
Central Nervous System Pericytes Contribute to Health and Disease

 ,
, 

Abstract
:1. Introduction
2. PC Identity, Multipotency and Differentiation Potential
3. PC Markers
4. PC Origin
5. PC Functions
6. CNS Microvascular PCs and Potential Biomarkers of NVU Damage
7. PC Dysfunction in Neurodevelopmental Disorders
8. PC Dysfunction in Alzheimer’s Disease (AD)
9. PC Dysfunction in Multiple Sclerosis (MS)
10. PC Dysfunction in Stroke and NeuroCOVID-19
11. Conclusions and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Aβ | beta-amyloid |
| α-SMA | alpha smooth muscle actin |
| ACE2 | angiotensin-converting enzyme |
| AD | Alzheimer’s disease |
| AGE | advanced glycation end products |
| AKAP12 | A-kinase anchor protein 12 |
| Ang-1 | angiopoietin-1 |
| ApoE | apolipoprotein E |
| AQP4 | aquaporin-4 |
| ATP | adenosine triphosphate |
| BBB | blood–brain barrier |
| BM | basement membrane |
| Ca2+ | calcium |
| CAA | cerebral amyloid angiopathy |
| CD146 | cluster of differentiation 146 |
| CNS | central nervous system |
| COVID-19 | coronavirus disease 19 |
| CSPGs | chondroitin sulfate proteoglycans |
| CypA | cyclophilin A |
| DLK1 | delta like non-canonical notch ligand 1 |
| EAE | experimental autoimmune encephalomyelitis |
| ECM | extracellular matrix |
| ECs | endothelial cells |
| GJ | gap junctions |
| LRP1 | lipoprotein receptor-related protein 1 |
| MFSD2a | major facilitator superfamily domain-containing protein 2a |
| MMP-9 | matrix metalloproteinase |
| MS | multiple sclerosis |
| MTs | microtubes |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NG2 | neural/glial antigen 2 |
| NVU | neurovascular unit |
| OPCs | oligodendrocyte precursor cells |
| P2X7R | ATP-gated P2X receptor cation channel |
| PCs | pericytes |
| PDGF | platelet-derived growth factor |
| PDGFRβ | PDGF receptor β |
| PGE2 | prostaglandin E2 |
| Ppar-γ | peroxisome proliferator-activated receptor γ |
| RAGE | AGE receptor |
| ROS | reactive oxygen species |
| Runx2 | runt-related transcription factor 2; |
| SARS-CoV-2 | severe acute respiratory syndrome coronavirus-2 |
| SMCs | smooth muscle cells |
| Sox10 | sry-related HMg-Box gene 10 |
| Sox-9 | SRY-box transcription factor 9; |
| Tbx18 | T-Box transcription factor 18 |
| TGFβ-1 | transforming growth factor beta-1 |
| TIMP-3 | tissue inhibitor of metalloproteinase-3 |
| TJ | tight junction |
| TNTs | tunnelling nanotubes |
| Wnt-1 | wingless-type MMTV integration site family, member 1 |
| ZFP423 | PDGFRβ+ zinc finger protein 423 |
References
- Hirschi, K.K.; D’Amore, P.A. Pericytes in the microvasculature. Cardiovasc. Res. 1996, 32, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Campagnolo, P.; Cesselli, D.; Al Haj Zen, A.; Beltrami, A.P.; Krankel, N.; Katare, R.; Angelini, G.; Emanueli, C.; Madeddu, P. Human adult vena saphena contains perivascular progenitor cells endowed with clonogenic and proangiogenic potential. Circulation 2010, 121, 1735–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montiel-Eulefi, E.; Nery, A.A.; Rodrigues, L.C.; Sanchez, R.; Romero, F.; Ulrich, H. Neural differentiation of rat aorta pericyte cells. Cytometry A 2012, 81, 65–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacomazzi, G.; Giovannelli, G.; Rotini, A.; Costamagna, D.; Quattrocelli, M.; Sampaolesi, M. Isolation of Mammalian Mesoangioblasts: A Subset of Pericytes with Myogenic Potential. Methods Mol. Biol. 2021, 2235, 155–167. [Google Scholar] [CrossRef]
- Armulik, A.; Genove, G.; Betsholtz, C. Pericytes: Developmental, physiological, and pathological perspectives, problems, and promises. Dev. Cell 2011, 21, 193–215. [Google Scholar] [CrossRef] [Green Version]
- Sims, D.E. The pericyte—A review. Tissue Cell 1986, 18, 153–174. [Google Scholar] [CrossRef]
- Armulik, A.; Abramsson, A.; Betsholtz, C. Endothelial/pericyte interactions. Circ. Res. 2005, 97, 512–523. [Google Scholar] [CrossRef] [Green Version]
- Tilton, R.G.; Kilo, C.; Williamson, J.R. Pericyte-endothelial relationships in cardiac and skeletal muscle capillaries. Microvasc. Res. 1979, 18, 325–335. [Google Scholar] [CrossRef]
- Nwadozi, E.; Rudnicki, M.; Haas, T.L. Metabolic Coordination of Pericyte Phenotypes: Therapeutic Implications. Front. Cell. Dev. Biol. 2020, 8, 77. [Google Scholar] [CrossRef]
- Lindahl, P.; Johansson, B.R.; Leveen, P.; Betsholtz, C. Pericyte loss and microaneurysm formation in PDGF-B-deficient mice. Science 1997, 277, 242–245. [Google Scholar] [CrossRef]
- Benjamin, L.E.; Hemo, I.; Keshet, E. A plasticity window for blood vessel remodelling is defined by pericyte coverage of the preformed endothelial network and is regulated by PDGF-B and VEGF. Development 1998, 125, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Majesky, M.W.; Dong, X.R.; Regan, J.N.; Hoglund, V.J. Vascular smooth muscle progenitor cells: Building and repairing blood vessels. Circ. Res. 2011, 108, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majesky, M.W. Developmental basis of vascular smooth muscle diversity. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1248–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias Moura Prazeres, P.H.; Sena, I.F.G.; Borges, I.D.T.; de Azevedo, P.O.; Andreotti, J.P.; de Paiva, A.E.; de Almeida, V.M.; de Paula Guerra, D.A.; Pinheiro Dos Santos, G.S.; Mintz, A.; et al. Pericytes are heterogeneous in their origin within the same tissue. Dev. Biol. 2017, 427, 6–11. [Google Scholar] [CrossRef]
- Kawaguchi, M.; Bader, D.M.; Wilm, B. Serosal mesothelium retains vasculogenic potential. Dev. Dyn. 2007, 236, 2973–2979. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; D’Souza, S.S.; Moskvin, O.V.; Toh, H.; Wang, B.; Zhang, J.; Swanson, S.; Guo, L.W.; Thomson, J.A.; Slukvin, I.I. Specification and Diversification of Pericytes and Smooth Muscle Cells from Mesenchymoangioblasts. Cell Rep. 2017, 19, 1902–1916. [Google Scholar] [CrossRef] [Green Version]
- Nehls, V.; Drenckhahn, D. Heterogeneity of microvascular pericytes for smooth muscle type alpha-actin. J. Cell Biol. 1991, 113, 147–154. [Google Scholar] [CrossRef]
- Hartmann, D.A.; Underly, R.G.; Grant, R.I.; Watson, A.N.; Lindner, V.; Shih, A.Y. Pericyte structure and distribution in the cerebral cortex revealed by high-resolution imaging of transgenic mice. Neurophotonics 2015, 2, 041402. [Google Scholar] [CrossRef] [Green Version]
- Girolamo, F.; Dallatomasina, A.; Rizzi, M.; Errede, M.; Walchli, T.; Mucignat, M.T.; Frei, K.; Roncali, L.; Perris, R.; Virgintino, D. Diversified expression of NG2/CSPG4 isoforms in glioblastoma and human foetal brain identifies pericyte subsets. PLoS ONE 2013, 8, e84883. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, T.; Nalbandian, A.; Uchida, Y.; Li, W.; Arnold, T.D.; Kubota, Y.; Yamamoto, S.; Ema, M.; Mukouyama, Y.S. Tissue Myeloid Progenitors Differentiate into Pericytes through TGF-beta Signaling in Developing Skin Vasculature. Cell Rep. 2017, 18, 2991–3004. [Google Scholar] [CrossRef]
- Birbrair, A.; Zhang, T.; Wang, Z.M.; Messi, M.L.; Mintz, A.; Delbono, O. Type-1 pericytes participate in fibrous tissue deposition in aged skeletal muscle. Am. J. Physiol. Cell Physiol. 2013, 305, C1098–C1113. [Google Scholar] [CrossRef] [PubMed]
