
Epidermal Growth Factor Receptor Expression in the Corneal Epithelium

Abstract
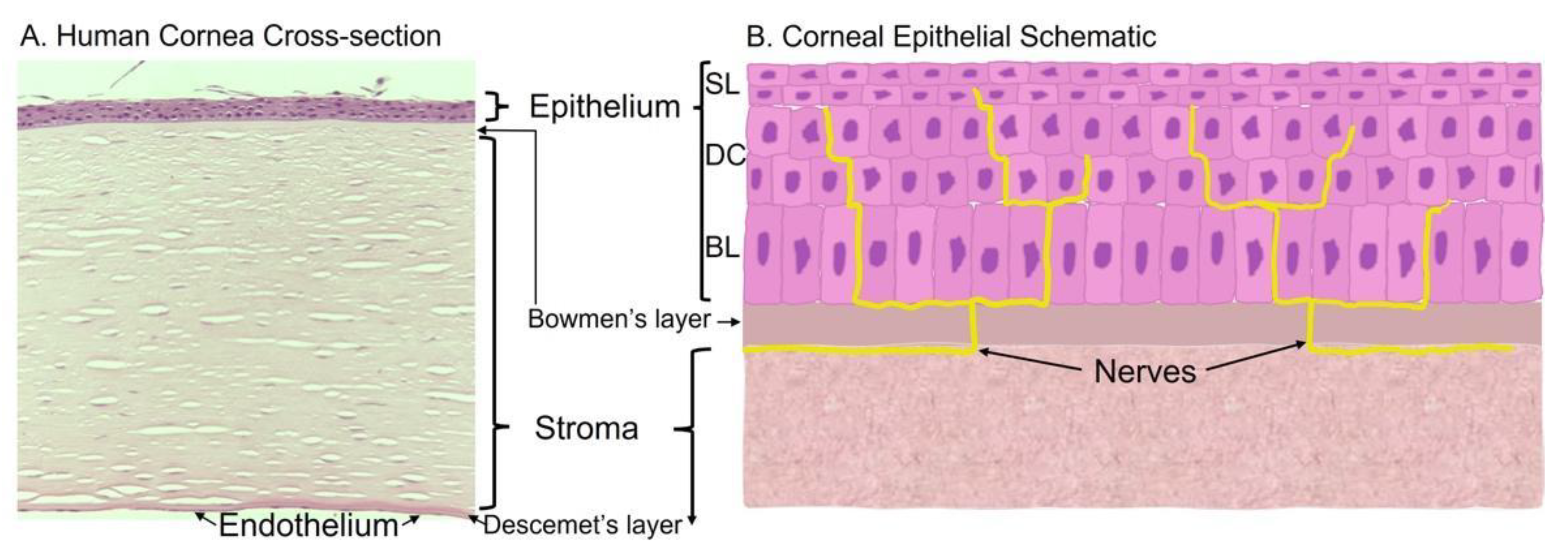
:1. Cornea Structure and Function
1.1. Corneal Epithelium
1.2. Stroma
1.3. Endothelium
1.4. Bowman and Descemet’s Layers
2. Corneal Epithelial Perturbations
2.1. Trauma
2.2. Infections
2.3. Corneal Diseases
3. Regulation of the Corneal Epithelial Homeostasis
3.1. EGFR Expression in Corneal Epithelial Cells
3.2. Ocular Expression of EGFR Ligands
4. Physiologic Role of the EGFR and ErbB Family in Corneal Epithelial Homeostasis
4.1. EGFR in the Corneal Epithelium
4.2. ErbB Family Members in the Corneal Epithelium
4.3. Specific Effects of EGFR Ligands on the Corneal Epithelial Cells
4.4. Negative Consequences of Sustained EGFR Activity
5. Pharmacological Approaches to Promote Corneal Epithelial Homeostasis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wilson, S.E. Bowman’s layer in the cornea—Structure and function and regeneration. Exp. Eye Res. 2020, 195, 108033. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, R.C.; Wilson, S.E. Descemet’s membrane development, structure, function and regeneration. Exp. Eye Res. 2020, 197, 108090. [Google Scholar] [CrossRef]
- Scott, A. Scott’s Anatomy of the Eye and Orbit; Ridgevue Publishing LLC: Boulder, CO, USA, 2016. [Google Scholar]
- Haddad, A. Renewal of the rabbit corneal epithelium as investigated by autoradiography after intravitreal injection of 3H-thymidine. Cornea 2000, 19, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Eghrari, A.O.; Riazuddin, S.A.; Gottsch, J.D. Overview of the cornea: Structure, function, and development. Mol. Biol. Eye Dis. 2015, 134, 7–23. [Google Scholar]
- Mantelli, F.; Mauris, J.; Argueso, P. The ocular surface epithelial barrier and other mechanisms of mucosal protection: From allergy to infectious diseases. Curr. Opin. Allergy Clin. Immunol. 2013, 13, 563–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugrue, S.P.; Zieske, J. ZO1 in corneal epithelium: Association to the zonula occludens and adherens junctions. Exp. Eye Res. 1997, 64, 11–20. [Google Scholar] [CrossRef]
- Patel, S.; McLaren, J.; Hodge, D.; Bourne, W. Normal human keratocyte density and corneal thickness measurement by using confocal microscopy in vivo. Investig. Ophthalmol. Vis. Sci. 2001, 42, 333–339. [Google Scholar]
- Reinstein, D.Z.; Archer, T.J.; Gobbe, M.; Silverman, R.H.; Coleman, D.J. Stromal thickness in the normal cornea: Three-dimensional display with artemis very high-frequency digital ultrasound. J. Refract. Surg. 2009, 25, 776–786. [Google Scholar] [CrossRef] [Green Version]
- Espana, E.M.; Birk, D.E. Composition, structure and function of the corneal stroma. Exp. Eye Res. 2020, 198, 108137. [Google Scholar] [CrossRef]
- Newsome, D.A.; Gross, J.; Hassell, J.R. Human corneal stroma contains 3 distinct collagens. Investig. Ophthalmol. Vis. Sci. 1982, 22, 376–381. [Google Scholar]
- Rózsa, A.J.; Beuerman, R.W. Density and organization of free nerve endings in the corneal epithelium of the rabbit. Pain 1982, 14, 105–120. [Google Scholar] [CrossRef]
- Robertson, D.M.; Winkelmann, R.K. A whole-mount cholinesterase technique for demonstrating corneal nerves: Observations in the albino rabbit. Investig. Ophthalmol. 1970, 9, 710–715. [Google Scholar]
- Tervo, T.; Palkama, A. Innervation of the rabbit cornea. A histochemical and electron-microscopic study. Acta Anat. 1978, 102, 164–175. [Google Scholar] [CrossRef]
- Marfurt, C.F.; Cox, J.; Deek, S.; Dvorscak, L. Anatomy of the human corneal innervation. Exp. Eye Res. 2010, 90, 478–492. [Google Scholar] [CrossRef]
