
The Impact of Escherichia coli Probiotic Strain O83:K24:H31 on the Maturation of Dendritic Cells and Immunoregulatory Functions In Vitro and In Vivo

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Preparation
2.2. Mice
2.3. Preparation and Stimulation of Bone Marrow–Derived Dendritic Cells
2.4. Coculture of EcO83-Primed BMDC with Naïve CD4+ T Cells
2.5. Flow Cytometry
2.6. Gene Expression Analyses
2.7. ELISA
2.8. In Vivo Bioluminiscence Imaging
2.9. Statistical Analyses
3. Results
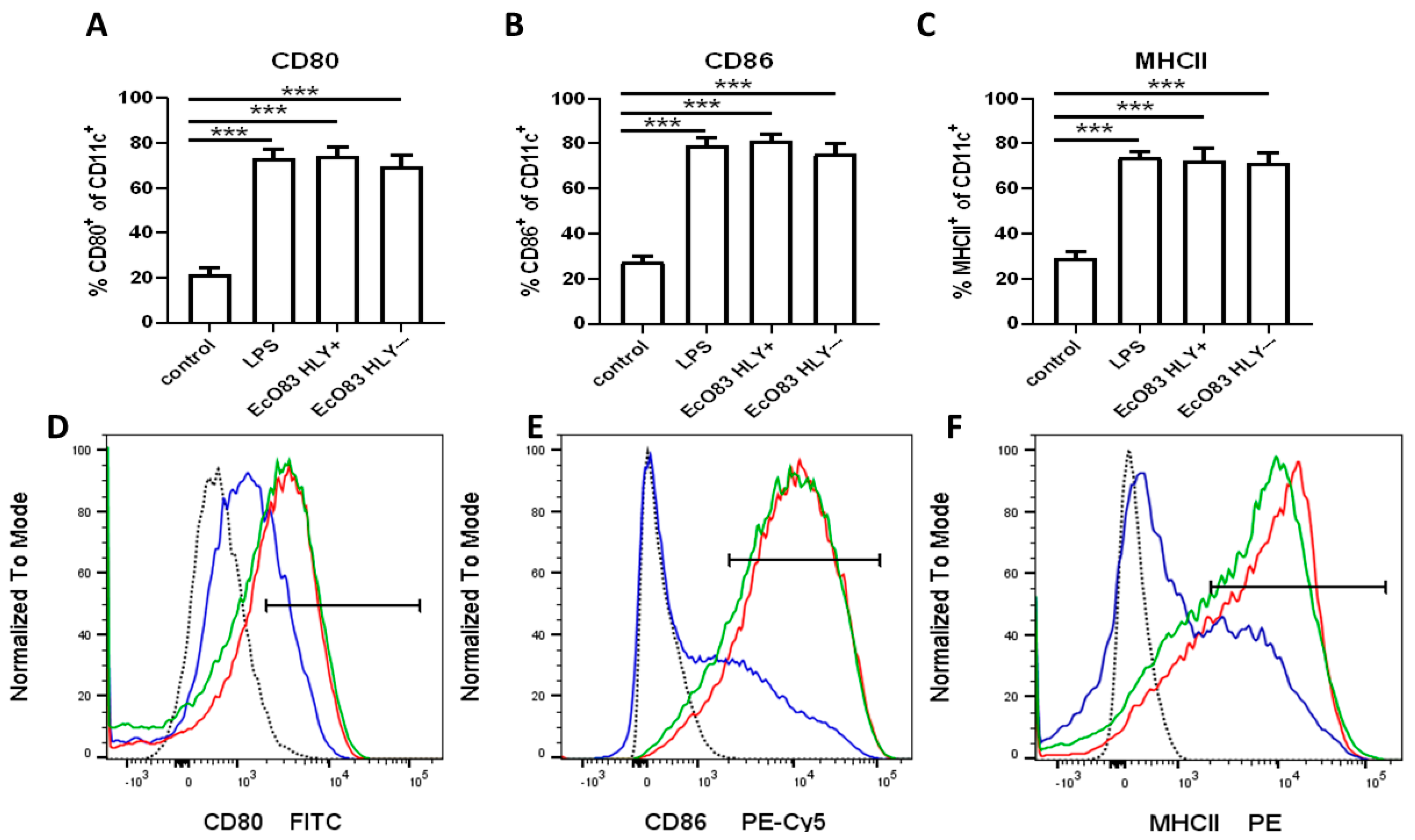
3.1. The Impact of EcO83 on BMDC Maturation
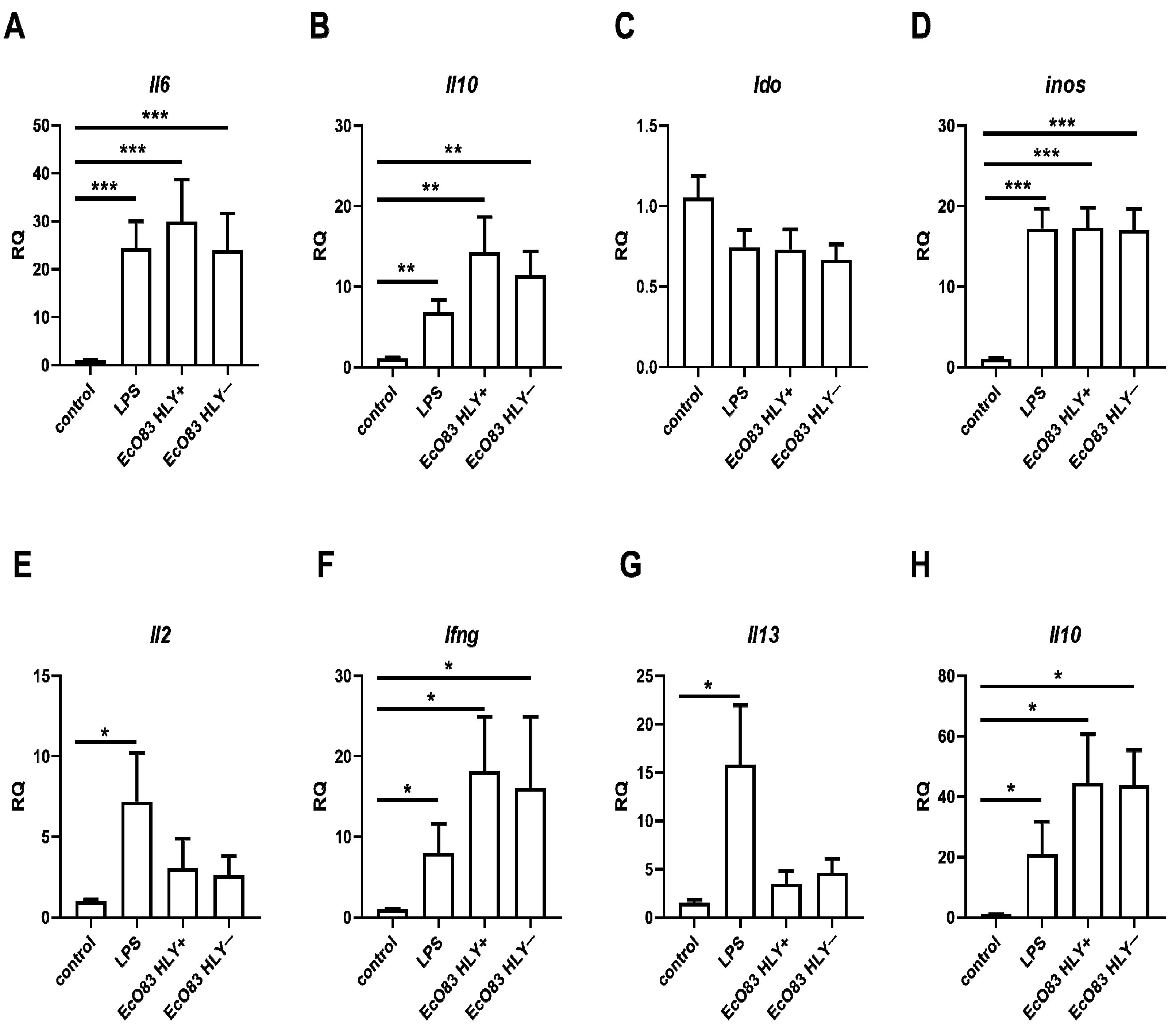
3.2. The Impact of EcO83 on Gene Expression in BMDC and CD4+ T Cells Cocultured with Primed BMDC
3.2.1. The Effect of EcO83 on Gene Expression in BMDC In Vitro
3.2.2. The Capacity of EcO83-Primed BMDC and Gut DC to Polarize CD4+ T Cells
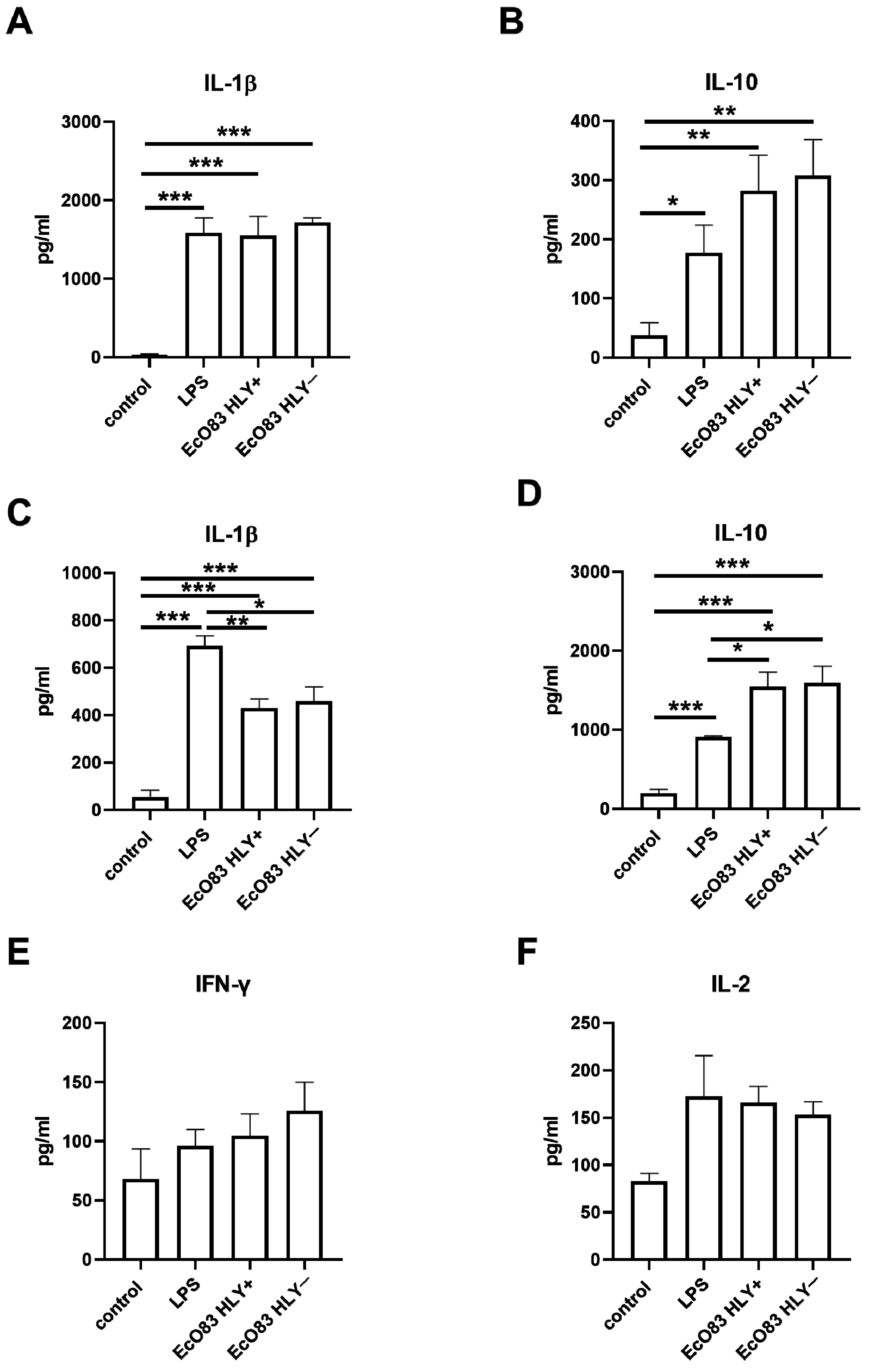
3.3. The Effect of EcO83 on Cytokine Production by BMDC and CD4+ T Cells Cocultured with Primed BMDC
3.3.1. The Capacity of EcO83 to Modulate Cytokine Production by BMDC
3.3.2. The Impact of Primed BMDC on Cytokine Release by CD4+ T Cells
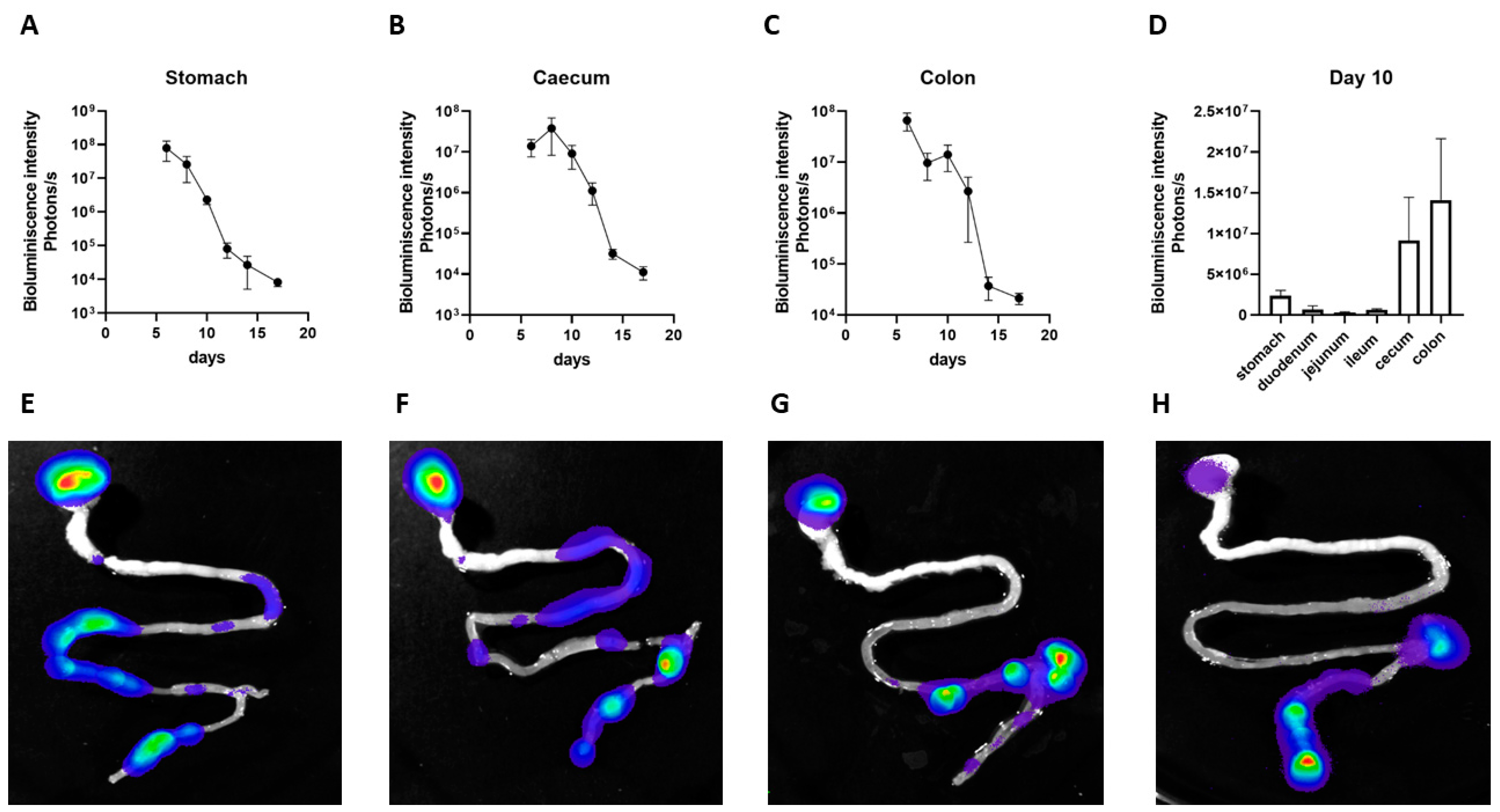
3.4. Persistence of EcO83 in Neonatal Gastrointestinal Tract
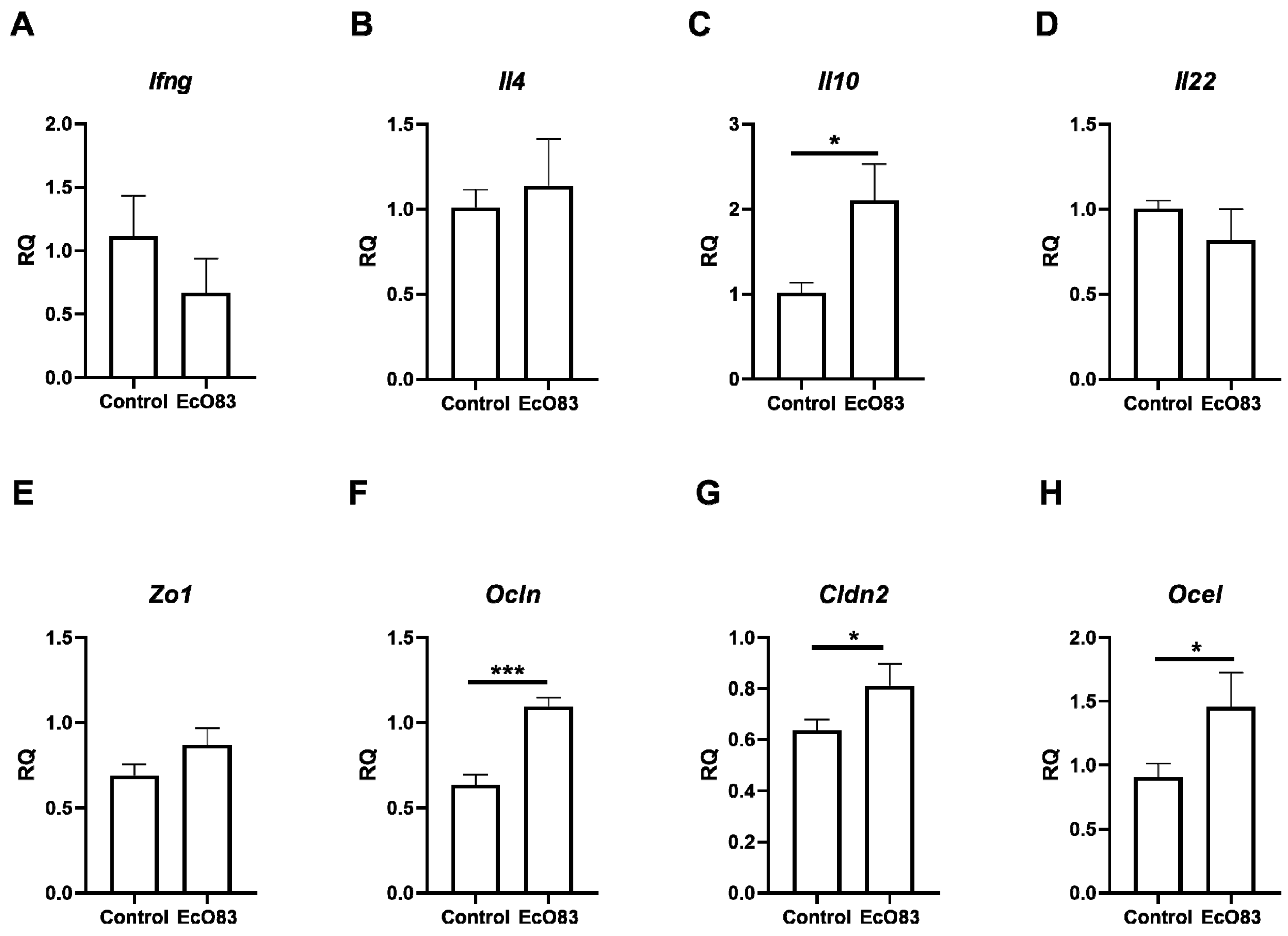
3.5. Effect of Early Postantal EcO83 on Cytokine Expression in Gut
3.6. The Effect of Early Postnatal Administration of EcO83 on Gut Barrier Functions
3.7. The Impact of Early Postnatal EcO83 Administration on Selected Cellular Subsets in Gut
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mishra, A.; Lai, G.C.; Yao, L.J.; Aung, T.T.; Shental, N.; Rotter-Maskowitz, A.; Shepherdson, E.; Singh, G.S.N.; Pai, R.; Shanti, A.; et al. Microbial exposure during early human development primes fetal immune cells. Cell 2021, 184, 3394–3409. [Google Scholar] [CrossRef] [PubMed]
- Pelzer, E.; Gomez-Arango, L.F.; Barrett, H.L.; Nitert, M.D. Review: Maternal health and the placental microbiome. Placenta 2017, 54, 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prince, A.L.; Ma, J.; Kannan, P.S.; Alvarez, M.; Gisslen, T.; Harris, R.A.; Sweeney, E.L.; Knox, C.L.; Lambers, D.S.; Jobe, A.H.; et al. The placental membrane microbiome is altered among subjects with spontaneous preterm birth with and without chorioamnionitis. Am. J. Obs. Gynecol. 2016, 214, 627.e1–627.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Gadir, A.; Massoud, A.H.; Chatila, T.A. Antigen-specific treg cells in immunological tolerance: Implications for allergic diseases. F1000Research 2018, 7, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiby, J.S.; McCormick, K.; Sherrill-Mix, S.; Clarke, E.L.; Kessler, L.R.; Taylor, L.J.; Hofstaedter, C.E.; Roche, A.M.; Mattei, L.M.; Bittinger, K.; et al. Lack of detection of a human placenta microbiome in samples from preterm and term deliveries. Microbiome 2018, 6, 196. [Google Scholar] [CrossRef] [PubMed]
