
Regional Variation of Gap Junctional Connections in the Mammalian Inner Retina


 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Sample Preparation
2.2. Immunohistochemistry and Confocal Microscopy
2.3. Measurement of Retinal Eccentricity and Feature Density
2.4. 3D Colocalization Analysis
2.5. Data Analysis
3. Results
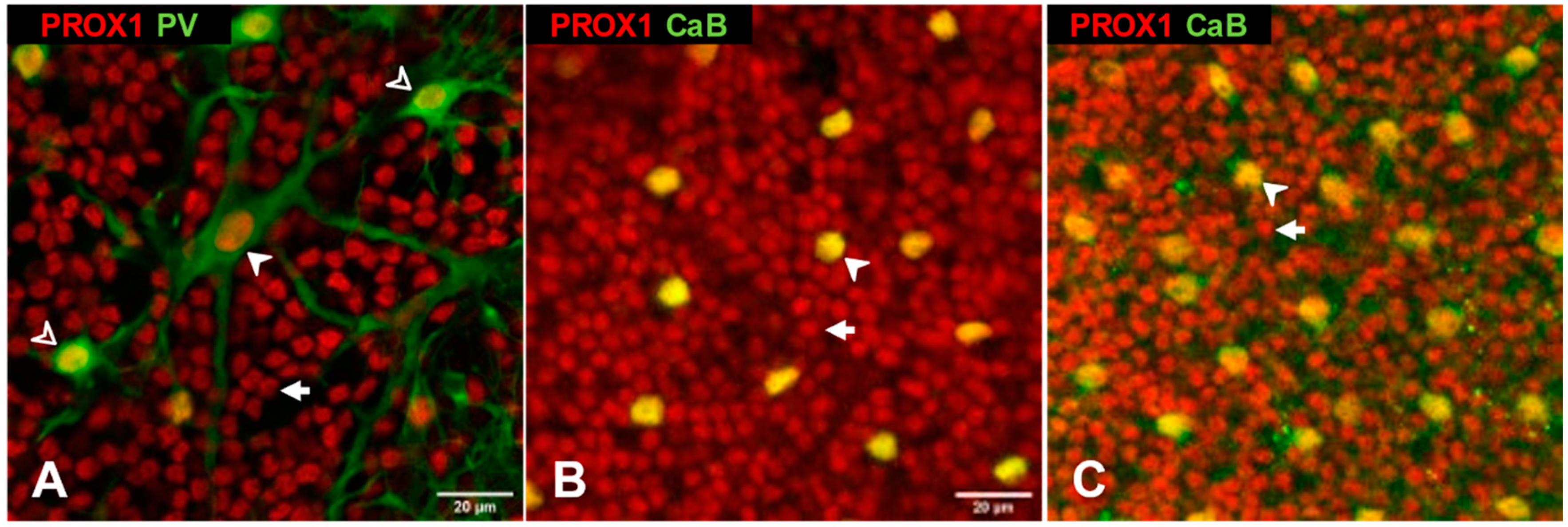
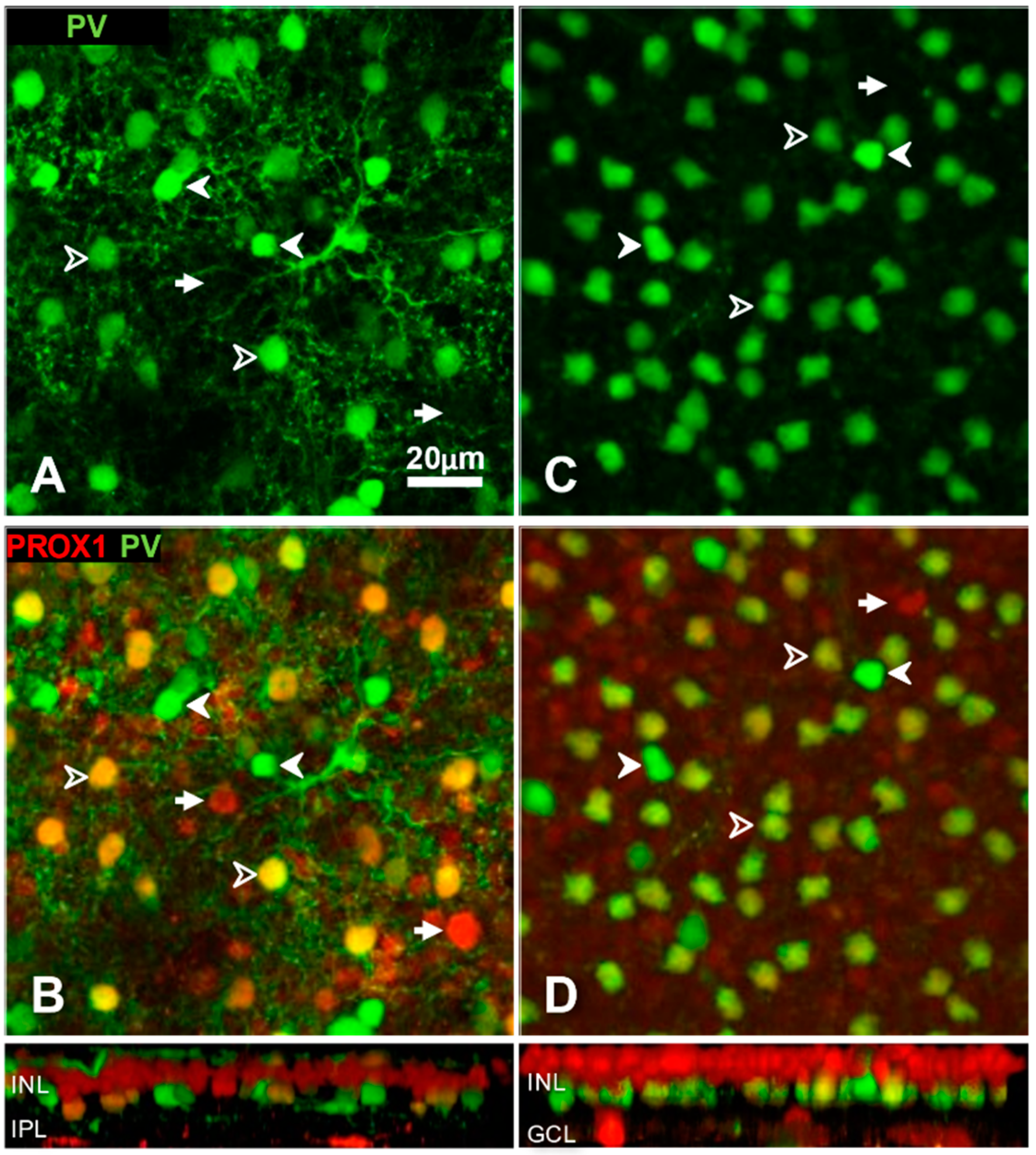
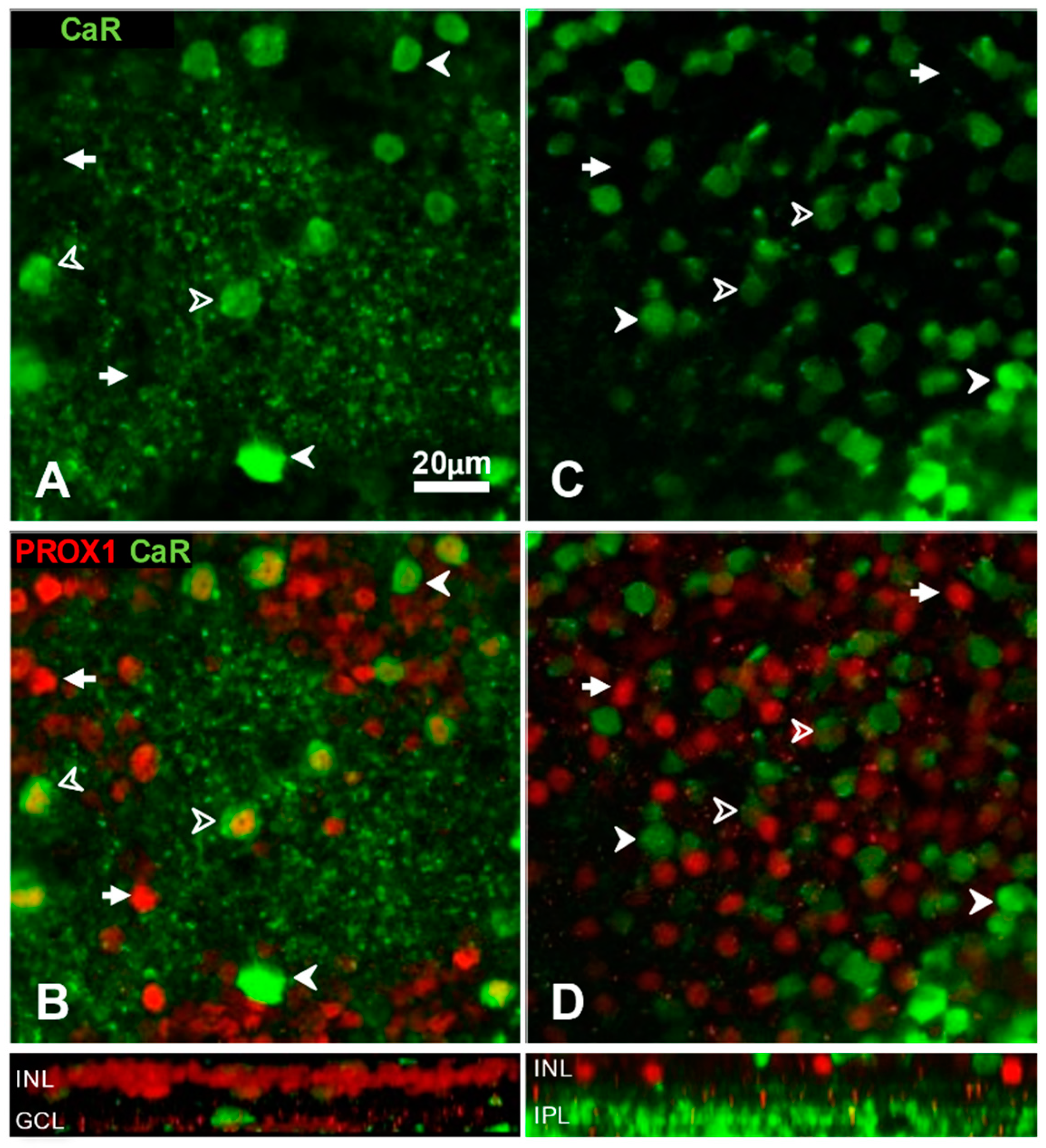
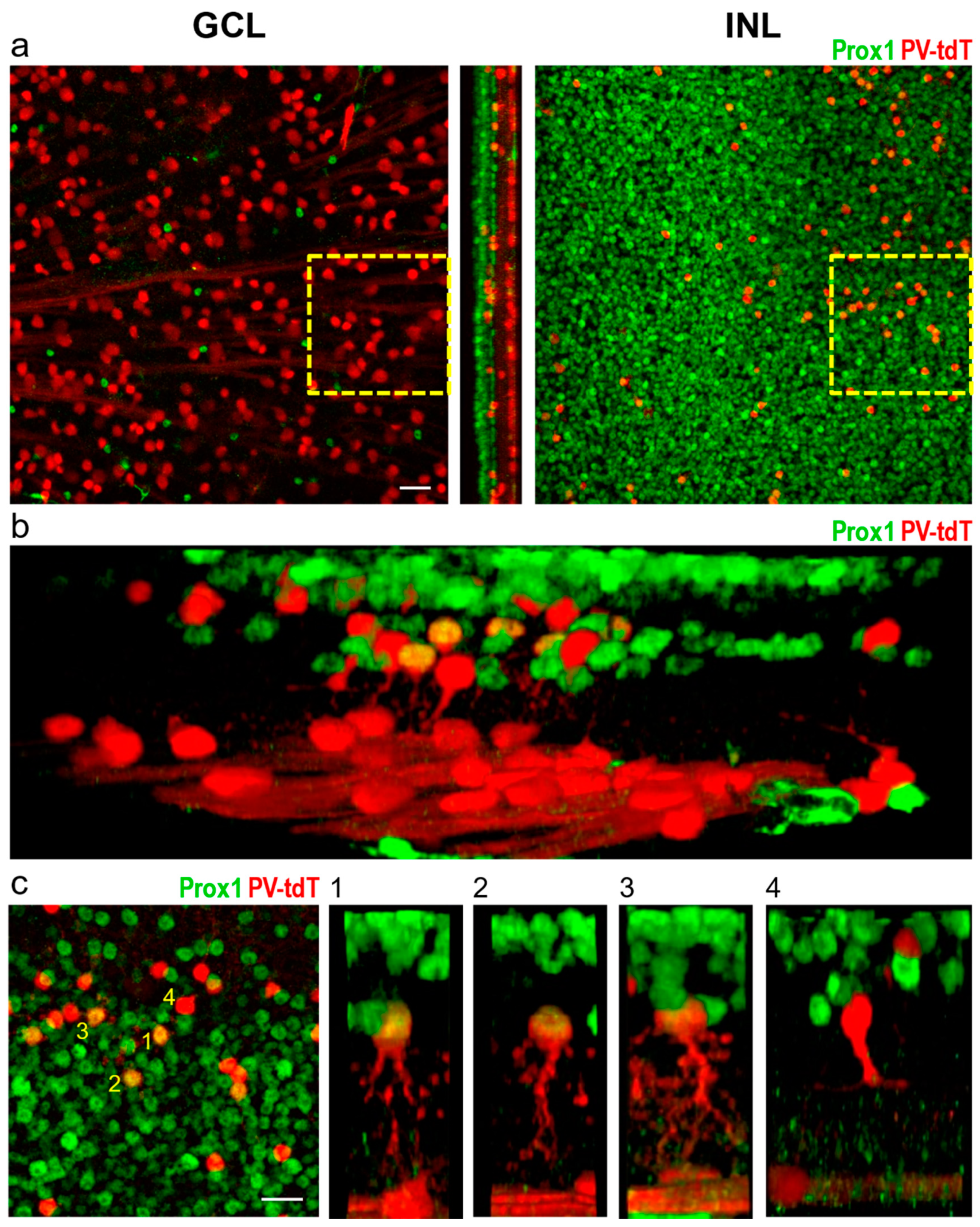
3.1. Prox1 Immunoreactive Cell Types in Cat, Rat and Mouse Retinas
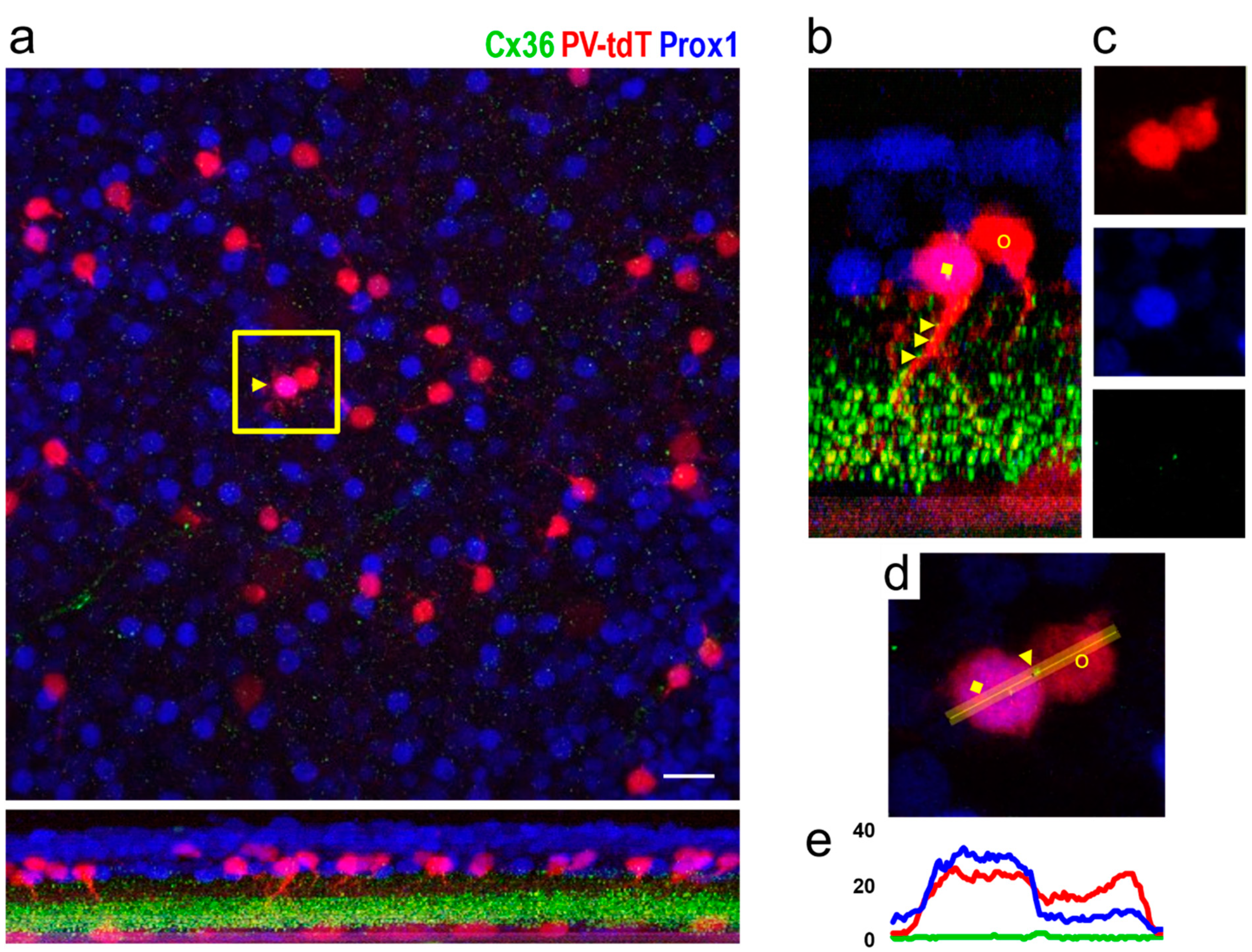
3.2. Identification of Individual AII Amacrine Cells in the PV-tdT Mouse Line
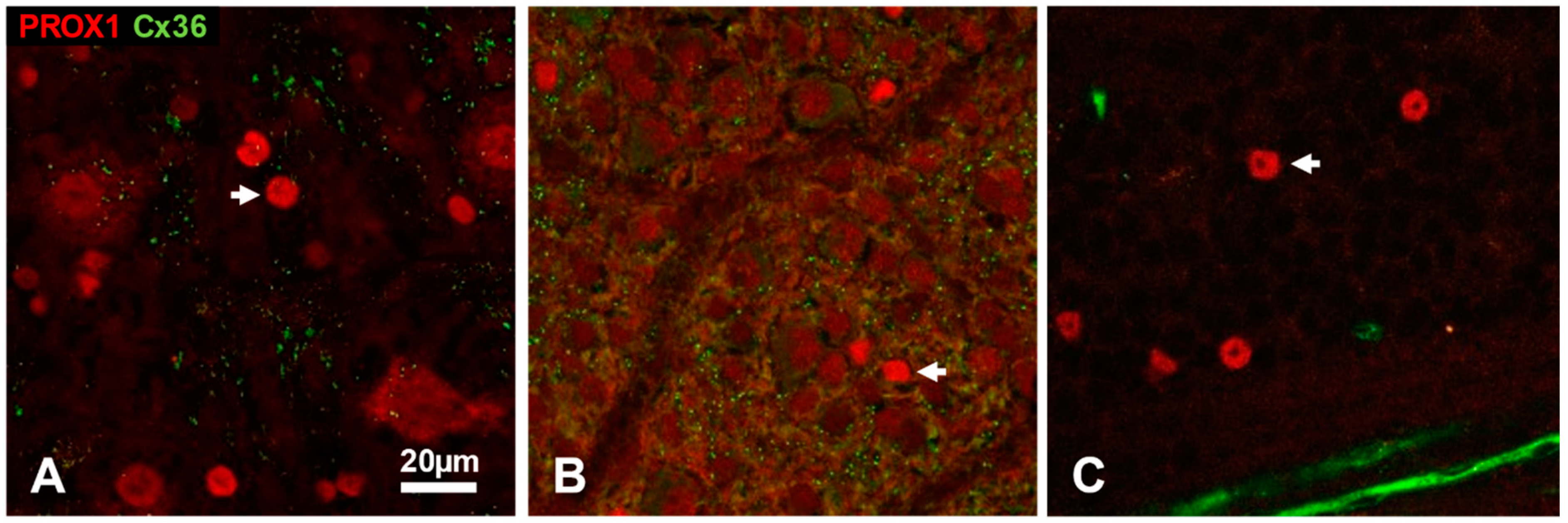
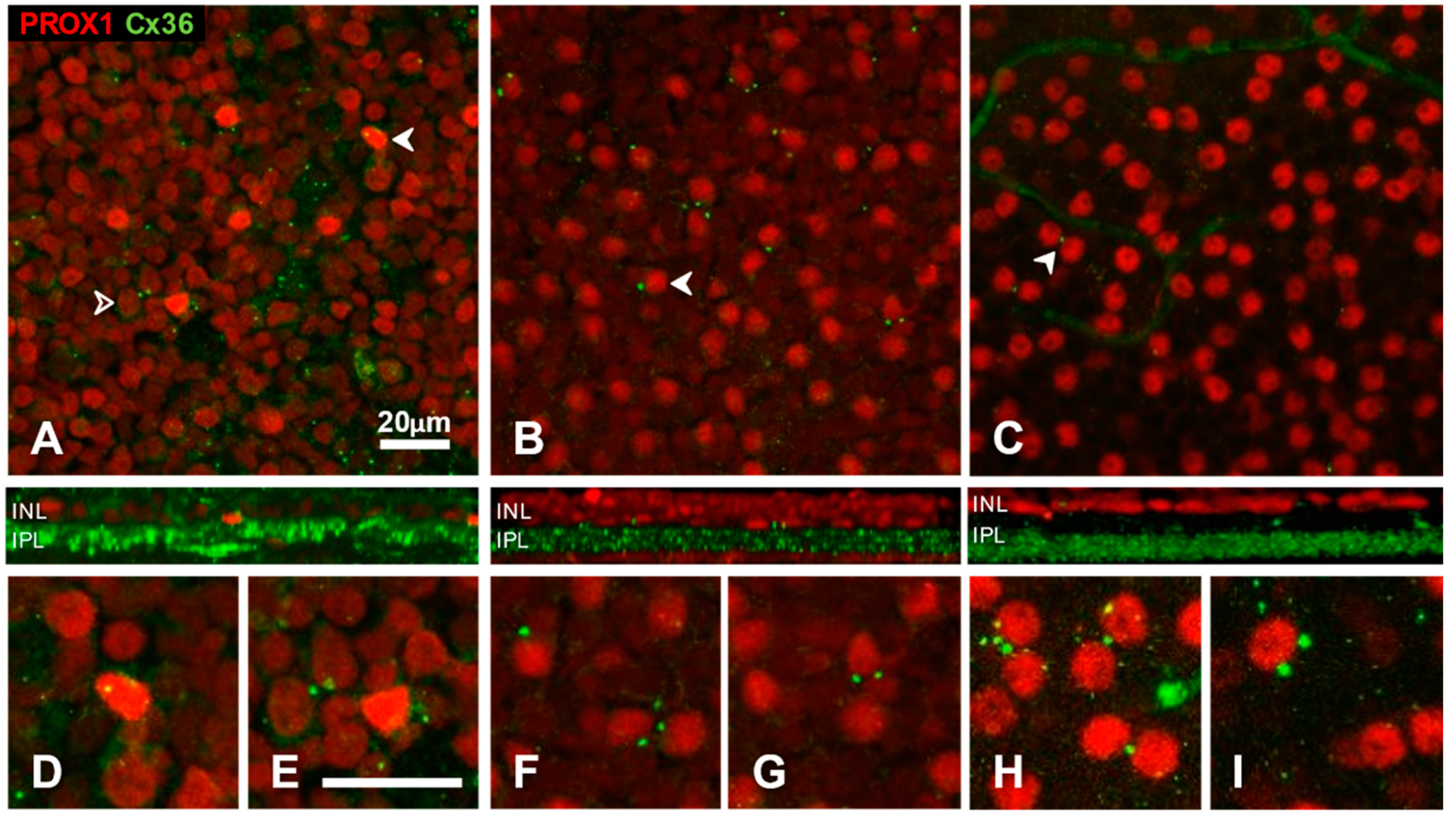
3.3. Somatic Cx36 Plaques on Amacrine Cells of the Cat, Rat and Mouse Retina
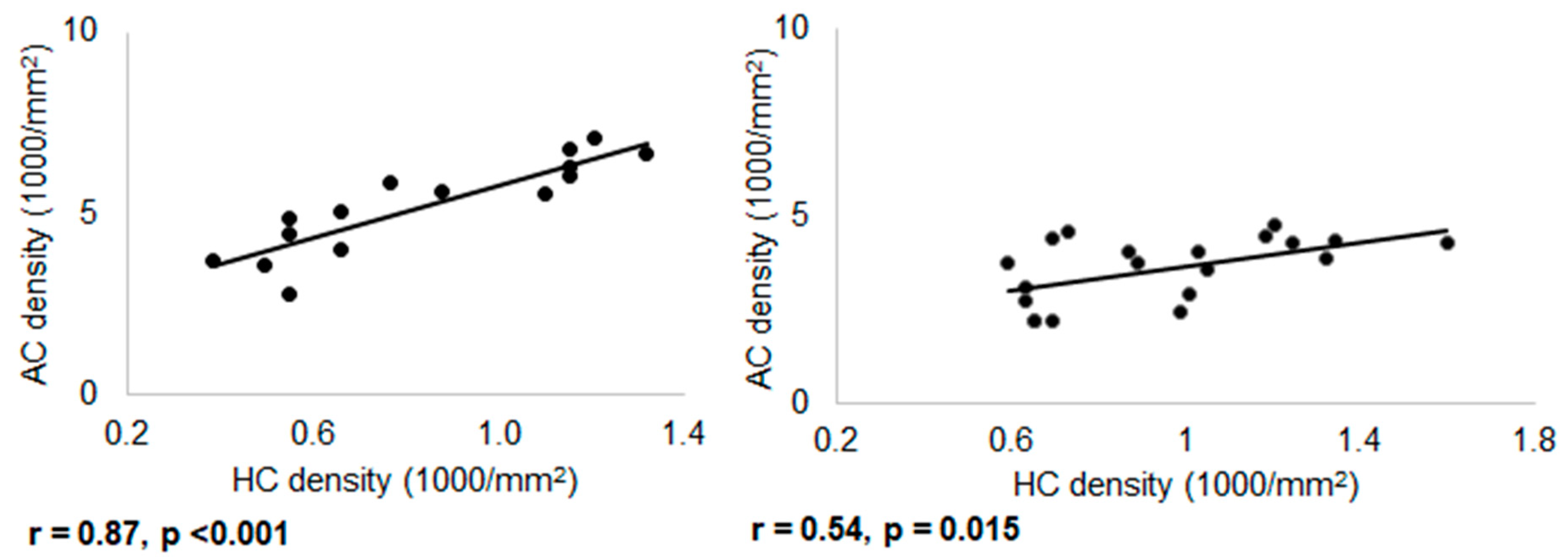
3.4. Regional Variation of Connexin-36 Density in the OFF- and ON-Sublaminae of the Inner Plexiform Layer
4. Discussion
4.1. Conservative Expression of Prox1 in Major Cell Types of the Inner Nuclear Layer
4.2. Somatic Gap Junctions of Retinal Amacrine Cells
4.3. Different Scaling Principles of Connexin-36 Gap Junction Density in ON and OFF Circuits with Retinal Position
4.4. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, B.A.; Tyrrell, L.P.; Kamilar, J.M.; Collin, S.P.; Dominy, N.J.; Hall, M.I.; Heesy, C.P.; Lisney, T.J.; Loew, E.R.; Moritz, G.L.; et al. Structure and Function of Regional Specializations in the Vertebrate Retina. In Evolution of Nervous Systems; Kaas, J.H., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 1, pp. 351–372. ISBN 978-0-12-804096-6. [Google Scholar]
- Ahnelt, P.K.; Kolb, H. The Mammalian Photoreceptor Mosaic-Adaptive Design. Prog. Retin. Eye Res. 2000, 19, 711–777. [Google Scholar] [CrossRef]
- Curcio, C.A.; Sloan, K.R.; Kalina, R.E.; Hendrickson, A.E. Human Photoreceptor Topography. J. Comp. Neurol. 1990, 292, 497–523. [Google Scholar] [CrossRef]
- Steinberg, R.H.; Reid, M.; Lacy, P.L. The Distribution of Rods and Cones in the Retina of the Cat (Felis Domesticus). J. Comp. Neurol. 1973, 148, 229–248. [Google Scholar] [CrossRef] [PubMed]
- Linberg, K.A.; Lewis, G.P.; Shaaw, C.; Rex, T.S.; Fisher, S.K. Distribution of S- and M-Cones in Normal and Experimentally Detached Cat Retina. J Comp Neurol 2001, 430, 343–356. [Google Scholar] [CrossRef]
- Curcio, C.A.; Allen, K.A. Topography of Ganglion Cells in Human Retina. J. Comp. Neurol. 1990, 300, 5–25. [Google Scholar] [CrossRef]
- Telkes, I.; Lee, S.C.; Jusuf, P.R.; Grünert, U. The Midget-Parvocellular Pathway of Marmoset Retina: A Quantitative Light Microscopic Study. J Comp Neurol 2008, 510, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Stewart, E.E.M.; Valsecchi, M.; Schütz, A.C. A Review of Interactions between Peripheral and Foveal Vision. J. Vis. 2020, 20, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Strasburger, H.; Rentschler, I.; Jüttner, M. Peripheral Vision and Pattern Recognition: A Review. J. Vis. 2011, 11, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, R. Cat Cones Have Rod Input: A Comparison of the Response Properties of Cones and Horizontal Cell Bodies in the Retina of the Cat. J. Comp. Neurol. 1977, 172, 109–135. [Google Scholar] [CrossRef]
- Schneeweis, D.M.; Schnapf, J.L. Photovoltage of Rods and Cones in the Macaque Retina. Science 1995, 268, 1053–1056. [Google Scholar] [CrossRef]
- Deans, M.R.; Völgyi, B.; Goodenough, D.A.; Bloomfield, S.A.; Paul, D.L. Connexin36 Is Essential for Transmission of Rod-Mediated Visual Signals in the Mammalian Retina. Neuron 2002, 36, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Güldenagel, M.; Ammermüller, J.; Feigenspan, A.; Teubner, B.; Degen, J.; Söhl, G.; Willecke, K.; Weiler, R.; Sohl, G.; Willecke, K.; et al. Visual Transmission Deficits in Mice with Targeted Disruption of the Gap Junction Gene Connexin. J. Neurosci. 2001, 21, 6036–6044. [Google Scholar] [CrossRef] [Green Version]
