
In Vitro Growth Conditions Boost Plant Lipid Remodelling and Influence Their Composition


Abstract
:1. Introduction
2. Materials and Methods
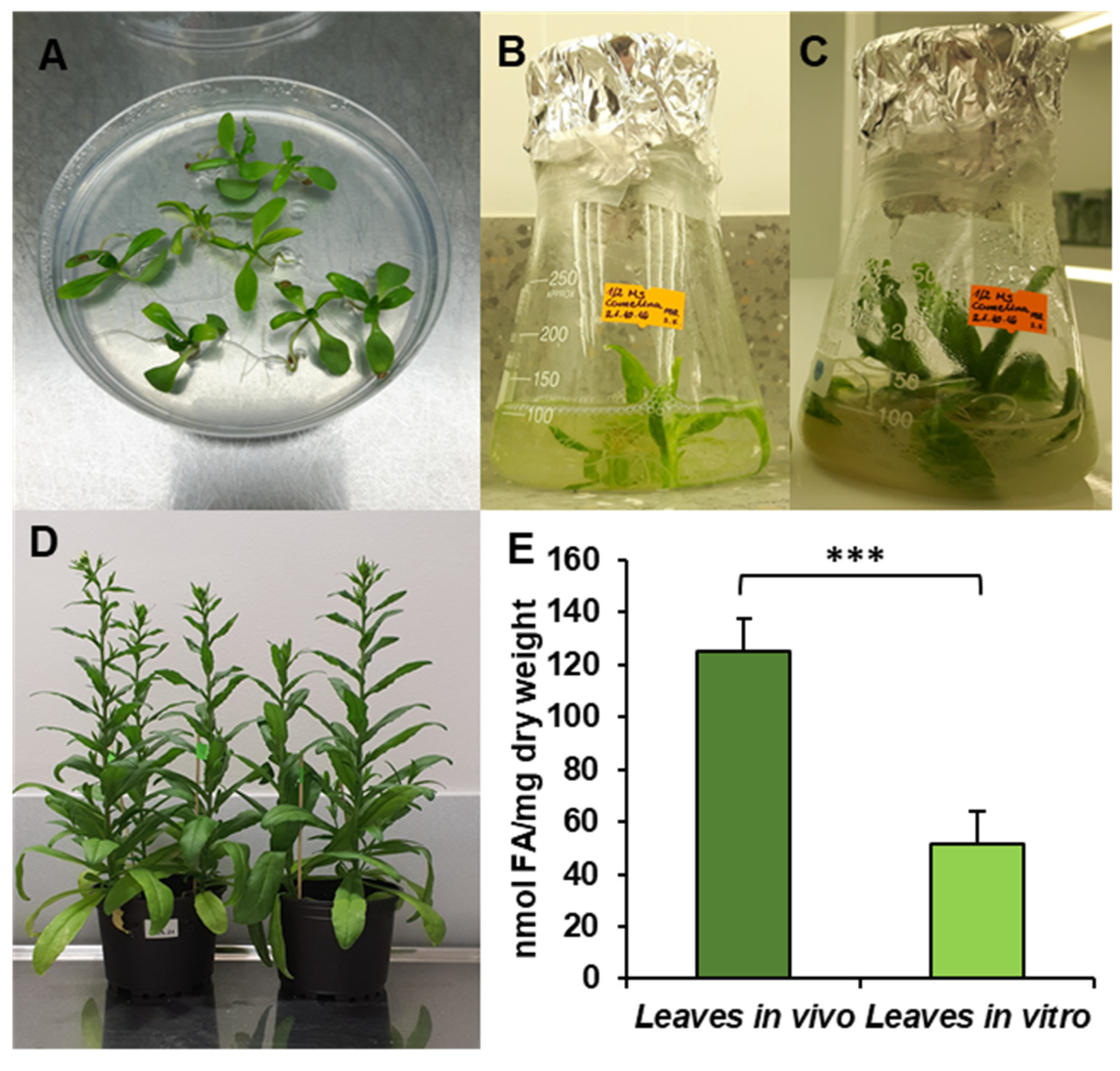
2.1. Plant Material and Growth Conditions
2.2. Lipid Analysis
2.3. Microsomal Membrane Preparation
2.4. Enzyme Assay
3. Results
3.1. Contents and Composition of Fatty Acids of Acyl-Lipids in Analysed Leaves
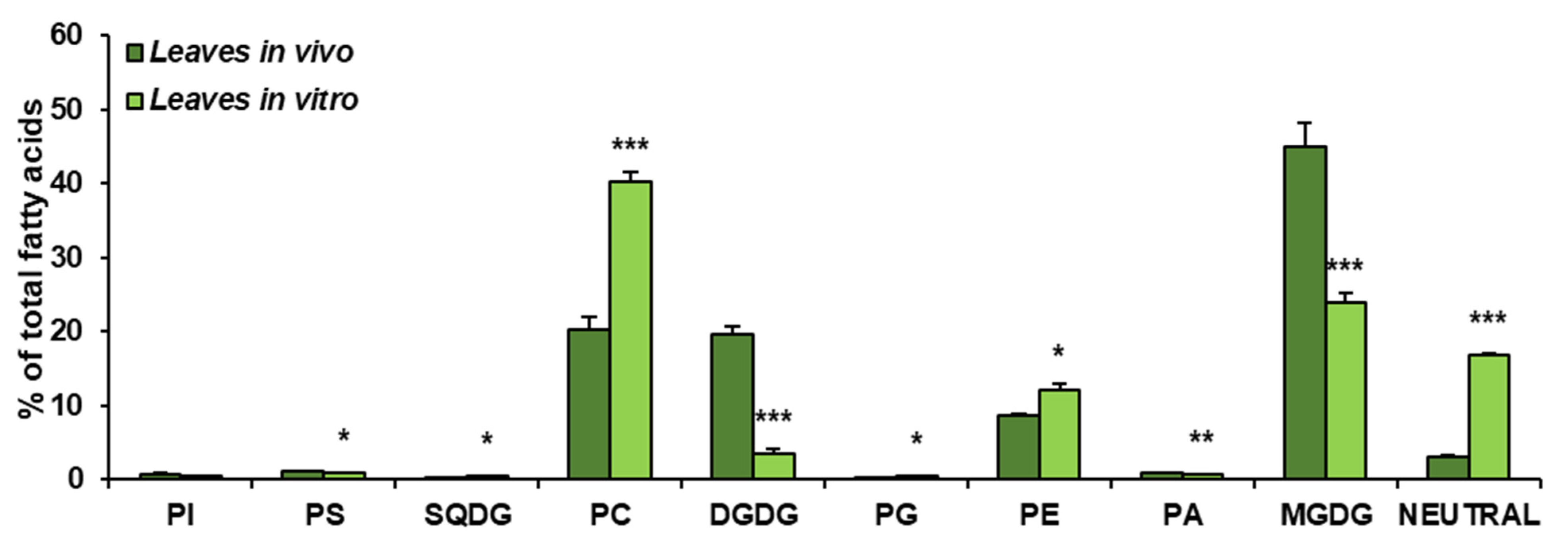
3.2. Lipid Classes in Analysed Leaves of C. sativa from In Vivo and In Vitro Conditions
3.3. Fatty Acid Composition of Main Lipid Classes of C. sativa Leaves from In Vivo and In Vitro Conditions
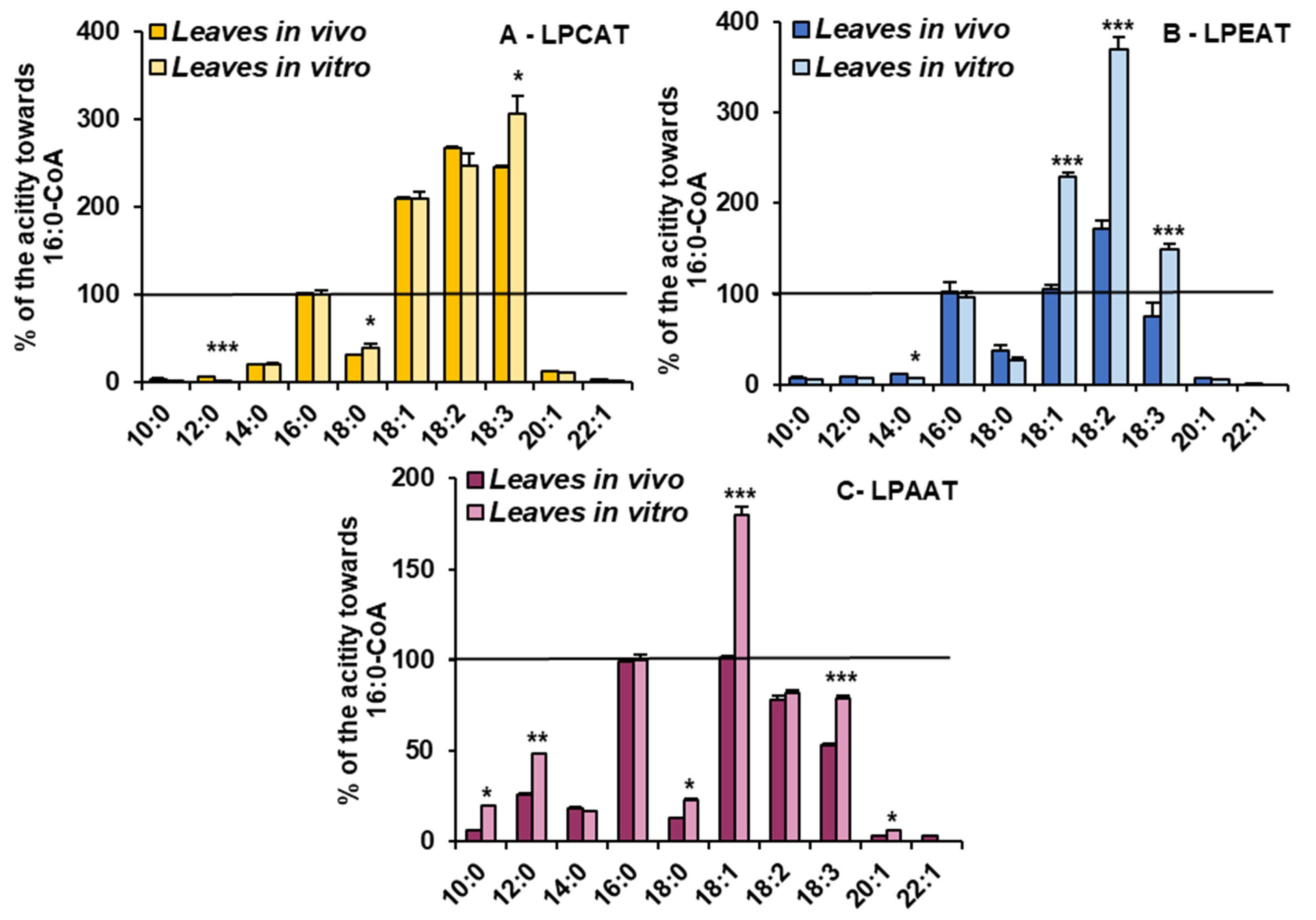
3.4. Activity and Substrate Specificity of Acyl-CoA:lysophospholipid Acyltransferases of C. sativa Leaves from In Vivo and In Vitro Conditions
3.5. Phospholipid Remodelling Intensity in C. sativa Leaves from In Vivo and In Vitro Conditions
4. Discussion
4.1. In Vitro Growth Conditions Clearly Decline Prokaryotic Pathway Activity
4.2. LPLAT Enzyme Activity and Remodelling Intensity Are Enhanced in In Vitro Cultivated Leaves
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Block, M.A.; Dorne, A.J.; Joyard, J.; Douce, R. Preparation and characterization of membrane fractions enriched in outer and inner envelope membranes from spinach chloroplasts. II. Biochemical characterization. J. Biol. Chem. 1983, 258, 13281–13286. [Google Scholar] [CrossRef]
- Mizusawa, N.; Wada, H. The role of lipids in photosystem II. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 194–208. [Google Scholar] [CrossRef] [Green Version]
- Dörmann, P. Galactolipids in Plant Membranes. In Enclopedia of Life Sciences; Jon Wiley & Sons Ltd.: Hobokern, NJ, USA, 2013. [Google Scholar]
- Kobayashi, K.; Endo, K.; Wada, H. Roles of lipids in photosynthesis. In Lipids in Plant and Algae Development; Nakamura, Y., Li-Beisson, Y., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 21–49. [Google Scholar]
- Ohlrogge, J.; Browse, J. Lipid biosynthesis. Plant Cell 1995, 7, 957–970. [Google Scholar]
- Birner, R.; Bürgermeister, M.; Schneiter, R.; Daum, G. Roles of phosphatidylethanolamine and of its several biosynthetic pathways in Saccharomyces cerevisiae. Mol. Biol. Cell 2001, 12, 997–1007. [Google Scholar] [CrossRef] [Green Version]