- Birbrair, A.; Zhang, T.; Wang, Z.M.; Messi, M.L.; Enikolopov, G.N.; Mintz, A.; Delbono, O. Role of pericytes in skeletal muscle regeneration and fat accumulation. Stem Cells Dev. 2013, 22, 2298–2314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Y.; Chen, Z.L.; Norris, E.H.; Strickland, S. Astrocytic laminin regulates pericyte differentiation and maintains blood brain barrier integrity. Nat. Commun. 2014, 5, 3413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Geevarghese, A.; Herman, I.M. Pericyte-endothelial crosstalk: Implications and opportunities for advanced cellular therapies. Transl. Res. 2014, 163, 296–306. [Google Scholar] [CrossRef] [Green Version]
- Barron, L.; Gharib, S.A.; Duffield, J.S. Lung Pericytes and Resident Fibroblasts: Busy Multitaskers. Am. J. Pathol. 2016, 186, 2519–2531. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Chen, S.; Xiang, B.; Li, Y.; Li, L.; Wang, Q. Jagged1/Notch3 Signaling Modulates Hemangioma-Derived Pericyte Proliferation and Maturation. Cell. Physiol. Biochem. 2016, 40, 895–907. [Google Scholar] [CrossRef]
- Birbrair, A.; Borges, I.D.T.; Gilson Sena, I.F.; Almeida, G.G.; da Silva Meirelles, L.; Goncalves, R.; Mintz, A.; Delbono, O. How Plastic Are Pericytes? Stem Cells Dev. 2017, 26, 1013–1019. [Google Scholar] [CrossRef]
- Siedlecki, J.; Asani, B.; Wertheimer, C.; Hillenmayer, A.; Ohlmann, A.; Priglinger, C.; Priglinger, S.; Wolf, A.; Eibl-Lindner, K. Combined VEGF/PDGF inhibition using axitinib induces alphaSMA expression and a pro-fibrotic phenotype in human pericytes. Graefes Arch. Clin. Exp. Ophthalmol. 2018, 256, 1141–1149. [Google Scholar] [CrossRef]
- Alarcon-Martinez, L.; Yilmaz-Ozcan, S.; Yemisci, M.; Schallek, J.; Kilic, K.; Villafranca-Baughman, D.; Can, A.; Di Polo, A.; Dalkara, T. Retinal ischemia induces alpha-SMA-mediated capillary pericyte contraction coincident with perivascular glycogen depletion. Acta Neuropathol. Commun. 2019, 7, 134. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, J.; Itoh, H.; Hirashima, M.; Ogawa, M.; Nishikawa, S.; Yurugi, T.; Naito, M.; Nakao, K.; Nishikawa, S. Flk1-positive cells derived from embryonic stem cells serve as vascular progenitors. Nature 2000, 408, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. Manipulating angiogenesis in medicine. J Intern. Med. 2004, 255, 538–561. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.A.; Mendelson, A.; Kunisaki, Y.; Birbrair, A.; Kou, Y.; Arnal-Estape, A.; Pinho, S.; Ciero, P.; Nakahara, F.; Ma’ayan, A.; et al. Fetal liver hematopoietic stem cell niches associate with portal vessels. Science 2016, 351, 176–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asada, N.; Kunisaki, Y.; Pierce, H.; Wang, Z.; Fernandez, N.F.; Birbrair, A.; Ma’ayan, A.; Frenette, P.S. Differential cytokine contributions of perivascular haematopoietic stem cell niches. Nat. Cell Biol. 2017, 19, 214–223. [Google Scholar] [CrossRef] [Green Version]
- Balabanov, R.; Beaumont, T.; Dore-Duffy, P. Role of central nervous system microvascular pericytes in activation of antigen-primed splenic T-lymphocytes. J. Neurosci. Res. 1999, 55, 578–587. [Google Scholar] [CrossRef]
- Balabanov, R.; Washington, R.; Wagnerova, J.; Dore-Duffy, P. CNS microvascular pericytes express macrophage-like function, cell surface integrin alpha M, and macrophage marker ED-2. Microvasc. Res. 1996, 52, 127–142. [Google Scholar] [CrossRef]
- Crisan, M.; Deasy, B.; Gavina, M.; Zheng, B.; Huard, J.; Lazzari, L.; Peault, B. Purification and long-term culture of multipotent progenitor cells affiliated with the walls of human blood vessels: Myoendothelial cells and pericytes. Methods Cell Biol. 2008, 86, 295–309. [Google Scholar] [CrossRef]
- Birbrair, A.; Zhang, T.; Wang, Z.M.; Messi, M.L.; Enikolopov, G.N.; Mintz, A.; Delbono, O. Skeletal muscle neural progenitor cells exhibit properties of NG2-glia. Exp. Cell Res. 2013, 319, 45–63. [Google Scholar] [CrossRef] [Green Version]
- Dore-Duffy, P.; Katychev, A.; Wang, X.; Van Buren, E. CNS microvascular pericytes exhibit multipotential stem cell activity. J. Cereb. Blood Flow Metab. 2006, 26, 613–624. [Google Scholar] [CrossRef] [Green Version]
- Karow, M.; Sanchez, R.; Schichor, C.; Masserdotti, G.; Ortega, F.; Heinrich, C.; Gascon, S.; Khan, M.A.; Lie, D.C.; Dellavalle, A.; et al. Reprogramming of pericyte-derived cells of the adult human brain into induced neuronal cells. Cell Stem Cell 2012, 11, 471–476. [Google Scholar] [CrossRef] [Green Version]
- Nakagomi, T.; Kubo, S.; Nakano-Doi, A.; Sakuma, R.; Lu, S.; Narita, A.; Kawahara, M.; Taguchi, A.; Matsuyama, T. Brain vascular pericytes following ischemia have multipotential stem cell activity to differentiate into neural and vascular lineage cells. Stem Cells 2015, 33, 1962–1974. [Google Scholar] [CrossRef] [PubMed]
- Paul, G.; Ozen, I.; Christophersen, N.S.; Reinbothe, T.; Bengzon, J.; Visse, E.; Jansson, K.; Dannaeus, K.; Henriques-Oliveira, C.; Roybon, L.; et al. The adult human brain harbors multipotent perivascular mesenchymal stem cells. PLoS ONE 2012, 7, e35577. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, R.; Kawahara, M.; Nakano-Doi, A.; Takahashi, A.; Tanaka, Y.; Narita, A.; Kuwahara-Otani, S.; Hayakawa, T.; Yagi, H.; Matsuyama, T.; et al. Brain pericytes serve as microglia-generating multipotent vascular stem cells following ischemic stroke. J. Neuroinflamm. 2016, 13, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva Meirelles, L.; Caplan, A.I.; Nardi, N.B. In search of the in vivo identity of mesenchymal stem cells. Stem Cells 2008, 26, 2287–2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellavalle, A.; Maroli, G.; Covarello, D.; Azzoni, E.; Innocenzi, A.; Perani, L.; Antonini, S.; Sambasivan, R.; Brunelli, S.; Tajbakhsh, S.; et al. Pericytes resident in postnatal skeletal muscle differentiate into muscle fibres and generate satellite cells. Nat. Commun. 2011, 2, 499. [Google Scholar] [CrossRef] [Green Version]
- Sacchetti, B.; Funari, A.; Remoli, C.; Giannicola, G.; Kogler, G.; Liedtke, S.; Cossu, G.; Serafini, M.; Sampaolesi, M.; Tagliafico, E.; et al. No Identical “Mesenchymal Stem Cells” at Different Times and Sites: Human Committed Progenitors of Distinct Origin and Differentiation Potential Are Incorporated as Adventitial Cells in Microvessels. Stem Cell Rep. 2016, 6, 897–913. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes-Camboa, N.; Cattaneo, P.; Sun, Y.; Moore-Morris, T.; Gu, Y.; Dalton, N.D.; Rockenstein, E.; Masliah, E.; Peterson, K.L.; Stallcup, W.B.; et al. Pericytes of Multiple Organs Do Not Behave as Mesenchymal Stem Cells In Vivo. Cell Stem Cell 2017, 20, 345–359.e5. [Google Scholar] [CrossRef] [Green Version]
- Worsdorfer, P.; Ergun, S. Do Vascular Mural Cells Possess Endogenous Plasticity In Vivo? Stem Cell Rev. Rep. 2014, 14, 144–147. [Google Scholar] [CrossRef]
- Yianni, V.; Sharpe, P.T. Molecular Programming of Perivascular Stem Cell Precursors. Stem Cells 2018, 36, 1890–1904. [Google Scholar] [CrossRef] [Green Version]