- de Goffau, M.C.; Lager, S.; Sovio, U.; Gaccioli, F.; Cook, E.; Peacock, S.J.; Parkhill, J.; Charnock-Jones, D.S.; Smith, G.C.S. Human placenta has no microbiome but can contain potential pathogens. Nature 2019, 572, 329–334. [Google Scholar] [CrossRef]
- Zhang, C.; Li, L.; Jin, B.; Xu, X.; Zuo, X.; Li, Y.; Li, Z. The effects of delivery mode on the gut microbiota and health: State of art. Front. Microbiol. 2021, 12, 724449. [Google Scholar] [CrossRef]
- Kolokotroni, O.; Middleton, N.; Gavatha, M.; Lamnisos, D.; Priftis, K.N.; Yiallouros, P.K. Asthma and atopy in children borns by caesarean section: Effect modification by family history of allergies—A population based cross-sectional study. BMC Pediatr. 2012, 12, 179. [Google Scholar] [CrossRef] [Green Version]
- Kindinger, L.M.; Bennett, P.R.; Lee, Y.S.; Marchesi, J.R.; Smith, A.; Cacciatore, S.; Holmes, E.; Nicholson, J.K.; Teoh, T.G.; MacIntyre, D.A. The interaction between vaginal microbiota, cervical length, and vaginal progesterone treatment for preterm birth risk. Microbiome 2017, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.; Makino, H.; Cetinyurek Yavuz, A.; Ben-Amor, K.; Roelofs, M.; Ishikawa, E.; Kubota, H.; Swinkels, S.; Sakai, T.; Oishi, K.; et al. Early-life events, including mode of delivery and type of feeding, siblings and gender, shape the developing gut microbiota. PLoS ONE 2016, 11, e0158498. [Google Scholar] [CrossRef] [Green Version]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dogra, S.; Sakwinska, O.; Soh, S.E.; Ngom-Bru, C.; Bruck, W.M.; Berger, B.; Brussow, H.; Lee, Y.S.; Yap, F.; Chong, Y.S.; et al. Dynamics of infant gut microbiota are influenced by delivery mode and gestational duration and are associated with subsequent adiposity. mBio 2015, 6, e02419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakwinska, O.; Foata, F.; Berger, B.; Brüssow, H.; Combremont, S.; Mercenier, A.; Dogra, S.; Soh, S.-E.; Yen, J.C.K.; Heong, G.Y.S.; et al. Does the maternal vaginal microbiota play a role in seeding the microbiota of neonatal gut and nose? Benef. Microbes 2017, 8, 763–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koren, O.; Goodrich, J.K.; Cullender, T.C.; Spor, A.; Laitinen, K.; Backhed, H.K.; Gonzalez, A.; Werner, J.J.; Angenent, L.T.; Knight, R.; et al. Host remodeling of the gut microbiome and metabolic changes during pregnancy. Cell 2012, 150, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Freitas, A.C.; Bocking, A.; Hill, J.E.; Money, D.M.; the VOGUE Research Group. Increased richness and diversity of the vaginal microbiota and spontaneous preterm birth. Microbiome 2018, 6, 117. [Google Scholar] [CrossRef]
- Romero, R.; Hassan, S.S.; Gajer, P.; Tarca, A.L.; Fadrosh, D.W.; Bieda, J.; Chaemsaithong, P.; Miranda, J.; Chaiworapongsa, T.; Ravel, J. The vaginal microbiota of pregnant women who subsequently have spontaneous preterm labor and delivery and those with a normal delivery at term. Microbiome 2014, 2, 18. [Google Scholar] [CrossRef] [Green Version]