- Völgyi, B.; Deans, M.R.; Paul, D.L.; Bloomfield, S.A. Convergence and Segregation of the Multiple Rod Pathways in Mammalian Retina. J. Neurosci. 2004, 24, 11182–11192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naka, K.I.; Nye, P.W. Role of Horizontal Cells in Organization of the Catfish Retinal Receptive Field. J. Neurophysiol. 1971, 34, 785–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelley, J.; Dedek, K.; Schubert, T.; Feigenspan, A.; Schultz, K.; Hombach, S.; Willecke, K.; Weiler, R. Horizontal Cell Receptive Fields Are Reduced in Connexin57-Deficient Mice. Eur. J. Neurosci. 2006, 23, 3176–3186. [Google Scholar] [CrossRef] [PubMed]
- Hu, E.H.; Bloomfield, S.A. Gap Junctional Coupling Underlies the Short-Latency Spike Synchrony of Retinal α Ganglion Cells. J. Neurosci. 2003, 23, 6768–6777. [Google Scholar] [CrossRef] [PubMed]
- Völgyi, B.; Pan, F.; Paul, D.L.; Wang, J.T.; Huberman, A.D.; Bloomfield, S.A. Gap Junctions Are Essential for Generating the Correlated Spike Activity of Neighboring Retinal Ganglion Cells. PLoS ONE 2013, 8, e69426. [Google Scholar] [CrossRef] [Green Version]
- Mastronarde, D.N. Interactions between Ganglion Cells in Cat Retina. J. Neurophysiol. 1983, 49, 350–365. [Google Scholar] [CrossRef]
- Brivanlou, I.H.; Warland, D.K.; Meister, M. Mechanisms of Concerted Firing among Retinal Ganglion Cells. Neuron 1998, 20, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Bloomfield, S.A.; Völgyi, B. The Diverse Functional Roles and Regulation of Neuronal Gap Junctions in the Retina. Nat. Rev. Neurosci. 2009, 10, 495–506. [Google Scholar] [CrossRef]
- Völgyi, B.; Kovács-Öller, T.; Atlasz, T.; Wilhelm, M.; Gábriel, R. Gap Junctional Coupling in the Vertebrate Retina: Variations on One Theme? Prog. Retin. Eye Res. 2013, 34, 1–18. [Google Scholar] [CrossRef]
- O’Brien, J.J.; Chen, X.; Macleish, P.R.; O’Brien, J.J.; Massey, S.C. Photoreceptor Coupling Mediated by Connexin36 in the Primate Retina. J. Neurosci. 2012, 32, 4675–4687. [Google Scholar] [CrossRef] [PubMed]
- Feigenspan, A.; Janssen-Bienhold, U.; Hormuzdi, S.; Monyer, H.; Degen, J.; Sohl, G.; Willecke, K.; Ammermuller, J.; Weiler, R. Expression of Connexin36 in Cone Pedicles and OFF-Cone Bipolar Cells of the Mouse Retina. J. Neurosci. 2004, 24, 3325–3334. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Han, J.W.; Kim, H.J.; Kim, I.B.; Lee, M.Y.; Oh, S.J.; Chung, J.W.; Chun, M.H. The Immunocytochemical Localization of Connexin 36 at Rod and Cone Gap Junctions in the Guinea Pig Retina. Eur J Neurosci 2003, 18, 2925–2934. [Google Scholar] [CrossRef]
- Asteriti, S.; Gargini, C.; Cangiano, L. Connexin 36 Expression Is Required for Electrical Coupling between Mouse Rods and Cones. Vis. Neurosci. 2017, 34, E006. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.G.; Freed, M.A.; Sterling, P. Microcircuitry of the Dark-Adapted Cat Retina: Functional Architecture of the Rod-Cone Network. J. Neurosci. 1986, 6, 3505–3517. [Google Scholar] [CrossRef] [PubMed]
- Kovács-Öller, T.; Debertin, G.; Balogh, M.; Ganczer, A.; Orbán, J.; Nyitrai, M.; Balogh, L.; Kántor, O.; Völgyi, B. Connexin36 Expression in the Mammalian Retina: A Multiple-Species Comparison. Front. Cell. Neurosci. 2017, 11, 65. [Google Scholar] [CrossRef] [Green Version]
- Mills, S.L.; Massey, S.C. Differential Properties of Two Gap Junctional Pathways Made by AII Amacrine Cells. Nature 1995, 377, 734–737. [Google Scholar] [CrossRef]
- Bloomfield, S.A.; Xin, D.; Osborne, T. Light-Induced Modulation of Coupling between AII Amacrine Cells in the Rabbit Retina. Vis. Neurosci. 1997, 14, 565–576. [Google Scholar] [CrossRef]
- Massey, S.C.; Mills, S.L. Gap Junctions between AII Amacrine Cells and Calbindin-Positive Bipolar Cells in the Rabbit Retina. Vis. Neurosci. 1999, 16, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Bloomfield, S.A.; Dacheux, R.F. Rod Vision: Pathways and Processing in the Mammalian Retina. Prog. Retin. Eye Res. 2001, 20, 351–384. [Google Scholar] [CrossRef]
