, R.C.J.; Webb, P.; Gustafsson, J.-Å.; Kushner, P.J.; Baxter, J.D.; Leitman, D.C. Estradiol repression of tumor necrosis factor-alpha transcription requires estrogen receptor activation function-2 and is enhanced by coactivators. Proc. Natl. Acad. Sci. USA 1999, 96, 15161–15166. [Google Scholar] [CrossRef] [Green Version]
- Arenas, I.A.; Armstrong, S.J.; Xu, Y.; Davidge, S.T. Chronic Tumor Necrosis Factor-α Inhibition Enhances NO Modulation of Vascular Function in Estrogen-Deficient Rats. Hypertension 2005, 46, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, Y.; Bai, Y.; Chao, J.; Hu, G.; Chen, X.; Yao, H. Involvement of PUMA in pericyte migration induced by methamphetamine. Exp. Cell Res. 2017, 356, 28–39. [Google Scholar] [CrossRef]
- Xiao, H.; Deng, M.; Yang, B.; Hu, Z.; Tang, J. Pretreatment with 17β-Estradiol Attenuates Cerebral Ischemia-Induced Blood-Brain Barrier Disruption in Aged Rats: Involvement of Antioxidant Signaling. Neuroendocrinology 2017, 106, 20–29. [Google Scholar] [CrossRef]
- Cvoro, A.; Tatomer, D.; Tee, M.-K.; Zogovic, T.; Harris, H.A.; Leitman, D.C. Selective estrogen receptor-β agonists repress transcription of proinflammatory genes. J. Immunol. 2008, 180, 630–636. [Google Scholar] [CrossRef]
- Tiwari-Woodruff, S.K.; Morales, L.B.; Lee, R.R.; Voskuhl, R.R. Differential neuroprotective and anti-inflammatory effects of estrogen receptor ligands. J. Neurochem. 2007, 102, 140. [Google Scholar]
- Xing, D.; Feng, W.; Miller, A.P.; Weathington, N.M.; Chen, Y.-F.; Novak, L.; Blalock, J.E.; Oparil, S. Estrogen modulates TNF-α-induced inflammatory responses in rat aortic smooth muscle cells through estrogen receptor-β activation. Am. J. Physiol. Circ. Physiol. 2007, 292, H2607–H2612. [Google Scholar] [CrossRef]
- Maggioli, E.; McArthur, S.; Mauro, C.; Kieswich, J.; Kusters, D.; Reutelingsperger, C.; Yaqoob, M.; Solito, E. Estrogen protects the blood–brain barrier from inflammation-induced disruption and increased lymphocyte trafficking. Brain Behav. Immun. 2016, 51, 212–222. [Google Scholar] [CrossRef] [Green Version]
- Vaccari, J.P.D.R.; Bramlett, H.M.; Perez-Pinzon, M.A.; Raval, A.P. Estrogen preconditioning: A promising strategy to reduce inflammation in the ischemic brain. Cond. Med. 2019, 2, 106–113. [Google Scholar]
- Asl, S.Z.; Khaksari, M.; Khachki, A.S.; Shahrokhi, N.; Nourizade, S. Contribution of estrogen receptors alpha and beta in the brain response to traumatic brain injury. J. Neurosurg. 2013, 119, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Spence, R.D.; Wisdom, A.; Cao, Y.; Hill, H.M.; Mongerson, C.R.L.; Stapornkul, B.; Itoh, N.; Sofroniew, M.V.; Voskuhl, R.R. Estrogen Mediates Neuroprotection and Anti-Inflammatory Effects during EAE through ER Signaling on Astrocytes But Not through ER Signaling on Astrocytes or Neurons. J. Neurosci. 2013, 33, 10924–10933. [Google Scholar] [CrossRef] [PubMed]
- Kenagy, R.D.; Civelek, M.; Kikuchi, S.; Chen, L.; Grieff, A.; Sobel, M.; Lusis, A.J.; Clowes, A.W. Scavenger receptor class A member 5 ( SCARA5 ) and suprabasin ( SBSN ) are hub genes of coexpression network modules associated with peripheral vein graft patency. J. Vasc. Surg. 2015, 64, 202–209.e6. [Google Scholar] [CrossRef] [Green Version]
- Špiranec, K.; Chen, W.; Werner, F.; Nikolaev, V.O.; Naruke, T.; Koch, F.; Werner, A.; Eder-Negrin, P.; Diéguez-Hurtado, R.; Adams, R.H.; et al. Endothelial C-Type Natriuretic Peptide Acts on Pericytes to Regulate Microcirculatory Flow and Blood Pressure. Circulation 2018, 138, 494–508. [Google Scholar] [CrossRef]
- Zechariah, A.; ElAli, A.; Doeppner, T.R.; Jin, F.; Hasan, M.R.; Helfrich, I.; Mies, G.; Hermann, D.M. Vascular Endothelial Growth Factor Promotes Pericyte Coverage of Brain Capillaries, Improves Cerebral Blood Flow During Subsequent Focal Cerebral Ischemia, and Preserves the Metabolic Penumbra. Stroke 2013, 44, 1690–1697. [Google Scholar] [CrossRef] [Green Version]
