
Osthole Inhibits Expression of Genes Associated with Toll-like Receptor 2 Signaling Pathway in an Organotypic 3D Skin Model of Human Epidermis with Atopic Dermatitis




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture
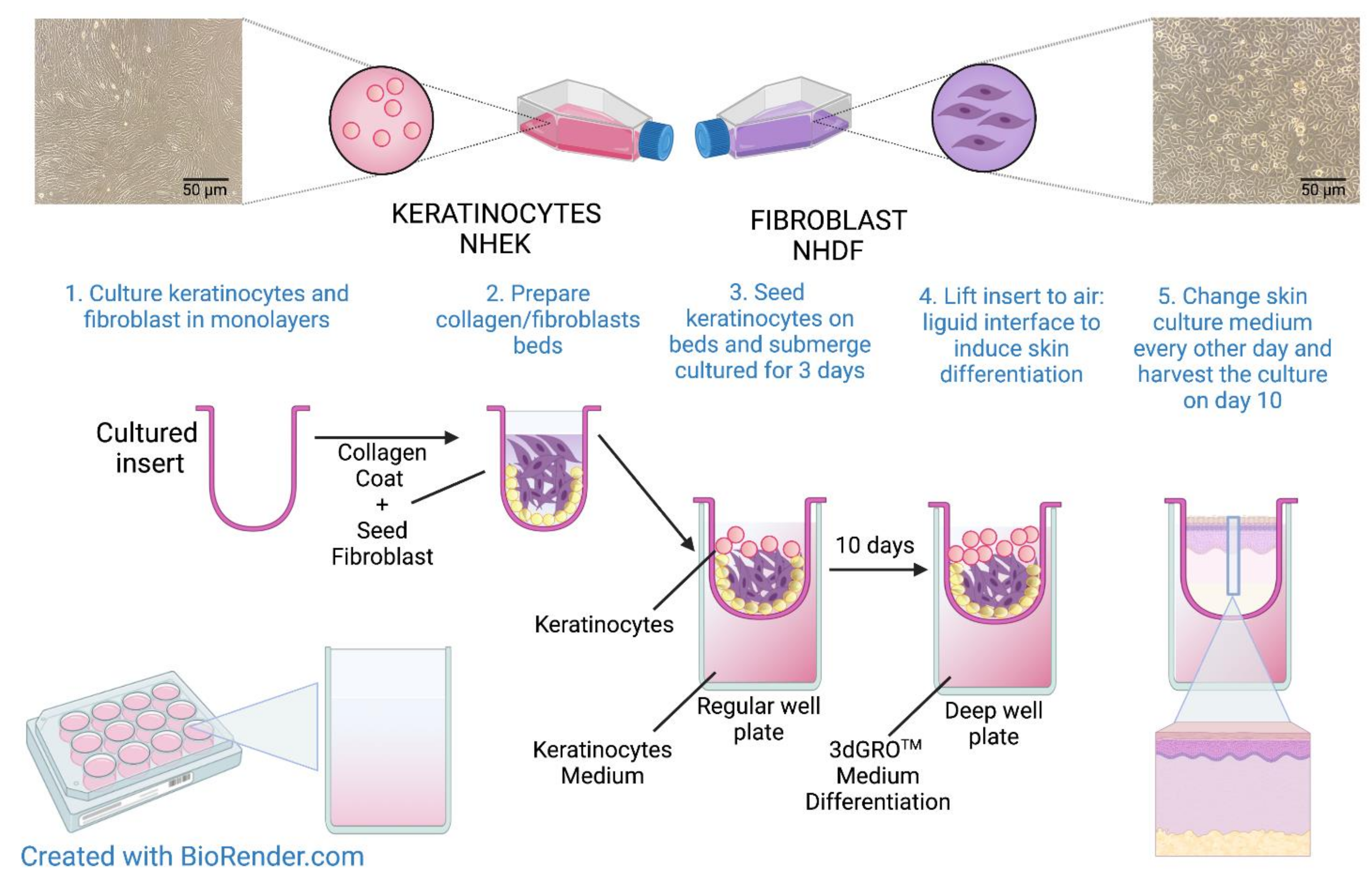
2.3. Organotypic 3D Skin Model
2.4. Incubation of the Organotypic 3D Skin Model with the Investigated Substances
2.5. RNA Isolation and Reverse Transcription
2.6. Quantitative Real-Time PCR (qPCR) and Data Analysis
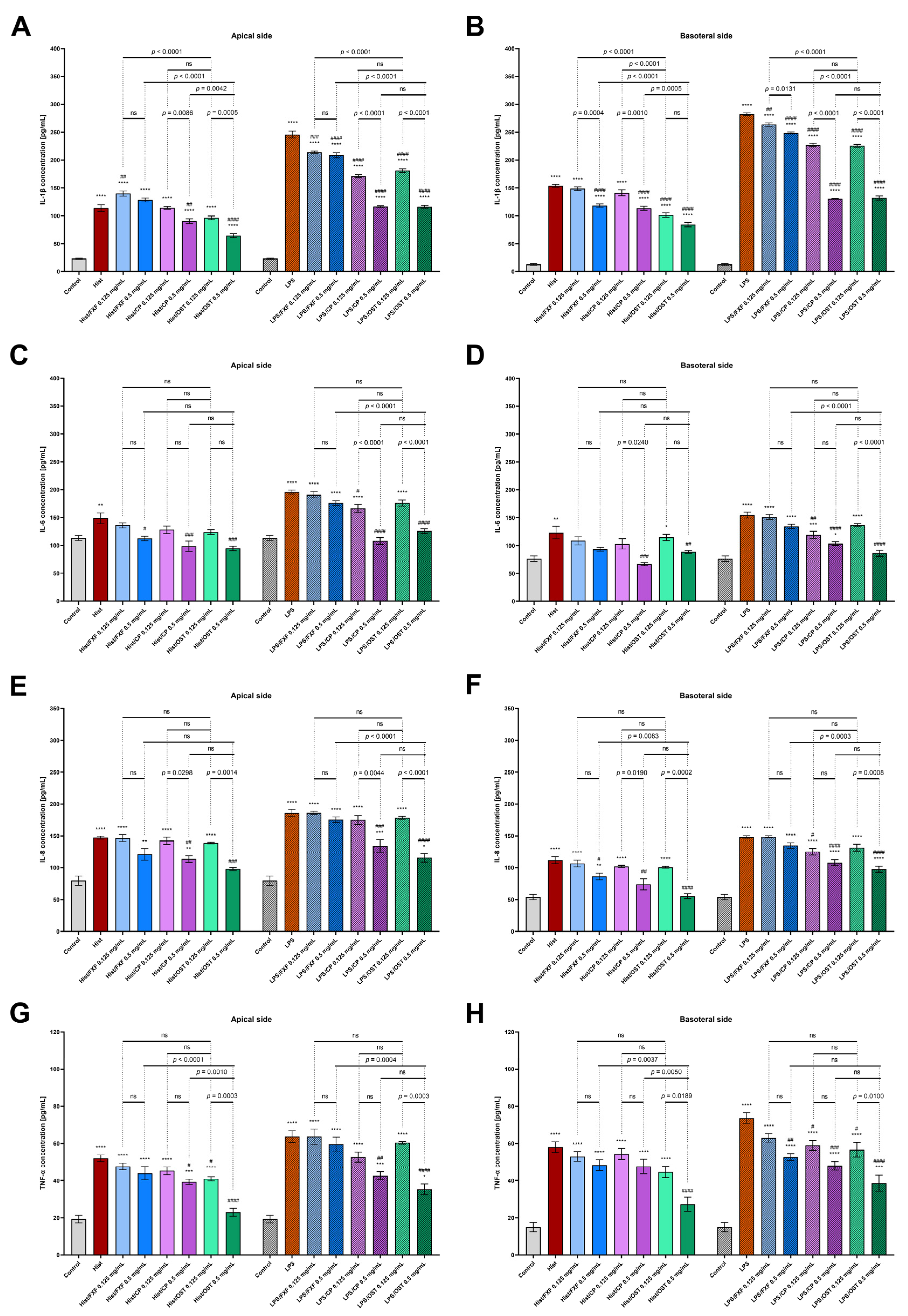
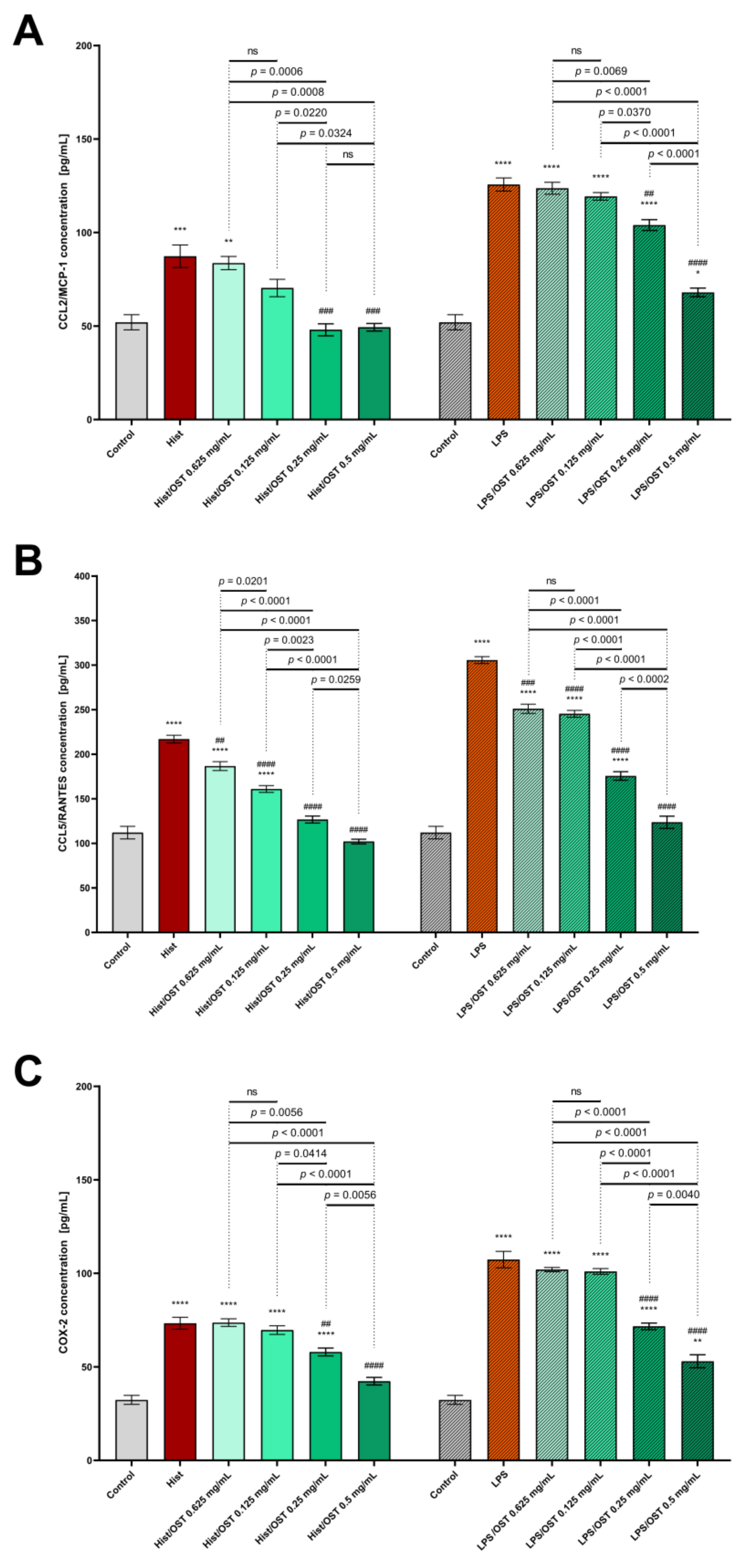
2.7. Analysis of Cytokine and Chemokine Levels
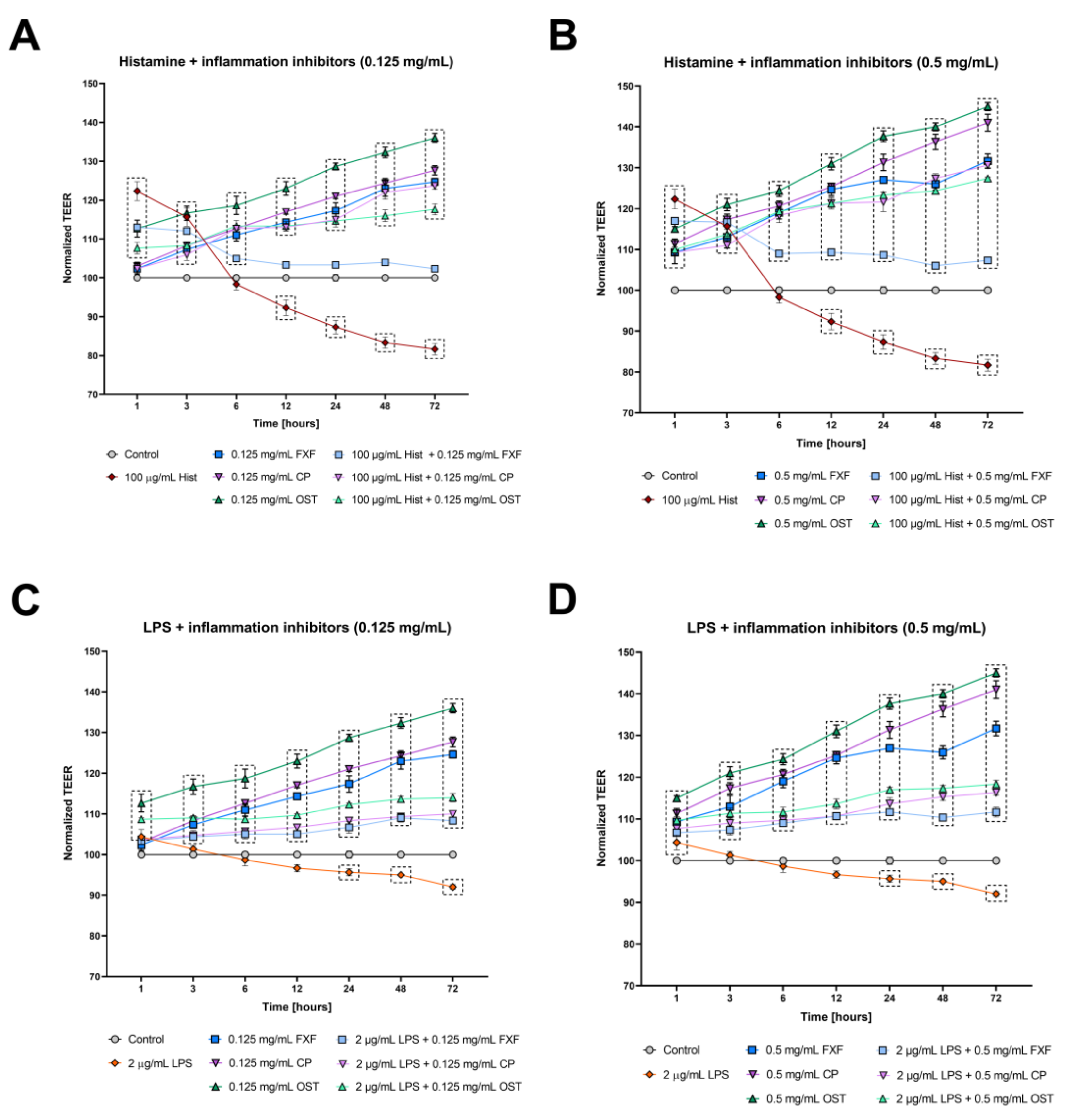
2.8. Measurement of Transepithelial Electrical Resistance
2.9. Statistical Analysis
3. Results
3.1. Osthole Prevents Histamine- and LPS-Induced Disruption of Tight Junctions in a 3D Organotypic Skin Model
3.2. Osthole Decreases Secretion of Pro-Inflammatory Cytokines, Chemokines and Proteins in a 3D Organotypic Skin Model Treated with Histamine and LPS
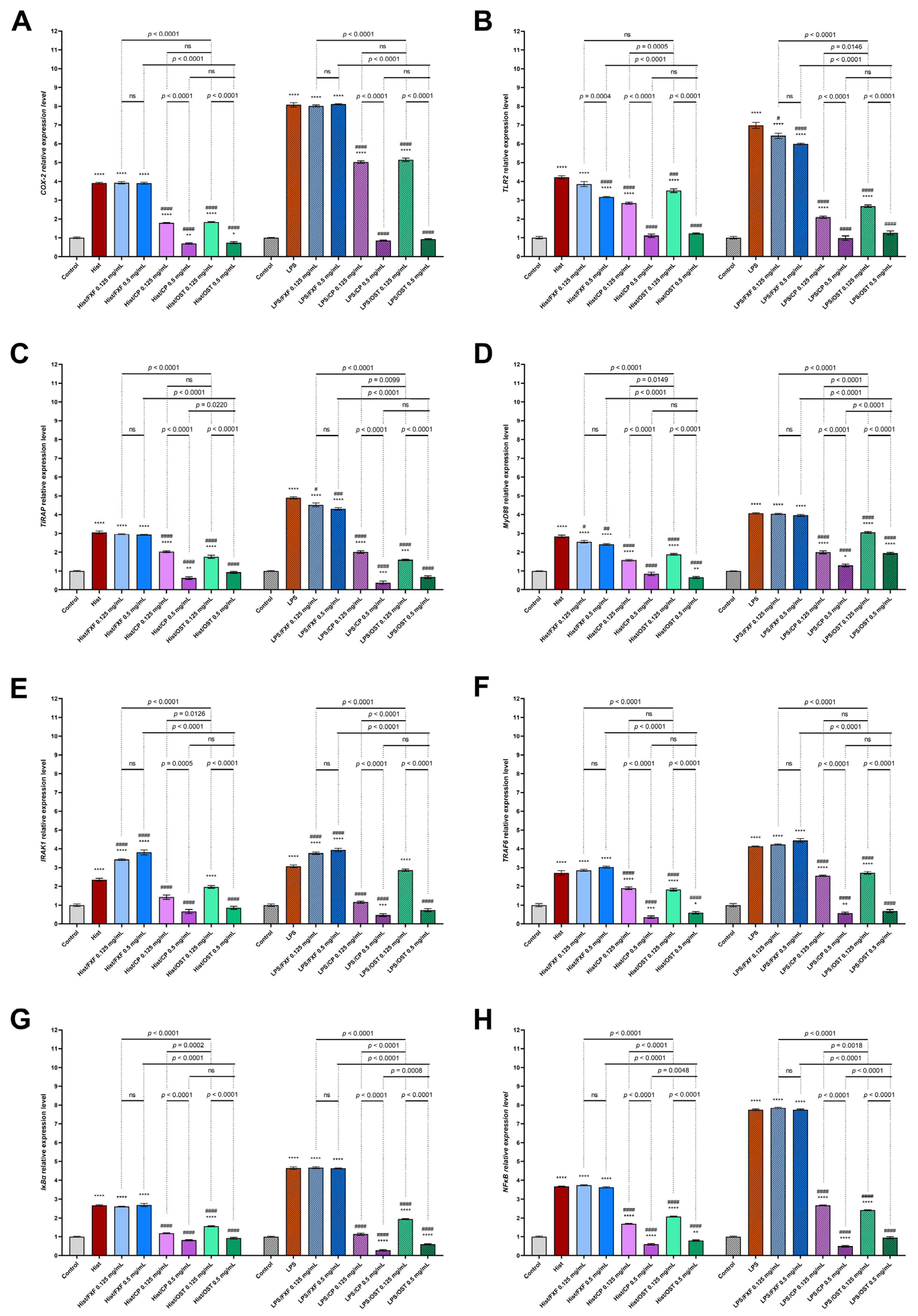
3.3. Osthole Regulates the Expression of TLR2, TIRAP, MyD88, IRAK1, TRAF6, IκBα and NFκB in Histamine/LPS-Induced Organotypic 3D Skin Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | Atopic dermatitis |
| CCL2/MCP-1 | Chemokine (C-C motif) ligand 2/monocyte chemoattractant protein 1 |
| CCL5/RANTES | Chemokine (C-C motif) ligand 5/(regulated on activation, normal T cell expressed and secreted |
| ChKs | Chemokines |
| CKs | Cytokines |
| COX-2 | Cyclooxygenase 2 |
| CP | Clobetasol propionate |
| DMSO | Dimethyl sulfoxide |
| ELISA | Enzyme-linked immunosorbent assay |
| FXF | Fexofenadine hydrochloride |
| IFN | Interferon |
| IRAK1 | Interleukin receptor-associated kinase 1 |
| IκB⍺ | NFκB inhibitor alpha |
| LPS | Lipopolysaccharides |
| MyD88 | Myeloid differentiation protein 88 |
| NFκB | Nuclear factor kappa B |
| NHDF | Normal human dermal fibroblasts |
| NHEK | Normal human epidermal keratinocytes |
| PBMCs | Peripheral blood mononuclear cells |
| TEER | Transepithelial electrical resistance |
| TIRAP | TIR domain containing adaptor protein |
| TLR2 | Toll-like receptor 2 |
| TNF-α | Tumor necrosis factor alpha |
| TRAF6 | TNF receptor-associated factor 6 |
References
- Kim, C.; Park, K.Y.; Ahn, S.; Kim, D.H.; Li, K.; Kim, D.W.; Kim, M.-B.; Jo, S.-J.; Yim, H.W.; Seo, S.J. Economic Impact of Atopic Dermatitis in Korean Patients. Ann. Dermatol. 2015, 27, 298–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gittler, J.K.; Krueger, J.G.; Guttman-Yassky, E. Atopic dermatitis results in intrinsic barrier and immune abnormalities: Implications for contact dermatitis. J. Allergy Clin. Immunol. 2013, 131, 300–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nettis, E.; Ortoncelli, M.; Pellacani, G.; Foti, C.; Di Leo, E.; Patruno, C.; Rongioletti, F.; Argenziano, G.; Ferrucci, S.M.; Macchia, L.; et al. A Multicenter Study on the Prevalence of Clinical Patterns and Clinical Phenotypes in Adult Atopic Dermatitis. J. Investig. Allergol. Clin. Immunol. 2020, 30, 448–450. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, S.; Novak, N. Atopic dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Nomura, I.; Gao, B.; Boguniewicz, M.; Darst, M.A.; Travers, J.B.; Leung, D.Y. Distinct patterns of gene expression in the skin lesions of atopic dermatitis and psoriasis: A gene microarray analysis. J. Allergy Clin. Immunol. 2003, 112, 1195–1202. [Google Scholar] [CrossRef]