- Hartveit, E.; Veruki, M.L. Electrical Synapses between AII Amacrine Cells in the Retina: Function and Modulation. Brain Res. 2012, 1487, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Schubert, T.; Degen, J.; Willecke, K.; Hormuzdi, S.G.; Monyer, H.; Weiler, R. Connexin36 Mediates Gap Junctional Coupling of Alpha-Ganglion Cells in Mouse Retina. J. Comp. Neurol. 2005, 485, 191–201. [Google Scholar] [CrossRef]
- Völgyi, B.; Abrams, J.; Paul, D.L.; Bloomfield, S.A. Morphology and Tracer Coupling Pattern of Alpha Ganglion Cells in the Mouse Retina. J. Comp. Neurol. 2005, 492, 66–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Völgyi, B.; Chheda, S.; Bloomfield, S.A. Tracer Coupling Patterns of the Ganglion Cell Subtypes in the Mouse Retina. J. Comp. Neurol. 2009, 512, 664–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, F.; Paul, D.L.; Bloomfield, S.A.; Völgyi, B. Connexin36 Is Required for Gap Junctional Coupling of Most Ganglion Cell Subtypes in the Mouse Retina. J. Comp. Neurol. 2010, 518, 911–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, S.L. Unusual Coupling Patterns of a Cone Bipolar Cell in the Rabbit Retina. Vis. Neurosci. 1999, 16, 1029–1035. [Google Scholar] [CrossRef]
- Kántor, O.; Varga, A.; Nitschke, R.; Naumann, A.; Énzsöly, A.; Lukáts, A.; Szabó, A.; Németh, J.; Völgyi, B. Bipolar Cell Gap Junctions Serve Major Signaling Pathways in the Human Retina. Brain Struct Funct 2017, 222, 2603–2624. [Google Scholar] [CrossRef] [PubMed]
- Manookin, M.B.; Patterson, S.S.; Linehan, C.M. Neural Mechanisms Mediating Motion Sensitivity in Parasol Ganglion Cells of the Primate Retina. Neuron 2018, 97, 1327–1340.e4. [Google Scholar] [CrossRef] [Green Version]
- Sigulinsky, C.L.; Anderson, J.R.; Kerzner, E.; Rapp, C.N.; Pfeiffer, R.L.; Rodman, T.M.; Emrich, D.P.; Rapp, K.D.; Nelson, N.T.; Lauritzen, J.S.; et al. Network Architecture of Gap Junctional Coupling among Parallel Processing Channels in the Mammalian Retina. J. Neurosci. 2020, 40, 4483–4511. [Google Scholar] [CrossRef]
- Xin, D.; Bloomfield, S.A. Tracer Coupling Pattern of Amacrine and Ganglion Cells in the Rabbit Retina. J. Comp. Neurol. 1997, 383, 512–528. [Google Scholar] [CrossRef]
- Völgyi, B.; Xin, D.; Amarillo, Y.; Bloomfield, S.A. Morphology and Physiology of the Polyaxonal Amacrine Cells in the Rabbit Retina. J. Comp. Neurol. 2001, 440, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, J.; Massey, S.C. Coupling Pattern of S1 and S2 Amacrine Cells in the Rabbit Retina. Vis. Neurosci. 2002, 19, 119–131. [Google Scholar] [CrossRef]
- Bloomfield, S.A.; Völgyi, B. Response Properties of a Unique Subtype of Wide-Field Amacrine Cell in the Rabbit Retina. Vis. Neurosci. 2007, 24, 459–469. [Google Scholar] [CrossRef]
- Jacoby, J.; Nath, A.; Jessen, Z.F.; Schwartz, G.W. A Self-Regulating Gap Junction Network of Amacrine Cells Controls Nitric Oxide Release in the Retina. Neuron 2018, 100, 1149–1162.e5. [Google Scholar] [CrossRef] [Green Version]
- Telkes, I.; Kóbor, P.; Orbán, J.; Kovács-Öller, T.; Völgyi, B.; Buzás, P. Connexin-36 Distribution and Layer-Specific Topography in the Cat Retina. Brain Struct. Funct. 2019, 224, 2183–2197. [Google Scholar] [CrossRef] [Green Version]
- Bishop, P.O.; Kozak, W.; Vakkur, G.J. Some Quantitative Aspects of the Cat’s Eye: Axis and Plane of Reference, Visual Field Co-Ordinates and Optics. J. Physiol. 1962, 163, 466–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, A. A Quantitative Analysis of the Cat Retinal Ganglion Cell Topography. J Comp Neurol 1975, 163, 107–128. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Dyer, M.A.; Livesey, F.J.; Cepko, C.L.; Oliver, G. Prox1 Function Controls Progenitor Cell Proliferation and Horizontal Cell Genesis in the Mammalian Retina. Nat. Genet. 2003, 34, 53–58. [Google Scholar] [CrossRef]