- Vanlandewijck, M.; He, L.; Mae, M.A.; Andrae, J.; Ando, K.; Del Gaudio, F.; Nahar, K.; Lebouvier, T.; Lavina, B.; Gouveia, L.; et al. A molecular atlas of cell types and zonation in the brain vasculature. Nature 2018, 554, 475–480. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Vanlandewijck, M.; Mae, M.A.; Andrae, J.; Ando, K.; Del Gaudio, F.; Nahar, K.; Lebouvier, T.; Lavina, B.; Gouveia, L.; et al. Single-cell RNA sequencing of mouse brain and lung vascular and vessel-associated cell types. Sci. Data 2018, 5, 180160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karow, M.; Camp, J.G.; Falk, S.; Gerber, T.; Pataskar, A.; Gac-Santel, M.; Kageyama, J.; Brazovskaja, A.; Garding, A.; Fan, W.; et al. Direct pericyte-to-neuron reprogramming via unfolding of a neural stem cell-like program. Nat. Neurosci. 2018, 21, 932–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakata, M.; Nakagomi, T.; Maeda, M.; Nakano-Doi, A.; Momota, Y.; Matsuyama, T. Induction of Perivascular Neural Stem Cells and Possible Contribution to Neurogenesis Following Transient Brain Ischemia/Reperfusion Injury. Transl. Stroke Res. 2017, 8, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Farahani, R.M.; Rezaei-Lotfi, S.; Simonian, M.; Xaymardan, M.; Hunter, N. Neural microvascular pericytes contribute to human adult neurogenesis. J. Comp. Neurol. 2019, 527, 780–796. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.C.; Baily, J.E.; Corselli, M.; Diaz, M.E.; Sun, B.; Xiang, G.; Gray, G.A.; Huard, J.; Peault, B. Human myocardial pericytes: Multipotent mesodermal precursors exhibiting cardiac specificity. Stem Cells 2015, 33, 557–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierantozzi, E.; Vezzani, B.; Badin, M.; Curina, C.; Severi, F.M.; Petraglia, F.; Randazzo, D.; Rossi, D.; Sorrentino, V. Tissue-Specific Cultured Human Pericytes: Perivascular Cells from Smooth Muscle Tissue Have Restricted Mesodermal Differentiation Ability. Stem Cells Dev. 2016, 25, 674–686. [Google Scholar] [CrossRef] [Green Version]
- Vishvanath, L.; MacPherson, K.A.; Hepler, C.; Wang, Q.A.; Shao, M.; Spurgin, S.B.; Wang, M.Y.; Kusminski, C.M.; Morley, T.S.; Gupta, R.K. Pdgfrbeta+ Mural Preadipocytes Contribute to Adipocyte Hyperplasia Induced by High-Fat-Diet Feeding and Prolonged Cold Exposure in Adult Mice. Cell Metab. 2016, 23, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Holm, A.; Heumann, T.; Augustin, H.G. Microvascular Mural Cell Organotypic Heterogeneity and Functional Plasticity. Trends Cell. Biol. 2018, 28, 302–316. [Google Scholar] [CrossRef]
- Errede, M.; Girolamo, F.; Virgintino, D. High-Resolution Confocal Imaging of Pericytes in Human Fetal Brain Microvessels. Methods Mol. Biol. 2021, 2206, 143–150. [Google Scholar] [CrossRef]
- van Dijk, C.G.; Nieuweboer, F.E.; Pei, J.Y.; Xu, Y.J.; Burgisser, P.; van Mulligen, E.; el Azzouzi, H.; Duncker, D.J.; Verhaar, M.C.; Cheng, C. The complex mural cell: Pericyte function in health and disease. Int. J. Cardiol. 2015, 190, 75–89. [Google Scholar] [CrossRef]
- Birbrair, A.; Zhang, T.; Wang, Z.M.; Messi, M.L.; Mintz, A.; Delbono, O. Pericytes at the intersection between tissue regeneration and pathology. Clin. Sci. 2015, 128, 81–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girolamo, F.; de Trizio, I.; Errede, M.; Longo, G.; d’Amati, A.; Virgintino, D. Neural crest cell-derived pericytes act as pro-angiogenic cells in human neocortex development and gliomas. Fluids Barriers CNS 2021, 18, 14. [Google Scholar] [CrossRef] [PubMed]
- Stallcup, W.B. The NG2 proteoglycan: Past insights and future prospects. J. Neurocytol. 2002, 31, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Horner, P.J.; Thallmair, M.; Gage, F.H. Defining the NG2-expressing cell of the adult CNS. J. Neurocytol. 2002, 31, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Dellavalle, A.; Sampaolesi, M.; Tonlorenzi, R.; Tagliafico, E.; Sacchetti, B.; Perani, L.; Innocenzi, A.; Galvez, B.G.; Messina, G.; Morosetti, R.; et al. Pericytes of human skeletal muscle are myogenic precursors distinct from satellite cells. Nat. Cell Biol. 2007, 9, 255–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virgintino, D.; Girolamo, F.; Errede, M.; Capobianco, C.; Robertson, D.; Stallcup, W.B.; Perris, R.; Roncali, L. An intimate interplay between precocious, migrating pericytes and endothelial cells governs human fetal brain angiogenesis. Angiogenesis 2007, 10, 35–45. [Google Scholar] [CrossRef]
- Heldin, C.H.; Westermark, B. Mechanism of action and in vivo role of platelet-derived growth factor. Physiol. Rev. 1999, 79, 1283–1316. [Google Scholar] [CrossRef]
- He, L.; Vanlandewijck, M.; Raschperger, E.; Andaloussi Mae, M.; Jung, B.; Lebouvier, T.; Ando, K.; Hofmann, J.; Keller, A.; Betsholtz, C. Analysis of the brain mural cell transcriptome. Sci. Rep. 2016, 6, 35108. [Google Scholar] [CrossRef] [Green Version]
- Girolamo, F.; Errede, M.; Longo, G.; Annese, T.; Alias, C.; Ferrara, G.; Morando, S.; Trojano, M.; Kerlero de Rosbo, N.; Uccelli, A.; et al. Defining the role of NG2-expressing cells in experimental models of multiple sclerosis. A biofunctional analysis of the neurovascular unit in wild type and NG2 null mice. PLoS ONE 2019, 14, e0213508. [Google Scholar] [CrossRef] [Green Version]
- Alliot, F.; Rutin, J.; Leenen, P.J.; Pessac, B. Pericytes and periendothelial cells of brain parenchyma vessels co-express aminopeptidase N, aminopeptidase A, and nestin. J. Neurosci. Res. 1999, 58, 367–378. [Google Scholar] [CrossRef]
- Smyth, L.C.D.; Rustenhoven, J.; Scotter, E.L.; Schweder, P.; Faull, R.L.M.; Park, T.I.H.; Dragunow, M. Markers for human brain pericytes and smooth muscle cells. J. Chem. Neuroanat. 2018, 92, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Errede, M.; Mangieri, D.; Longo, G.; Girolamo, F.; de Trizio, I.; Vimercati, A.; Serio, G.; Frei, K.; Perris, R.; Virgintino, D. Tunneling nanotubes evoke pericyte/endothelial communication during normal and tumoral angiogenesis. Fluids Barriers CNS 2018, 15, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, M.J.; Muller, N.; Korschenhausen, D.; Kirsch, K.H.; Penning, R.; Ackenheil, M.; Johnson, J.P.; Hampel, H. Melanoma-associated adhesion molecule MUC18/MCAM (CD146) and transcriptional regulator mader in normal human CNS. Neuroimmunomodulation 1998, 5, 270–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacFadyen, J.; Savage, K.; Wienke, D.; Isacke, C.M. Endosialin is expressed on stromal fibroblasts and CNS pericytes in mouse embryos and is downregulated during development. Gene Expr. Patterns 2007, 7, 363–369. [Google Scholar] [CrossRef]
- Kim, S.Y.; Choi, J.; Roh, J.; Kim, C.H. NeuroTrace 500/525 identifies human induced pluripotent stem cell-derived brain pericyte-like cells. Mol. Brain 2022, 15, 11. [Google Scholar] [CrossRef]
- Damisah, E.C.; Hill, R.A.; Tong, L.; Murray, K.N.; Grutzendler, J. A fluoro-Nissl dye identifies pericytes as distinct vascular mural cells during in vivo brain imaging. Nat. Neurosci. 2017, 20, 1023–1032. [Google Scholar] [CrossRef] [Green Version]
- Eberth, C.J. Handbuch der Lehre von der Gewegen des Menschen und der Tiere; W. Engelman: Leipzig, Germany, 1871. [Google Scholar]
- Rouget, C. Note Sur Le Developpement De La Tunique Contractile Des Vaisseaux. Compt Rend Acad Sci 1874, 59, 559–562. [Google Scholar]
- Zimmermann, K.W. Der feinere Bau der Blutcapillaren. Z. Anat. Entwickl. 1923, 68, 29–109. [Google Scholar] [CrossRef]
- Bergwerff, M.; Verberne, M.E.; DeRuiter, M.C.; Poelmann, R.E.; Gittenberger-de Groot, A.C. Neural crest cell contribution to the developing circulatory system: Implications for vascular morphology? Circ. Res. 1998, 82, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Etchevers, H.C.; Vincent, C.; Le Douarin, N.M.; Couly, G.F. The cephalic neural crest provides pericytes and smooth muscle cells to all blood vessels of the face and forebrain. Development 2001, 128, 1059–1068. [Google Scholar] [CrossRef]