- Zommiti, M.; Chikindas, M.L.; Ferchichi, M. Probiotics-live biotherapeutics: A story of success, limitations, and future prospects-not only for humans. Probiotics Antimicrob. Proteins 2020, 12, 1266–1289. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Amenyogbe, N.; Kollmann, T.R.; Ben-Othman, R. Early-life host-microbiome interphase: The key frontier for immune development. Front. Pediatr. 2017, 5, 111. [Google Scholar] [CrossRef] [Green Version]
- Nash, M.J.; Frank, D.N.; Friedman, J.E. Early microbes modify immune system development and metabolic homeostasis-the “restaurant” hypothesis revisited. Front. Endocrinol. 2017, 8, 349. [Google Scholar] [CrossRef]
- Walker, W.A. The importance of appropriate initial bacterial colonization of the intestine in newborn, child, and adult health. Pediatr. Res. 2017, 82, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Dzidic, M.; Boix-Amoros, A.; Selma-Royo, M.; Mira, A.; Collado, M.C. Gut microbiota and mucosal immunity in the neonate. Med. Sci. 2018, 6, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozakova, H.; Schwarzer, M.; Tuckova, L.; Srutkova, D.; Czarnowska, E.; Rosiak, I.; Hudcovic, T.; Schabussova, I.; Hermanova, P.; Zakostelska, Z.; et al. Colonization of germ-free mice with a mixture of three lactobacillus strains enhances the integrity of gut mucosa and ameliorates allergic sensitization. Cell. Mol. Immunol. 2016, 13, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Ohnmacht, C. Microbiota, regulatory t cell subsets, and allergic disorders. Allergo J. Int. 2016, 25, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Koga, M.M.; Engel, A.; Pigni, M.; Lavanchy, C.; Stevanin, M.; Laversenne, V.; Schneider, B.L.; Acha-Orbea, H. Il10- and il35-secreting mutudc lines act in cooperation to inhibit memory t cell activation through lag-3 expression. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Domogalla, M.P.; Rostan, P.V.; Raker, V.K.; Steinbrink, K. Tolerance through education: How tolerogenic dendritic cells shape immunity. Front. Immunol. 2017, 8, 1764. [Google Scholar] [CrossRef] [Green Version]
- Olson, B.; Sullivan, J.; Burlingham, W. Interleukin 35: A key mediator of suppression and the propagation of infectious tolerance. Front. Immunol. 2013, 4, 315. [Google Scholar] [CrossRef] [Green Version]
- Shevach, E.M.; Thornton, A.M. Ttregs, ptregs, and itregs: Similarities and differences. Immunol. Rev. 2014, 259, 88–102. [Google Scholar] [CrossRef] [Green Version]
- Shevach, E.M. Mechanisms of foxp3+ t regulatory cell-mediated suppression. Immunity 2009, 30, 636–645. [Google Scholar] [CrossRef] [Green Version]