- Goyal, S.; Hamrah, P. Understanding neuropathic corneal pain—Gaps and current therapeutic approaches. Semin. Ophthalmol. 2016, 31, 59–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klyce, S.D. Endothelial pump and barrier function. Exp. Eye Res. 2020, 198, 108068. [Google Scholar] [CrossRef] [PubMed]
- Maurice, D.M. The location of the fluid pump in the cornea. J. Physiol. 1972, 221, 43–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meek, K.M.; Leonard, D.W.; Connon, C.J.; Dennis, S.; Khan, S. Transparency, swelling and scarring in the corneal stroma. Eye 2003, 17, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Ong Tone, S.; Kocaba, V.; Böhm, M.; Wylegala, A.; White, T.; Jurkunas, U.V. Fuchs endothelial corneal dystrophy: The vicious cycle of Fuchs pathogenesis. Prog. Retin. Eye Res. 2020, 80, 100863. [Google Scholar] [CrossRef] [PubMed]
- Dewitt, E.N. The Histopathology of Bowman’s Membrane. Trans. Am. Ophthalmol. Soc. 1931, 29, 461–485. [Google Scholar]
- Germundsson, J.; Karanis, G.; Fagerholm, P.; Lagali, N. Age-related thinning of Bowman’s layer in the human cornea in vivo. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6143–6149. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, S.; Osawa, T.; Tohyama, K. Comparative observations on corneas, with special reference to Bowman’s layer and Descemet’s membrane in mammals and amphibians. J. Morphol. 2002, 254, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E.; He, Y.G.; Weng, J.; Li, Q.; McDowall, A.W.; Vital, M.; Chwang, E.L. Epithelial injury induces keratocyte apoptosis: Hypothesized role for the interleukin-1 system in the modulation of corneal tissue organization and wound healing. Exp. Eye Res. 1996, 62, 325–337. [Google Scholar] [CrossRef]
- Paralkar, V.M.; Vukicevic, S.; Reddi, A.H. Transforming growth-factor β type-1 binds to collagen IV of basement-membrane Matrix: Implications for development. Dev. Biol. 1991, 143, 303–308. [Google Scholar] [CrossRef]
- Göhring, W.; Sasaki, T.; Heldin, C.-H.; Timpl, R. Mapping of the binding of platelet-derived growth factor to distinct domains of the basement membrane proteins BM-40 and perlecan and distinction from the BM-40 collagen-binding epitope. Eur. J. Biochem. 1998, 255, 60–66. [Google Scholar] [CrossRef]
- Sakamoto, T.; Ueno, H.; Sonoda, K.; Hisatomi, T.; Shimizu, K.; Ohashi, H.; Inomata, H. Blockade of TGF-beta by in vivo gene transfer of a soluble TGF-beta type II receptor in the muscle inhibits corneal opacification, edema and angiogenesis. Gene Ther. 2000, 7, 1915–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepp, M.A.; Zieske, J.; Trinkaus-Randall, V.; Kyne, B.M.; Pal-Ghosh, S.; Tadvalkar, G.; Pajoohesh-Ganji, A. Wounding the cornea to learn how it heals. Exp. Eye Res. 2014, 121, 178–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, R.I.; Savitsky, E. Ocular trauma. In Combat Casualty Care—Lessons Learned from OEF and OIF; Savitsky, E., Eastridge, B., Eds.; Office of The Surgeon General: Fort Detrick, MD, USA, 2011; pp. 299–342. [Google Scholar]
- Negussie, D.; Bejiga, A. Ocular emergencies presenting to Menelik II Hospital. Ethiop. Med. J. 2011, 49, 17–24. [Google Scholar] [PubMed]
- Azar, D.T.; Spurr-Michaud, S.J.; Tisdale, A.S.; Gipson, I.K. Altered epithelial-basement membrane interactions in diabetic corneas. Arch. Ophthalmol. 1992, 110, 537–540. [Google Scholar] [CrossRef]
- Friend, J.; Ishii, Y.; Thoft, R.A. Corneal epithelial changes in diabetic rats. Ophthalmic Res. 1982, 14, 269–278. [Google Scholar] [CrossRef]