- Pérez De Sevilla Müller, L.; Azar, S.S.; de los Santos, J.; Brecha, N.C. Prox1 Is a Marker for Aii Amacrine Cells in the Mouse Retina. Front. Neuroanat. 2017, 11, 1–12. [Google Scholar] [CrossRef]
- Röhrenbeck, J.; Wässle, H.; Heizmann, C.W. Immunocytochemical Labelling of Horizontal Cells in Mammalian Retina Using Antibodies against Calcium-Binding Proteins. Neurosci Lett 1987, 77, 255–260. [Google Scholar] [CrossRef]
- Gábriel, R.; Straznicky, C. Immunocytochemical Localization of Parvalbumin- and Neurofilament Triplet Protein Immunoreactivity in the Cat Retina: Colocalization in a Subpopulation of AII Amacrine Cells. Brain Res. 1992, 595, 133–136. [Google Scholar] [CrossRef]
- Peichl, L.; González-Soriano, J. Morphological Types of Horizontal Cell in Rodent Retinae: A Comparison of Rat, Mouse, Gerbil, and Guinea Pig. Vis. Neurosci. 1994, 11, 501–517. [Google Scholar] [CrossRef]
- Pasteels, B.; Rogers, J.; Blachier, F.; Pochet, R. Calbindin and Calretinin Localization in Retina from Different Species. Vis. Neurosci. 1990, 5, 1–16. [Google Scholar] [CrossRef]
- Wässle, H.; Grünert, U.; Röhrenbeck, J. Immunocytochemical Staining of AII-Amacrine Cells in the Rat Retina with Antibodies against Parvalbumin. J Comp Neurol 1993, 332, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Goebel, D.J.; Pourcho, R.G. Calretinin in the Cat Retina: Colocalizations with Other Calcium-Binding Proteins, GABA and Glycine. Vis. Neurosci. 1997, 14, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Kolb, H.; Nelson, R.; Mariani, A. Amacrine Cells, Bipolar Cells and Ganglion Cells of the Cat Retina: A Golgi Study. Vision Res. 1981, 21, 1081–1114. [Google Scholar] [CrossRef]
- Jeon, M.H.; Jeon, C.J. Immunocytochemical Localization of Calretinin Containing Neurons in Retina from Rabbit, Cat, and Dog. Neurosci Res 1998, 32, 75–84. [Google Scholar] [CrossRef]
- Gábriel, R.; Witkovsky, P. Cholinergic, but Not the Rod Pathway-Related Glycinergic (AII), Amacrine Cells Contain Calretinin in the Rat Retina. Neurosci. Lett. 1998, 247, 179–182. [Google Scholar] [CrossRef]
- Hwang, I.K.; Yoo, K.Y.; Kim, D.S.; Jung, J.Y.; Shin, M.C.; Seo, K.; Kim, K.S.; Kang, T.C.; Won, M.H. Comparative Study on Calretinin Immunoreactivity in Gerbil and Rat Retina. Anat Histol Embryol 2005, 34, 129–131. [Google Scholar] [CrossRef]
- Kovács-Öller, T.; Szarka, G.; Ganczer, A.; Tengölics, Á.; Balogh, B.; Völgyi, B. Expression of Ca2+-Binding Buffer Proteins in the Human and Mouse Retinal Neurons. Int. J. Mol. Sci. 2019, 20, 2229. [Google Scholar] [CrossRef] [Green Version]
- Haverkamp, S.; Wässle, H. Immunocytochemical Analysis of the Mouse Retina. J. Comp. Neurol. 2000, 424, 1–23. [Google Scholar] [CrossRef]
- Kolb, H.; Famiglietti, E.V. Rod and Cone Pathways in the Inner Plexiform Layer of Cat Retina. Science 1974, 186, 47–49. [Google Scholar] [CrossRef] [Green Version]
- Famiglietti, E.V.; Kolb, H. A Bistratified Amacrine Cell and Synaptic Circuitry in the Inner Plexiform Layer of the Retina. Brain Res. 1975, 84, 293–300. [Google Scholar] [CrossRef]
- Nelson, R. AII Amacrine Cells Quicken Time Course of Rod Signals in the Cat Retina. J. Neurophysiol. 1982, 47, 928–947. [Google Scholar] [CrossRef]
- Kovács-Öller, T.; Raics, K.; Orbán, J.; Nyitrai, M.; Völgyi, B. Developmental Changes in the Expression Level of Connexin36 in the Rat Retina. Cell Tissue Res. 2014, 358, 289–302. [Google Scholar] [CrossRef]
- Stone, J. A Quantitative Analysis of the Distribution of Ganglion Cells in the Cat’s Retina. J. Comp. Neurol. 1965, 124, 337–352. [Google Scholar] [CrossRef]
- Dräger, U.C.; Olsen, J.F. Ganglion Cell Distribution in the Retina of the Mouse. Investig. Ophthalmol. Vis. Sci. 1981, 20, 285–293. [Google Scholar]