- Gibellini, F.; Smith, T.K. The Kennedy pathway-De novo synthesis of phosphatidylethanolamine and phosphatidylcholine. IUBMB Life 2010, 62, 414–428. [Google Scholar] [CrossRef]
- Jasieniecka-Gazarkiewicz, K.; Demski, K.; Gidda, S.K.; Klińska, S.; Niedojadło, J.; Lager, I.; Carlsson, A.S.; Minina, E.A.; Mullen, R.T.; Bozhkov, P.V.; et al. Subcellular Localization of Acyl-CoA: Lysophosphatidylethanolamine Acyltransferases (LPEATs) and the Effects of Knocking-Out and Overexpression of Their Genes on Autophagy Markers Level and Life Span of A. thaliana. Int. J. Mol. Sci. 2021, 22, 3006. [Google Scholar] [CrossRef]
- Testerink, C.; Munnik, T. Molecular, cellular, and physiological responses to phosphatidic acid formation in plants. J. Exp. Bot. 2011, 62, 2349–2361. [Google Scholar] [CrossRef] [Green Version]
- Meijer, H.J.; Munnik, T. Phospholipid-based signaling in plants. Annu. Rev. Plant Biol. 2003, 54, 265–306. [Google Scholar] [CrossRef]
- Dörmann, P. Membrane lipids. In Plant Lipids. Biology, Utilisation and Manipulation; Murphy, D.J., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2005; pp. 123–161. [Google Scholar]
- Kennedy, E.P. Biosynthesis of complex lipids. Fed. Proc. 1961, 20, 934–940. [Google Scholar]
- Frentzen, M. Acyltransferases from basic science to modified seed oils. Fett/Lipid 1998, 100, 161–166. [Google Scholar] [CrossRef]
- Slack, C.R.; Campbell, L.C.; Browse, J.A.; Roughan, P.G. Some evidence for the reversibility of the cholinephosphtranferase-catalysed reaction in developing linseed cotyledons in vivo. Biochem Biophys. Acta 1983, 754, 10–20. [Google Scholar] [CrossRef]
- Somerville, C.; Browse, J.; Jaworski, J.G.; Ohlorgge, J.B. Lipids. In Biochemistry and Molecular Biology of Plants; Buchnanan, B.B., Gruissem, W., Jones, J.L., Eds.; American Society of Plant Biologists: Rockville, MD, USA, 2001; pp. 456–527. [Google Scholar]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; Debono, A.; Durrett, T.P.; et al. Arabidopsis Book; Robt, L., Ed.; American Society of Plant Biologist: Rockville, MD, USA, 2013; pp. 1–70. [Google Scholar]
