
PTX-3 Secreted by Intra-Articular-Injected SMUP-Cells Reduces Pain in an Osteoarthritis Rat Model

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
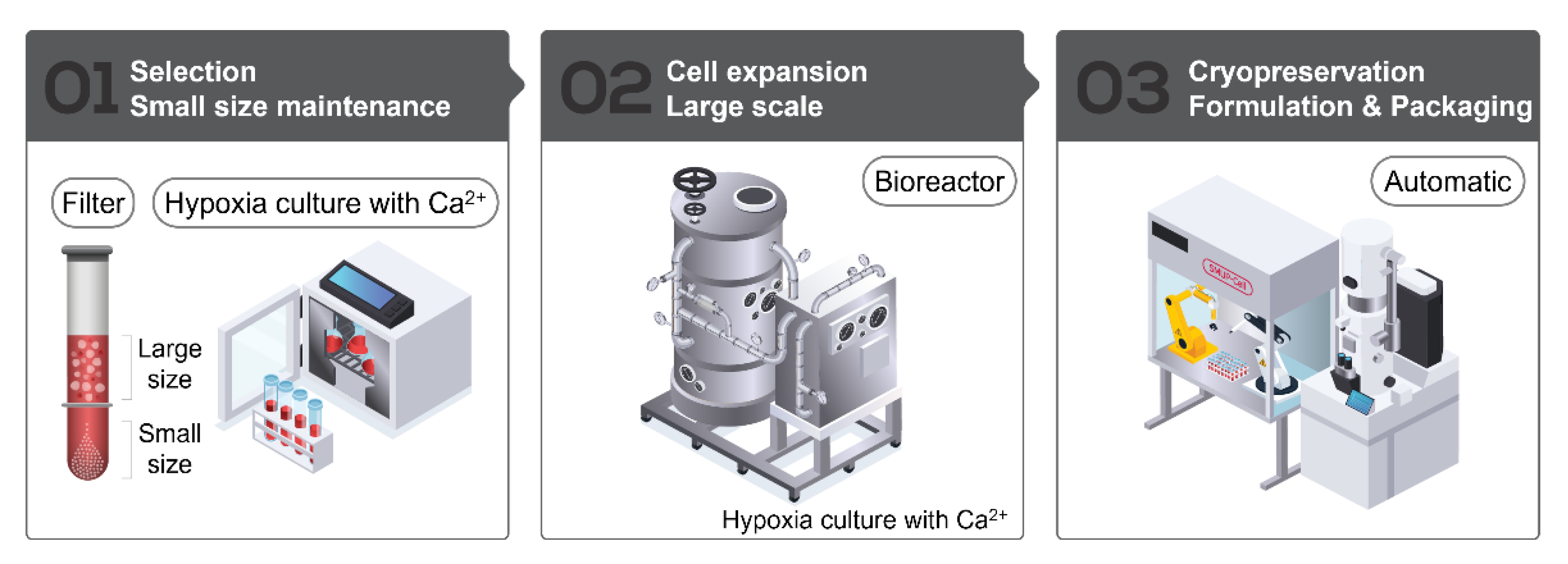
2.1. SMUP-Cell Platform
2.2. SMUP-Cell Characterization
2.3. Inflammation Status under the Co-Culture System
2.4. Immunofluorescence Staining
2.5. Cytokine Array and Western Blotting
2.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) and Small-Interfering (si)RNA Assay
2.7. Animals and Induction of OA
2.8. Pain Assessment: Measurement of Hind-limb Weight Distribution
2.9. Statistical Analysis
3. Results
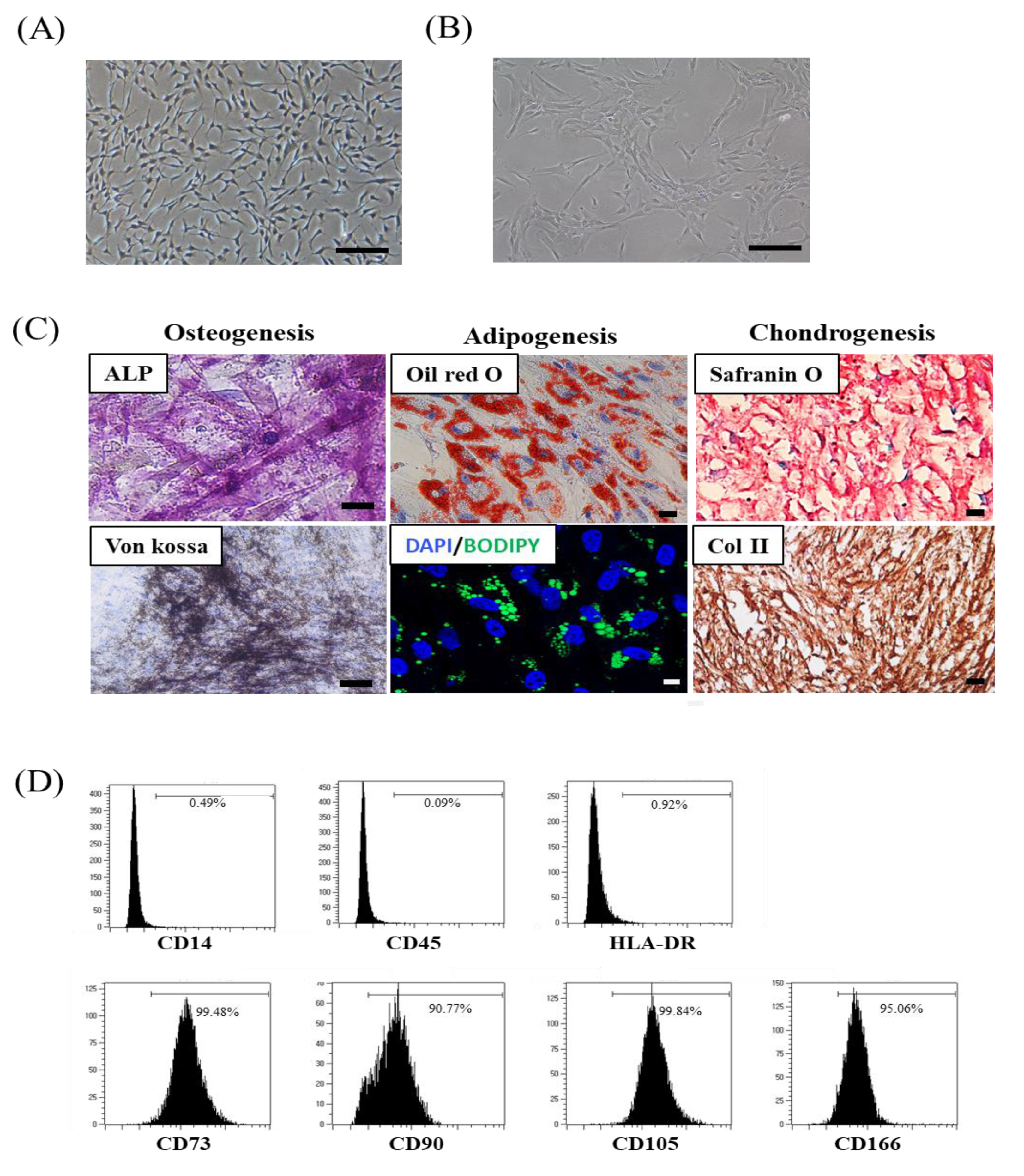
3.1. SMUP-Cells Possess Stem Cell Characteristics
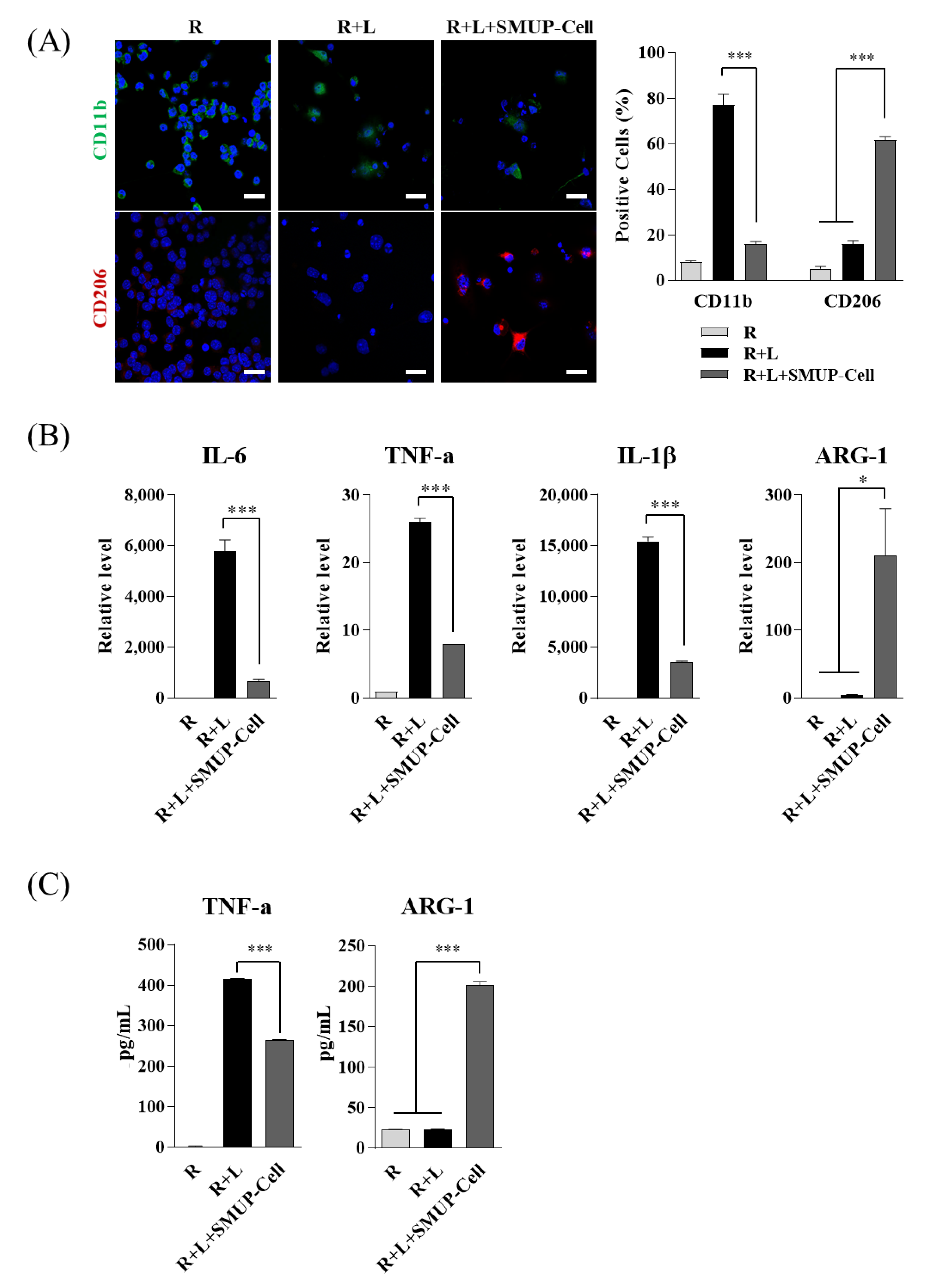
3.2. SMUP-Cells Activate Macrophage Polarization
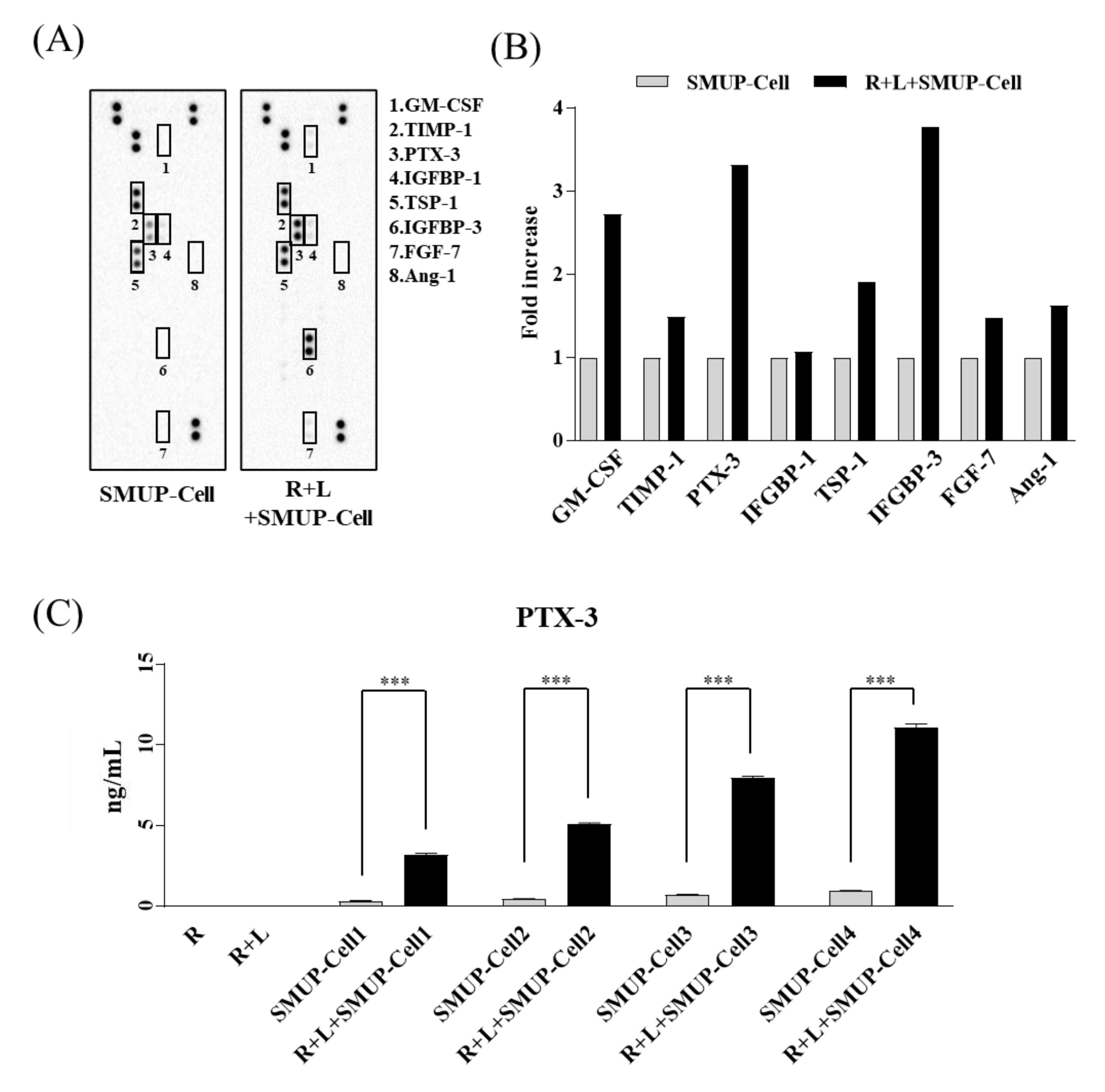
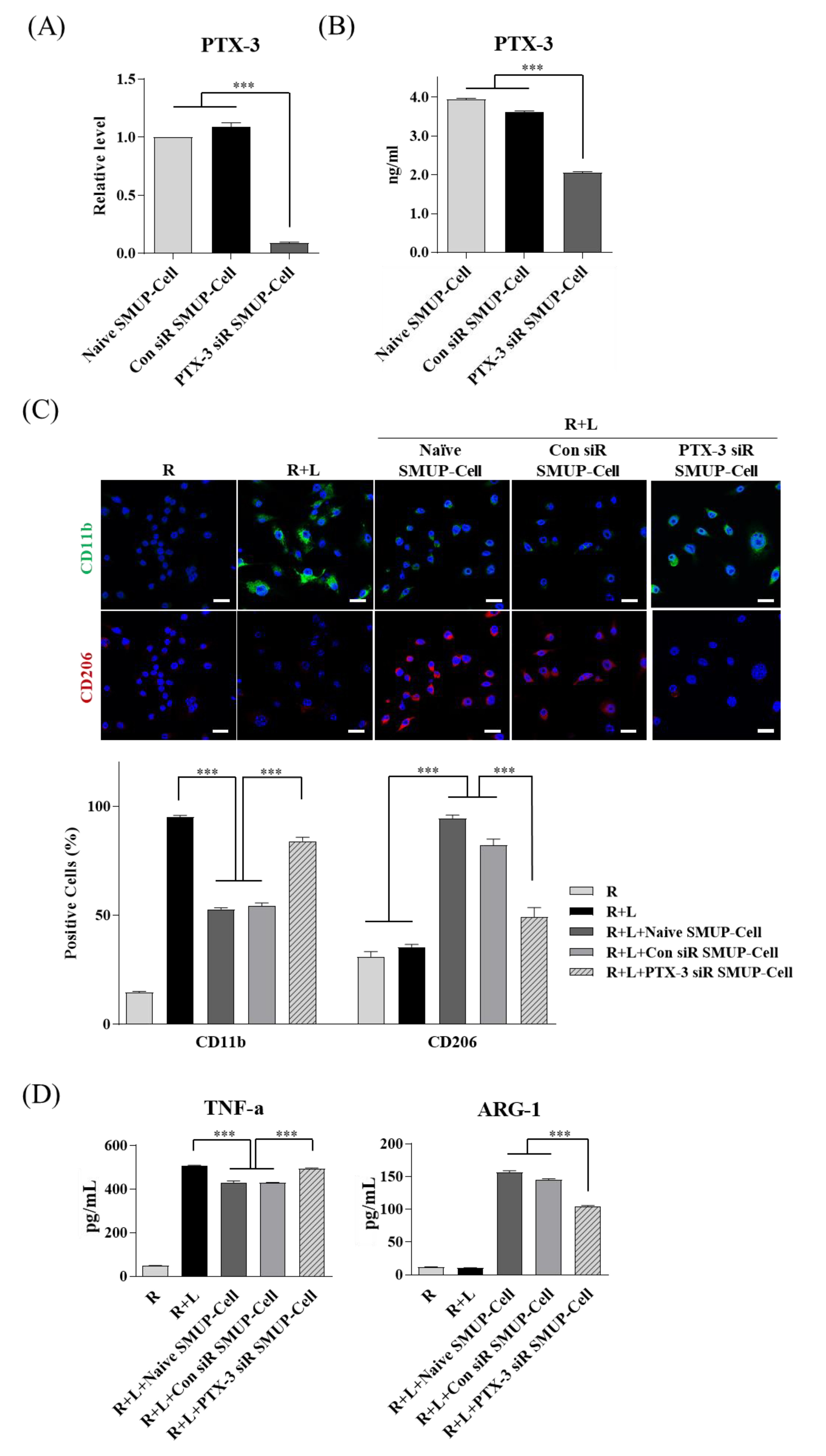
3.3. PTX-3 Secreted by SMUP-Cells Is Critical to Macrophage Polarization
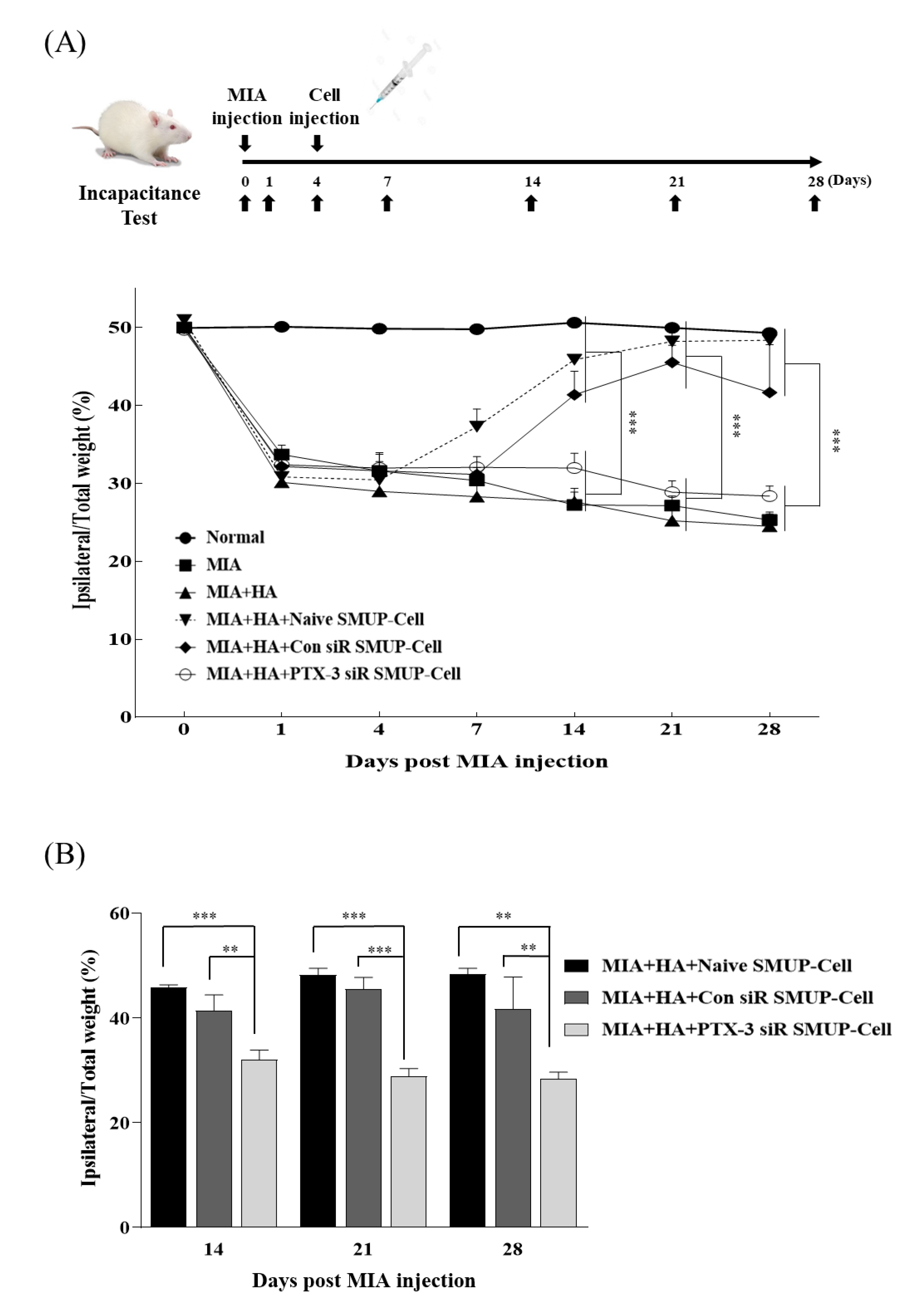
3.4. PTX-3 Improves the Therapeutic Capacity of SMUP-Cells in an OA Rat Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal Stem Cell Perspective: Cell Biology to Clinical Progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Song, J.S.; Hong, K.T.; Kim, N.M.; Park, H.S.; Choi, N.H. Human Umbilical Cord Blood-Derived Mesenchymal Stem Cell Implantation for Osteoarthritis of the Knee. Arch. Orthop. Trauma. Surg. 2020, 140, 503–509. [Google Scholar] [CrossRef]
- Ichiseki, T.; Shimazaki, M.; Ueda, Y.; Ueda, S.; Tsuchiya, M.; Souma, D.; Kaneuji, A.; Kawahara, N. Intraarticularly-Injected Mesenchymal Stem Cells Stimulate Anti-Inflammatory Molecules and Inhibit Pain Related Protein and Chondrolytic Enzymes in a Monoiodoacetate-Induced Rat Arthritis Model. Int. J. Mol. Sci. 2018, 19, 203. [Google Scholar] [CrossRef] [Green Version]