- Korn, J.; Christ, B.; Kurz, H. Neuroectodermal origin of brain pericytes and vascular smooth muscle cells. J. Comp. Neurol. 2002, 442, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.; Sheridan, J.; Veiga-Fernandes, H.; Roderick, K.; Pachnis, V.; Adams, R.; Blackburn, C.; Kioussis, D.; Coles, M. Contribution of neural crest-derived cells in the embryonic and adult thymus. J. Immunol. 2008, 180, 3183–3189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, S.M.; Stolt, C.C.; Terszowski, G.; Blum, C.; Amagai, T.; Kessaris, N.; Iannarelli, P.; Richardson, W.D.; Wegner, M.; Rodewald, H.R. Neural crest origin of perivascular mesenchyme in the adult thymus. J. Immunol. 2008, 180, 5344–5351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilm, B.; Ipenberg, A.; Hastie, N.D.; Burch, J.B.; Bader, D.M. The serosal mesothelium is a major source of smooth muscle cells of the gut vasculature. Development 2005, 132, 5317–5328. [Google Scholar] [CrossRef] [Green Version]
- Que, J.; Wilm, B.; Hasegawa, H.; Wang, F.; Bader, D.; Hogan, B.L. Mesothelium contributes to vascular smooth muscle and mesenchyme during lung development. Proc. Natl. Acad. Sci. USA 2008, 105, 16626–16630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asahina, K.; Zhou, B.; Pu, W.T.; Tsukamoto, H. Septum transversum-derived mesothelium gives rise to hepatic stellate cells and perivascular mesenchymal cells in developing mouse liver. Hepatology 2011, 53, 983–995. [Google Scholar] [CrossRef] [Green Version]
- Faal, T.; Phan, D.T.T.; Davtyan, H.; Scarfone, V.M.; Varady, E.; Blurton-Jones, M.; Hughes, C.C.W.; Inlay, M.A. Induction of Mesoderm and Neural Crest-Derived Pericytes from Human Pluripotent Stem Cells to Study Blood-Brain Barrier Interactions. Stem Cell Rep. 2019, 12, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Gage, P.J.; Rhoades, W.; Prucka, S.K.; Hjalt, T. Fate maps of neural crest and mesoderm in the mammalian eye. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4200–4208. [Google Scholar] [CrossRef]
- Trost, A.; Schroedl, F.; Lange, S.; Rivera, F.J.; Tempfer, H.; Korntner, S.; Stolt, C.C.; Wegner, M.; Bogner, B.; Kaser-Eichberger, A.; et al. Neural crest origin of retinal and choroidal pericytes. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7910–7921. [Google Scholar] [CrossRef] [Green Version]
- Lamagna, C.; Bergers, G. The bone marrow constitutes a reservoir of pericyte progenitors. J. Leukoc. Biol. 2006, 80, 677–681. [Google Scholar] [CrossRef] [Green Version]
- Prazeres, P.; Almeida, V.M.; Lousado, L.; Andreotti, J.P.; Paiva, A.E.; Santos, G.S.P.; Azevedo, P.O.; Souto, L.; Almeida, G.G.; Filev, R.; et al. Macrophages Generate Pericytes in the Developing Brain. Cell Mol. Neurobiol. 2018, 38, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Ozerdem, U.; Alitalo, K.; Salven, P.; Li, A. Contribution of bone marrow-derived pericyte precursor cells to corneal vasculogenesis. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3502–3506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sierra, R.; Gomez Bustillo, S.; Kameneva, P.; Fiore, E.J.; Mazzone, G.L.; Borda, M.; Blanco, M.V.; Usuardi, C.; Furlan, A.; Ernfors, P.; et al. Contribution of neural crest and GLAST(+) Wnt1(+) bone marrow pericytes with liver fibrogenesis and/or regeneration. Liver Int. 2020, 40, 977–987. [Google Scholar] [CrossRef]
- Miller, S.R.; Perera, S.N.; Baker, C.V. Constitutively active Notch1 converts cranial neural crest-derived frontonasal mesenchyme to perivascular cells in vivo. Biol. Open 2017, 6, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Leveen, P.; Pekny, M.; Gebre-Medhin, S.; Swolin, B.; Larsson, E.; Betsholtz, C. Mice deficient for PDGF B show renal, cardiovascular, and hematological abnormalities. Genes Dev. 1994, 8, 1875–1887. [Google Scholar] [CrossRef] [Green Version]
- Soriano, P. Abnormal kidney development and hematological disorders in PDGF beta-receptor mutant mice. Genes Dev. 1994, 8, 1888–1896. [Google Scholar] [CrossRef] [Green Version]
- Pallone, T.L.; Silldorff, E.P.; Turner, M.R. Intrarenal blood flow: Microvascular anatomy and the regulation of medullary perfusion. Clin. Exp. Pharmacol. Physiol. 1998, 25, 383–392. [Google Scholar] [CrossRef]
- Hellstrom, M.; Gerhardt, H.; Kalen, M.; Li, X.; Eriksson, U.; Wolburg, H.; Betsholtz, C. Lack of pericytes leads to endothelial hyperplasia and abnormal vascular morphogenesis. J. Cell Biol. 2001, 153, 543–553. [Google Scholar] [CrossRef] [Green Version]
- Enge, M.; Bjarnegard, M.; Gerhardt, H.; Gustafsson, E.; Kalen, M.; Asker, N.; Hammes, H.P.; Shani, M.; Fassler, R.; Betsholtz, C. Endothelium-specific platelet-derived growth factor-B ablation mimics diabetic retinopathy. EMBO J. 2002, 21, 4307–4316. [Google Scholar] [CrossRef] [Green Version]
- Peppiatt, C.M.; Howarth, C.; Mobbs, P.; Attwell, D. Bidirectional control of CNS capillary diameter by pericytes. Nature 2006, 443, 700–704. [Google Scholar] [CrossRef] [Green Version]
- Hall, C.N.; Reynell, C.; Gesslein, B.; Hamilton, N.B.; Mishra, A.; Sutherland, B.A.; O’Farrell, F.M.; Buchan, A.M.; Lauritzen, M.; Attwell, D. Capillary pericytes regulate cerebral blood flow in health and disease. Nature 2014, 508, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, D.Y.; Lee, J.; Kim, J.; Kim, K.; Hong, S.; Han, S.; Kubota, Y.; Augustin, H.G.; Ding, L.; Kim, J.W.; et al. Plastic roles of pericytes in the blood-retinal barrier. Nat. Commun. 2017, 8, 15296. [Google Scholar] [CrossRef] [PubMed]
- Papapetropoulos, A.; Fulton, D.; Mahboubi, K.; Kalb, R.G.; O’Connor, D.S.; Li, F.; Altieri, D.C.; Sessa, W.C. Angiopoietin-1 inhibits endothelial cell apoptosis via the Akt/survivin pathway. J. Biol. Chem. 2000, 275, 9102–9105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, W.B.; Bohnsack, B.L.; Faske, J.B.; Anthis, N.J.; Bayless, K.J.; Hirschi, K.K.; Davis, G.E. Coregulation of vascular tube stabilization by endothelial cell TIMP-2 and pericyte TIMP-3. J. Cell Biol. 2006, 175, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Rucker, H.K.; Wynder, H.J.; Thomas, W.E. Cellular mechanisms of CNS pericytes. Brain Res. Bull. 2000, 51, 363–369. [Google Scholar] [CrossRef]
- Dohgu, S.; Takata, F.; Yamauchi, A.; Nakagawa, S.; Egawa, T.; Naito, M.; Tsuruo, T.; Sawada, Y.; Niwa, M.; Kataoka, Y. Brain pericytes contribute to the induction and up-regulation of blood-brain barrier functions through transforming growth factor-beta production. Brain Res. 2005, 1038, 208–215. [Google Scholar] [CrossRef]
- Hawkins, B.T.; Davis, T.P. The blood-brain barrier/neurovascular unit in health and disease. Pharmacol. Rev. 2005, 57, 173–185. [Google Scholar] [CrossRef]
- Nakagawa, S.; Deli, M.A.; Nakao, S.; Honda, M.; Hayashi, K.; Nakaoke, R.; Kataoka, Y.; Niwa, M. Pericytes from brain microvessels strengthen the barrier integrity in primary cultures of rat brain endothelial cells. Cell Mol. Neurobiol. 2007, 27, 687–694. [Google Scholar] [CrossRef] [Green Version]
- Armulik, A.; Genove, G.; Mae, M.; Nisancioglu, M.H.; Wallgard, E.; Niaudet, C.; He, L.; Norlin, J.; Lindblom, P.; Strittmatter, K.; et al. Pericytes regulate the blood-brain barrier. Nature 2010, 468, 557–561. [Google Scholar] [CrossRef] [Green Version]
- Daneman, R.; Zhou, L.; Kebede, A.A.; Barres, B.A. Pericytes are required for blood-brain barrier integrity during embryogenesis. Nature 2010, 468, 562–566. [Google Scholar] [CrossRef] [Green Version]
- Al Ahmad, A.; Taboada, C.B.; Gassmann, M.; Ogunshola, O.O. Astrocytes and pericytes differentially modulate blood-brain barrier characteristics during development and hypoxic insult. J. Cereb. Blood Flow Metab. 2011, 31, 693–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonkowski, D.; Katyshev, V.; Balabanov, R.D.; Borisov, A.; Dore-Duffy, P. The CNS microvascular pericyte: Pericyte-astrocyte crosstalk in the regulation of tissue survival. Fluids Barriers CNS 2011, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villabona-Rueda, A.; Erice, C.; Pardo, C.A.; Stins, M.F. The Evolving Concept of the Blood Brain Barrier (BBB): From a Single Static Barrier to a Heterogeneous and Dynamic Relay Center. Front. Cell. Neurosci. 2019, 13, 405. [Google Scholar] [CrossRef]