- Han, H.; Roan, F.; Ziegler, S.F. The atopic march: Current insights into skin barrier dysfunction and epithelial cell-derived cytokines. Immunol. Rev. 2017, 278, 116–130. [Google Scholar] [CrossRef]
- Werfel, T.; Allam, J.-P.; Biedermann, T.; Eyerich, K.; Gilles, S.; Guttman-Yassky, E.; Hoetzenecker, W.; Knol, E.; Simon, H.-U.; Wollenberg, A.; et al. Cellular and molecular immunologic mechanisms in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2016, 138, 336–349. [Google Scholar] [CrossRef] [Green Version]
- Wollenberg, A.; Barbarot, S.; Bieber, T.; Christen-Zaech, S.; Deleuran, M.; Fink-Wagner, A.; Gieler, U.; Girolomoni, G.; Lau, S.; Muraro, A.; et al. Consensus-based European guidelines for treatment of atopic eczema (atopic dermatitis) in adults and children: Part I. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 657–682. [Google Scholar] [CrossRef] [Green Version]
- Dattola, A.; Bennardo, L.; Silvestri, M.; Nisticò, S.P. What’s new in the treatment of atopic dermatitis? Dermatol. Ther. 2019, 32, e12787. [Google Scholar] [CrossRef]
- Sato, M.; Yamamoto-Hanada, K.; Yang, L.; Irahara, M.; Ishikawa, F.; Iwama-Mitsui, M.; Saito-Abe, M.; Miyaji, Y.; Inagaki, S.; Fukuie, T.; et al. Complementary and alternative medicine and atopic dermatitis in children. J. Dermatol. Sci. 2020, 97, 80–82. [Google Scholar] [CrossRef]
- Tyurin, Y.A.; Shamsutdinov, A.F.; Kalinin, N.N.; Sharifullina, A.A.; Reshetnikova, I.D. Association of Toll-Like Cell Receptors TLR2 (p.Arg753GLN) and TLR4 (p.Asp299GLY) Polymorphisms with Indicators of General and Local Immunity in Patients with Atopic Dermatitis. J. Immunol. Res. 2017, 2017, 8493545. [Google Scholar] [CrossRef]
- Salpietro, C.; Rigoli, L.; Miraglia Del Giudice, M.; Cuppari, C.; Di Bella, C.; Salpietro, A.; Maiello, N.; La Rosa, M.; Marseglia, G.L.; Leonardi, S.; et al. TLR2 and TLR4 gene polymorphisms and atopic dermatitis in Italian children: A multicenter study. Int. J. Immunopathol. Pharmacol. 2011, 24, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Hasannejad, H.; Takahashi, R.; Kimishima, M.; Hayakawa, K.; Shiohara, T. Selective impairment of Toll-like receptor 2-mediated proinflammatory cytokine production by monocytes from patients with atopic dermatitis. J. Allergy Clin. Immunol. 2007, 120, 69–75. [Google Scholar] [CrossRef]
- Panzer, R.; Blobel, C.; Fölster-Holst, R.; Proksch, E. TLR2 and TLR4 expression in atopic dermatitis, contact dermatitis and psoriasis. Exp. Dermatol. 2014, 23, 364–366. [Google Scholar] [CrossRef]
- Anwar, M.A.; Basith, S.; Choi, S. Negative regulatory approaches to the attenuation of Toll-like receptor signaling. Exp. Mol. Med. 2013, 45, e11. [Google Scholar] [CrossRef]
- Guo, J.; Liang, W.; Li, J.; Long, J. Knockdown of FSTL1 inhibits oxLDL-induced inflammation responses through the TLR4/MyD88/NF-κB and MAPK pathway. Biochem. Biophys. Res. Commun. 2016, 478, 1528–1533. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, D.; Xi, Y.; Li, J.; Liu, B.; Li, J. Upregulation of miRNA-140-5p inhibits inflammatory cytokines in acute lung injury through the MyD88/NF-κB signaling pathway by targeting TLR4. Exp. Ther. Med. 2018, 16, 3913–3920. [Google Scholar] [CrossRef]