- Um, S.; Ha, J.; Choi, S.J.; Oh, W.; Jin, H.J. Prospects for the Therapeutic Development of Umbilical Cord Blood-Derived Mesenchymal Stem Cells. World J. Stem Cells 2020, 12, 1511–1528. [Google Scholar] [CrossRef]
- Zhou, X.; Hong, Y.; Zhang, H.; Li, X. Mesenchymal Stem Cell Senescence and Rejuvenation: Current Status and Challenges. Front. Cell Dev. Biol. 2020, 8, 364. [Google Scholar] [CrossRef]
- Cunha, B.; Aguiar, T.; Carvalho, S.B.; Silva, M.M.; Gomes, R.A.; Carrondo, M.J.T.; Gomes-Alves, P.; Peixoto, C.; Serra, M.; Alves, P.M. Bioprocess Integration for Human Mesenchymal Stem Cells: From up to Downstream Processing Scale-Up to Cell Proteome Characterization. J. Biotechnol. 2017, 248, 87–98. [Google Scholar] [CrossRef]
- Phelps, J.; Sanati-Nezhad, A.; Ungrin, M.; Duncan, N.A.; Sen, A. Bioprocessing of Mesenchymal Stem Cells and Their Derivatives: Toward Cell-Free Therapeutics. Stem Cells Int. 2018, 2018, 9415367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, W.; Kwon, S.J.; Jin, H.J.; Jeong, S.Y.; Choi, S.J.; Oh, W.; Yang, Y.S.; Jeon, H.B.; Jeon, E.S. Optimization of Culture Conditions for Rapid Clinical-Scale Expansion of Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells. Clin. Transl. Med. 2017, 6, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Jin, H.J.; Heo, J.; Ju, H.; Lee, H.Y.; Kim, S.; Lee, S.; Lim, J.; Jeong, S.Y.; Kwon, J.; et al. Small Hypoxia-Primed Mesenchymal Stem Cells Attenuate Graft-Versus-Host Disease. Leukemia 2018, 32, 2672–2684. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Bae, Y.K.; Um, S.; Kwon, J.H.; Kim, G.H.; Choi, S.J.; Oh, W.; Jin, H.J. A Small-Sized Population of Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells Shows High Stemness Properties and Therapeutic Benefit. Stem Cells Int. 2020, 2020, 5924983. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.H.; Kim, M.; Um, S.; Lee, H.J.; Bae, Y.K.; Choi, S.J.; Hwang, H.H.; Oh, W.; Jin, H.J. Senescence-Associated Secretory Phenotype Suppression Mediated by Small-Sized Mesenchymal Stem Cells Delays Cellular Senescence Through TLR2 and TLR5 Signaling. Cells 2021, 10, 63. [Google Scholar] [CrossRef]
- Okubo, M.; Okada, Y. Destruction of the Articular Cartilage in Osteoarthritis. Clin. Calcium. 2013, 23, 1705–1713. [Google Scholar] [PubMed]
- Chen, D.; Shen, J.; Zhao, W.; Wang, T.; Han, L.; Hamilton, J.L.; Im, H.J. Osteoarthritis: Toward a Comprehensive Understanding of Pathological Mechanism. Bone Res. 2017, 5, 16044. [Google Scholar] [CrossRef] [PubMed]
- Neogi, T. The Epidemiology and Impact of Pain in Osteoarthritis. Osteoarthr. Cartil. 2013, 21, 1145–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeser, R.F.; Collins, J.A.; Diekman, B.O. Ageing and the Pathogenesis of Osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 412–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado, M.; Nam, J. The Role of Changes in Extracellular Matrix of Cartilage in the Presence of Inflammation on the Pathology of Osteoarthritis. BioMed Res. Int. 2013, 2013, 284873. [Google Scholar] [CrossRef] [Green Version]
- Riegger, J.; Brenner, R.E. Pathomechanisms of Posttraumatic Osteoarthritis: Chondrocyte Behavior and Fate in a Precarious Environment. Int. J. Mol. Sci. 2020, 21, 1560. [Google Scholar] [CrossRef] [Green Version]
- Lambrechts, T.; Papantoniou, I.; Viazzi, S.; Bovy, T.; Schrooten, J.; Luyten, F.P.; Aerts, J.-M. Evaluation of a Monitored Multiplate Bioreactor for Large-Scale Expansion of Human Periosteum Derived Stem Cells for Bone Tissue Engineering Applications. Biochem. Eng. J. 2016, 108, 58–68. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.J.; Kwon, J.H.; Kim, M.; Bae, Y.K.; Choi, S.J.; Oh, W.; Yang, Y.S.; Jeon, H.B. Downregulation of Melanoma Cell Adhesion Molecule (MCAM/CD146) Accelerates Cellular Senescence in Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells. Stem Cells Transl. Med. 2016, 5, 427–439. [Google Scholar] [CrossRef]
