
Early Life Irradiation-Induced Hypoplasia and Impairment of Neurogenesis in the Dentate Gyrus and Adult Depression Are Mediated by MicroRNA- 34a-5p/T-Cell Intracytoplasmic Antigen-1 Pathway

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Radiation Exposure
2.2. Behavioral Tests
2.2.1. Open Field (Locomotor) Test
2.2.2. Tail Suspension Test
2.2.3. Forced Swim Test
2.3. Immunohistochemical Staining
2.4. RNA Extraction from the Mouse Brain
2.5. Systematic miR Sequencing (miRSeq) Analysis
2.6. Real-Time Quantitative Reverse Transcription PCR (qRT-PCR) Analysis of miR
2.7. Predication of miR-34a-5p Targets and Luciferase Reporter Assay
2.8. qRT-PCR Analysis of Tia1 mRNA
2.9. Western Blot
2.10. Culture of Neural Stem Cells
2.11. RNA Isolation from NSCs and qRT-PCR Analysis for miR and mRNA
2.12. Western Blot for NSCs
2.13. Overexpression of Tia1 in NSCs
2.14. Statistical Analysis
3. Results
3.1. ϒ-Irradiation at P3 Induced Depression-Like Behavior in Adult Mice
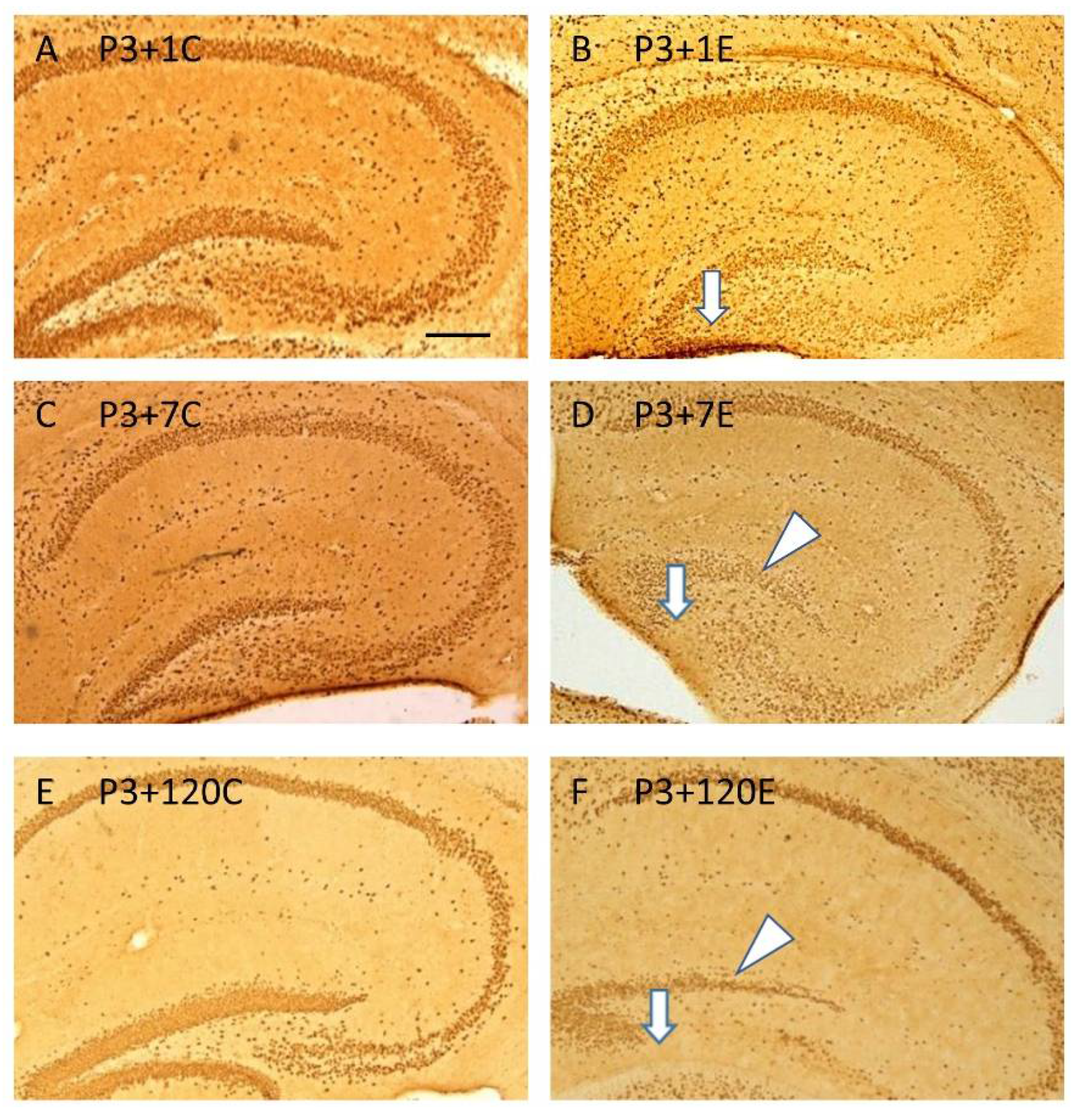
3.2. ϒ-Irradiation with 5Gy Induced Hypoplasia of The Infrapyramidal Blade of The Stratum Granulosum, and Aberrant and Impaired Neurogenesis in the Subgranular Zone of the Dentate Gyrus
3.3. Systematic miRSeq and Real Time RT-PCR
3.4. miR-34a-5p Targeted on mRNA of Tia1
3.5. mRNA and Protein Expression of Tia1 in Mice Brain after Γ-Irradiation with 5Gy
3.6. Γ-Irradiation Increased the Expression of miR-34a-5p in NSCs and Decreased the mRNA Expression of Tia1
3.7. Γ-Irradiation Dose- and Time-Dependently Decreased Protein Expression of Tia1 in NSCs
3.8. Overexpression of Tia1 Partially Blocked Γ-Irradiation-Induced Impairment of Neurogenesis
4. Discussion
4.1. Radiation, Hippocampal Neuropathology and Depression
4.2. miR-34a-5p in Neurogenesis and Depression
4.3. miR-34a-5p Targets Tia1 to Prevent Neurogenesis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| Atg9a | Autophagy-related 9a |
| BDNF | Brain-derived neurotrophic factor |
| bFGF | Basic fibroblast growth factor |
| CREB | Cyclic AMP response-element binding protein |
| DCX | Doublecortin |
| EGF | Epidermal growth factor |
| ERK | Extracellular signal-regulated kinase ½ |
| HEK | Human embryonic kidney |
| HMGA2 | High Mobility Group AT-Hook 2) |
| MAP2 | Activity and microtubule-associated protein 2 |
| MDD | Major depressive disorder |
| miR | MicroRNA |
| miRSeq | MicroRNA sequencing |
| MMP9 | Matrix metalloproteinase 9 |
| NF-κB | Nuclear factor kappa light chain enhancer of activated B |
| NLRC5 | A central nucleotide-binding oligomerization domain (NOD)-, C-terminal leucine rich repeat domain (LRR)- and caspase activation and recruitment domains (CARDs) -containing 5 |
| NMDA | N-methyl D-aspartate |
| NUMBL | NUMB Like Endocytic Adaptor Protein |
| NSC | Neural stem cell |
| PDGFRA | Platelet-derived growth factor receptor A |
| qRT-PCR | Real-time quantitative reverse transcription polymerase chain reaction |
| SEM | Standard error of the mean |
| SIRT1 | Sirtuin 1 |
| Syt1 | Synaptotagmin 1 |