- Rosenberg, M.E.; Tervo, T.M.; Immonen, I.J.; Müller, L.J.; Grönhagen-Riska, C.; Vesaluoma, M.H. Corneal structure and sensitivity in type 1 diabetes mellitus. Investig. Ophthalmol. Vis. Sci. 2000, 41, 2915–2921. [Google Scholar]
- Taylor, H.R.; A Kimsey, R. Corneal epithelial basement membrane changes in diabetes. Investig. Ophthalmol. Vis. Sci. 1981, 20, 548–553. [Google Scholar]
- Johnson, K.S.; Levin, F.; Chu, D.S. Persistent corneal epithelial defect associated with erlotinib treatment. Cornea 2009, 28, 706–707. [Google Scholar] [CrossRef] [PubMed]
- Morishige, N.; Hatabe, N.; Morita, Y.; Yamada, N.; Kimura, K.; Sonoda, K.-H. Spontaneous healing of corneal perforation after temporary discontinuation of erlotinib treatment. Case Rep. Ophthalmol. 2014, 5, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Saint-Jean, A.; De la Maza, M.S.; Morral, M.; Torras, J.; Quintana, R.; Molina, J.J.; Molina-Prat, N. Ocular adverse events of systemic inhibitors of the epidermal growth factor receptor: Report of 5 cases. Ophthalmology 2012, 119, 1798–1802. [Google Scholar] [CrossRef] [PubMed]
- Liesegang, T.J. Herpes simplex virus epidemiology and ocular importance. Cornea 2001, 20, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Vrcek, I.; Choudhury, E.; Durairaj, V. Herpes zoster ophthalmicus: A review for the internist. Am. J. Med. 2016, 130, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Asbell, P.; Stenson, S. Ulcerative keratitis. Survey of 30 years’ laboratory experience. Arch. Ophthalmol. 1982, 100, 77–80. [Google Scholar] [CrossRef]
- Thomas, P.A. Fungal infections of the cornea. Eye 2003, 17, 852–862. [Google Scholar] [CrossRef]
- Marquart, M.E.; O’Callaghan, R.J. Infectious keratitis: Secreted bacterial proteins that mediate corneal damage. J. Ophthalmol. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lalitha, P.; Prajna, N.V.; Kabra, A.; Mahadevan, K.; Srinivasan, M. Risk factors for treatment outcome in fungal keratitis. Ophthalmology 2006, 113, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Voss, K.; Nguyen, A.; Heur, M. Non-infectious and non-hereditary diseases of the corneal epithelium. Exp. Eye Res. 2021, 202, 108316. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.-S.; Yin, J.; Xu, K.; Huang, J. Growth factors and corneal epithelial wound healing. Brain Res. Bull. 2010, 81, 229–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoppenreijs, V.P.; Pels, E.; Vrensen, G.F.; Felten, P.C.; Treffers, W.F. Platelet-derived growth factor: Receptor expression in corneas and effects on corneal cells. Investig. Ophthalmol. Vis. Sci. 1993, 34, 637–649. [Google Scholar]
- Wilson, S.E.; He, Y.G.; Weng, J.; Zieske, J.D.; Jester, J.V.; Schultz, G.S. Effect of epidermal growth factor, hepatocyte growth factor, and keratinocyte growth factor, on proliferation, motility and differentiation of human corneal epithelial cells. Exp. Eye Res. 1994, 59, 665–678. [Google Scholar] [CrossRef]
- Rho, C.R.; Park, M.Y.; Kang, S. Effects of granulocyte-macrophage colony-stimulating (GM-CSF) factor on corneal epithelial cells in corneal wound healing model. PLoS ONE 2015, 10, e0138020. [Google Scholar]
- Chandrasekher, G.; Kakazu, A.H.; Bazan, H.E. HGF- and KGF-induced activation of PI-3K/p70 s6 kinase pathway in corneal epithelial cells: Its relevance in wound healing. Exp. Eye Res. 2001, 73, 191–202. [Google Scholar] [CrossRef]