- Bae, Y.K.; Kwon, J.H.; Kim, M.; Kim, G.H.; Choi, S.J.; Oh, W.; Jeon, H.B. Intracellular Calcium Determines the Adipogenic Differentiation Potential of Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells via the Wnt5a/β-Catenin Signaling Pathway. Stem Cells International 2018, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, Y.; Fujita, M.; Kawasaki, S.; Sanaki, T.; Yoshioka, T.; Higashino, K.; Tofukuji, S.; Yoneda, S.; Takahashi, T.; Koda, K.; et al. Contribution of Synovial Macrophages to Rat Advanced Osteoarthritis Pain Resistant to Cyclooxygenase Inhibitors. Pain 2019, 160, 895–907. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Voog, J.; Jones, D.L. Stem Cells and the Niche: A Dynamic Duo. Cell Stem Cell 2010, 6, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Guo, H.; Bai, F.; Zhang, M.; Yang, L.; Deng, J.; Xiong, L. A Rat Model of Knee Osteoarthritis Suitable for Electroacupuncture Study. Exp. Anim. 2018, 67, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.; Yu, H.Y.; Heo, J.; Song, M.; Shin, J.H.; Lim, J.; Yoon, S.J.; Kim, Y.; Lee, S.; Kim, S.W.; et al. Mesenchymal Stem Cells Protect Against the Tissue Fibrosis of Ketamine-Induced Cystitis in Rat Bladder. Sci. Rep. 2016, 6, 30881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jossen, V.; van den Bos, C.; Eibl, R.; Eibl, D. Manufacturing Human Mesenchymal Stem Cells at Clinical Scale: Process and Regulatory Challenges. Appl. Microbiol. Biotechnol. 