| Tia1 | T-cell intracytoplasmic antigen-1 |
| UTR | Untranslated region |
References
- Loganovsky, K.N.; Vasilenko, Z.L. Depression and ionizing radiation. Probl. Radiatsiinoi Medytsyny Ta Radiobiolohii 2013, 18, 200–219. [Google Scholar]
- Remennick, L.I. Immigrants from Chernobyl-affected areas in Israel: The link between health and social adjustment. Soc. Sci. Med. 2002, 54, 309–317. [Google Scholar] [CrossRef]
- Loganovsky, K.N.; Loganovskaja, T.K.; Nechayev, S.Y.; Antipchuk, Y.Y.; Bomko, M.A. Disrupted development of the dominant hemisphere following prenatal irradiation. J. Neuropsychiatry. Clin. Neurosci. 2008, 20, 274–291. [Google Scholar] [CrossRef] [PubMed]
- Contis, G.; Foley, T.P., Jr. Depression, suicide ideation, and thyroid tumors among ukrainian adolescents exposed as children to chernobyl radiation. J. Clin. Med. Res. 2015, 7, 332–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, S.M.; Kern, K.C.; O’Connor, M.F.; Montag, M.J.; Kim, A.; Yoo, Y.S.; Giesser, B.S.; Sicotte, N.L. Smaller cornu ammonis 2-3/dentate gyrus volumes and elevated cortisol in multiple sclerosis patients with depressive symptoms. Biol. Psychiatry 2010, 68, 553–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapolsky, R.M. Why stress is bad for your brain. Science 1996, 273, 749–750. [Google Scholar] [CrossRef]
- Satterthwaite, T.D.; Cook, P.A.; Bruce, S.E.; Conway, C.; Mikkelsen, E.; Satchell, E.; Vandekar, S.N.; Durbin, T.; Shinohara, R.T.; Sheline, Y.I. Dimensional depression severity in women with major depression and post-traumatic stress disorder correlates with fronto-amygdalar hypoconnectivty. Mol. Psychiatry. 2016, 21, 894–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheline, Y.I.; Wang, P.W.; Gado, M.H.; Csernansky, J.G.; Vannier, M.W. Hippocampal atrophy in recurrent major depression. Proc. Natl. Acad. Sci. USA 1996, 93, 3908–3913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheline, Y.I. 3D MRI studies of neuroanatomic changes in unipolar major depression: The role of stress and medical comorbidity. Biol. Psychiatry 2000, 48, 791–800. [Google Scholar] [CrossRef]
- Zheng, L.S.; Hitoshi, S.; Kaneko, N.; Takao, K.; Miyakawa, T.; Tanaka, Y.; Xia, H.; Kalinke, U.; Kudo, K.; Kanba, S.; et al. Mechanisms for interferon-α-induced depression and neural stem cell dysfunction. Stem Cell Rep. 2014, 3, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Santarelli, L.; Saxe, M.; Gross, C.; Surget, A.; Battaglia, F.; Dulawa, S.; Weisstaub, N.; Lee, J.; Duman, R.; Arancio, O.; et al. Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science 2003, 301, 805–809. [Google Scholar] [CrossRef] [Green Version]