- De Luca, C.; Colangelo, A.M.; Virtuoso, A.; Alberghina, L.; Papa, M. Neurons, Glia, Extracellular Matrix and Neurovascular Unit: A Systems Biology Approach to the Complexity of Synaptic Plasticity in Health and Disease. Int. J. Mol. Sci. 2020, 21, 1539. [Google Scholar] [CrossRef] [Green Version]
- Uemura, M.T.; Maki, T.; Ihara, M.; Lee, V.M.Y.; Trojanowski, J.Q. Brain Microvascular Pericytes in Vascular Cognitive Impairment and Dementia. Front. Aging Neurosci. 2020, 12, 80. [Google Scholar] [CrossRef] [Green Version]
- Winkler, E.A.; Sagare, A.P.; Zlokovic, B.V. The Pericyte: A Forgotten Cell Type with Important Implications for Alzheimer’s Disease? Brain Pathol 2014, 24, 371–386. [Google Scholar] [CrossRef]
- Balabanov, R.; Dore-Duffy, P. Role of the Cns Microvascular Pericyte in the Blood-Brain Barrier. J. Neurosci. Res. 1998, 53, 637–644. [Google Scholar] [CrossRef]
- Attwell, D.; Mishra, A.; Hall, C.N.; O’Farrell, F.M.; Dalkara, T. What Is a Pericyte? J. Cereb. Blood Flow Metab. 2016, 36, 451–455. [Google Scholar] [CrossRef] [Green Version]
- Winkler, E.A.; Bell, R.D.; Zlokovic, B.V. Central Nervous System Pericytes in Health and Disease. Nat. Neurosci. 2011, 14, 1398–1405. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, N.B.; Attwell, D.; Hall, C.N. Pericyte-mediated regulation of capillary diameter: A component of neurovascular coupling in health and disease. Front. Neuroenergetics 2010, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Kovacs-Oller, T.; Ivanova, E.; Bianchimano, P.; Sagdullaev, B.T. The pericyte connectome: Spatial precision of neurovascular coupling is driven by selective connectivity maps of pericytes and endothelial cells and is disrupted in diabetes. Cell Discov. 2020, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.; Kovacs-Oller, T.; Sagdullaev, B.T. Vascular Pericyte Impairment and Connexin43 Gap Junction Deficit Contribute to Vasomotor Decline in Diabetic Retinopathy. J. Neurosci. 2017, 37, 7580–7594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gundersen, G.A.; Vindedal, G.F.; Skare, O.; Nagelhus, E.A. Evidence that pericytes regulate aquaporin-4 polarization in mouse cortical astrocytes. Brain Struct. Funct. 2014, 219, 2181–2186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaegebeur, A.; Segura, I.; Carmeliet, P. Pericytes: Blood-brain barrier safeguards against neurodegeneration? Neuron 2010, 68, 321–323. [Google Scholar] [CrossRef] [Green Version]
- Baumann, J.; Tsao, C.C.; Patkar, S.; Huang, S.F.; Francia, S.; Magnussen, S.N.; Gassmann, M.; Vogel, J.; Koster-Hegmann, C.; Ogunshola, O.O. Pericyte, but not astrocyte, hypoxia inducible factor-1 (HIF-1) drives hypoxia-induced vascular permeability in vivo. Fluids Barriers CNS 2022, 19, 6. [Google Scholar] [CrossRef]
- Bell, R.D.; Winkler, E.A.; Sagare, A.P.; Singh, I.; LaRue, B.; Deane, R.; Zlokovic, B.V. Pericytes Control Key Neurovascular Functions and Neuronal Phenotype in the Adult Brain and During Brain Aging. Neuron 2010, 68, 409–427. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D.; Nico, B.; Crivellato, E. The role of pericytes in angiogenesis. Int. J. Dev. Biol. 2011, 55, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D.; Tamma, R.; Annese, T. The role of vascular niche and endothelial cells in organogenesis and regeneration. Exp. Cell Res. 2021, 398, 112398. [Google Scholar] [CrossRef]
- Ozerdem, U.; Stallcup, W.B. Early Contribution of Pericytes to Angiogenic Sprouting and Tube Formation. Angiogenesis 2003, 6, 241–249. [Google Scholar] [CrossRef] [Green Version]
- Virgintino, D.; Ozerdem, U.; Girolamo, F.; Roncali, L.; Stallcup, W.B.; Perris, R. Reversal of cellular roles in angiogenesis: Implications for anti-angiogenic therapy. J. Vasc. Res. 2008, 45, 129–131. [Google Scholar] [CrossRef] [Green Version]
- Nehls, V.; Denzer, K.; Drenckhahn, D. Pericyte involvement in capillary sprouting during angiogenesis in situ. Cell Tissue Res. 1992, 270, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Nystrom, H.C.; Lindblom, P.; Wickman, A.; Andersson, I.; Norlin, J.; Faldt, J.; Lindahl, P.; Skott, O.; Bjarnegard, M.; Fitzgerald, S.M.; et al. Platelet-derived growth factor B retention is essential for development of normal structure and function of conduit vessels and capillaries. Cardiovasc. Res. 2006, 71, 557–565. [Google Scholar] [CrossRef] [Green Version]
- Hori, S.; Ohtsuki, S.; Hosoya, K.; Nakashima, E.; Terasaki, T. A pericyte-derived angiopoietin-1 multimeric complex induces occludin gene expression in brain capillary endothelial cells through Tie-2 activation in vitro. J. Neurochem. 2004, 89, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Dore-Duffy, P. Pericytes: Pluripotent Cells of the Blood Brain Barrier. Curr. Pharm. Des. 2008, 14, 1581–1593. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.R.; Mayanil, C.S.; Kim, K.S.; Tomita, T. Angiopoietin-1 Regulates Brain Endothelial Permeability through Ptpn-2 Mediated Tyrosine Dephosphorylation of Occludin. PLoS ONE 2015, 10, e0130857. [Google Scholar] [CrossRef]
- Dore-Duffy, P.; Cleary, K. Morphology and properties of pericytes. Methods Mol. Biol. 2011, 686, 49–68. [Google Scholar] [CrossRef]
- Jung, B.; Arnold, T.D.; Raschperger, E.; Gaengel, K.; Betsholtz, C. Visualization of vascular mural cells in developing brain using genetically labeled transgenic reporter mice. J. Cereb. Blood Flow Metab. 2018, 38, 456–468. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Ling, T.W.; Rewell, S.S.; Hare, D.L.; Howells, D.W.; Kourakis, A.; Wookey, P.J. A novel population of alpha-smooth muscle actin-positive cells activated in a rat model of stroke: An analysis of the spatio-temporal distribution in response to ischemia. J. Cereb. Blood Flow Metab. 2012, 32, 2055–2065. [Google Scholar] [CrossRef] [Green Version]
- Ben-Zvi, A.; Lacoste, B.; Kur, E.; Andreone, B.J.; Mayshar, Y.; Yan, H.; Gu, C. Mfsd2a is critical for the formation and function of the blood-brain barrier. Nature 2014, 509, 507–511. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Zlokovic, B.V. Blood-brain barrier: A dual life of MFSD2A? Neuron 2014, 82, 728–730. [Google Scholar] [CrossRef] [Green Version]
- Trost, A.; Lange, S.; Schroedl, F.; Bruckner, D.; Motloch, K.A.; Bogner, B.; Kaser-Eichberger, A.; Strohmaier, C.; Runge, C.; Aigner, L.; et al. Brain and Retinal Pericytes: Origin, Function and Role. Front. Cell. Neurosci. 2016, 10, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmina, A.B.; Morgun, A.V.; Kuvacheva, N.V.; Lopatina, O.L.; Komleva, Y.K.; Malinovskaya, N.A.; Pozhilenkova, E.A. Establishment of neurogenic microenvironment in the neurovascular unit: The connexin 43 story. Rev. Neurosci. 2014, 25, 97–111. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, A.G.; Lange, S.; Silva, M.E.; Gonzalez, G.A.; Tempfer, H.; van Wijngaarden, P.; Zhao, C.; Di Canio, L.; Trost, A.; Bieler, L.; et al. Pericytes Stimulate Oligodendrocyte Progenitor Cell Differentiation during CNS Remyelination. Cell Rep. 2017, 20, 1755–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, T.; Maeda, M.; Uemura, M.; Lo, E.K.; Terasaki, Y.; Liang, A.C.; Shindo, A.; Choi, Y.K.; Taguchi, A.; Matsuyama, T.; et al. Potential interactions between pericytes and oligodendrocyte precursor cells in perivascular regions of cerebral white matter. Neurosci. Lett. 2015, 597, 164–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozhilenkova, E.A.; Lopatina, O.L.; Komleva, Y.K.; Salmin, V.V.; Salmina, A.B. Blood-brain barrier-supported neurogenesis in healthy and diseased brain. Rev. Neurosci. 2017, 28, 397–415. [Google Scholar] [CrossRef] [PubMed]