- Shokoohinia, Y.; Jafari, F.; Mohammadi, Z.; Bazvandi, L.; Hosseinzadeh, L.; Chow, N.; Bhattacharyya, P.; Farzaei, M.H.; Farooqi, A.A.; Nabavi, S.M.; et al. Potential Anticancer Properties of Osthol: A Comprehensive Mechanistic Review. Nutrients 2018, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Ayudhya, C.C.N.; Thapaliya, M.; Deepak, V.; Ali, H. Multifaceted MRGPRX2: New insight into the role of mast cells in health and disease. J. Allergy Clin. Immunol. 2021, 148, 293–308. [Google Scholar] [CrossRef]
- Kordulewska, N.K.; Cieślińska, A.; Fiedorowicz, E.; Jarmołowska, B.; Piskorz-Ogórek, K.; Kostyra, E. Cytokines concentrations in serum samples from allergic children-Multiple analysis to define biomarkers for better diagnosis of allergic inflammatory process. Immunobiology 2018, 223, 648–657. [Google Scholar] [CrossRef]
- Kordulewska, N.K.; Cieślińska, A.; Fiedorowicz, E.; Jarmołowska, B.; Kostyra, E. High Expression of IL-1RI and EP₂ Receptors in the IL-1β/COX-2 Pathway, and a New Alternative to Non-Steroidal Drugs-Osthole in Inhibition COX-2. Int. J. Mol. Sci. 2019, 20, 186. [Google Scholar] [CrossRef] [Green Version]
- Kordulewska, N.K.; Kostyra, E.; Chwała, B.; Moszyńska, M.; Cieślińska, A.; Fiedorowicz, E.; Jarmołowska, B. A novel concept of immunological and allergy interactions in autism spectrum disorders: Molecular, anti-inflammatory effect of osthole. Int. Immunopharmacol. 2019, 72, 1–11. [Google Scholar] [CrossRef]
- Kordulewska, N.K.; Kostyra, E.; Cieślińska, A.; Fiedorowicz, E.; Jarmołowska, B. Cytokine production by PBMC and serum from allergic and non-allergic subjects following in vitro histamine stimulation to test fexofenadine and osthole anti-allergic properties. Eur. J. Pharmacol. 2016, 791, 763–772. [Google Scholar] [CrossRef]
- Kordulewska, N.K.; Kostyra, E.; Cieślińska, A.; Matysiewicz, M.; Fiedorowicz, E.; Sienkiewicz-Szłapka, E. Changes in gene expression induced by histamine, fexofenadine and osthole: Expression of histamine H1 receptor, COX-2, NF-κB, CCR1, chemokine CCL5/RANTES and interleukin-1β in PBMC allergic and non-allergic patients. Immunobiology 2017, 222, 571–581. [Google Scholar] [CrossRef]
- Kordulewska, N.K.; Kostyra, E.; Matysiewicz, M.; Cieślińska, A.; Jarmołowska, B. Impact of fexofenadine, osthole and histamine on peripheral blood mononuclear cell proliferation and cytokine secretion. Eur. J. Pharmacol. 2015, 761, 254–261. [Google Scholar] [CrossRef]
- Kordulewska, N.K.; Kostyra, E.; Piskorz-Ogórek, K.; Moszyńska, M.; Cieślińska, A.; Fiedorowicz, E.; Jarmołowska, B. Serum cytokine levels in children with spectrum autism disorder: Differences in pro- and anti-inflammatory balance. J. Neuroimmunol. 2019, 337, 577066. [Google Scholar] [CrossRef]
- Fan, H.; Gao, Z.; Ji, K.; Li, X.; Wu, J.; Liu, Y.; Wang, X.; Liang, H.; Liu, Y.; Li, X.; et al. The in vitro and in vivo anti-inflammatory effect of osthole, the major natural coumarin from Cnidium monnieri (L.) Cuss, via the blocking of the activation of the NF-κB and MAPK/p38 pathways. Phytomedicine 2019, 58, 152864. [Google Scholar] [CrossRef]
- Yang, S.-M.; Chan, Y.-L.; Hua, K.-F.; Chang, J.-M.; Chen, H.-L.; Tsai, Y.-J.; Hsu, Y.-J.; Chao, L.K.; Feng-Ling, Y.; Tsai, Y.-L.; et al. Osthole improves an accelerated focal segmental glomerulosclerosis model in the early stage by activating the Nrf2 antioxidant pathway and subsequently inhibiting NF-κB-mediated COX-2 expression and apoptosis. Free Radic. Biol. Med. 2014, 73, 260–269. [Google Scholar] [CrossRef]