- Boldrini, M.; Galfalvy, H.; Dwork, A.J.; Rosoklija, G.B.; Trencevska-Ivanovska, I.; Pavlovski, G.; Hen, R.; Arango, V.; Mann, J.J. Resilience Is Associated With Larger Dentate Gyrus, While Suicide Decedents With Major Depressive Disorder Have Fewer Granule Neurons. Biol. Psychiatry 2019, 85, 850–862. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, B.L.; van Praag, H.; Gage, F.H. Adult brain neurogenesis and psychiatry: A novel theory of depression. Mol. Psychiatry 2000, 5, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G.; Kronenberg, G. Depressed new neurons—Adult hippocampal neurogenesis and a cellular plasticity hypothesis of major depression. Biol. Psychiatry 2003, 54, 499–503. [Google Scholar] [CrossRef]
- Vogel, G. Neuroscience. Depression drugs’ powers may rest on new neurons. Science 2003, 301, 757. [Google Scholar] [CrossRef]
- Warner-Schmidt, J.L.; Duman, R.S. Hippocampal neurogenesis: Opposing effects of stress and antidepressant treatment. Hippocampus 2006, 16, 239–249. [Google Scholar] [CrossRef]
- Kang, J.; Kim, W.; Seo, H.; Kim, E.; Son, B.; Lee, S.; Park, G.; Jo, S.; Moon, C.; Youn, H.; et al. Radiation-induced overexpression of transthyretin inhibits retinol-mediated hippocampal neurogenesis. Sci. Rep. 2018, 8, 8394. [Google Scholar] [CrossRef] [Green Version]
- Snyder, J.S.; Soumier, A.; Brewer, M.; Pickel, J.; Cameron, H.A. Adult hippocampal neurogenesis buffers stress responses and depressive behaviour. Nature 2011, 476, 458–461. [Google Scholar] [CrossRef]
- Son, Y.; Yang, M.; Kim, J.S.; Kim, J.; Kim, S.H.; Kim, J.C.; Shin, T.; Wang, H.; Jo, S.K.; Jung, U.; et al. Hippocampal dysfunction during the chronic phase following a single exposure to cranial irradiation. Exp. Neurol. 2014, 254, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Yang, M.; Wang, H.; Moon, C. Hippocampal dysfunctions caused by cranial irradiation: A review of the experimental evidence. Brain Behav. Immun. 2015, 45, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.R.; Loke, W.K.; Wong, P.; Khoo, B.C. Radioprotective effect of ursolic acid in radiation-induced impairment of neurogenesis, learning and memory in adolescent BALB/c mouse. Physiol. Behav. 2017, 175, 37–46. [Google Scholar] [CrossRef]
- Guo, Y.R.; Liu, Z.W.; Peng, S.; Duan, M.Y.; Feng, J.W.; Wang, W.F.; Xu, Y.H.; Tang, X.; Zhang, X.Z.; Ren, B.X.; et al. The Neuroprotective Effect of Amitriptyline on Radiation-Induced Impairment of Hippocampal Neurogenesis. Dose-Response A Publ. Int. Hormesis Soc. 2019, 17, 1559325819895912. [Google Scholar] [CrossRef]
- Wang, S.W.; Ren, B.X.; Qian, F.; Luo, X.Z.; Tang, X.; Peng, X.C.; Huang, J.R.; Tang, F.R. Radioprotective effect of epimedium on neurogenesis and cognition after acute radiation exposure. Neurosci. Res. 2019, 145, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Prabu, P.; Poongothai, S.; Shanthirani, C.S.; Anjana, R.M.; Mohan, V.; Balasubramanyam, M. Altered circulatory levels of miR-128, BDNF, cortisol and shortened telomeres in patients with type 2 diabetes and depression. Acta Diabetol. 2020, 57, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Wingo, T.S.; Yang, J.; Fan, W.; Min Canon, S.; Gerasimov, E.S.; Lori, A.; Logsdon, B.; Yao, B.; Seyfried, N.T.; Lah, J.J.; et al. Brain microRNAs associated with late-life depressive symptoms are also associated with cognitive trajectory and dementia. NPJ Genom. Med. 2020, 5, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.P.; Liu, X.L.; Chen, J.J.; Cheng, K.; Bai, S.J.; Zheng, P.; Zhou, C.J.; Wang, W.; Wang, H.Y.; Zhong, L.M.; et al. Circulating microRNA 134 sheds light on the diagnosis of major depressive disorder. Transl. Psychiatry 2020, 10, 95. [Google Scholar] [CrossRef]