- Joyard, J.; Block, M.A.; Malherbe, A.; Marachal, E.; Douce, R. Origin and synthesis of galactolipid and sulfolipid headgroups. In Lipid Metabolism in Plants; Moore, T.S., Ed.; CRC Press: Boca Raton, FA, USA, 1993; pp. 231–258. [Google Scholar]
- Lands, W.E. Metabolism of glycerolipides; a comparison of lecithin and triglyceride synthesis. J. Biol. Chem. 1958, 231, 883–888. [Google Scholar] [CrossRef]
- Lager, I.; Yilmaz, J.L.; Zhou, X.R.; Jasieniecka, K.; Kazachkov, M.; Wang, P.; Zou, J.; Weselake, R.; Smith, M.A.; Bayon, S.; et al. Plant acyl-CoA:lysophosphatidylcholine acyltransferases (LPCATs) have different specificities in their forward and reverse reactions. J. Biol. Chem. 2013, 288, 36902–36914. [Google Scholar] [CrossRef] [Green Version]
- Jasieniecka-Gazarkiewicz, K.; Demski, K.; Lager, I.; Stymne, S.; Banaś, A. Possible role of different yeast and plant lysophospholipid:acyl-CoA acyltransferases (LPLATs) in acyl remodelling of phospholipids. Lipids 2016, 51, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Klińska, S.; Jasieniecka-Gazarkiewicz, K.; Banaś, A. Acyl-CoA: AzdLysophosphatidylcholine acyltransferases (LPCATs) of Camelina sativa seeds: Biochemical properties and function. Planta 2019, 250, 1655–1670. [Google Scholar] [CrossRef] [Green Version]
- Klińska, S.; Jasieniecka-Gazarkiewicz, K.; Demski, K.; Banaś, A. Editing of phosphatidic acid and phosphatidylethanolamine by acyl-CoA: Lysophospholipid acyltransferases in developing Camelina sativa seeds. Planta 2020, 10, 4. [Google Scholar] [CrossRef]
- Klińska, S.; Demski, K.; Jasieniecka-Gazarkiewicz, K.; Banaś, A. LPEATs Tailor Plant Phospholipid Composition through Adjusting Substrate Preferences to Temperature. Int. J. Mol. Sci. 2021, 22, 8137. [Google Scholar] [CrossRef]
- Cardoso, J.C.; de Oliveira, B.E.B.S.; Cardoso, F.d.I. Advances and challenges on the in vitro production of secondary metabolites from medicinal plants. Hortic. Bras. 2019, 37, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Verpoorte, R.; Contin, A.; Memelink, J. Biotechnology for the production of plant secondary metabolites. Phytochem. Rev. 2002, 1, 13–25. [Google Scholar] [CrossRef]