- Shah, K.; McCormack, C.E.; Bradbury, N.A. Do you know the sex of your cells? Am. J. Physiol. Cell Physiol. 2014, 306, C3–C18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardot-Leccia, N.; Hubiche, T.; Dellamonica, J.; Burel-Vandenbos, F.; Passeron, T. Pericyte alteration sheds light on micro-vasculopathy in COVID-19 infection. Intensiv. Care Med. 2020, 46, 1777–1778. [Google Scholar] [CrossRef] [PubMed]
- Galani, I.-E.; Rovina, N.; Lampropoulou, V.; Triantafyllia, V.; Manioudaki, M.; Pavlos, E.; Koukaki, E.; Fragkou, P.C.; Panou, V.; Rapti, V.; et al. Untuned antiviral immunity in COVID-19 revealed by temporal type I/III interferon patterns and flu comparison. Nat. Immunol. 2021, 22, 32–40. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Du, K.Y.; Peng, C. Incidence, clinical features, and outcomes of COVID-19 in Canada: Impact of sex and age. J. Ovarian Res. 2020, 13, 137. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.-H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Description | Log2 FC (Co- vs. Mono-Culture) | FDR p-Value |
|---|---|---|---|
| FADS1 [44]; MIR1908 [45] | Fatty acid desaturase 1; microRNA 1908 | −3.1 | 0.208 |
| ZFYVE16 [46,47] | Zinc finger, FYVE domain containing 16 | −3.1 | 0.058 |
| SIX4 [48] | SIX homeobox 4 | −2.8 | 0.333 |
| RBBP9 [49] | Retinoblastoma binding protein 9 | −2.5 | 0.315 |
| CPSF2 [50] | Cleavage and polyadenylation specific factor 2 | −2.3 | 0.208 |
| RAB33B [51] | RAB33B, member RAS oncogene family | −2.3 | 0.208 |
| GUF1 [52] | GUF1 homolog, GTPase | −2.2 | 0.333 |
| TOPORS [53,54] | Topoisomerase I binding, arginine/serine-rich, E3 ubiquitin protein ligase | −2.2 | 0.249 |
| NUP58 [55] | Nucleoporin 58kDa | −2.1 | 0.208 |
| PPP4R2 [56] | Protein phosphatase 4, regulatory subunit 2 | −2.1 | 0.315 |
| EREG [57] | Epiregulin | −2.1 | 0.315 |
| B3GNT5 [58] | UDP-GlcNAc:betaGal beta-1,3-N-acetylglucosaminyltransferase 5 | −2.0 | 0.208 |
| REXO4 [59,60] | REX4 homolog, 3’-5’ exonuclease | 2.8 | 0.249 |
| NDUFAF6 [61] | NADH dehydrogenase (ubiquinone) complex I, assembly factor 6 | 2.4 | 0.249 |
| ZNF582-AS1 [62] | ZNF582 antisense RNA 1 (head to head) | 2.2 | 0.315 |
| DACT1 [63] | Dishevelled Binding Antagonist Of Beta Catenin 1 | 2.1 | 0.332 |
| GLIPR1L1 [64] | GLI pathogenesis-related 1 like 1 | 2.1 | 0.315 |
| Gene | Gene Description | Log2 FC (Co- vs. Mono-Culture) | FDR p-Value |
|---|---|---|---|
| TARSL2 [68] | Threonyl-tRNA synthetase-like 2 | 2.7 | 0.335 |
| BEND6 [69,70] | BEN domain containing 6 | 2.3 | 0.335 |
| HIST1H3I [71,72,73] | Histone cluster 1, H3i | −3.3 | 0.335 |
| HIST1H4D [71,72,73] | Histone cluster 1, H4d | −2.0 | 0.335 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurmann, L.; Okoniewski, M.; Dubey, R.K. Estradiol Inhibits Human Brain Vascular Pericyte Migration Activity: A Functional and Transcriptomic Analysis. Cells 2021, 10, 2314. https://doi.org/10.3390/cells10092314
Kurmann L, Okoniewski M, Dubey RK. Estradiol Inhibits Human Brain Vascular Pericyte Migration Activity: A Functional and Transcriptomic Analysis. Cells. 2021; 10(9):2314. https://doi.org/10.3390/cells10092314
Chicago/Turabian StyleKurmann, Lisa, Michal Okoniewski, and Raghvendra K. Dubey. 2021. "Estradiol Inhibits Human Brain Vascular Pericyte Migration Activity: A Functional and Transcriptomic Analysis" Cells 10, no. 9: 2314. https://doi.org/10.3390/cells10092314
APA StyleKurmann, L., Okoniewski, M., & Dubey, R. K. (2021). Estradiol Inhibits Human Brain Vascular Pericyte Migration Activity: A Functional and Transcriptomic Analysis. Cells, 10(9), 2314. https://doi.org/10.3390/cells10092314