- Maffioletti, E.; Cattaneo, A.; Rosso, G.; Maina, G.; Maj, C.; Gennarelli, M.; Tardito, D.; Bocchio-Chiavetto, L. Peripheral whole blood microRNA alterations in major depression and bipolar disorder. J. Affect. Disord. 2016, 200, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Segaran, R.C.; Chan, L.Y.; Wang, H.; Sethi, G.; Tang, F.R. Neuronal Development-Related miRNAs as Biomarkers for Alzheimer’s disease, Depression, Schizophrenia and Ionizing Radiation Exposure. Curr. Med. Chem. 2020. [Google Scholar] [CrossRef]
- Guo, L.; Zhu, Z.; Wang, G.; Cui, S.; Shen, M.; Song, Z.; Wang, J.H. microRNA-15b contributes to depression-like behavior in mice by affecting synaptic protein levels and function in the nucleus accumbens. J. Biol. Chem. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwivedi, Y. microRNA-124: A putative therapeutic target and biomarker for major depression. Expert Opin. Ther. Targets 2017, 21, 653–656. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.S.; Mu, R.H.; Li, C.F.; Dong, S.Q.; Geng, D.; Liu, Q.; Yi, L.T. microRNA-124 targets glucocorticoid receptor and is involved in depression-like behaviors. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 79, 417–425. [Google Scholar] [CrossRef]
- Gu, Z.; Pan, J.; Chen, L. MiR-124 suppression in the prefrontal cortex reduces depression-like behavior in mice. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Liu, M.; Zhang, Q.; Zhang, J.; Chen, J.; Suo, L.; Chen, Q. Knockdown of miR-124 reduces depression-like behavior in rats by targeting CREB1 and BDNF. Curr. Neurovascular Res. 2020, 17, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Lo Iacono, L.; Ielpo, D.; Accoto, A.; Di Segni, M.; Babicola, L.; D’Addario, S.L.; Ferlazzo, F.; Pascucci, T.; Ventura, R.; Andolina, D. MicroRNA-34a Regulates the Depression-like Behavior in Mice by Modulating the Expression of Target Genes in the Dorsal Raphe. Mol. Neurobiol. 2020, 57, 823–836. [Google Scholar] [CrossRef]
- Kuang, W.H.; Dong, Z.Q.; Tian, L.T.; Li, J. MicroRNA-451a, microRNA-34a-5p, and microRNA-221-3p as predictors of response to antidepressant treatment. Braz. J. Med. Biol. Res. Rev. Bras. Pesqui. Med. Biol. 2018, 51, e7212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arimoto-Matsuzaki, K.; Saito, H.; Takekawa, M. TIA1 oxidation inhibits stress granule assembly and sensitizes cells to stress-induced apoptosis. Nat. Commun. 2016, 7, 10252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apicco, D.J.; Ash, P.E.A.; Maziuk, B.; LeBlang, C.; Medalla, M.; Al Abdullatif, A.; Ferragud, A.; Botelho, E.; Ballance, H.I.; Dhawan, U.; et al. Reducing the RNA binding protein TIA1 protects against tau-mediated neurodegeneration in vivo. Nat. Neurosci. 