- Grzegorczyk-Karolak, I.; Matkowski, A.; Wysokińska, H. Antioxidant activity of extracts from in vitro cultures of Salvia officinalis L. Food Chem. 2007, 104, 536–541. [Google Scholar] [CrossRef]
- Khorasani, A.; Winardi, S.; Koshy, P.; Rosna, T.; Arash, R. Antioxidant and antibacterial activities of ethanolic extracts of Asparagus officinalis cv. Mary Washington: Comparison of in vivo and in vitro grown plant bioactivities. Afr. J. Biotechnol. 2010, 9, 8460–8466. [Google Scholar]
- Park, W.; Feng, Y.; Ahn, S.J. Alteration of leaf shape, improved metaltolerance, and productivity of seed by overexpression of CsHMA3 in Camelina sativa. Biotechnol. Biofuels. 2014, 7, 96. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Mao, X.; Zhao, K.; Ji, X.; Ji, C.; Xue, J.; Li, R. Characterisation of phospholipid: Diacylglycerol acyltransferases (PDATs) from Camelina sativa and their roles in stress responses. Biol. Open 2017, 6, 1024–1034. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Mao, X.; Zhao, K.; Sun, Y.; Ji, C.; Xue, J.; Li, R. Spatio-temporal expression and stress responses of DGAT1, DGAT2 and PDAT responsible for TAG biosynthesis in Camelina sativa. Emir. J. Food Agric. 2017, 29, 274–284. [Google Scholar] [CrossRef]
- Demski, K.; Łosiewska, A.; Jasieniecka-Gazarkiewicz, K.; Klińska, S.; Banaś, A. Phospholipid:diacylglycerol acyltransferase1 overexpression delays senescence and enhances post-heat and cold exposure fitness. Front. Plant Sci. 2020, 11, 611897. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. 1959, 37, 911–917. [Google Scholar]
- Sánchez, M.; David, G.N.; David, N.B. The relationship between palmitoyl-coenzyme A synthetase activity and esterifcation of sn-glycerol 3-phosphate in rat liver mitochondria. Biochem. J. 1973, 132, 697–706. [Google Scholar] [CrossRef] [Green Version]
- Miquel, M.; Browse, J. Arabidopsis mutants deficient in polyunsaturated fatty acid synthesis. Biochemical and genetic characterization of a plant oleoyl-phosphatidylcholine desaturase. J. Biol. Chem. 1992, 267, 1502–1509. [Google Scholar] [CrossRef]