- Cai, Y.; Sun, W.; Zhang, X.-X.; Lin, Y.-D.; Chen, H.; Li, H. Osthole prevents acetaminophen-induced liver injury in mice. Acta Pharmacol. Sin. 2018, 39, 74–84. [Google Scholar] [CrossRef]
- Jiao, Y.; Kong, L.; Yao, Y.; Li, S.; Tao, Z.; Yan, Y.; Yang, J. Osthole decreases beta amyloid levels through up-regulation of miR-107 in Alzheimer’s disease. Neuropharmacology 2016, 108, 332–344. [Google Scholar] [CrossRef]
- Tang, D.-Z.; Hou, W.; Zhou, Q.; Zhang, M.; Holz, J.; Sheu, T.-J.; Li, T.-F.; Cheng, S.-D.; Shi, Q.; Harris, S.E.; et al. Osthole stimulates osteoblast differentiation and bone formation by activation of beta-catenin-BMP signaling. J. Bone Miner. Res. 2010, 25, 1234–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueroa, M.; Rivero-Cruz, I.; Rivero-Cruz, B.; Bye, R.; Navarrete, A.; Mata, R. Constituents, biological activities and quality control parameters of the crude extract and essential oil from Arracacia tolucensis var. multifida. J. Ethnopharmacol. 2007, 113, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Hener, P.; Zhang, Z.; Kato, S.; Metzger, D.; Chambon, P. Topical vitamin D3 and low-calcemic analogs induce thymic stromal lymphopoietin in mouse keratinocytes and trigger an atopic dermatitis. Proc. Natl. Acad. Sci. USA 2006, 103, 11736–11741. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Hener, P.; Frossard, N.; Kato, S.; Metzger, D.; Li, M.; Chambon, P. Thymic stromal lymphopoietin overproduced by keratinocytes in mouse skin aggravates experimental asthma. Proc. Natl. Acad. Sci. USA 2009, 106, 1536–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, J.; Omori, M.; Gyarmati, D.; Zhou, B.; Aye, T.; Brewer, A.; Comeau, M.R.; Campbell, D.J.; Ziegler, S.F. Spontaneous atopic dermatitis in mice expressing an inducible thymic stromal lymphopoietin transgene specifically in the skin. J. Exp. Med. 2005, 202, 541–549. [Google Scholar] [CrossRef]
- Nakajima, S.; Igyártó, B.Z.; Honda, T.; Egawa, G.; Otsuka, A.; Hara-Chikuma, M.; Watanabe, N.; Ziegler, S.F.; Tomura, M.; Inaba, K.; et al. Langerhans cells are critical in epicutaneous sensitization with protein antigen via thymic stromal lymphopoietin receptor signaling. J. Allergy Clin. Immunol. 2012, 129, 1048–1055.e6. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [Green Version]
- Raymond, S.L.; López, M.C.; Baker, H.V.; Larson, S.D.; Efron, P.A.; Sweeney, T.E.; Khatri, P.; Moldawer, L.L.; Wynn, J.L. Unique transcriptomic response to sepsis is observed among patients of different age groups. PLoS ONE 2017, 12, e0184159. [Google Scholar] [CrossRef]
- Kordulewska, N.K.; Topa, J.; Tańska, M.; Cieślińska, A.; Fiedorowicz, E.; Savelkoul, H.F.J.; Jarmołowska, B. Modulatory Effects of Osthole on Lipopolysaccharides-Induced Inflammation in Caco-2 Cell Monolayer and Co-Cultures with THP-1 and THP-1-Derived Macrophages. Nutrients 2020, 13, 123. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 45e. [Google Scholar] [CrossRef]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER measurement techniques for in vitro barrier model systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef] [Green Version]
- Murayama, Y.; Sakuma, T.; Udagawa, H.; Utsunomiya, J.; Okamoto, R.; Asano, K. Sex hormone-binding globulin and estrogen receptor in breast cancer: Technique and preliminary clinical results. J. Clin. Endocrinol. Metab. 1978, 46, 998–1006. [Google Scholar] [CrossRef]