- Shin, E.; Lim, D.H.; Han, J.; Nam, D.-H.; Park, K.; Ahn, M.-J.; Kang, W.K.; Lee, J.; Ahn, J.S.; Lee, S.-H.; et al. Markedly increased ocular side effect causing severe vision deterioration after chemotherapy using new or investigational epidermal or fibroblast growth factor receptor inhibitors. BMC Ophthalmol. 2020, 20, 19. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Upadhya, D.; Lu, L.; Reneker, L.W. Fibroblast growth factor receptor 2 (FGFR2) is required for corneal epithelial cell proliferation and differentiation during embryonic development. PLoS ONE 2015, 10, e0117089. [Google Scholar] [CrossRef] [Green Version]
- Carrington, L.M.; Albon, J.; Anderson, I.; Kamma, C.; Boulton, M. Differential regulation of key stages in early corneal wound healing by TGF-beta isoforms and their inhibitors. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1886–1894. [Google Scholar] [CrossRef] [Green Version]
- Lambiase, A.; Manni, L.; Bonini, S.; Rama, P.; Micera, A.; Aloe, L. Nerve growth factor promotes corneal healing: Structural, biochemical, and molecular analyses of rat and human corneas. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1063–1069. [Google Scholar]
- Wilson, S.E.; Esposito, A. Focus on molecules: Interleukin-1: A master regulator of the corneal response to injury. Exp. Eye Res. 2009, 89, 124–125. [Google Scholar] [CrossRef] [Green Version]
- Nishida, T.; Nakamura, M.; Mishima, H.; Otori, T.; Hikida, M. Interleukin 6 facilitates corneal epithelial wound closure in vivo. Arch. Ophthalmol. 1992, 110, 1292–1294. [Google Scholar] [CrossRef]
- Peterson, J.L.; Phelps, E.D.; Doll, M.A.; Schaal, S.; Ceresa, B.P. The role of endogenous epidermal growth factor receptor ligands in mediating corneal epithelial homeostasis. Investig. Ophthalmol. Vis. Sci. 2014, 55, 2870–2880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zieske, J.; Takahashi, H.; E Hutcheon, A.; Dalbone, A.C. Activation of epidermal growth factor receptor during corneal epithelial migration. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1346–1355. [Google Scholar]
- Rush, J.S.; Bingaman, D.P.; Chaney, P.G.; Wax, M.B.; Ceresa, B.P. Administration of menadione, vitamin K3, ameliorates off-target effects on corneal epithelial wound healing due to receptor tyrosine kinase inhibition. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5864–5871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foerster, C.G.; Cursiefen, C.; E Kruse, F. Persisting corneal erosion under cetuximab (Erbitux) treatment (epidermal growth factor receptor antibody). Cornea 2008, 27, 612–614. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.; Dean, W.H.; Price, N.; Gomaa, A.; Ayre, G.; Guglani, S.; Sallam, A. Perforating corneal ulceration in a patient with lung metastatic adenocarcinoma treated with gefitinib: A case report. Case Rep. Ophthalmol. Med. 2012, 2012, 379132. [Google Scholar] [CrossRef]
- Kawakami, H.; Sugioka, K.; Yonesaka, K.; Satoh, T.; Shimomura, Y.; Nakagawa, K. Human epidermal growth factor eyedrops for cetuximab-related filamentary keratitis. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2011, 29, e678–e679. [Google Scholar] [CrossRef]
- McClintock, J.L.; Ceresa, B. Transforming growth factor-α (TGF-α) enhances corneal epithelial cell migration by promoting EGFR recycling. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3455–3461. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, J.L.; Pike, L.J. Heterogeneity in EGF-binding affinities arises from negative cooperativity in an aggregating system. Proc. Natl. Acad. Sci. USA 2008, 105, 112–117. [Google Scholar] [CrossRef] [Green Version]
- Lax, I.; Bellot, F.; Howk, R.; Ullrich, A.; Givol, D.; Schlessinger, J. Functional analysis of the ligand binding site of EGF-receptor utilizing chimeric chicken/human receptor molecules. EMBO J. 1989, 8, 421–427. [Google Scholar] [CrossRef]