- Falcone, D.L.; Ogas, J.P.; Somerville, C.R. Regulation of membrane fatty acid composition by temperature in mutants of Arabidopsis with alterations in membrane lipid composition. BMC Plant Biol. 2004, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Murakami, Y.; Tsuyama, M.; Kobayashi, Y.; Kodama, H.; Iba, K. Trienoic Fatty Acids and Plant Tolerance of High Temperature. Science 2000, 287, 476–479. [Google Scholar] [CrossRef]
- Yaeno, T.; Matsuda, O.; Iba, K. Role of chloroplast trienoic fatty acids in plant disease defense responses. Plant J. 2004, 40, 931–941. [Google Scholar] [CrossRef]
- Wang, K.; Guo, Q.; Froehlich, J.E.; Hersh, H.L.; Zienkiewicz, A.; Howe, G.A.; Benning, C. Two abscisic acid-responsive plastid lipase genes involved in jasmonic acid biosynthesis in Arabidopsis thaliana. Plant Cell. 2018, 30, 1006–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.; Ding, N.-Z. Plant Unsaturated Fatty Acids: Multiple Roles in Stress Response. Front. Plant Sci. 2020, 11, 562785. [Google Scholar] [CrossRef] [PubMed]
- Higashi, Y.; Okazaki, Y.; Takano, K.; Myouga, F.; Shinozaki, K.; Knoch, E.; Fukushima, A.; Saito, K. Remodels chloroplastic monogalactosyldiacylglycerol by liberating α-linolenic acid in Arabidopsis leaves under heat stress. Plant Cell. 2018, 30, 1887–1905. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Chen, G.; Kazachkov, M.; Greer, M.S.; Caldo, K.M.; Zou, J.; Weselake, R.J. In vivo and in vitro evidence for biochemical coupling of reactions catalyzed by lysophosphatidylcholine acyltransferase and diacylglycerol acyltransferase. J. Biol. Chem. 2015, 290, 18068–18078. [Google Scholar] [CrossRef] [Green Version]
- Stålberg, K.; Ståhl, U.; Stymne, S.; Ohlrogge, J. Characterization of two Arabidopsis thaliana acyltransferases with preference for lysophosphatidylethanolamine. BMC Plant Biol. 2009, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Jasieniecka-Gazarkiewicz, K.; Lager, I.; Carlsson, A.S.; Gutbrod, K.; Peisker, H.; Dörmann, P.; Stymne, S.; Banaś, A. Acyl-CoA:Lysophosphatidylethanolamine Acyltransferase Activity Regulates Growth of Arabidopsis. Plant Physiol. 2017, 174, 986–998. [Google Scholar] [CrossRef] [Green Version]