2018, 102, 3981–3994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mennan, C.; Garcia, J.; Roberts, S.; Hulme, C.; Wright, K. A Comprehensive Characterisation of Large-Scale Expanded Human Bone Marrow and Umbilical Cord Mesenchymal Stem Cells. Stem Cell Res. Ther. 2019, 10, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, A.L.; Lefavor, R.C.; Zubair, A.C. Characterization and Cost–Benefit Analysis of Automated Bioreactor-Expanded Mesenchymal Stem Cells for Clinical Applications. Transfusion 2018, 58, 2374–2382. [Google Scholar] [CrossRef]
- Salehi-Nik, N.; Amoabediny, G.; Pouran, B.; Tabesh, H.; Shokrgozar, M.A.; Haghighipour, N.; Khatibi, N.; Anisi, F.; Mottaghy, K.; Zandieh-Doulabi, B. Engineering Parameters in Bioreactor’s Design: A Critical Aspect in Tissue Engineering. BioMed. Res. Int. 2013, 2013, 762132. [Google Scholar] [CrossRef]
- Robb, K.P.; Fitzgerald, J.C.; Barry, F.; Viswanathan, S. Mesenchymal Stromal Cell Therapy: Progress in Manufacturing and Assessments of Potency. Cytotherapy. 2019, 21, 289–306. [Google Scholar] [CrossRef]
- Mesquita, T.J.B.; Sargo, C.R.; Fuzer, J.R.N.; Paredes, S.A.H.; Giordano, R.C.; Horta, A.C.L.; Zangirolami, T.C. Metabolic Fluxes-Oriented Control of Bioreactors: A Novel Approach to Tune Micro-Aeration and Substrate Feeding in Fermentations. Microb. Cell Factories 2019, 18, 150. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, S.; Radhi, M.; Gowler, P.; Burston, J.J.; Gandhi, R.D.; Thorn, G.J.; Piccinini, A.M.; Walsh, D.A.; Chapman, V.; de Moor, C.H. The Polyadenylation Inhibitor Cordycepin Reduces Pain, Inflammation and Joint Pathology in Rodent Models of Osteoarthritis. Sci. Rep. 2019, 9, 4696. [Google Scholar] [CrossRef]
- Jin, H.J.; Bae, Y.K.; Kim, M.; Kwon, S.J.; Jeon, H.B.; Choi, S.J.; Kim, S.W.; Yang, Y.S.; Oh, W.; Chang, J.W. Comparative Analysis of Human Mesenchymal Stem Cells from Bone Marrow, Adipose Tissue, and Umbilical Cord Blood as Sources of Cell Therapy. Int. J. Mol. Sci. 2013, 14, 17986–18001. [Google Scholar] [CrossRef]