- Lemmon, M.A. Ligand-induced ErbB receptor dimerization. Exp. Cell Res. 2009, 315, 638–648. [Google Scholar] [CrossRef] [Green Version]
- Pike, L.J. Negative co-operativity in the EGF receptor. Biochem. Soc. Trans. 2012, 40, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.W.; Hsu, J.M.; Xia, W.; Wang, H.L.; Wang, Y.N.; Chang, W.C.; Arold, S.T.; Chou, C.K.; Tsou, P.H.; Yamaguchi, H.; et al. PRMT1-mediated methylation of the EGF receptor regulates signaling and cetuximab response. J. Clin. Investig. 2015, 125, 4529–4543. [Google Scholar] [CrossRef] [Green Version]
- Ringerike, T.; Stang, E.; Johannessen, L.E.; Sandnes, D.; Levy, F.O.; Madshus, I.H. High-affinity binding of epidermal growth factor (EGF) to EGF receptor is disrupted by overexpression of mutant dynamin (K44A). J. Biol. Chem. 1998, 273, 16639–16642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rush, J.S.; Boeving, M.A.; Berry, W.L.; Ceresa, B.P. Antagonizing c-Cbl enhances EGFR-dependent corneal epithelial homeostasis. Investig. Ophthalmol. Vis. Sci. 2014, 55, 4691–4699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, D.M.; Li, L.; Fisher, S.; Pearce, V.P.; Shay, J.W.; Wright, W.E.; Cavanagh, H.D.; Jester, J.V. Characterization of growth and differentiation in a telomerase-immortalized human corneal epithelial cell line. Investig. Ophthalmol. Vis. Sci. 2005, 46, 470–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, G.; Lembach, K.J.; Morrison, M.M.; Cohen, S. Characterization of the binding of 125-I-labeled epidermal growth factor to human fibroblasts. J. Biol. Chem. 1975, 250, 4297–4304. [Google Scholar] [CrossRef]
- Limbird, L.E. A Short Course on Receptor Theory; Martinus Nyhoff/Kluwer Academic Publishing: Boston, MA, USA; Leiden, The Netherlands, 1985. [Google Scholar]
- Charoenrat, P.; Rhys-Evans, P.; Modjtahedi, H.; Court, W.; Box, G.; Eccles, S. Overexpression of epidermal growth factor receptor in human head and neck squamous carcinoma cell lines correlates with matrix metalloproteinase-9 expression and in vitro invasion. Int. J. Cancer 2000, 86, 307–317. [Google Scholar] [CrossRef]
- Filmus, J.; Pollak, M.N.; Cailleau, R.; Buick, R.N. MDA-468, a human breast cancer cell line with a high number of epidermal growth factor (EGF) receptors, has an amplified EGF receptor gene and is growth inhibited by EGF. Biochem. Biophys. Res. Commun. 1985, 128, 898–905. [Google Scholar] [CrossRef] [Green Version]
- Krupp, M.N.; Connolly, D.T.; Lane, M.D. Synthnesis, turnover, and down-regulation of epidermal growth factor receptors in human A431 epidermoid carcinoma cells and skin fibroblasts. J. Biol. Chem. 1982, 257, 11489–11496. [Google Scholar] [CrossRef]
- Fabricant, R.N.; Alpar, A.J.; Centifanto, Y.M.; Kaufman, H.E. Epidermal growth factor receptors on corneal endothelium. Arch. Ophthalmol. 1981, 99, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Engelmann, B.; Schumacher, U.; Haen, E. Epidermal growth factor binding sites on human erythrocytes in donors with different ABO blood groups. Am. J. Hematol. 1992, 39, 239–241. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.G.; Hollenberg, M.D. Distinct receptors for epidermal growth factor-urogastrone in cultured gastric smooth muscle cells. Am. J. Physiol. Content 1991, 260, G827–G834. [Google Scholar] [CrossRef] [PubMed]