- Oo, K.C.; Huang, A.H. Lysophosphatidate acyltransferase activities in the microsomes from palm endosperm, maize scutellum, and rapeseed cotyledon of maturing seeds. Plant Physiol. 1989, 91, 1288–1295. [Google Scholar] [CrossRef] [Green Version]
- Lassner, M.W.; Levering, C.K.; Davies, H.M.; Knutzon, D.S. Lysophosphatidic acid acyltransferase from meadowfoam mediates insertion of erucic acid at the sn-2 position of triacylglycerol in transgenic rapeseed oil. Plant Physiol. 1995, 109, 1389–1394. [Google Scholar] [CrossRef] [Green Version]
- Arroyo-Caro, J.M.; Chileh, T.; Kazachkov, M.; Zou, J.; Alonso, D.L.; García-Maroto, F. The multigene family of lysophosphatidate acyltransferase (LPAT)-related enzymes in Ricinus communis: Cloning and molecular characterization of two LPAT genes that are expressed in castor seeds. Plant Sci. 2013, 199–200, 29–40. [Google Scholar] [CrossRef]
- Fahs, Z.; Rossez, Y.; Guénin, S.; Gutierrez, L.; Thomasset, B.; Perrin, Y. Cloning and molecular characterization of three lysophosphatidic acid acyltransferases expressed in fax seeds. Plant Sci. 2019, 280, 41–50. [Google Scholar] [CrossRef]
- Abdullah, H.M.; Akbari, P.; Paulose, B.; Schnell, D.; Qi, W.; Park, Y.; Pareek, A.; Dhankher, O.P. Transcriptome profling of Camelina sativa to identify genes involved in triacylglycerol biosynthesis and accumulation in the developing seeds. Biotechnol Biofuels 2016, 9, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagale, S.; Koh, C.; Nixon, J.; Bollina, V.; Clarke, W.E.; Tuteja, R.; Spillane, C.; Robinson, S.J.; Links, M.G.; Clarke, C.; et al. The emerging biofuel crop Camelina sativa retains a highly undifferentiated hexaploid genome structure. Nat. Commun. 2014, 5, 3706. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Class | Type of Cultivation Condition | Fatty Acids (mol%) | ||||||
|---|---|---|---|---|---|---|---|---|
| 16:0 | 16:3 | 18:0 | 18:1 | 18:2 | 18:3 | Others | ||
| PE | In vitro | 25.5 | - | 4.0 | 10.6 *** | 26.4 | 28.4 | 5.0 ** |
| ±0.2 | ±0.01 | ±0.1 | ±0.2 | ±0.8 | ±0.2 | |||
| In vivo | 37.4 | - | 4.8 | 5.9 | 24.0 | 26.3 | 1.5 | |
| ±3.0 | ±0.4 | ±0.3 | ±1.8 | ±1.7 | ±0.01 | |||
| PC | In vitro | 20.8 | - | 4.2 | 9.7 *** | 17.7 *** | 46.5 *** | 1.2 |