2018, 21, 72–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderweyde, T.; Apicco, D.J.; Youmans-Kidder, K.; Ash, P.E.A.; Cook, C.; Lummertz da Rocha, E.; Jansen-West, K.; Frame, A.A.; Citro, A.; Leszyk, J.D.; et al. Interaction of tau with the RNA-Binding Protein TIA1 Regulates tau Pathophysiology and Toxicity. Cell Rep. 2016, 15, 1455–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arisi, I.; D’Onofrio, M.; Brandi, R.; Felsani, A.; Capsoni, S.; Drovandi, G.; Felici, G.; Weitschek, E.; Bertolazzi, P.; Cattaneo, A. Gene expression biomarkers in the brain of a mouse model for Alzheimer’s disease: Mining of microarray data by logic classification and feature selection. J. Alzheimer’s Dis. JAD 2011, 24, 721–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, A.; Zhang, H.; Chen, S.; Xia, F.; Yang, Y.; Dong, F.; Sun, D.; Xiong, S.; Zhang, J. miR-34a expands myeloid-derived suppressor cells via apoptosis inhibition. Exp. Cell Res. 2014, 326, 259–266. [Google Scholar] [CrossRef]
- Crowley, J.J.; Jones, M.D.; O’Leary, O.F.; Lucki, I. Automated tests for measuring the effects of antidepressants in mice. Pharmacol. Biochem. Behav. 2004, 78, 269–274. [Google Scholar] [CrossRef]
- Karalok, H.M.; Aydin, E.; Saglam, O.; Torun, A.; Guzeloglu-Kayisli, O.; Lalioti, M.D.; Kristiansson, H.; Duke, C.M.; Choe, G.; Flannery, C.; et al. mRNA-binding protein TIA-1 reduces cytokine expression in human endometrial stromal cells and is down-regulated in ectopic endometrium. J. Clin. Endocrinol. Metab. 2014, 99, E2610–E2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeBlang, C.J.; Medalla, M.; Nicoletti, N.W.; Hays, E.C.; Zhao, J.; Shattuck, J.; Cruz, A.L.; Wolozin, B.; Luebke, J.I. Reduction of the RNA Binding Protein TIA1 Exacerbates Neuroinflammation in Tauopathy. Front. Neurosci. 2020, 14, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, K.; Kedersha, N.; Shen, L.; Blackshear, P.J.; Anderson, P. Arthritis suppressor genes TIA-1 and TTP dampen the expression of tumor necrosis factor alpha, cyclooxygenase 2, and inflammatory arthritis. Proc. Natl. Acad. Sci. USA. 2004, 101, 2011–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piecyk, M.; Wax, S.; Beck, A.R.; Kedersha, N.; Gupta, M.; Maritim, B.; Chen, S.; Gueydan, C.; Kruys, V.; Streuli, M.; et al. TIA-1 is a translational silencer that selectively regulates the expression of TNF-alpha. EMBO J. 2000, 19, 4154–4163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Liu, T.; Huang, W.; Liu, H.; Zhang, H.M.; Li, Q.; Chen, Z.; Guo, A.Y. MicroRNA regulatory pathway analysis identifies miR-142-5p as a negative regulator of TGF-beta pathway via targeting SMAD3. Oncotarget 2016, 7, 71504–71513. [Google Scholar] [CrossRef] [Green Version]
- Zonenberg, A.; Leoniak, M.; Zarzycki, W. The effect of Chernobyl accident on the development of non malignant diseases. Endokrynol. Pol. 2006, 57, 38–44. [Google Scholar]