- Haigler, H.; Ash, J.F.; Singer, S.J.; Cohen, S. Visualization by fluorescence of the binding and internalization of epidermal growth factor in human carcinoma cells A-431. Proc. Natl. Acad. Sci. USA 1978, 75, 3317–3321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajagopal, S.; Huang, S.; Moskal, T.L.; Lee, B.-N.; el-Naggar, A.K.; Chakrabarty, S. Epidermal growth factor expression in human colon and colon carcinomas: Anti-sense epidermal growth factor receptor RNA down-regulates the proliferation of human colon cancer cells. Int. J. Cancer 1995, 62, 661–667. [Google Scholar] [CrossRef]
- Yang, E.B.; Wang, D.F.; Mack, P.; Cheng, L.Y. EGF receptor in Chang liver and hepatoma HepG2 cells. Biochem. Mol. Boil. Int. 1996, 38, 813–820. [Google Scholar]
- Lee, L.S.; Weinstein, I.B. Tumor-promoting phorbol esters inhibit binding of epidermal growth factor to cellular receptors. Science 1978, 202, 313–315. [Google Scholar] [CrossRef]
- Imai, Y.; Leung, C.K.; Friesen, H.G.; Shiu, R.P. Epidermal growth factor receptors and effect of epidermal growth factor on growth of human breast cancer cells in long-term tissue culture. Cancer Res. 1982, 42, 4394–4398. [Google Scholar]
- De Souza, G.A.; Godoy, L.M.; Mann, M. Identification of 491 proteins in the tear fluid proteome reveals a large number of proteases and protease inhibitors. Genome Biol. 2006, 7, R72. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.E.; Liang, Q.; Kim, W.J. Lacrimal gland HGF, KGF, and EGF mRNA levels increase after corneal epithelial wounding. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2185–2190. [Google Scholar]
- Xu, K.-P.; Ding, Y.; Ling, J.; Dong, Z.; Yu, F.-S.X. Wound-induced HB-EGF ectodomain shedding and EGFR activation in corneal epithelial cells. Investig. Opthalmology Vis. Sci. 2004, 45, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Begley, C.G.; Port, N.; Bradley, A.; Braun, R.; King-Smith, E. The effects of increasing ocular surface stimulation on blinking and tear secretion. Investig. Ophthalmol. Vis. Sci. 2015, 56, 4211–4220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohashi, Y.; Motokura, M.; Kinoshita, Y.; Mano, T.; Watanabe, H.; Kinoshita, S.; Manabe, R.; Oshiden, K.; Yanaihara, C. Presence of epidermal growth factor in human tears. Investig. Ophthalmol. Vis. Sci. 1989, 30, 1879–1882. [Google Scholar]
- Van Setten, G.B.; Schultz, G.S.; Macauley, S. Growth factors in human tear fluid and in lacrimal glands. Adv. Exp. Med. Biol. 1994, 350, 315–319. [Google Scholar] [PubMed]
- Van Setten, G.B.; Tervo, K.; Virtanen, I.; Tarkkanen, A.; Tervo, T. Immunohistochemical demonstration of epidermal growth factor in the lacrimal and submandibular glands of rats. Acta Ophthalmol. 1990, 68, 477–480. [Google Scholar] [CrossRef] [PubMed]
- Block, E.R.; Matela, A.R.; SundarRaj, N.; Iszkula, E.R.; Klarlund, J.K. Wounding induces motility in sheets of corneal epithelial cells through loss of spatial constraints. J. Biol. Chem. 2004, 279, 36166. [Google Scholar] [CrossRef]
- Boucher, I.; Yang, L.; Mayo, C.; Klepeis, V.; Trinkaus-Randall, V. Injury and nucleotides induce phosphorylation of epidermal growth factor receptor: MMP and HB-EGF dependent pathway. Exp. Eye Res. 2007, 85, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Block, E.R.; Klarlund, J.K. Wounding sheets of epithelial cells activates the epidermal growth factor receptor through distinct short- and long-range mechanisms. Mol. Biol. Cell 2008, 19, 4909–4917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, S.E.; Chen, L.; Mohanab, R.R.; Liang, Q.; Liu, J. Expression of HGF, KGF, EGF and receptor messenger RNAs following corneal epithelial wounding. Exp. Eye Res. 1999, 68, 377–397. [Google Scholar] [CrossRef]