- Norris, D.A. Mechanisms of action of topical therapies and the rationale for combination therapy. J. Am. Acad. Dermatol. 2005, 53, S17–S25. [Google Scholar] [CrossRef]
- Kim, B.E.; Leung, D.Y.M. Significance of Skin Barrier Dysfunction in Atopic Dermatitis. Allergy. Asthma Immunol. Res. 2018, 10, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Hajar, T.; Gontijo, J.R.V.; Hanifin, J.M. New and developing therapies for atopic dermatitis. An. Bras. Dermatol. 2018, 93, 104–107. [Google Scholar] [CrossRef] [Green Version]
- Man, G.; Hu, L.-Z.; Elias, P.M.; Man, M.-Q. Therapeutic Benefits of Natural Ingredients for Atopic Dermatitis. Chin. J. Integr. Med. 2018, 24, 308–314. [Google Scholar] [CrossRef]
- Meyle, J.; Gültig, K.; Rascher, G.; Wolburg, H. Transepithelial electrical resistance and tight junctions of human gingival keratinocytes. J. Periodontal Res. 1999, 34, 214–222. [Google Scholar] [CrossRef]
- Ollmar, S.; Eek, A.; Sundström, F.; Emtestam, L. Electrical impedance for estimation of irritation in oral mucosa and skin. Med. Prog. Technol. 1995, 21, 29–37. [Google Scholar]
- Trautmann, A.; Akdis, M.; Schmid-Grendelmeier, P.; Disch, R.; Bröcker, E.B.; Blaser, K.; Akdis, C.A. Targeting keratinocyte apoptosis in the treatment of atopic dermatitis and allergic contact dermatitis. J. Allergy Clin. Immunol. 2001, 108, 839–846. [Google Scholar] [CrossRef]
- Leyva-Castillo, J.M.; Hener, P.; Jiang, H.; Li, M. TSLP produced by keratinocytes promotes allergen sensitization through skin and thereby triggers atopic march in mice. J. Investig. Dermatol. 2013, 133, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Niebuhr, M.; Lutat, C.; Sigel, S.; Werfel, T. Impaired TLR-2 expression and TLR-2-mediated cytokine secretion in macrophages from patients with atopic dermatitis. Allergy 2009, 64, 1580–1587. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, Y.; Zhang, J.; Dou, X.; Yang, H.; Shao, Y.; Wang, K.; Yu, B.; Zhang, W.; Lau, H.Y.A. Impaired Toll-like receptor 2-mediated Th1 and Th17/22 cytokines secretion in human peripheral blood mononuclear cells from patients with atopic dermatitis. J. Transl. Med. 2015, 13, 384. [Google Scholar] [CrossRef] [Green Version]
- Kaesler, S.; Volz, T.; Skabytska, Y.; Köberle, M.; Hein, U.; Chen, K.-M.; Guenova, E.; Wölbing, F.; Röcken, M.; Biedermann, T. Toll-like receptor 2 ligands promote chronic atopic dermatitis through IL-4-mediated suppression of IL-10. J. Allergy Clin. Immunol. 2014, 134, 92–99. [Google Scholar] [CrossRef]
- Jin, H.; Kumar, L.; Mathias, C.; Zurakowski, D.; Oettgen, H.; Gorelik, L.; Geha, R. Toll-like receptor 2 is important for the T(H)1 response to cutaneous sensitization. J. Allergy Clin. Immunol. 2009, 123, 875–882.e1. [Google Scholar] [CrossRef] [Green Version]
- Horng, T.; Barton, G.M.; Flavell, R.A.; Medzhitov, R. The adaptor molecule TIRAP provides signalling specificity for Toll-like receptors. Nature 2002, 420, 329–333. [Google Scholar] [CrossRef]
- Yamamoto, M.; Sato, S.; Hemmi, H.; Sanjo, H.; Uematsu, S.; Kaisho, T.; Hoshino, K.; Takeuchi, O.; Kobayashi, M.; Fujita, T.; et al. Essential role for TIRAP in activation of the signalling cascade shared by TLR2 and TLR4. Nature 2002, 420, 324–329. [Google Scholar] [CrossRef]