- Loganovsky, K.N.; Bomko, M.O.; Abramenko, I.V.; Kuts, K.V.; Belous, N.I.; Masiuk, S.V.; Gresko, M.V.; Loganovska, T.K.; Antypchuk, K.Y.; Perchuk, I.V.; et al. Neuropsychobiological mechanisms of affective and cognitive disorders in the chornobyl clean-up workers taking into account the specific gene polymorphisms. Probl. Radiatsiinoi Medytsyny Ta radiobiolohii 2018, 23, 373–409. [Google Scholar] [CrossRef]
- Aranha, M.M.; Santos, D.M.; Sola, S.; Steer, C.J.; Rodrigues, C.M. miR-34a regulates mouse neural stem cell differentiation. PLoS ONE 2011, 6, e21396. [Google Scholar] [CrossRef]
- Jauhari, A.; Singh, T.; Singh, P.; Parmar, D.; Yadav, S. Regulation of miR-34 Family in Neuronal Development. Mol. Neurobiol. 2018, 55, 936–945. [Google Scholar] [CrossRef]
- Jauhari, A.; Yadav, S. MiR-34 and MiR-200: Regulator of Cell Fate Plasticity and Neural Development. Neuromolecular Med. 2019, 21, 97–109. [Google Scholar] [CrossRef]
- Liang, T.Y.; Lou, J.Y. Increased Expression of mir-34a-5p and Clinical Association in Acute Ischemic Stroke Patients and in a Rat Model. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2016, 22, 2950–2955. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Liu, Y.; Wang, X.; Wu, J.; Liu, K.; Zhou, J.; Liu, L.; Zhang, C. Identification of differential microRNAs in cerebrospinal fluid and serum of patients with major depressive disorder. PLoS ONE 2015, 10, e0121975. [Google Scholar] [CrossRef]
- Cosín-Tomás, M.; Antonell, A.; Lladó, A.; Alcolea, D.; Fortea, J.; Ezquerra, M.; Lleó, A.; Martí, M.J.; Pallàs, M.; Sanchez-Valle, R.; et al. Plasma miR-34a-5p and miR-545-3p as Early Biomarkers of Alzheimer’s Disease: Potential and Limitations. Mol. Neurobiol. 2017, 54, 5550–5562. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Xiao, H.; Li, Y.; Dong, J.; Luo, D.; Li, H.; Feng, G.; Wang, H.; Fan, S. Total abdominal irradiation exposure impairs cognitive function involving miR-34a-5p/BDNF axis. Biochim. Biophys. Acta Mol. Basis. Dis. 2017, 1863, 2333–2341. [Google Scholar] [CrossRef]
- Joilin, G.; Guévremont, D.; Ryan, B.; Claudianos, C.; Cristino, A.S.; Abraham, W.C.; Williams, J.M. Rapid regulation of microRNA following induction of long-term potentiation in vivo. Front. Mol. Neurosci. 2014, 7, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.J.; Weng, S.L.; Hsieh, J.Y.; Wang, T.Y.; Chang, M.D.; Wang, H.W. MicroRNA-34a modulates genes involved in cellular motility and oxidative phosphorylation in neural precursors derived from human umbilical cord mesenchymal stem cells. BMC Med. Genom. 2011, 4, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fineberg, S.K.; Datta, P.; Stein, C.S.; Davidson, B.L. MiR-34a represses Numbl in murine neural progenitor cells and antagonizes neuronal differentiation. PLoS ONE 2012, 7, e38562. [Google Scholar] [CrossRef]
- Morgado, A.L.; Xavier, J.M.; Dionísio, P.A.; Ribeiro, M.F.; Dias, R.B.; Sebastião, A.M.; Solá, S.; Rodrigues, C.M. MicroRNA-34a Modulates Neural Stem Cell Differentiation by Regulating Expression of Synaptic and Autophagic Proteins. Mol. Neurobiol. 2015, 51, 1168–1183. [Google Scholar] [CrossRef] [PubMed]