- Konieczna, P.; Groeger, D.; Ziegler, M.; Frei, R.; Ferstl, R.; Shanahan, F.; Quigley, E.M.; Kiely, B.; Akdis, C.A.; O’Mahony, L. Bifidobacterium infantis 35624 administration induces foxp3 t regulatory cells in human peripheral blood: Potential role for myeloid and plasmacytoid dendritic cells. Gut 2012, 61, 354–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, S.G.; Kayama, H.; Ueda, Y.; Takahashi, T.; Asahara, T.; Tsuji, H.; Tsuji, N.M.; Kiyono, H.; Ma, J.S.; Kusu, T.; et al. Probiotic bifidobacterium breve induces il-10-producing tr1 cells in the colon. PLoS Pathog. 2012, 8, e1002714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macho Fernandez, E.; Valenti, V.; Rockel, C.; Hermann, C.; Pot, B.; Boneca, I.G.; Grangette, C. Anti-inflammatory capacity of selected lactobacilli in experimental colitis is driven by nod2-mediated recognition of a specific peptidoglycan-derived muropeptide. Gut 2011, 60, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Zakostelska, Z.; Kverka, M.; Klimesova, K.; Rossmann, P.; Mrazek, J.; Kopecny, J.; Hornova, M.; Srutkova, D.; Hudcovic, T.; Ridl, J.; et al. Lysate of probiotic lactobacillus casei dn-114 001 ameliorates colitis by strengthening the gut barrier function and changing the gut microenvironment. PLoS ONE 2011, 6, e27961. [Google Scholar] [CrossRef] [PubMed]
- Hrdy, J.; Kocourkova, I.; Lodinova-Zadnikova, R.; Kolarova, L.; Prokesova, L. The effect of a probiotic Escherichia coli strain on regulatory t-cells in six year-old children. Benef. Microbes 2016, 7, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Hrdy, J.; Vlasakova, K.; Cerny, V.; Sukenikova, L.; Novotna, O.; Petraskova, P.; Borakova, K.; Lodinova-Zadnikova, R.; Kolarova, L.; Prokesova, L. Decreased allergy incidence in children supplemented with E. coli o83:K24:H31 and its possible modes of action. Eur. J. Immunol. 2018, 48, 2015–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodinova-Zadnikova, R.; Cukrowska, B.; Tlaskalova-Hogenova, H. Oral administration of probiotic Escherichia coli after birth reduces frequency of allergies and repeated infections later in life (after 10 and 20 years). Int. Arch. Allergy Immunol. 2003, 131, 209–211. [Google Scholar] [CrossRef]
- Lodinova-Zadnikova, R.; Prokesova, L.; Kocourkova, I.; Hrdy, J.; Zizka, J. Prevention of allergy in infants of allergic mothers by probiotic Escherichia coli. Int. Arch. Allergy Immunol. 2010, 153, 201–206. [Google Scholar] [CrossRef]
- Wassenaar, T.M. Insights from 100 years of research with probiotic E. coli. Eur. J. Microbiol. Immunol. 2016, 6, 147–161. [Google Scholar] [CrossRef] [Green Version]
- Sheshko, V.; Hejnova, J.; Rehakova, Z.; Sinkora, J.; Faldyna, M.; Alexa, P.; Felsberg, J.; Nemcova, R.; Bomba, A.; Sebo, P. Hlya knock out yields a safer Escherichia coli a0 34/86 variant with unaffected colonization capacity in piglets. FEMS Immunol. Med. Microbiol. 2006, 48, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Hrdy, J.; Alard, J.; Couturier-Maillard, A.; Boulard, O.; Boutillier, D.; Delacre, M.; Lapadatescu, C.; Cesaro, A.; Blanc, P.; Pot, B.; et al. Lactobacillus reuteri 5454 and Bifidobacterium animalis ssp. Lactis 5764 improve colitis while differentially impacting dendritic cells maturation and antimicrobial responses. Sci. Rep. 2020, 10, 5345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majer, M.; Machacek, T.; Sukenikova, L.; Hrdy, J.; Horak, P. The peripheral immune response of mice infected with a neuropathogenic schistosome. Parasite Immunol. 2020, 42, e12710. [Google Scholar] [CrossRef] [PubMed]