- Salinas-Navarro, M.; Jiménez-López, M.; Valiente-Soriano, F.J.; Alarcón-Martínez, L.; Avilés-Trigueros, M.; Mayor, S.; Holmes, T.; Lund, R.D.; Villegas-Pérez, M.P.; Vidal-Sanz, M. Retinal Ganglion Cell Population in Adult Albino and Pigmented Mice: A Computerized Analysis of the Entire Population and Its Spatial Distribution. Vision Res. 2009, 49, 637–647. [Google Scholar] [CrossRef] [Green Version]
- Salinas-Navarro, M.; Mayor-Torroglosa, S.; Jiménez-López, M.; Avilés-Trigueros, M.; Holmes, T.M.; Lund, R.D.; Villegas-Pérez, M.P.; Vidal-Sanz, M. A Computerized Analysis of the Entire Retinal Ganglion Cell Population and Its Spatial Distribution in Adult Rats. Vision Res. 2009, 49, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Ortín-Martínez, A.; Jiménez-López, M.; Nadal-Nicolás, F.M.; Salinas-Navarro, M.; Alarcón-Martínez, L.; Sauvé, Y.; Villegas-Pérez, M.P.; Vidal-Sanz, M.; Agudo-Barriuso, M. Automated Quantification and Topographical Distribution of the Whole Population of S- and L-Cones in Adult Albino and Pigmented Rats. Investig. Opthalmology Vis. Sci. 2010, 51, 3171. [Google Scholar] [CrossRef] [PubMed]
- Voigt, T. Cholinergic Amacrine Cells in the Rat Retina. J. Comp. Neurol. 1986, 248, 19–35. [Google Scholar] [CrossRef]
- Oliver, G.; Sosa-Pineda, B.; Geisendorf, S.; Spana, E.P.; Doe, C.Q.; Gruss, P. Prox 1, a Prospero-Related Homeobox Gene Expressed during Mouse Development. Mech. Dev. 1993, 44, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Lavado, A.; Oliver, G. Prox1 Expression Patterns in the Developing and Adult Murine Brain. Dev. Dyn. 2007, 236, 518–524. [Google Scholar] [CrossRef]
- Vaney, D.I. The Morphology and Topographic Distribution of AII Amacrine Cells in the Cat Retina. Proc. R. Soc. London - Biol. Sci. 1985, 224, 475–488. [Google Scholar] [CrossRef]
- Cook, T. Cell Diversity in the Retina: More than Meets the Eye. BioEssays 2003, 25, 921–925. [Google Scholar] [CrossRef]
- Boycott, B.B.; Peichl, L.; Wassle, H. Morphological Types of Horizontal Cell in the Retina of the Domestic Cat. Proc. R. Soc. London - Biol. Sci. 1978, 203, 229–245. [Google Scholar] [CrossRef]
- Röhrenbeck, J.; Wässle, H.; Boycott, B.B. Horizontal Cells in the Monkey Retina: Immunocytochemical Staining with Antibodies against Calcium Binding Proteins. Eur J Neurosci 1989, 1, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Fyk-Kolodziej, B.; Qin, P.; Dzhagaryan, A.; Pourcho, R.G. Differential Cellular and Subcellular Distribution of Glutamate Transporters in the Cat Retina. Vis. Neurosci. 2004, 21, 551–565. [Google Scholar] [CrossRef]
- Lyser, K.M.; Li, A.I.; Nuñez, M. Horizontal Cells in the Rabbit Retina: Differentiation of Subtypes at Neonatal and Postnatal Stages. Int. J. Dev. Neurosci. 1994, 12, 673–682. [Google Scholar] [CrossRef]
- Sotelo, C.; Llinás, R. Specialized Membrane Junctions between Neurons in the Vertebrate Cerebellar Cortex. J. Cell Biol. 1972, 53, 271–289. [Google Scholar] [CrossRef] [Green Version]
- Galarreta, M.; Hestrin, S. Electrical Synapses between Gaba-Releasing Interneurons. Nat. Rev. Neurosci. 2001, 2, 425–433. [Google Scholar] [CrossRef]
- Hinrichsen, C.F.L.; Larramendi, L.M.H. Synapses and Cluster Formation of the Mouse Mesencephalic Fifth Nucleus. Brain Res. 1968, 7, 296–299. [Google Scholar] [CrossRef]
- Pedroarena, C.M.; Pose, I.E.; Yamuy, J.; Chase, M.H.; Morales, F.R. Oscillatory Membrane Potential Activity in the Soma of a Primary Afferent Neuron. J. Neurophysiol. 1999, 82, 1465–1476. [Google Scholar] [CrossRef]
- Curti, S.; Hoge, G.; Nagy, J.I.; Pereda, A.E. Synergy between Electrical Coupling and Membrane Properties Promotes Strong Synchronization of Neurons of the Mesencephalic Trigeminal Nucleus. J. Neurosci. 2012, 32, 4341–4359. [Google Scholar] [CrossRef]