- Bernard, N.J.; O’Neill, L.A. Mal, more than a bridge to MyD88. IUBMB Life 2013, 65, 777–786. [Google Scholar] [CrossRef]
- Tao, Y.; Wang, Y.; Wang, X.; Wang, C.; Bao, K.; Ji, L.; Jiang, G.; Hong, M. Calycosin Suppresses Epithelial Derived Initiative Key Factors and Maintains Epithelial Barrier in Allergic Inflammation via TLR4 Mediated NF-κB Pathway. Cell. Physiol. Biochem. 2017, 44, 1106–1119. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Guan, X.-K.; Liu, R.-X. Role of Chinese Herbal Medicines in Regulation of Energy Metabolism in Treating Cardiovascular Diseases. Chin. J. Integr. Med. 2019, 25, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Gallo, R. Toll-Like Receptors in Skin Infections and Inflammatory Diseases. Infect. Disord. Drug Targets 2008, 8, 144–155. [Google Scholar] [CrossRef]
- Jiao, D.; Wong, C.-K.; Qiu, H.-N.; Dong, J.; Cai, Z.; Chu, M.; Hon, K.-L.; Tsang, M.S.-M.; Lam, C.W.-K. NOD2 and TLR2 ligands trigger the activation of basophils and eosinophils by interacting with dermal fibroblasts in atopic dermatitis-like skin inflammation. Cell. Mol. Immunol. 2016, 13, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kim, B.E.; Leung, D.Y.M. Pathophysiology of atopic dermatitis: Clinical implications. Allergy Asthma Proc. 2019, 40, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Liu, W.; Zhang, L.-J. The Role of Toll-Like Receptors in Skin Host Defense, Psoriasis, and Atopic Dermatitis. J. Immunol. Res. 2019, 2019, 1824624. [Google Scholar] [CrossRef]
- Kuo, I.-H.; Carpenter-Mendini, A.; Yoshida, T.; McGirt, L.Y.; Ivanov, A.I.; Barnes, K.C.; Gallo, R.L.; Borkowski, A.W.; Yamasaki, K.; Leung, D.Y.; et al. Activation of epidermal toll-like receptor 2 enhances tight junction function: Implications for atopic dermatitis and skin barrier repair. J. Investig. Dermatol. 2013, 133, 988–998. [Google Scholar] [CrossRef] [Green Version]
- Pastore, S.; Mascia, F.; Mariani, V.; Girolomoni, G. The epidermal growth factor receptor system in skin repair and inflammation. J. Investig. Dermatol. 2008, 128, 1365–1374. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-J.; Lee, S.-H. Epidermal permeability barrier defects and barrier repair therapy in atopic dermatitis. Allergy. Asthma Immunol. Res. 2014, 6, 276–287. [Google Scholar] [CrossRef] [Green Version]
- De Benedetto, A.; Agnihothri, R.; McGirt, L.Y.; Bankova, L.G.; Beck, L.A. Atopic Dermatitis: A Disease Caused by Innate Immune Defects? J. Investig. Dermatol. 2009, 129, 14–30. [Google Scholar] [CrossRef] [Green Version]
- Chieosilapatham, P.; Kiatsurayanon, C.; Umehara, Y.; Trujillo-Paez, J.V.; Peng, G.; Yue, H.; Nguyen, L.T.H.; Niyonsaba, F. Keratinocytes: Innate immune cells in atopic dermatitis. Clin. Exp. Immunol. 2021, 204, 296–309. [Google Scholar] [CrossRef]
- Park, G.; Moon, B.C.; Choi, G.; Lim, H.-S. Cera Flava Alleviates Atopic Dermatitis by Activating Skin Barrier Function via Immune Regulation. Int. J. Mol. Sci. 2021, 22, 7531. [Google Scholar] [CrossRef]
- Gong, L.; Wang, H.; Sun, X.; Liu, C.; Duan, C.; Cai, R.; Gu, X.; Zhu, S. Toll-Interleukin 1 Receptor domain-containing adaptor protein positively regulates BV2 cell M1 polarization. Eur. J. Neurosci. 2016, 43, 1674–1682. [Google Scholar] [CrossRef]
- Belhaouane, I.; Hoffmann, E.; Chamaillard, M.; Brodin, P.; Machelart, A. Paradoxical Roles of the MAL/Tirap Adaptor in Pathologies. Front. Immunol. 2020, 11, 569127. [Google Scholar] [CrossRef]