- Daniele, S.; Frati, L.; Fiore, C.; Santoni, G. The effect of the epidermal growth factor (EGF) on the corneal epithelium in humans. Albrecht Graefes Arch. Klin. Exp. Ophthalmol. 1979, 210, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Scardovi, C.; De Felice, G.; Gazzaniga, A. Epidermal growth factor in the topical treatment of traumatic corneal ulcers. Ophthalmologica 1993, 206, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Dellaert, M.M.M.J.; A Casey, T.; Wiffen, S.; Gordon, J.; Johnson, P.; Geerards, A.J.M.; Rijneveld, W.J.; Remeijer, L.; Mulder, P.G.H.; Beekhuis, W.H. Influence of topical human epidermal growth factor on postkeratoplasty re-epithelialisation. Br. J. Ophthalmol. 1997, 81, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Kandarakis, A.S.; Page, C.; Kaufman, H.E. The effect of epidermal growth factor on epithelial healing after penetrating keratoplasty in human eyes. Am. J. Ophthalmol. 1984, 98, 411–415. [Google Scholar] [CrossRef]
- Koehn, D.; Meyer, K.J.; Syed, N.A.; Anderson, M.G. Ketamine/xylazine-induced corneal damage in mice. PLoS ONE 2015, 10, e0132804. [Google Scholar] [CrossRef]
- Ghaffari, M.S.; Malmasi, A.; Bokaie, S. Effect of acepromazine or xylazine on tear production as measured by Schirmer tear test in normal cats. Vet. Ophthalmol. 2010, 13, 1–3. [Google Scholar] [CrossRef]
- Ceresa, B.P.; Peterson, J.L. Cell and molecular biology of epidermal growth factor receptor. Int. Rev. Cell Mol. Biol. 2014, 313, 145–178. [Google Scholar]
- Liu, Z.; Carvajal, M.; Carraway, C.A.C.; Carraway, K.; Pflugfelder, S.C. Expression of the receptor tyrosine kinases, epidermal growth factor receptor, ErbB2, and ErbB3, in human ocular surface epithelia. Cornea 2001, 20, 81–85. [Google Scholar] [CrossRef]
- Swan, J.S.; Arango, M.E.; Carothers-Carraway, C.A.C.; Carraway, K.L. An ErbB-2-Muc4 complex in rat ocular surface epithelia. Curr. Eye Res. 2002, 24, 397–402. [Google Scholar] [CrossRef]
- Xu, K.-P.; Riggs, A.; Ding, Y.; Yu, F.-S.X. Role of ErbB2 in corneal epithelial wound healing. Investig. Ophthalmol. Vis. Sci. 2004, 45, 4277–4283. [Google Scholar] [CrossRef] [Green Version]
- Kreps, E.O.; Derveaux, T.; Denys, H. Corneal changes in trastuzumab emtansine treatment. Clin. Breast Cancer 2018, 18, e427–e429. [Google Scholar] [CrossRef]
- Orlandi, A.; Fasciani, R.; Cassano, A.; Agresta, A.; Calegari, M.A.; Caporossi, A.; Barone, C. Trastuzumab-induced corneal ulceration: Successful no-drug treatment of a “blind” side effect in a case report. BMC Cancer 2015, 15, 973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinkamp, M.P.; Low-Nam, S.T.; Yang, S.; Lidke, K.A.; Lidke, D.S.; Wilson, B.S. erbB3 is an active tyrosine kinase capable of homo- and heterointeractions. Mol. Cell. Biol. 2014, 34, 965–977. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Wang, S.; Lyu, H.; Cai, B.; Yang, X.; Wang, J.; Liu, B. The anti-erbB3 antibody MM-121/SAR256212 in combination with trastuzumab exerts potent antitumor activity against trastuzumab-resistant breast cancer cells. Mol. Cancer 2013, 12, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rush, J.S.; Peterson, J.L.; Ceresa, B.P. Betacellulin (BTC) biases the EGFR to dimerize with ErbB3. Mol. Pharmacol. 2018, 94, 1382–1390. [Google Scholar] [CrossRef] [Green Version]
- Schlessinger, J. Allosteric regulation of the epidermal growth factor receptor kinase. Cell Biol. 1986, 103, 2067–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Shintani, A.; Nakata, M.; Shing, Y.; Folkman, J.; Igarashi, K.; Sasada, R. Recombinant human betacellulin. Molecular structure, biological activities, and receptor interaction. J. Biol. Chem. 1994, 269, 9966–9973. [Google Scholar] [CrossRef]