- Connors, B.W.; Long, M.A. Electrical Synapses in the Mammalian Brain. Annu. Rev. Neurosci. 2004, 27, 393–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smedowski, A.; Akhtar, S.; Liu, X.; Pietrucha-Dutczak, M.; Podracka, L.; Toropainen, E.; Alkanaan, A.; Ruponen, M.; Urtti, A.; Varjosalo, M.; et al. Electrical Synapses Interconnecting Axons Revealed in the Optic Nerve Head – a Novel Model of Gap Junctions’ Involvement in Optic Nerve Function. Acta Ophthalmol. 2020, 98, 408–417. [Google Scholar] [CrossRef] [Green Version]
- Vardi, N.; Smith, R.G. The AII Amacrine Network: Coupling Can Increase Correlated Activity. Vis. Res 1996, 36, 3743–3757. [Google Scholar] [CrossRef] [Green Version]
- Zandt, B.-J.; Liu, J.H.; Veruki, M.L.; Hartveit, E. AII Amacrine Cells: Quantitative Reconstruction and Morphometric Analysis of Electrophysiologically Identified Cells in Live Rat Retinal Slices Imaged with Multi-Photon Excitation Microscopy. Brain Struct. Funct. 2017, 222, 151–182. [Google Scholar] [CrossRef] [Green Version]
- Sterling, P. Microcircuitry of the Cat Retina. Annu. Rev. Neurosci. 1983, 6, 149–185. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, Y.; Omi, N. Functional Allocation of Synaptic Contacts in Microcircuits from Rods via Rod Bipolar to AII Amacrine Cells in the Mouse Retina. J. Comp. Neurol. 2013, 521, 3541–3555. [Google Scholar] [CrossRef] [Green Version]
- Bloomfield, S.A.; Völgyi, B. Function and Plasticity of Homologous Coupling between AII Amacrine Cells. Vision Res. 2004, 44, 3297–3306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Song, J.-H.; Denisova, J.V.; Park, W.-M.; Fontes, J.D.; Belousov, A.B. Neuronal Gap Junction Coupling Is Regulated by Glutamate and Plays Critical Role in Cell Death during Neuronal Injury. J. Neurosci. 2012, 32, 713–725. [Google Scholar] [CrossRef] [Green Version]
- Szarka, G.; Balogh, M.; Tengölics, Á.; Ganczer, A.; Völgyi, B.; Kovács-Öller, T. The Role of Gap Junctions in Cell Death and Neuromodulation in the Retina. Neural Regen. Res. 2021, 16, 1911. [Google Scholar] [CrossRef]
- Wang, H.Y.; Lin, Y.-P.P.; Mitchell, C.K.; Ram, S.; O’Brien, J. Two-Color Fluorescent Analysis of Connexin 36 Turnover: Relationship to Functional Plasticity. J Cell Sci 2015, 128, 3888–3897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hervé, J.-C.; Derangeon, M.; Bahbouhi, B.; Mesnil, M.; Sarrouilhe, D. The Connexin Turnover, an Important Modulating Factor of the Level of Cell-to-Cell Junctional Communication: Comparison with Other Integral Membrane Proteins. J. Membr. Biol. 2007 2171 2007, 217, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Feigenspan, A.; Teubner, B.; Willecke, K.; Weiler, R. Expression of Neuronal Connexin36 in AII Amacrine Cells of the Mammalian Retina. J. Neurosci. 2001, 21, 230–239. [Google Scholar] [CrossRef]
- Veruki, M.L.; Hartveit, E. AII (Rod) Amacrine Cells Form a Network of Electrically Coupled Interneurons in the Mammalian Retina. Neuron 2002, 33, 935–946. [Google Scholar] [CrossRef] [Green Version]
- Vaney, D.I. Many Diverse Types of Retinal Neurons Show Tracer Coupling When Injected with Biocytin or Neurobiotin. Neurosci. Lett. 1991, 125, 187–190. [Google Scholar] [CrossRef]
- Strettoi, E.; Raviola, E.; Dacheux, R.F. Synaptic Connections of the Narrow-Field, Bistratified Rod Amacrine Cell (AII) in the Rabbit Retina. J Comp Neurol 1992, 325, 152–168. [Google Scholar] [CrossRef]
- Zandt, B.-J.; Veruki, M.L.; Hartveit, E. Electrotonic Signal Processing in AII Amacrine Cells: Compartmental Models and Passive Membrane Properties for a Gap Junction-Coupled Retinal Neuron. Brain Struct. Funct. 2018, 223, 3383–3410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, S.L.; O’Brien, J.J.; Li, W.; O’Brien, J.; Massey, S.C. Rod Pathways in the Mammalian Retina Use Connexin. J. Comp. Neurol. 2001, 436, 336–350. [Google Scholar] [CrossRef]
- Sterling, P.; Freed, M.A.; Smith, R.G. Architecture of Rod and Cone Circuits to the On-Beta Ganglion Cell. J. Neurosci. 1988, 8, 623–642. [Google Scholar] [CrossRef]