- Ottone, C.; Krusche, B.; Whitby, A.; Clements, M.; Quadrato, G.; Pitulescu, M.E.; Adams, R.H.; Parrinello, S. Direct cell-cell contact with the vascular niche maintains quiescent neural stem cells. Nat. Cell Biol. 2014, 16, 1045–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crouch, E.E.; Liu, C.; Silva-Vargas, V.; Doetsch, F. Regional and stage-specific effects of prospectively purified vascular cells on the adult V-SVZ neural stem cell lineage. J. Neurosci. 2015, 35, 4528–4539. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.E.; Lange, S.; Hinrichsen, B.; Philp, A.R.; Reyes, C.R.; Halabi, D.; Mansilla, J.B.; Rotheneichner, P.; Guzman de la Fuente, A.; Couillard-Despres, S.; et al. Pericytes Favor Oligodendrocyte Fate Choice in Adult Neural Stem Cells. Front. Cell. Neurosci. 2019, 13, 85. [Google Scholar] [CrossRef] [Green Version]
- Shibahara, T.; Ago, T.; Nakamura, K.; Tachibana, M.; Yoshikawa, Y.; Komori, M.; Yamanaka, K.; Wakisaka, Y.; Kitazono, T. Pericyte-Mediated Tissue Repair through PDGFRbeta Promotes Peri-Infarct Astrogliosis, Oligodendrogenesis, and Functional Recovery after Acute Ischemic Stroke. eNeuro 2020, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Tsai, H.H.; Niu, J.; Munji, R.; Davalos, D.; Chang, J.; Zhang, H.; Tien, A.C.; Kuo, C.J.; Chan, J.R.; Daneman, R.; et al. Oligodendrocyte precursors migrate along vasculature in the developing nervous system. Science 2016, 351, 379–384. [Google Scholar] [CrossRef] [Green Version]
- Gaceb, A.; Ozen, I.; Padel, T.; Barbariga, M.; Paul, G. Pericytes secrete pro-regenerative molecules in response to platelet-derived growth factor-BB. J. Cereb. Blood Flow Metab. 2018, 38, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinholm, J.E.; Hamilton, N.B.; Kessaris, N.; Richardson, W.D.; Bergersen, L.H.; Attwell, D. Regulation of oligodendrocyte development and myelination by glucose and lactate. J. Neurosci. 2011, 31, 538–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montagne, A.; Nikolakopoulou, A.M.; Zhao, Z.; Sagare, A.P.; Si, G.; Lazic, D.; Barnes, S.R.; Daianu, M.; Ramanathan, A.; Go, A.; et al. Pericyte degeneration causes white matter dysfunction in the mouse central nervous system. Nat. Med. 2018, 24, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Greenwood-Goodwin, M.; Yang, J.; Hassanipour, M.; Larocca, D. A novel lineage restricted, pericyte-like cell line isolated from human embryonic stem cells. Sci. Rep. 2016, 6, 24403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; Brookes, O.; Battaglia, G. Pericytes from Mesenchymal Stem Cells as a model for the blood-brain barrier. Sci. Rep. 2017, 7, 39676. [Google Scholar] [CrossRef]
- Yamamizu, K.; Iwasaki, M.; Takakubo, H.; Sakamoto, T.; Ikuno, T.; Miyoshi, M.; Kondo, T.; Nakao, Y.; Nakagawa, M.; Inoue, H.; et al. In Vitro Modeling of Blood-Brain Barrier with Human iPSC-Derived Endothelial Cells, Pericytes, Neurons, and Astrocytes via Notch Signaling. Stem Cell Rep. 2017, 8, 634–647. [Google Scholar] [CrossRef]
- Chin, E.; Goh, E. Blood-brain barrier on a chip. Methods Cell Biol. 2018, 146, 159–182. [Google Scholar] [CrossRef]
- Campisi, M.; Lim, S.H.; Chiono, V.; Kamm, R.D. 3D Self-Organized Human Blood-Brain Barrier in a Microfluidic Chip. Methods Mol. Biol. 2021, 2258, 205–219. [Google Scholar] [CrossRef]
- Pediaditakis, I.; Kodella, K.R.; Manatakis, D.V.; Le, C.Y.; Hinojosa, C.D.; Tien-Street, W.; Manolakos, E.S.; Vekrellis, K.; Hamilton, G.A.; Ewart, L.; et al. Modeling alpha-synuclein pathology in a human brain-chip to assess blood-brain barrier disruption. Nat. Commun. 2021, 12, 5907. [Google Scholar] [CrossRef]
- Persidsky, Y.; Ramirez, S.H.; Haorah, J.; Kanmogne, G.D. Blood-brain barrier: Structural components and function under physiologic and pathologic conditions. J. Neuroimmune Pharmacol. 2006, 1, 223–236. [Google Scholar] [CrossRef]
- Montagne, A.; Barnes, S.R.; Sweeney, M.D.; Halliday, M.R.; Sagare, A.P.; Zhao, Z.; Toga, A.W.; Jacobs, R.E.; Liu, C.Y.; Amezcua, L.; et al. Blood-brain barrier breakdown in the aging human hippocampus. Neuron 2015, 85, 296–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, P.T.; Jicha, G.A.; Wang, W.X.; Ighodaro, E.; Artiushin, S.; Nichols, C.G.; Fardo, D.W. ABCC9/SUR2 in the brain: Implications for hippocampal sclerosis of aging and a potential therapeutic target. Ageing Res. Rev. 2015, 24, 111–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanimirovic, D.B.; Friedman, A. Pathophysiology of the neurovascular unit: Disease cause or consequence? J. Cereb. Blood Flow Metab. 2012, 32, 1207–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronson, J.K.; Ferner, R.E. Biomarkers—A General Review. Curr. Protoc. Pharmacol. 2017, 76, 9.23.1–9.23.17. [Google Scholar] [CrossRef]
- Olsson, B.; Lautner, R.; Andreasson, U.; Ohrfelt, A.; Portelius, E.; Bjerke, M.; Holtta, M.; Rosen, C.; Olsson, C.; Strobel, G.; et al. CSF and blood biomarkers for the diagnosis of Alzheimer’s disease: A systematic review and meta-analysis. Lancet Neurol. 2016, 15, 673–684. [Google Scholar] [CrossRef]
- Kuhle, J.; Barro, C.; Andreasson, U.; Derfuss, T.; Lindberg, R.; Sandelius, A.; Liman, V.; Norgren, N.; Blennow, K.; Zetterberg, H. Comparison of three analytical platforms for quantification of the neurofilament light chain in blood samples: ELISA, electrochemiluminescence immunoassay and Simoa. Clin. Chem. Lab. Med. 2016, 54, 1655–1661. [Google Scholar] [CrossRef]
- Barthelemy, N.R.; Horie, K.; Sato, C.; Bateman, R.J. Blood plasma phosphorylated-tau isoforms track CNS change in Alzheimer’s disease. J. Exp. Med. 2020, 217, e20200861. [Google Scholar] [CrossRef]
- Karikari, T.K.; Pascoal, T.A.; Ashton, N.J.; Janelidze, S.; Benedet, A.L.; Rodriguez, J.L.; Chamoun, M.; Savard, M.; Kang, M.S.; Therriault, J.; et al. Blood phosphorylated tau 181 as a biomarker for Alzheimer’s disease: A diagnostic performance and prediction modelling study using data from four prospective cohorts. Lancet Neurol. 2020, 19, 422–433. [Google Scholar] [CrossRef]
- Atici, Y.; Alehan, F.; Sezer, T.; Tuygun, N.; Haberal, A.; Yazici, A.C.; Karacan, C.D. Serum S100B levels in children with simple febrile seizures. Seizure 2012, 21, 175–177. [Google Scholar] [CrossRef] [Green Version]
- Mikkonen, K.; Pekkala, N.; Pokka, T.; Romner, B.; Uhari, M.; Rantala, H. S100B proteins in febrile seizures. Seizure 2012, 21, 144–146. [Google Scholar] [CrossRef] [Green Version]
- Calik, M.; Abuhandan, M.; Sonmezler, A.; Kandemir, H.; Oz, I.; Taskin, A.; Selek, S.; Iscan, A. Elevated serum S-100B levels in children with temporal lobe epilepsy. Seizure 2013, 22, 99–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calik, M.; Abuhandan, M.; Kandemir, H.; Guzel, B.; Solmaz, A.; Celik, H.; Taskin, A.; Iscan, A. Interictal serum S-100B protein levels in intractable epilepsy: A case-control study. Neurosci. Lett. 2014, 558, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Arango-Lievano, M.; Boussadia, B.; De Terdonck, L.D.T.; Gault, C.; Fontanaud, P.; Lafont, C.; Mollard, P.; Marchi, N.; Jeanneteau, F. Topographic Reorganization of Cerebrovascular Mural Cells under Seizure Conditions. Cell Rep. 2018, 23, 1045–1059. [Google Scholar] [CrossRef] [Green Version]