- Almalki, S.G.; Agrawal, D.K. Effects of Matrix Metalloproteinases on the Fate of Mesenchymal Stem Cells. Stem Cell Res. Ther. 2016, 7, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Lim, H.; Lee, D.; Choi, S.J.; Oh, W.; Yang, Y.S.; Oh, J.S.; Hwang, H.H.; Jeon, H.B. Thrombospondin-1 Secreted by Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells Rescues Neurons from Synaptic Dysfunction in Alzheimer’s Disease Model. Sci. Rep. 2018, 8, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Kwon, J.H.; Bae, Y.K.; Kim, G.H.; Um, S.; Ha, J.; Choi, S.J.; Oh, W.; Jin, H.J. Soluble PTX3 of Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells Attenuates Hyperoxic Lung Injury by Activating Macrophage Polarization in Neonatal Rat Model. Stem Cells Int. 2020, 2020, 1802976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, T.L.; Gomoll, A.H.; Lattermann, C.; Hernandez, A.J.; Bueno, D.F.; Amano, M.T. Macrophage: A Potential Target on Cartilage Regeneration. Front. Immunol. 2020, 11, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, F.O.; Gordon, S. The M1 and M2 Paradigm of Macrophage Activation: Time for Reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.E.; Jang, J.E.; Kim, H.S.; Jung, M.K.; Ko, M.S.; Kim, M.O.; Koh, E.H. Mesenchymal stem cells prevent the progression of diabetic nephropathy by improving mitochondrial function in tubular epithelial cells. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Shiraki, A.; Kotooka, N.; Komoda, H.; Hirase, T.; Oyama, J.I.; Node, K. Pentraxin-3 Regulates the Inflammatory Activity of Macrophages. Biochem. Biophys. Rep. 2016, 5, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Elcombe, S.E.; Naqvi, S.; Van Den Bosch, M.W.M.; MacKenzie, K.F.; Cianfanelli, F.; Brown, G.D.; Arthur, J.S.C. Dectin-1 Regulates IL-10 Production via a MSK1/2 and CREB Dependent Pathway and Promotes the Induction of Regulatory Macrophage Markers. PLoS ONE 2013, 8, e60086. [Google Scholar] [CrossRef] [Green Version]