- Marc, R.E.; Anderson, J.R.; Jones, B.W.; Sigulinsky, C.L.; Lauritzen, J.S. The AII Amacrine Cell Connectome: A Dense Network Hub. Front Neural Circuits 2014, 8, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, A.; Hilgen, G.; Dorgau, B.; Sammler, E.M.; Weiler, R.; Monyer, H.; Dedek, K.; Hormuzdi, S.G. AII Amacrine Cells Discriminate between Heterocellular and Homocellular Locations When Assembling Connexin36-Containing Gap Junctions. J. Cell Sci. 2014, 127, 1190–1202. [Google Scholar] [CrossRef] [Green Version]
- Macneil, M.A.; Purrier, S.; Rushmore, R.J. The Composition of the Inner Nuclear Layer of the Cat Retina. Vis. Neurosci. 2009, 26, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Teeter, C.M.; Stevens, C.F. A General Principle of Neural Arbor Branch Density. Curr. Biol. 2011, 21, 2105–2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kier, C.; Buchsbaum, G.; Sterling, P. How Retinal Microcircuits Scale for Ganglion Cells of Different Size. J. Neurosci. 1995, 15, 7673–7683. [Google Scholar] [CrossRef]
- Freed, M.A.; Smith, R.G.; Sterling, P. Computational Model of the On-Alpha Ganglion Cell Receptive Field Based on Bipolar Cell Circuitry. Proc. Natl. Acad. Sci. USA 1992, 89, 236–240. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Vasudeva, V.; Vardi, N.; Sterling, P.; Freed, M.A. Different Types of Ganglion Cell Share a Synaptic Pattern. J. Comp. Neurol. 2008, 507, 1871–1878. [Google Scholar] [CrossRef]
- Percival, K.A.; Jusuf, P.R.; Martin, P.R.; Grünert, U. Synaptic Inputs onto Small Bistratified (Blue-ON/Yellow-OFF) Ganglion Cells in Marmoset Retina. J Comp Neurol 2009, 517, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Fournel, R.; Hartveit, E.; Veruki, M.L. Differential Contribution of Gap Junctions to the Membrane Properties of ON- and OFF-Bipolar Cells of the Rat Retina. Cell. Mol. Neurobiol. 2021, 41, 229–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wässle, H. Parallel Processing in the Mammalian Retina. Nat. Rev. Neurosci. 2004, 5, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Chichilnisky, E.J.; Kalmar, R.S. Functional Asymmetries in ON and OFF Ganglion Cells of Primate Retina. J. Neurosci. 2002, 22, 2737–2747. [Google Scholar] [CrossRef] [PubMed]
- Pandarinath, C.; Victor, J.D.; Nirenberg, S. Symmetry Breakdown in the ON and OFF Pathways of the Retina at Night: Functional Implications. J. Neurosci. 2010, 30, 10006–10014. [Google Scholar] [CrossRef] [Green Version]
- Ravi, S.; Ahn, D.; Greschner, M.; Chichilnisky, E.J.; Field, G.D. Pathway-Specific Asymmetries between ON and OFF Visual Signals. J. Neurosci. 2018, 38, 9728–9740. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Wang, Y.; Lashgari, R.; Swadlow, H.A.; Alonso, J.M. Faster Thalamocortical Processing for Dark than Light Visual Targets. J. Neurosci. 2011, 31, 17471–17479. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Purushothaman, G.; Casagrande, V.A. The Functional Asymmetry of ON and OFF Channels in the Perception of Contrast. J. Neurophysiol. 2015, 114, 2816–2829. [Google Scholar] [CrossRef]
- Escobar, M.J.; Reyes, C.; Herzog, R.; Araya, J.; Otero, M.; Ibaceta, C.; Palacios, A.G. Characterization of Retinal Functionality at Different Eccentricities in a Diurnal Rodent. Front. Cell. Neurosci. 2018, 12, 444. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent or Resource | Source | Identifier |
|---|---|---|
| Antibodies | ||
| rabbit polyclonal anti-calretinin | Merck Hungary, Budapest, Hungary | AB 5054 |
| mouse monoclonal anti-connexin-36 | Merck Hungary, Budapest, Hungary | MAB 3045 |
| rabbit polyclonal anti-Prox1 | Merck Hungary, Budapest, Hungary | ABN 278 |
| mouse monoclonal anti-parvalbumin | Swant, Burgdorf, Switzerland | PV 235 |
| mouse monoclonal anti-calbindin | Synaptic Systems, Göttingen, German | 214011 |
| biotinylated anti-mouse IgG (H + L) | Vector Laboratories, Burlingame, CA, USA | BA 2001 |