- Korc, M.; Magun, B.E. Recycling of epidermal growth factor in human pancreatic carcinoma cell line. Proc. Natl. Acad. Sci. USA 1985, 82, 6172–6175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roepstorff, K.; Grandal, M.V.; Henriksen, L.; Knudsen, S.L.J.; Lerdrup, M.; Grøvdal, L.; Willumsen, B.M.; Van Deurs, B. Differential effects of EGFR ligands on endocytic sorting of the receptor. Traffic 2009, 10, 1115–1127. [Google Scholar] [CrossRef]
- Hennessey, P.J.; Nirgiotis, J.G.; Shinn, M.N.; Andrassy, R.J. Continuous EGF application impairs long-term collagen accumulation during wound-healing in rats. J. Pediatr. Surg. 1991, 26, 362–366. [Google Scholar] [CrossRef]
- Nezu, E.; Ohashi, Y.; Kinoshita, S.; Manabe, R. Recombinant human epidermal growth-factor and corneal neovascularization. Jpn. J. Ophthalmol. 1992, 36, 401–406. [Google Scholar]
- Rao, K.; Farley, W.J.; Pflugfelder, S.C. Association between high tear epidermal growth factor levels and corneal subepithelial fibrosis in dry eye conditions. Investig. Opthalmology Vis. Sci. 2010, 51, 844–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceresa, B. Spatial regulation of epidermal growth factor receptor signaling by endocytosis. Int. J. Mol. Sci. 2012, 14, 72–87. [Google Scholar] [CrossRef] [Green Version]
- Dinneen, J.L.; Ceresa, B. Constitutive activation of rab5 results in a ligand independent redistribution of the EGFR and attenuates its ability to signal. Traffic 2004, 5, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Rush, J.S.; Ceresa, B. RAB7 and TSG101 are required for the constitutive recycling of unliganded EGFRs via distinct mechanisms. Mol. Cell. Endocrinol. 2013, 381, 188–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crotchett, B.L.M.; Ceresa, B. Knockout of c-Cbl slows EGFR endocytic trafficking and enhances EGFR signaling despite incompletely blocking receptor ubiquitylation. Pharmacol. Res. Perspect. 2021, 9, e00756. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cells | EGFRs/Cell | Reference |
|---|---|---|
| Non-cancerous cell lines | ||
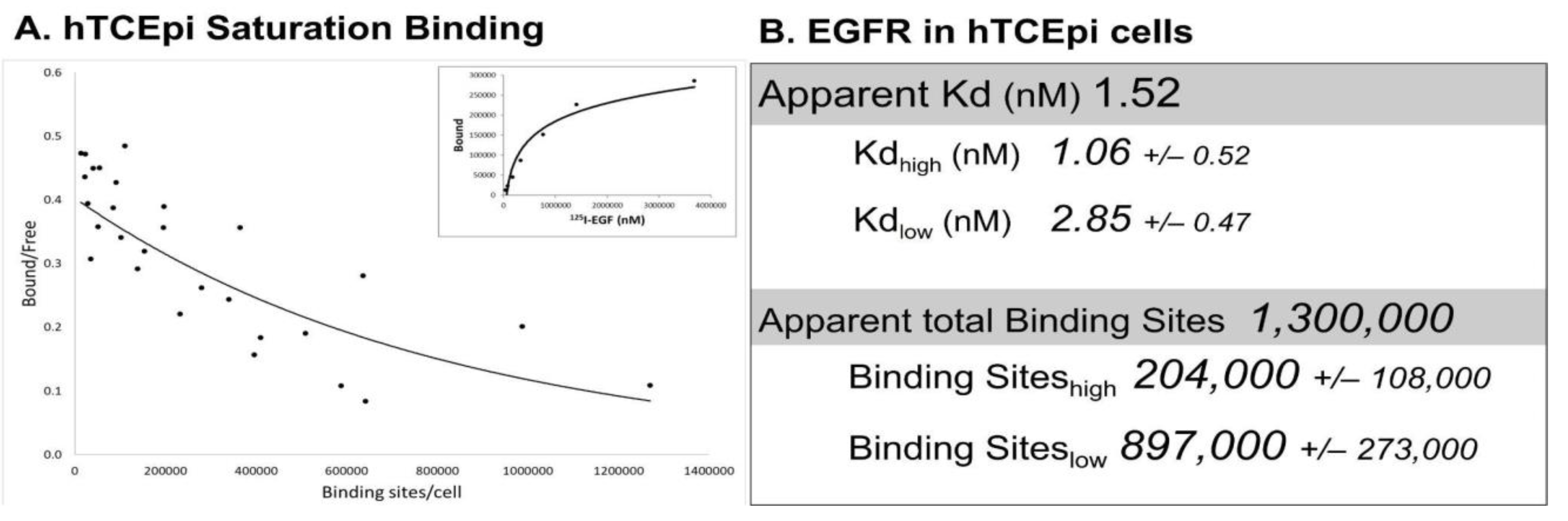
| hTCEpi (human) | 1,300,000 | Figure 2 |
| Oral mucosa | 200,000 | [73] |
| Skin fibroblasts | 51,000–70,000 | [71,74,75] |
| Primary corneal endothelium | 40,000 | [76] |
| Blood cells | 7800–25,400 | [77] |
| Gastric smooth muscle | 24,000 | [78] |
| Cancer cell lines | ||
| A431 cell (epidermoid carcinoma) | 1,500,000–2,600,000 | [74,75,79] |
| MDA-MB-468 cells (mammary gland) | 1,900,000 | [74] |
| Moser-1 (colon cancer) | 295,700 | [80] |
| HepG2 (liver carcinoma) | 180,000 | [81] |
| HT29 (colon cancer) | 120,000 | [80] |
| HeLa (cervical adenocarcinoma) | 43,500 | [82] |
| MCF-7 (mammary epithelial) | 2800–10,000 | [83] |
| KM12SM (colon cancer) | 7000 | [80] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peterson, J.L.; Ceresa, B.P. Epidermal Growth Factor Receptor Expression in the Corneal Epithelium. Cells 2021, 10, 2409. https://doi.org/10.3390/cells10092409
Peterson JL, Ceresa BP. Epidermal Growth Factor Receptor Expression in the Corneal Epithelium. Cells. 2021; 10(9):2409. https://doi.org/10.3390/cells10092409
Chicago/Turabian StylePeterson, Joanne L., and Brian P. Ceresa. 2021. "Epidermal Growth Factor Receptor Expression in the Corneal Epithelium" Cells 10, no. 9: 2409. https://doi.org/10.3390/cells10092409
APA StylePeterson, J. L., & Ceresa, B. P. (2021). Epidermal Growth Factor Receptor Expression in the Corneal Epithelium. Cells, 10(9), 2409. https://doi.org/10.3390/cells10092409