- Gulati, G.; Iffland, P.H., 2nd; Janigro, D.; Zhang, B.; Luggen, M.E. Anti-NR2 antibodies, blood-brain barrier, and cognitive dysfunction. Clin. Rheumatol. 2016, 35, 2989–2997. [Google Scholar] [CrossRef]
- Blyth, B.J.; Farhavar, A.; Gee, C.; Hawthorn, B.; He, H.; Nayak, A.; Stocklein, V.; Bazarian, J.J. Validation of serum markers for blood-brain barrier disruption in traumatic brain injury. J. Neurotrauma 2009, 26, 1497–1507. [Google Scholar] [CrossRef]
- Dadas, A.; Washington, J.; Diaz-Arrastia, R.; Janigro, D. Biomarkers in traumatic brain injury (TBI): A review. Neuropsychiatr. Dis. Treat. 2018, 14, 2989–3000. [Google Scholar] [CrossRef] [Green Version]
- Janigro, D.; Bailey, D.M.; Lehmann, S.; Badaut, J.; O’Flynn, R.; Hirtz, C.; Marchi, N. Peripheral Blood and Salivary Biomarkers of Blood-Brain Barrier Permeability and Neuronal Damage: Clinical and Applied Concepts. Front. Neurol. 2020, 11, 577312. [Google Scholar] [CrossRef]
- Malik, V.A.; Di Benedetto, B. The Blood-Brain Barrier and the EphR/Ephrin System: Perspectives on a Link Between Neurovascular and Neuropsychiatric Disorders. Front. Mol. Neurosci. 2018, 11, 127. [Google Scholar] [CrossRef]
- Kanner, A.A.; Marchi, N.; Fazio, V.; Mayberg, M.R.; Koltz, M.T.; Siomin, V.; Stevens, G.H.; Masaryk, T.; Aumayr, B.; Vogelbaum, M.A.; et al. Serum S100beta: A noninvasive marker of blood-brain barrier function and brain lesions. Cancer 2003, 97, 2806–2813. [Google Scholar] [CrossRef] [Green Version]
- Vogelbaum, M.A.; Masaryk, T.; Mazzone, P.; Mekhail, T.; Fazio, V.; McCartney, S.; Marchi, N.; Kanner, A.; Janigro, D. S100beta as a predictor of brain metastases: Brain versus cerebrovascular damage. Cancer 2005, 104, 817–824. [Google Scholar] [CrossRef]
- Azmitia, E.C.; Saccomano, Z.T.; Alzoobaee, M.F.; Boldrini, M.; Whitaker-Azmitia, P.M. Persistent Angiogenesis in the Autism Brain: An Immunocytochemical Study of Postmortem Cortex, Brainstem and Cerebellum. J. Autism Dev. Disord. 2016, 46, 1307–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zengeler, K.E.; Lukens, J.R. Innate immunity at the crossroads of healthy brain maturation and neurodevelopmental disorders. Nat. Rev. Immunol. 2021, 21, 454–468. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Itoh, H.; Tsugawa, Y.; Nishida, Y.; Kurata, K.; Uemura, A.; Miyata, T. Embryonic pericytes promote microglial homeostasis and their effects on neural progenitors in the developing cerebral cortex. J. Neurosci. 2022, 42, 362–376. [Google Scholar] [CrossRef] [PubMed]
- Kadhim, H.; Tabarki, B.; Verellen, G.; De Prez, C.; Rona, A.M.; Sebire, G. Inflammatory cytokines in the pathogenesis of periventricular leukomalacia. Neurology 2001, 56, 1278–1284. [Google Scholar] [CrossRef] [PubMed]
- Back, S.A.; Han, B.H.; Luo, N.L.; Chricton, C.A.; Xanthoudakis, S.; Tam, J.; Arvin, K.L.; Holtzman, D.M. Selective vulnerability of late oligodendrocyte progenitors to hypoxia-ischemia. J. Neurosci. 2002, 22, 455–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Y.; Cai, Z.; Rhodes, P.G. Disturbance of oligodendrocyte development, hypomyelination and white matter injury in the neonatal rat brain after intracerebral injection of lipopolysaccharide. Brain Res. Dev. Brain Res. 2003, 140, 205–214. [Google Scholar] [CrossRef]
- Rousset, C.I.; Chalon, S.; Cantagrel, S.; Bodard, S.; Andres, C.; Gressens, P.; Saliba, E. Maternal exposure to LPS induces hypomyelination in the internal capsule and programmed cell death in the deep gray matter in newborn rats. Pediatr. Res. 2006, 59, 428–433. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Gao, C.; Gao, D.; Sun, R.; Li, W.; Wang, F.; Wang, Y.; Cao, H.; Zhou, G.; Zhang, J.; et al. Reduction in pericyte coverage leads to blood-brain barrier dysfunction via endothelial transcytosis following chronic cerebral hypoperfusion. Fluids Barriers CNS 2021, 18, 21. [Google Scholar] [CrossRef]
- Castillo-Melendez, M.; Yawno, T.; Allison, B.J.; Jenkin, G.; Wallace, E.M.; Miller, S.L. Cerebrovascular adaptations to chronic hypoxia in the growth restricted lamb. Int. J. Dev. Neurosci. 2015, 45, 55–65. [Google Scholar] [CrossRef]
- Farkas, E.; Luiten, P.G. Cerebral microvascular pathology in aging and Alzheimer’s disease. Prog. Neurobiol. 2001, 64, 575–611. [Google Scholar] [CrossRef] [Green Version]
- Baloyannis, S.J.; Baloyannis, I.S. The vascular factor in Alzheimer’s disease: A study in Golgi technique and electron microscopy. J. Neurol. Sci. 2012, 322, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Sengillo, J.D.; Winkler, E.A.; Walker, C.T.; Sullivan, J.S.; Johnson, M.; Zlokovic, B.V. Deficiency in mural vascular cells coincides with blood-brain barrier disruption in Alzheimer’s disease. Brain Pathol. 2013, 23, 303–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliday, M.R.; Rege, S.V.; Ma, Q.; Zhao, Z.; Miller, C.A.; Winkler, E.A.; Zlokovic, B.V. Accelerated pericyte degeneration and blood-brain barrier breakdown in apolipoprotein E4 carriers with Alzheimer’s disease. J. Cereb. Blood Flow Metab. 2016, 36, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Schultz, N.; Byman, E.; Fex, M.; Wennstrom, M. Amylin alters human brain pericyte viability and NG2 expression. J. Cereb. Blood Flow Metab. 2017, 37, 1470–1482. [Google Scholar] [CrossRef]
- Miners, J.S.; Schulz, I.; Love, S. Differing associations between Abeta accumulation, hypoperfusion, blood-brain barrier dysfunction and loss of PDGFRB pericyte marker in the precuneus and parietal white matter in Alzheimer’s disease. J. Cereb. Blood Flow Metab. 2018, 38, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Valbuena, I.; Valenti-Azcarate, R.; Amat-Villegas, I.; Riverol, M.; Marcilla, I.; de Andrea, C.E.; Sanchez-Arias, J.A.; Del Mar Carmona-Abellan, M.; Marti, G.; Erro, M.E.; et al. Amylin as a potential link between type 2 diabetes and alzheimer disease. Ann. Neurol. 2019, 86, 539–551. [Google Scholar] [CrossRef]
- Hardy, J.A.; Higgins, G.A. Alzheimer’s disease: The amyloid cascade hypothesis. Science 1992, 256, 184–185. [Google Scholar] [CrossRef]
- Arranz, A.M.; De Strooper, B. The role of astroglia in Alzheimer’s disease: Pathophysiology and clinical implications. Lancet Neurol. 2019, 18, 406–414. [Google Scholar] [CrossRef]
- Calvo-Rodriguez, M.; Bacskai, B.J. Mitochondria and Calcium in Alzheimer’s Disease: From Cell Signaling to Neuronal Cell Death. Trends Neurosci. 2021, 44, 136–151. [Google Scholar] [CrossRef]
- Biffi, A.; Greenberg, S.M. Cerebral amyloid angiopathy: A systematic review. J. Clin. Neurol. 2011, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, S.M.; Bacskai, B.J.; Hernandez-Guillamon, M.; Pruzin, J.; Sperling, R.; van Veluw, S.J. Cerebral amyloid angiopathy and Alzheimer disease—One peptide, two pathways. Nat. Rev. Neurol. 2020, 16, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Grammas, P.; Moore, P.; Weigel, P.H. Microvessels from Alzheimer’s disease brains kill neurons in vitro. Am. J. Pathol. 1999, 154, 337–342. [Google Scholar] [CrossRef]
- Pluta, R.; Jablonski, M.; Ulamek-Koziol, M.; Kocki, J.; Brzozowska, J.; Januszewski, S.; Furmaga-Jablonska, W.; Bogucka-Kocka, A.; Maciejewski, R.; Czuczwar, S.J. Sporadic Alzheimer’s disease begins as episodes of brain ischemia and ischemically dysregulated Alzheimer’s disease genes. Mol. Neurobiol. 2013, 48, 500–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thal, D.R.; Walter, J.; Saido, T.C.; Fandrich, M. Neuropathology and biochemistry of Abeta and its aggregates in Alzheimer’s disease. Acta Neuropathol. 2015, 129, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Raz, L.; Knoefel, J.; Bhaskar, K. The neuropathology and cerebrovascular mechanisms of dementia. J. Cereb. Blood Flow Metab. 2016, 36, 172–186. [Google Scholar] [CrossRef] [Green Version]