- Achek, A.; Kwon, H.-K.; Patra, M.C.; Shah, M.; Hong, R.; Lee, W.H.; Baek, W.-Y.; Choi, Y.S.; Kim, G.-Y.; Pham, T.L.H.; et al. A peptide derived from the core β-sheet region of TIRAP decoys TLR4 and reduces inflammatory and autoimmune symptoms in murine models. EBioMedicine 2020, 52, 102645. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, R.; Dainichi, T.; Tsuchiya, S.; Nomura, T.; Kitoh, A.; Hayden, M.S.; Ishii, K.J.; Tanaka, M.; Honda, T.; Egawa, G.; et al. Epithelial TRAF6 drives IL-17-mediated psoriatic inflammation. JCI Insight 2018, 3. [Google Scholar] [CrossRef]
- Gohda, J.; Matsumura, T.; Inoue, J. Cutting edge: TNFR-associated factor (TRAF) 6 is essential for MyD88-dependent pathway but not toll/IL-1 receptor domain-containing adaptor-inducing IFN-beta (TRIF)-dependent pathway in TLR signaling. J. Immunol. 2004, 173, 2913–2917. [Google Scholar] [CrossRef] [Green Version]
- Monaco, C.; Nanchahal, J.; Taylor, P.; Feldmann, M. Anti-TNF therapy: Past, present and future. Int. Immunol. 2015, 27, 55–62. [Google Scholar] [CrossRef]
- Nishikomori, R.; Izawa, K.; Kambe, N.; Ohara, O.; Yasumi, T. Low-frequency mosaicism in cryopyrin-associated periodic fever syndrome: Mosaicism in systemic autoinflammatory diseases. Int. Immunol. 2019, 31, 649–655. [Google Scholar] [CrossRef]
- Lai, Y.; Dong, C. Therapeutic antibodies that target inflammatory cytokines in autoimmune diseases. Int. Immunol. 2016, 28, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Kaminska, B. MAPK signalling pathways as molecular targets for anti-inflammatory therapy--from molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta 2005, 1754, 253–262. [Google Scholar] [CrossRef]
- Hommes, D.W.; Peppelenbosch, M.P.; van Deventer, S.J.H. Mitogen activated protein (MAP) kinase signal transduction pathways and novel anti-inflammatory targets. Gut 2003, 52, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Dumitru, C.D.; Ceci, J.D.; Tsatsanis, C.; Kontoyiannis, D.; Stamatakis, K.; Lin, J.H.; Patriotis, C.; Jenkins, N.A.; Copeland, N.G.; Kollias, G.; et al. TNF-alpha induction by LPS is regulated posttranscriptionally via a Tpl2/ERK-dependent pathway. Cell 2000, 103, 1071–1083. [Google Scholar] [CrossRef] [Green Version]
- Peroval, M.Y.; Boyd, A.C.; Young, J.R.; Smith, A.L. A critical role for MAPK signalling pathways in the transcriptional regulation of toll like receptors. PLoS ONE 2013, 8, e51243. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.-Y.; Schumann, R.R.; Hamann, L.; Neumann, K.; Worm, M.; Heine, G. Association of the toll-like receptor 2 A-16934T promoter polymorphism with severe atopic dermatitis. Allergy 2009, 64, 1608–1615. [Google Scholar] [CrossRef]
- Chan, C.W.H.; Leung, T.F.; Chow, K.M.; Wong, C.L.; Choi, K.C.; Chan, J.Y.W. Association of toll-like receptor 5 polymorphisms (rs5744174 and rs851139) and eczema: A pilot birth cohort study in Hong Kong newborn babies. Int. J. Dermatol. 2018, 57, e134–e135. [Google Scholar] [CrossRef]