- Cuffaro, B.; Assohoun, A.L.W.; Boutillier, D.; Sukenikova, L.; Desramaut, J.; Boudebbouze, S.; Salome-Desnoulez, S.; Hrdy, J.; Waligora-Dupriet, A.J.; Maguin, E.; et al. In vitro characterization of gut microbiota-derived commensal strains: Selection of parabacteroides distasonis strains alleviating tnbs-induced colitis in mice. Cells 2020, 9, 2104. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J.; Gomez-Llorente, C.; Campana-Martin, L.; Matencio, E.; Ortuno, I.; Martinez-Silla, R.; Gomez-Gallego, C.; Periago, M.J.; Ros, G.; Chenoll, E.; et al. Safety and immunomodulatory effects of three probiotic strains isolated from the feces of breast-fed infants in healthy adults: Setoprob study. PLoS ONE 2013, 8, e78111. [Google Scholar] [CrossRef]
- Hrdý, J.; Zanvit, P.; Novotná, O.; Kocourková, I.; Zižka, J.; Prokešová, L. Cytokine expression in cord blood cells of children of healthy and allergic mothers. Folia Microbiol. 2010, 55, 515–519. [Google Scholar] [CrossRef]
- Sukenikova, L.; Cerny, V.; Novotna, O.; Petraskova, P.; Borakova, K.; Kolarova, L.; Prokesova, L.; Hrdy, J. Different capacity of in vitro generated myeloid dendritic cells of newborns of healthy and allergic mothers to respond to probiotic strain E. coli o83:K24:H31. Immunol. Lett. 2017, 189, 82–89. [Google Scholar] [CrossRef]
- Meyer, M.P.; Chow, S.S.W.; Alsweiler, J.; Bourchier, D.; Broadbent, R.; Knight, D.; Lynn, A.M.; Patel, H. Probiotics for prevention of severe necrotizing enterocolitis: Experience of new zealand neonatal intensive care units. Front. Pediatr. 2020, 8, 119. [Google Scholar] [CrossRef]
- Wu, B.B.; Yang, Y.; Xu, X.; Wang, W.P. Effects of bifidobacterium supplementation on intestinal microbiota composition and the immune response in healthy infants. World J. Pediatr. WJP 2016, 12, 177–182. [Google Scholar] [CrossRef]
- Kim, H.K.; Rutten, N.B.; Besseling-van der Vaart, I.; Niers, L.E.; Choi, Y.H.; Rijkers, G.T.; van Hemert, S. Probiotic supplementation influences faecal short chain fatty acids in infants at high risk for eczema. Benef. Microbes 2015, 6, 783–790. [Google Scholar] [CrossRef]
- Simpson, M.R.; Dotterud, C.K.; Storro, O.; Johnsen, R.; Oien, T. Perinatal probiotic supplementation in the prevention of allergy related disease: 6 year follow up of a randomised controlled trial. BMC Derm. 2015, 15, 13. [Google Scholar] [CrossRef] [Green Version]