- Reyskens, K.M.S.E.; Arthur, J.S.C. Emerging Roles of the Mitogen and Stress Activated Kinases MSK1 and MSK2. Front. Cell Dev. Biol. 2016, 4, 56. [Google Scholar] [CrossRef] [Green Version]
- Schou, W.S.; Ashina, S.; Amin, F.M.; Goadsby, P.J.; Ashina, M. Calcitonin gene-related peptide and pain: A systematic review. J. Headache. Pain. 2017, 18, 34. [Google Scholar] [CrossRef] [Green Version]
- Rogoz, K.; Andersen, H.H.; Lagerstrom, M.C.; Kullander, K. Multimodal Use of Calcitonin Gene-Related Peptide and Substance P in Itch and Acute Pain Uncovered by the Elimination of Vesicular Glutamate Transporter 2 from Transient Receptor Potential Cation Channel Subfamily V Member 1 Neurons. J. Neurosci. 2014, 34, 14055–14068. [Google Scholar] [CrossRef] [Green Version]
- Cappuzzello, C.; Doni, A.; Dander, E.; Pasqualini, F.; Nebuloni, M.; Bottazzi, B.; Mantovani, A.; Biondi, A.; Garlanda, C.; D’Amico, G. Mesenchymal Stromal Cell-Derived PTX3 Promotes Wound Healing via Fibrin Remodeling. J. Invest. Dermatol. 2016, 136, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.W.; Moon, H.E.; Kim, H.S.; Paek, S.L.; Kim, Y.; Chang, J.W.; Yang, Y.S.; Kim, K.; Oh, W.; Hwang, J.H.; et al. Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells Improve Functional Recovery Through Thrombospondin1, pantraxin3, and Vascular Endothelial Growth Factor in the Ischemic Rat Brain. J. Neurosci. Res. 2015, 93, 1814–1825. [Google Scholar] [CrossRef] [PubMed]