- Novak, N.; Yu, C.-F.; Bussmann, C.; Maintz, L.; Peng, W.-M.; Hart, J.; Hagemann, T.; Diaz-Lacava, A.; Baurecht, H.-J.; Klopp, N.; et al. Putative association of a TLR9 promoter polymorphism with atopic eczema. Allergy 2007, 62, 766–772. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Bryant, C.E.; Doyle, S.L. Therapeutic targeting of Toll-like receptors for infectious and inflammatory diseases and cancer. Pharmacol. Rev. 2009, 61, 177–197. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Bao, K.; Wu, P.; Yu, X.; Wang, C.; Ji, L.; Hong, M. Integrative Analysis of lncRNAs, miRNAs, and mRNA-Associated ceRNA Network in an Atopic Dermatitis Recurrence Model. Int. J. Mol. Sci. 2018, 19, 3263. [Google Scholar] [CrossRef] [Green Version]
- Legat, F.J. Itch in Atopic Dermatitis—What Is New? Front. Med. 2021, 8. [Google Scholar] [CrossRef]
- Debnath, T.; Lee, Y.M.; Lim, J.H.; Lim, B.O. Anti-allergic and anti-atopic dermatitis effects of Gardenia Fructus extract. Food Agric. Immunol. 2018, 29, 665–674. [Google Scholar] [CrossRef]
- Huang, G.-J.; Bhaskar Reddy, M.V.; Kuo, P.-C.; Huang, C.-H.; Shih, H.-C.; Lee, E.-J.; Yang, M.-L.; Leu, Y.-L.; Wu, T.-S. A concise synthesis of viscolin, and its anti-inflammatory effects through the suppression of iNOS, COX-2, ERK phosphorylation and proinflammatory cytokines expressions. Eur. J. Med. Chem. 2012, 48, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Yatsuzuka, R.; Jiang, S.; Ueda, Y.; Kamei, C. Involvement of cyclooxygenase-2 in allergic nasal inflammation in rats. Int. Immunopharmacol. 2006, 6, 1736–1742. [Google Scholar] [CrossRef] [PubMed]
- Wee, J.H.; Zhang, Y.-L.; Rhee, C.-S.; Kim, D.-Y. Inhibition of Allergic Response by Intranasal Selective NF-κB Decoy Oligodeoxynucleotides in a Murine Model of Allergic Rhinitis. Allergy. Asthma Immunol. Res. 2017, 9, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Iannone, M.; Tonini, G.; Janowska, A.; Dini, V.; Romanelli, M. Definition of treatment goals in terms of clinician-reported disease severity and patient-reported outcomes in moderate-to-severe adult atopic dermatitis: A systematic review. Curr. Med. Res. Opin. 2021, 37, 1295–1301. [Google Scholar] [CrossRef]
- Siegels, D.; Heratizadeh, A.; Abraham, S.; Binnmyr, J.; Brockow, K.; Irvine, A.D.; Halken, S.; Mortz, C.G.; Flohr, C.; Schmid-Grendelmeier, P.; et al. Systemic treatments in the management of atopic dermatitis: A systematic review and meta-analysis. Allergy 2021, 76, 1053–1076. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kordulewska, N.K.; Topa, J.; Stryiński, R.; Jarmołowska, B. Osthole Inhibits Expression of Genes Associated with Toll-like Receptor 2 Signaling Pathway in an Organotypic 3D Skin Model of Human Epidermis with Atopic Dermatitis. Cells 2022, 11, 88. https://doi.org/10.3390/cells11010088
Kordulewska NK, Topa J, Stryiński R, Jarmołowska B. Osthole Inhibits Expression of Genes Associated with Toll-like Receptor 2 Signaling Pathway in an Organotypic 3D Skin Model of Human Epidermis with Atopic Dermatitis. Cells. 2022; 11(1):88. https://doi.org/10.3390/cells11010088
Chicago/Turabian StyleKordulewska, Natalia Karolina, Justyna Topa, Robert Stryiński, and Beata Jarmołowska. 2022. "Osthole Inhibits Expression of Genes Associated with Toll-like Receptor 2 Signaling Pathway in an Organotypic 3D Skin Model of Human Epidermis with Atopic Dermatitis" Cells 11, no. 1: 88. https://doi.org/10.3390/cells11010088
APA StyleKordulewska, N. K., Topa, J., Stryiński, R., & Jarmołowska, B. (2022). Osthole Inhibits Expression of Genes Associated with Toll-like Receptor 2 Signaling Pathway in an Organotypic 3D Skin Model of Human Epidermis with Atopic Dermatitis. Cells, 11(1), 88. https://doi.org/10.3390/cells11010088