- Namgyal, D.; Chandan, K.; Sultan, A.; Aftab, M.; Ali, S.; Mehta, R.; El-Serehy, H.A.; Al-Misned, F.A.; Sarwat, M. Dim Light at Night Induced Neurodegeneration and Ameliorative Effect of Curcumin. Cells 2020, 9, 2093. [Google Scholar] [CrossRef] [PubMed]
- Welch, C.; Chen, Y.; Stallings, R.L. MicroRNA-34a functions as a potential tumor suppressor by inducing apoptosis in neuroblastoma cells. Oncogene 2007, 26, 5017–5022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Zheng, Y.L.; Yin, X.; Xu, S.J.; Tian, D.; Zhang, C.Y.; Wang, S.; Ma, J.Z. Excessive Treadmill Training Enhances Brain-Specific MicroRNA-34a in the Mouse Hippocampus. Front. Mol. Neurosci. 2020, 13, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Córdova-Rivas, S.; Fraire-Soto, I.; Mercado-Casas Torres, A.; Servín-González, L.S.; Granados-López, A.J.; López-Hernández, Y.; Reyes-Estrada, C.A.; Gutiérrez-Hernández, R.; Castañeda-Delgado, J.E.; Ramírez-Hernández, L.; et al. 5p and 3p Strands of miR-34 Family Members Have Differential Effects in Cell Proliferation, Migration, and Invasion in Cervical Cancer Cells. Int. J. Mol. Sci. 2019, 20, 545. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Hao, L.; Yang, H.; Chen, J.; Liu, J. miRNA-34a suppresses colon carcinoma proliferation and induces cell apoptosis by targeting SYT1. Int. J. Clin. Exp. Pathol. 2019, 12, 2887–2897. [Google Scholar] [PubMed]
- Wang, B.; He, G.; Xu, G.; Wen, J.; Yu, X. miRNA-34a inhibits cell adhesion by targeting CD44 in human renal epithelial cells: Implications for renal stone disease. Urolithiasis 2020, 48, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Kiang, J.G.; Smith, J.T.; Anderson, M.N.; Elliott, T.B.; Gupta, P.; Balakathiresan, N.S.; Maheshwari, R.K.; Knollmann-Ritschel, B. Hemorrhage enhances cytokine, complement component 3, and caspase-3, and regulates microRNAs associated with intestinal damage after whole-body gamma-irradiation in combined injury. PLoS ONE 2017, 12, e0184393. [Google Scholar] [CrossRef]
- Ji, Q.; Han, J.; Wang, L.; Liu, J.; Dong, Y.; Zhu, K.; Shi, L. MicroRNA-34a promotes apoptosis of retinal vascular endothelial cells by targeting SIRT1 in rats with diabetic retinopathy. Cell Cycle. 2020, 19, 1–11. [Google Scholar] [CrossRef]
- Khanna, A.; Muthusamy, S.; Liang, R.; Sarojini, H.; Wang, E. Gain of survival signaling by down-regulation of three key miRNAs in brain of calorie-restricted mice. Aging 2011, 3, 223–236. [Google Scholar] [CrossRef] [Green Version]
- Periyasamy, P.; Thangaraj, A.; Bendi, V.S.; Buch, S. HIV-1 Tat-mediated microglial inflammation involves a novel miRNA-34a-NLRC5-NFκB signaling axis. Brain Behav. Immun. 2019, 80, 227–237. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, Y.; Qi, L.; Ding, L.; Jiang, H.; Yu, H. NFIX Circular RNA Promotes Glioma Progression by Regulating miR-34a-5p via Notch Signaling Pathway. Front. Mol. Neurosci. 2018, 11, 225. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Fu, T.; Zhao, S.; Gao, M. MicroRNA-34a-5p suppresses tumorigenesis and progression of glioma and potentiates Temozolomide-induced cytotoxicity for glioma cells by targeting HMGA2. Eur. J. Pharmacol. 2019, 852, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Chen, P.; Wang, X.; Yao, J.; Zhuang, S. miR-34a deficiency in APP/PS1 mice promotes cognitive function by increasing synaptic plasticity via AMPA and NMDA receptors. Neurosci. Lett. 2018, 670, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Rayman, J.B.; Hijazi, J.; Li, X.; Kedersha, N.; Anderson, P.J.; Kandel, E.R. Genetic Perturbation of TIA1 Reveals a Physiological Role in Fear Memory. Cell Rep. 2019, 26, 2970–2983.e2974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowin, B.; French, L.; Martinou, J.C.; Tschopp, J. Expression of the CTL-associated protein TIA-1 during murine embryogenesis. J. Immunol. 1996, 157, 1448–1454. [Google Scholar] [PubMed]
- Bi, J.W.; Zou, Y.L.; Qian, J.T.; Chen, W.B. MiR-599 serves a suppressive role in anaplastic thyroid cancer by activating the T-cell intracellular antigen. Exp. Ther. Med. 2019, 18, 2413–2420. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, R.; Yang, F.; Cheng, R.; Chen, X.; Cui, S.; Gu, Y.; Sun, W.; You, C.; Liu, Z.; et al. miR-19a promotes colorectal cancer proliferation and migration by targeting TIA1. Mol. Cancer 2017, 16, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Wang, M.; Lin, B.; Yao, D.; Li, J.; Tang, X.; Li, S.; Liu, Y.; Xie, R.; Yu, S. miR-487a promotes progression of gastric cancer by targeting TIA1. Biochime 2018, 154, 119–126. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Ma, Z.; Shen, H.; Wu, Z.; Liu, L.; Ren, B.; Wong, P.; Sethi, G.; Tang, F. Early Life Irradiation-Induced Hypoplasia and Impairment of Neurogenesis in the Dentate Gyrus and Adult Depression Are Mediated by MicroRNA- 34a-5p/T-Cell Intracytoplasmic Antigen-1 Pathway. Cells 2021, 10, 2476. https://doi.org/10.3390/cells10092476
Wang H, Ma Z, Shen H, Wu Z, Liu L, Ren B, Wong P, Sethi G, Tang F. Early Life Irradiation-Induced Hypoplasia and Impairment of Neurogenesis in the Dentate Gyrus and Adult Depression Are Mediated by MicroRNA- 34a-5p/T-Cell Intracytoplasmic Antigen-1 Pathway. Cells. 2021; 10(9):2476. https://doi.org/10.3390/cells10092476
Chicago/Turabian StyleWang, Hong, Zhaowu Ma, Hongyuan Shen, Zijun Wu, Lian Liu, Boxu Ren, Peiyan Wong, Gautam Sethi, and Fengru Tang. 2021. "Early Life Irradiation-Induced Hypoplasia and Impairment of Neurogenesis in the Dentate Gyrus and Adult Depression Are Mediated by MicroRNA- 34a-5p/T-Cell Intracytoplasmic Antigen-1 Pathway" Cells 10, no. 9: 2476. https://doi.org/10.3390/cells10092476
APA StyleWang, H., Ma, Z., Shen, H., Wu, Z., Liu, L., Ren, B., Wong, P., Sethi, G., & Tang, F. (2021). Early Life Irradiation-Induced Hypoplasia and Impairment of Neurogenesis in the Dentate Gyrus and Adult Depression Are Mediated by MicroRNA- 34a-5p/T-Cell Intracytoplasmic Antigen-1 Pathway. Cells, 10(9), 2476. https://doi.org/10.3390/cells10092476