| streptavidin-Alexa Fluor 488 conjugate | Invitrogen, Waltham, MA, USA | S 32354 |
| donkey anti-mouse IgG (H + L) Alexa Fluor 488 conjugate | Jackson ImmunoResearch, West Grove, PA, USA | 715-545-151 |
| donkey anti-rabbit IgG (H + L) Alexa Fluor 594 conjugate | Jackson ImmunoResearch, West Grove, PA, USA | 711-585-152 |
| goat anti-rabbit IgG (H + L) Texas Red conjugate | Jackson ImmunoResearch, West Grove, PA, USA | 111-075-003 |
| Animals | ||
| domestic cat (Felis catus) | Animal house of the Institute of Physiology, Medical School, University of Pécs, Hungary | N/A |
| rat (Rattus norvegicus), Wistar strain | Animal house of the Institute of Physiology, Medical School, University of Pécs, Hungary | N/A |
| mouse (Mus musculus), C57BL/6J strain | The Jackson Laboratory, Bar Harbor, USA | 000664 |
| mouse (Mus musculus), PV-Cre line | The Jackson Laboratory, Bar Harbor, USA | B6.129P2-Pvalbtm1(cre)Arbr/J, JAX #017320 |
| mouse (Mus musculus), tdTomato line | The Jackson Laboratory, Bar Harbor, USA | B6.Cg-Gt(ROSA)26Sortm9(CAG-tdTomato)Hze/J, JAX #007909 |
| Equipment | ||
| Zeiss LSM 710 confocal laser scanning microscope | Carl Zeiss, Jena, Germany | |
| Software | ||
| Fiji | https://imagej.net/software/fiji/, accessed on 27 July 2018 | N/A |
| Imaris 8.12 | Oxford Instruments, Zurich, Switzerland | N/A |
| SPSS 24.0 | IBM Corporation, Armonk, NY, USA | N/A |
| Excel for Office 365 | Microsoft, Redmond, WA, USA | N/A |
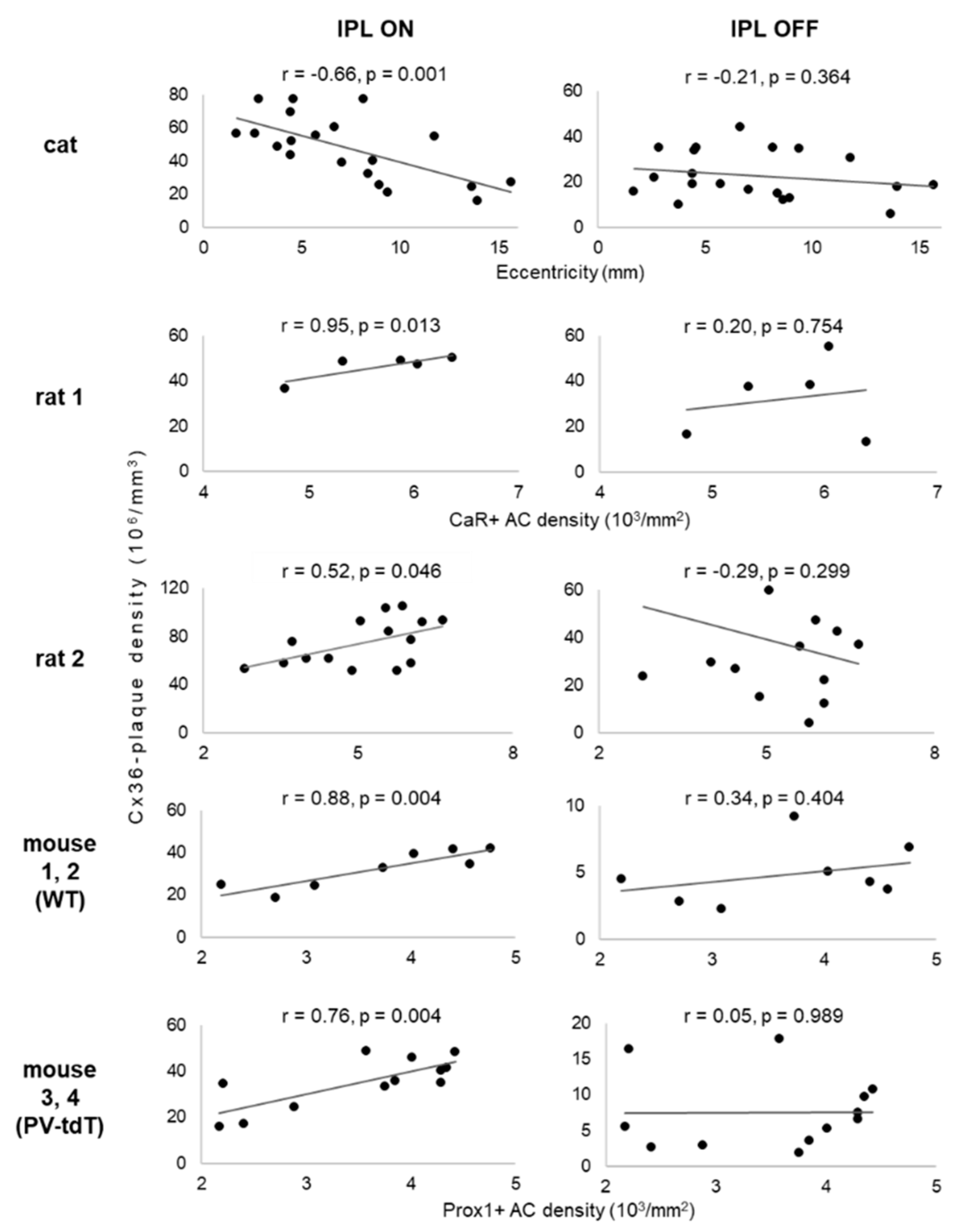
| Ample (Number of Retinal Locations) | Cat (n = 20) | Rat 1 (n = 5) | Rat 2 (n = 15) | Mouse 1 and 2, Wild Type (n = 8) | Mouse 3 and 4, PV-tdT (n = 12) | ||
|---|---|---|---|---|---|---|---|
| Marker used for measurement of cell density | none | CaR | Prox1 | Prox1 | Prox1 | ||
| Measure of retinal position | Eccentricity (mm) | Amacrine cell density (mm−2) | |||||
| Mean ± SD | 7.34 ± 4.01 | 5766 ± 598 | 5085 ± 1139 | 3687 ± 904 | 3520 ± 836 | ||
| IPL sublamina | OFF | Cx36-density (1/mm–3) | 22.8 × 106 ±10.5 × 106 | 32.3 × 106 ±17.4 × 106 | 38.7 × 106 ±24.5 × 106 | 4.84 × 106 ±2.29 × 106 | 7.55 × 106 ±5.53 × 106 |
| Correlation with retinal position | r = −0.21 p = 0.364 | r = 0.20 p = 0.754 | r = −0.29 p = 0.299 | r = 0.34 p = 0.404 | r = 0.05 p = 0.989 | ||
| ON | Cx36-density (mm–3) | 47.7 × 106 ±11.8 × 106 | 46.2 × 106 ±5.55 × 106 | 74.3 × 106 ±19.4 × 106 | 32.3 × 106 ±13.2 × 106 | 35.0 × 106 ±15.5 × 106 | |
| Correlation with retinal position | r = −0.66 p = 0.001 | r = 0.95 p = 0.013 | r = 0.52 p = 0.046 | r = 0.88 p = 0.004 | r = 0.76 p = 0.004 | ||
| Difference of Cx36-density between IPL OFF and IPL ON | t = −10.05 p < 0.001 | t = −2.44 p = 0.024 | t = −5.62 p < 0.001 | t = −47.95 p < 0.001 | t = −24.33 p < 0.001 | ||
| Correlation of Cx36-density between IPL OFF and IPL ON | r = 0.11 p = 0.325 | r = 0.33 p = 0.587 | r = 0.42 p = 0.121 | r = 0.54 p = 0.463 | r = 0.77 p = 0.075 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fusz, K.; Kovács-Öller, T.; Kóbor, P.; Szabó-Meleg, E.; Völgyi, B.; Buzás, P.; Telkes, I. Regional Variation of Gap Junctional Connections in the Mammalian Inner Retina. Cells 2021, 10, 2396. https://doi.org/10.3390/cells10092396
Fusz K, Kovács-Öller T, Kóbor P, Szabó-Meleg E, Völgyi B, Buzás P, Telkes I. Regional Variation of Gap Junctional Connections in the Mammalian Inner Retina. Cells. 2021; 10(9):2396. https://doi.org/10.3390/cells10092396
Chicago/Turabian StyleFusz, Katalin, Tamás Kovács-Öller, Péter Kóbor, Edina Szabó-Meleg, Béla Völgyi, Péter Buzás, and Ildikó Telkes. 2021. "Regional Variation of Gap Junctional Connections in the Mammalian Inner Retina" Cells 10, no. 9: 2396. https://doi.org/10.3390/cells10092396
APA StyleFusz, K., Kovács-Öller, T., Kóbor, P., Szabó-Meleg, E., Völgyi, B., Buzás, P., & Telkes, I. (2021). Regional Variation of Gap Junctional Connections in the Mammalian Inner Retina. Cells, 10(9), 2396. https://doi.org/10.3390/cells10092396