- Harden, J.L.; Egilmez, N.K. Indoleamine 2,3-dioxygenase and dendritic cell tolerogenicity. Immunol. Investig. 2012, 41, 738–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esmaeili, S.A.; Mahmoudi, M.; Rezaieyazdi, Z.; Sahebari, M.; Tabasi, N.; Sahebkar, A.; Rastin, M. Generation of tolerogenic dendritic cells using Lactobacillus rhamnosus and Lactobacillus delbrueckii as tolerogenic probiotics. J. Cell. Biochem. 2018, 119, 7865–7872. [Google Scholar] [CrossRef] [PubMed]
- Di Giacinto, C.; Marinaro, M.; Sanchez, M.; Strober, W.; Boirivant, M. Probiotics ameliorate recurrent th1-mediated murine colitis by inducing il-10 and il-10-dependent tgf-beta-bearing regulatory cells. J. Immunol. 2005, 174, 3237–3246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powrie, F.; Leach, M.W.; Mauze, S.; Menon, S.; Caddle, L.B.; Coffman, R.L. Inhibition of th1 responses prevents inflammatory bowel disease in scid mice reconstituted with CD45RBhi CD4+ T cells. Immunity 1994, 1, 553–562. [Google Scholar] [CrossRef]
- Asseman, C.; Mauze, S.; Leach, M.W.; Coffman, R.L.; Powrie, F. An essential role for interleukin 10 in the function of regulatory t cells that inhibit intestinal inflammation. J. Exp. Med. 1999, 190, 995–1004. [Google Scholar] [CrossRef]
- So, J.S.; Lee, C.G.; Kwon, H.K.; Yi, H.J.; Chae, C.S.; Park, J.A.; Hwang, K.C.; Im, S.H. Lactobacillus casei potentiates induction of oral tolerance in experimental arthritis. Mol. Immunol. 2008, 46, 172–180. [Google Scholar] [CrossRef]
- Aoki-Yoshida, A.; Yamada, K.; Hachimura, S.; Sashihara, T.; Ikegami, S.; Shimizu, M.; Totsuka, M. Enhancement of oral tolerance induction in do11.10 mice by Lactobacillus gasseri oll2809 via increase of effector regulatory T cells. PLoS ONE 2016, 11, e0158643. [Google Scholar] [CrossRef] [Green Version]
- Lavasani, S.; Dzhambazov, B.; Nouri, M.; Fåk, F.; Buske, S.; Molin, G.; Thorlacius, H.; Alenfall, J.; Jeppsson, B.; Weström, B. A novel probiotic mixture exerts a therapeutic effect on experimental autoimmune encephalomyelitis mediated by il-10 producing regulatory t cells. PLoS ONE 2010, 5, e9009. [Google Scholar] [CrossRef]
- Drakes, M.; Blanchard, T.; Czinn, S. Bacterial probiotic modulation of dendritic cells. Infect. Immun. 2004, 72, 3299–3309. [Google Scholar] [CrossRef] [Green Version]
- Hart, A.L.; Lammers, K.; Brigidi, P.; Vitali, B.; Rizzello, F.; Gionchetti, P.; Campieri, M.; Kamm, M.A.; Knight, S.C.; Stagg, A.J. Modulation of human dendritic cell phenotype and function by probiotic bacteria. Gut 2004, 53, 1602–1609. [Google Scholar] [CrossRef]
- Choi, S.-H.; Lee, S.-H.; Kim, M.G.; Lee, H.J.; Kim, G.-B. Lactobacillus plantarum cau1055 ameliorates inflammation in lipopolysaccharide-induced raw264.7 cells and a dextran sulfate sodium–induced colitis animal model. J. Dairy Sci. 2019, 102, 6718–6725. [Google Scholar] [CrossRef] [PubMed]