| ±0.3 | ±0.01 | ±0.1 | ±0.1 | ±0.2 | ±0.1 | |||
| In vivo | 21.9 | - | 3.2 | 0.9 | 4.3 | 68.0 | 1.4 | |
| ±2.5 | ±1.01 | ±0.1 | ±0.2 | ±3.0 | ±0.2 | |||
| MGDG | In vitro | 6.2 ** | 19.4 ** | - | 3.6 *** | 11.4 *** | 58.8 ** | 0.6 |
| ±0.1 | ±0.3 | ±0.1 | ±1.3 | ±0.9 | ±0.01 | |||
| In vivo | 4.0 | 30.0 | - | 0.5 | 3.0 | 62.6 | - | |
| ±0.2 | ±1.4 | ±0.4 | ±0.4 | ±0.6 | ||||
| DGDG | In vitro | 37.4 | 4.8 | - | 8.6* | 23.4 * | 25.4 ** | 0.5 |
| ±1.3 | ±0.1 | ±0.6 | ±0.6 | ±0.1 | ±0.01 | |||
| In vivo | 33.4 | 5.7 | - | 12.2 | 17.9 | 30.8 | - | |
| ±1.0 | ±0.3 | ±1.0 | ±0.8 | ±0.8 | ||||
| Type of Cultivation Condition | Fatty Acid Donors | Remodelling Intensity (pmol [14C]PL/nmol Microsomal PC/min) | ||
|---|---|---|---|---|
| PC | PE | PA | ||
| In vitro | [14C]18:1-CoA | 3.18 *** ±0.07 | 0.59 *** ±0.01 | 0.26 *** ±0.02 |
| [14C]18:2-CoA | 1.56 *** ±0.05 | 0.31 *** ±0.02 | 0.1 ** ±0.008 | |
| [14C18:3-CoA | 1.3 *** ±0.09 | 0.21 *** ±0.007 | 0.12 *** ±0.009 | |
| In vivo | [14C]18:1-CoA | 0.85 ±0.01 | 0.15 ±0.01 | 0.03 ±0.002 |
| [14C]18:2-CoA | 0.47 ±0.01 | 0.09 ±0.005 | 0.03 ±0.001 | |
| [14C18:3-CoA | 0.43 ±0.02 | 0.07 ±0.005 | 0.06 ±0.001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klińska, S.; Kędzierska, S.; Jasieniecka-Gazarkiewicz, K.; Banaś, A. In Vitro Growth Conditions Boost Plant Lipid Remodelling and Influence Their Composition. Cells 2021, 10, 2326. https://doi.org/10.3390/cells10092326
Klińska S, Kędzierska S, Jasieniecka-Gazarkiewicz K, Banaś A. In Vitro Growth Conditions Boost Plant Lipid Remodelling and Influence Their Composition. Cells. 2021; 10(9):2326. https://doi.org/10.3390/cells10092326
Chicago/Turabian StyleKlińska, Sylwia, Sara Kędzierska, Katarzyna Jasieniecka-Gazarkiewicz, and Antoni Banaś. 2021. "In Vitro Growth Conditions Boost Plant Lipid Remodelling and Influence Their Composition" Cells 10, no. 9: 2326. https://doi.org/10.3390/cells10092326
APA StyleKlińska, S., Kędzierska, S., Jasieniecka-Gazarkiewicz, K., & Banaś, A. (2021). In Vitro Growth Conditions Boost Plant Lipid Remodelling and Influence Their Composition. Cells, 10(9), 2326. https://doi.org/10.3390/cells10092326