- Mauri, T.; Zambelli, V.; Cappuzzello, C.; Bellani, G.; Dander, E.; Sironi, M.; Castiglioni, V.; Doni, A.; Mantovani, A.; Biondi, A.; et al. Intraperitoneal Adoptive Transfer of Mesenchymal Stem Cells Enhances Recovery from Acid Aspiration Acute Lung Injury in Mice. Intensive Care Med. Exp. 2017, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghouri, A.; Conaghan, P.G. Update on Novel Pharmacological Therapies for Osteoarthritis. Ther. Adv. Musculoskelet. Dis. 2019, 11, 1759720X19864492. [Google Scholar] [CrossRef]
- Grässel, S.; Muschter, D. Recent Advances in the Treatment of Osteoarthritis. F1000Research 2020, 9, F1000. [Google Scholar] [CrossRef] [PubMed]
- Im, G.I. Perspective on Intra-articular Injection Cell Therapy for Osteoarthritis Treatment. Tissue. Eng. Regen. Med. 2019, 16, 357–363. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.; Kim, G.-H.; Kim, M.; Seo, J.M.; Kim, Y.M.; Seon, M.R.; Um, S.; Choi, S.J.; Oh, W.; Song, B.R.; et al. PTX-3 Secreted by Intra-Articular-Injected SMUP-Cells Reduces Pain in an Osteoarthritis Rat Model. Cells 2021, 10, 2420. https://doi.org/10.3390/cells10092420
Lee M, Kim G-H, Kim M, Seo JM, Kim YM, Seon MR, Um S, Choi SJ, Oh W, Song BR, et al. PTX-3 Secreted by Intra-Articular-Injected SMUP-Cells Reduces Pain in an Osteoarthritis Rat Model. Cells. 2021; 10(9):2420. https://doi.org/10.3390/cells10092420
Chicago/Turabian StyleLee, Minju, Gee-Hye Kim, Miyeon Kim, Ji Min Seo, Yu Mi Kim, Mi Ra Seon, Soyoun Um, Soo Jin Choi, Wonil Oh, Bo Ram Song, and et al. 2021. "PTX-3 Secreted by Intra-Articular-Injected SMUP-Cells Reduces Pain in an Osteoarthritis Rat Model" Cells 10, no. 9: 2420. https://doi.org/10.3390/cells10092420
APA StyleLee, M., Kim, G. -H., Kim, M., Seo, J. M., Kim, Y. M., Seon, M. R., Um, S., Choi, S. J., Oh, W., Song, B. R., & Jin, H. J. (2021). PTX-3 Secreted by Intra-Articular-Injected SMUP-Cells Reduces Pain in an Osteoarthritis Rat Model. Cells, 10(9), 2420. https://doi.org/10.3390/cells10092420