- Alard, J.; Peucelle, V.; Boutillier, D.; Breton, J.; Kuylle, S.; Pot, B.; Holowacz, S.; Grangette, C. New probiotic strains for inflammatory bowel disease management identified by combining in vitro and in vivo approaches. Benef. Microbes 2018, 9, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Abrahamsson, T.R.; Sandberg Abelius, M.; Forsberg, A.; Bjorksten, B.; Jenmalm, M.C. A th1/th2-associated chemokine imbalance during infancy in children developing eczema, wheeze and sensitization. Clin. Exp. Allergy 2011, 41, 1729–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, C.E.; Hammarstrom, M.L.; Hernell, O. Probiotics during weaning reduce the incidence of eczema. Pediatr. Allergy Immunol. 2009, 20, 430–437. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, J.; Wu, C. Modulation of gut microbiota and immune system by probiotics, pre-biotics, and post-biotics. Front. Nutr. 2021, 8, 634897. [Google Scholar] [CrossRef]
- Kocourková, I.; Žádníková, R.; Žižka, J.; Rosová, V. Effect of oral application of a probiotic E. coli strain on the intestinal microflora of children of allergic mothers during the first year of life. Folia Microbiol. 2007, 52, 189–193. [Google Scholar] [CrossRef]
- Sarate, P.J.; Heinl, S.; Poiret, S.; Drinić, M.; Zwicker, C.; Schabussova, I.; Daniel, C.; Wiedermann, U.E. Coli nissle 1917 is a safe mucosal delivery vector for a birch-grass pollen chimera to prevent allergic poly-sensitization. Mucosal Immunol. 2019, 12, 132–144. [Google Scholar] [CrossRef]
- Sanders, D.S.A. Mucosal integrity and barrier function in the pathogenesis of early lesions in crohn’s disease. J. Clin. Pathol. 2005, 58, 568–572. [Google Scholar] [CrossRef] [Green Version]
- Tlaskalová-Hogenová, H.; Stěpánková, R.; Kozáková, H.; Hudcovic, T.; Vannucci, L.; Tučková, L.; Rossmann, P.; Hrnčíř, T.; Kverka, M.; Zákostelská, Z.; et al. The role of gut microbiota (commensal bacteria) and the mucosal barrier in the pathogenesis of inflammatory and autoimmune diseases and cancer: Contribution of germ-free and gnotobiotic animal models of human diseases. Cell. Mol. Immunol 2011, 8, 110–120. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, Y.; Fang, K.; Mao, K.; Dou, J.; Fan, H.; Zhou, C.; Wang, H. Lactobacillus acidophilus and clostridium butyricum ameliorate colitis in murine by strengthening the gut barrier function and decreasing inflammatory factors. Benef. Microbes 2018, 9, 775–787. [Google Scholar] [CrossRef]
- Kangwan, N.; Kongkarnka, S.; Boonkerd, N.; Unban, K.; Shetty, K.; Khanongnuch, C. Protective effect of probiotics isolated from traditional fermented tea leaves (miang) from northern thailand and role of synbiotics in ameliorating experimental ulcerative colitis in mice. Nutrients 2022, 14, 227. [Google Scholar] [CrossRef] [PubMed]
- Bein, A.; Eventov-Friedman, S.; Arbell, D.; Schwartz, B. Intestinal tight junctions are severely altered in nec preterm neonates. Pediatr. Neonatol. 2018, 59, 464–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpern, M.D.; Denning, P.W. The role of intestinal epithelial barrier function in the development of NEC. Tissue Barriers 2015, 3, e100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokešová, A.; Frolová, L.; Kverka, M.; Sokol, D.; Rossmann, P.; Bártová, J.; Tlaskalová-Hogenová, H. Oral administration of probiotic bacteria (E. coli Nissle, E. coli O83, Lactobacillus casei) influences the severity of dextran sodium sulfate-induced colitis in BALB/c mice. Folia Microbiol. 2006, 51, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Gronbach, K.; Flade, I.; Holst, O.; Lindner, B.; Ruscheweyh, H.J.; Wittmann, A.; Menz, S.; Schwiertz, A.; Adam, P.; Stecher, B.; et al. Endotoxicity of lipopolysaccharide as a determinant of T-cell-mediated colitis induction in mice. Gastroenterology 2014, 146, 765–775. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name (Abbreviation) | Catatogue Number of TaqMan Gene Expression Assay |
|---|---|
| Beta-actin (Actb) | Mm00607939_s1 |
| Interleukin 2 (Il2) | Mm00434256_m1 |
| Interleukin 4 (Il4) | Mm99999154_m1 |
| Interleukin 6 (Il6) | Mm99999064_m1 |
| Interleukin 10 (Il10) | Mm00439616_m1 |
| Interleukin 13 (Il13) | Mm99999190_m1 |
| Interleukin 22 (Il22) | Mm00444241_m1 |
| Interferon-gamma (Ifng) | Mm00801778_m1 |
| Inducible NO synthase (inos) | Mm00440485_m1 |
| Indol amine 2,3 dioxygenase (Ido) | Mm00492590_m1 |
| occludin/ELL domain containing 1 (Ocel) | Mm01349279_m1 |
| Zonulin 1 (Tjp1, Zo1) | Mm00493699_m1 |
| Occludin (Ocln) | Mm00500912_m1 |
| Claudin 2 (Cldn2) | Mm00516703_s1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Súkeníková, L.; Černý, V.; Věcek, J.; Petrásková, P.; Novotná, O.; Vobruba, Š.; Michalčíková, T.; Procházka, J.; Kolářová, L.; Prokešová, L.; et al. The Impact of Escherichia coli Probiotic Strain O83:K24:H31 on the Maturation of Dendritic Cells and Immunoregulatory Functions In Vitro and In Vivo. Cells 2022, 11, 1624. https://doi.org/10.3390/cells11101624
Súkeníková L, Černý V, Věcek J, Petrásková P, Novotná O, Vobruba Š, Michalčíková T, Procházka J, Kolářová L, Prokešová L, et al. The Impact of Escherichia coli Probiotic Strain O83:K24:H31 on the Maturation of Dendritic Cells and Immunoregulatory Functions In Vitro and In Vivo. Cells. 2022; 11(10):1624. https://doi.org/10.3390/cells11101624
Chicago/Turabian StyleSúkeníková, Lenka, Viktor Černý, Jan Věcek, Petra Petrásková, Olga Novotná, Šimon Vobruba, Tereza Michalčíková, Jan Procházka, Libuše Kolářová, Ludmila Prokešová, and et al. 2022. "The Impact of Escherichia coli Probiotic Strain O83:K24:H31 on the Maturation of Dendritic Cells and Immunoregulatory Functions In Vitro and In Vivo" Cells 11, no. 10: 1624. https://doi.org/10.3390/cells11101624
APA StyleSúkeníková, L., Černý, V., Věcek, J., Petrásková, P., Novotná, O., Vobruba, Š., Michalčíková, T., Procházka, J., Kolářová, L., Prokešová, L., & Hrdý, J. (2022). The Impact of Escherichia coli Probiotic Strain O83:K24:H31 on the Maturation of Dendritic Cells and Immunoregulatory Functions In Vitro and In Vivo. Cells, 11(10), 1624. https://doi.org/10.3390/cells11101624