- Damodarasamy, M.; Vernon, R.B.; Pathan, J.L.; Keene, C.D.; Day, A.J.; Banks, W.A.; Reed, M.J. The microvascular extracellular matrix in brains with Alzheimer’s disease neuropathologic change (ADNC) and cerebral amyloid angiopathy (CAA). Fluids Barriers CNS 2020, 17, 60. [Google Scholar] [CrossRef]
- Thomas, D.X.; Bajaj, S.; McRae-McKee, K.; Hadjichrysanthou, C.; Anderson, R.M.; Collinge, J. Association of TDP-43 proteinopathy, cerebral amyloid angiopathy, and Lewy bodies with cognitive impairment in individuals with or without Alzheimer’s disease neuropathology. Sci. Rep. 2020, 10, 14579. [Google Scholar] [CrossRef]
- Kisler, K.; Nelson, A.R.; Rege, S.V.; Ramanathan, A.; Wang, Y.; Ahuja, A.; Lazic, D.; Tsai, P.S.; Zhao, Z.; Zhou, Y.; et al. Pericyte degeneration leads to neurovascular uncoupling and limits oxygen supply to brain. Nat. Neurosci. 2017, 20, 406–416. [Google Scholar] [CrossRef] [Green Version]
- Berthiaume, A.A.; Grant, R.I.; McDowell, K.P.; Underly, R.G.; Hartmann, D.A.; Levy, M.; Bhat, N.R.; Shih, A.Y. Dynamic Remodeling of Pericytes In Vivo Maintains Capillary Coverage in the Adult Mouse Brain. Cell Rep. 2018, 22, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Giannoni, P.; Arango-Lievano, M.; Neves, I.D.; Rousset, M.C.; Baranger, K.; Rivera, S.; Jeanneteau, F.; Claeysen, S.; Marchi, N. Cerebrovascular pathology during the progression of experimental Alzheimer’s disease. Neurobiol. Dis. 2016, 88, 107–117. [Google Scholar] [CrossRef]
- Procter, T.V.; Williams, A.; Montagne, A. Interplay between Brain Pericytes and Endothelial Cells in Dementia. Am. J. Pathol. 2021, 191, 1917–1931. [Google Scholar] [CrossRef] [PubMed]
- Sagare, A.P.; Bell, R.D.; Zhao, Z.; Ma, Q.; Winkler, E.A.; Ramanathan, A.; Zlokovic, B.V. Pericyte loss influences Alzheimer-like neurodegeneration in mice. Nat. Commun. 2013, 4, 2932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Rodriguez, J.J.; Infante-Garcia, C.; Galindo-Gonzalez, L.; Garcia-Molina, Y.; Lechuga-Sancho, A.; Garcia-Alloza, M. Increased Spontaneous Central Bleeding and Cognition Impairment in APP/PS1 Mice with Poorly Controlled Diabetes Mellitus. Mol. Neurobiol. 2016, 53, 2685–2697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nortley, R.; Korte, N.; Izquierdo, P.; Hirunpattarasilp, C.; Mishra, A.; Jaunmuktane, Z.; Kyrargyri, V.; Pfeiffer, T.; Khennouf, L.; Madry, C.; et al. Amyloid beta oligomers constrict human capillaries in Alzheimer’s disease via signaling to pericytes. Science 2019, 365, eaav9518. [Google Scholar] [CrossRef]
- Han, B.H.; Zhou, M.L.; Johnson, A.W.; Singh, I.; Liao, F.; Vellimana, A.K.; Nelson, J.W.; Milner, E.; Cirrito, J.R.; Basak, J.; et al. Contribution of reactive oxygen species to cerebral amyloid angiopathy, vasomotor dysfunction, and microhemorrhage in aged Tg2576 mice. Proc. Natl. Acad. Sci. USA 2015, 112, E881–E890. [Google Scholar] [CrossRef] [Green Version]
- Yamagishi, S.; Takeuchi, M.; Matsui, T.; Nakamura, K.; Imaizumi, T.; Inoue HAngiotensin, I.I. augments advanced glycation end product-induced pericyte apoptosis through RAGE overexpression. FEBS Lett. 2005, 579, 4265–4270. [Google Scholar] [CrossRef] [Green Version]
- Daschil, N.; Kniewallner, K.M.; Obermair, G.J.; Hutter-Paier, B.; Windisch, M.; Marksteiner, J.; Humpel, C. L-type calcium channel blockers and substance P induce angiogenesis of cortical vessels associated with beta-amyloid plaques in an Alzheimer mouse model. Neurobiol. Aging 2015, 36, 1333–1341. [Google Scholar] [CrossRef] [Green Version]
- Miners, J.S.; Palmer, J.C.; Love, S. Pathophysiology of Hypoperfusion of the Precuneus in Early Alzheimer’s Disease. Brain Pathol. 2016, 26, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Montagne, A.; Huuskonen, M.T.; Rajagopal, G.; Sweeney, M.D.; Nation, D.A.; Sepehrband, F.; D’Orazio, L.M.; Harrington, M.G.; Chui, H.C.; Law, M.; et al. Undetectable gadolinium brain retention in individuals with an age-dependent blood-brain barrier breakdown in the hippocampus and mild cognitive impairment. Alzheimers Dement. 2019, 15, 1568–1575. [Google Scholar] [CrossRef]
- Nation, D.A.; Sweeney, M.D.; Montagne, A.; Sagare, A.P.; D’Orazio, L.M.; Pachicano, M.; Sepehrband, F.; Nelson, A.R.; Buennagel, D.P.; Harrington, M.G.; et al. Blood-brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat. Med. 2019, 25, 270–276. [Google Scholar] [CrossRef]
- Montagne, A.; Nation, D.A.; Sagare, A.P.; Barisano, G.; Sweeney, M.D.; Chakhoyan, A.; Pachicano, M.; Joe, E.; Nelson, A.R.; D’Orazio, L.M.; et al. APOE4 leads to blood-brain barrier dysfunction predicting cognitive decline. Nature 2020, 581, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.D.; Sagare, A.P.; Pachicano, M.; Harrington, M.G.; Joe, E.; Chui, H.C.; Schneider, L.S.; Montagne, A.; Ringman, J.M.; Fagan, A.M.; et al. A novel sensitive assay for detection of a biomarker of pericyte injury in cerebrospinal fluid. Alzheimers Dement. 2020, 16, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Mazzei, G.; Ikegami, R.; Abolhassani, N.; Haruyama, N.; Sakumi, K.; Saito, T.; Saido, T.C.; Nakabeppu, Y. A high-fat diet exacerbates the Alzheimer’s disease pathology in the hippocampus of the App(NL-F/NL-F) knock-in mouse model. Aging Cell 2021, 20, e13429. [Google Scholar] [CrossRef]
- Theriault, P.; El Ali, A.; Rivest, S. High fat diet exacerbates Alzheimer’s disease-related pathology in APPswe/PS1 mice. Oncotarget 2016, 7, 67808–67827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tachibana, M.; Yamazaki, Y.; Liu, C.C.; Bu, G.; Kanekiyo, T. Pericyte implantation in the brain enhances cerebral blood flow and reduces amyloid-beta pathology in amyloid model mice. Exp. Neurol. 2018, 300, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A.; Rhea, E.M. The Blood-Brain Barrier, Oxidative Stress, and Insulin Resistance. Antioxidants 2021, 10, 1695. [Google Scholar] [CrossRef]
- Takahashi, H.; Takata, F.; Matsumoto, J.; Machida, T.; Yamauchi, A.; Dohgu, S.; Kataoka, Y. Brain pericyte-derived soluble factors enhance insulin sensitivity in GT1-7 hypothalamic neurons. Biochem. Biophys. Res. Commun. 2015, 457, 532–537. [Google Scholar] [CrossRef]
- Casey, C.S.; Atagi, Y.; Yamazaki, Y.; Shinohara, M.; Tachibana, M.; Fu, Y.; Bu, G.; Kanekiyo, T. Apolipoprotein E Inhibits Cerebrovascular Pericyte Mobility through a RhoA Protein-mediated Pathway. J. Biol. Chem. 2015, 290, 14208–14217. [Google Scholar] [CrossRef] [Green Version]
- Bell, R.D.; Winkler, E.A.; Singh, I.; Sagare, A.P.; Deane, R.; Wu, Z.; Holtzman, D.M.; Betsholtz, C.; Armulik, A.; Sallstrom, J.; et al. Apolipoprotein E controls cerebrovascular integrity via cyclophilin A. Nature 2012, 485, 512–516. [Google Scholar] [CrossRef]
- Desai, M.K.; Mastrangelo, M.A.; Ryan, D.A.; Sudol, K.L.; Narrow, W.C.; Bowers, W.J. Early oligodendrocyte/myelin pathology in Alzheimer’s disease mice constitutes a novel therapeutic target. Am. J. Pathol. 2010, 177, 1422–1435. [Google Scholar] [CrossRef]
- Rivera, F.J.; Hinrichsen, B.; Silva, M.E. Pericytes in Multiple Sclerosis. Adv. Exp. Med. Biol. 2019, 1147, 167–187. [Google Scholar] [CrossRef] [PubMed]
- Salmina, A.B.; Komleva, Y.K.; Lopatina, O.L.; Birbrair, A. Pericytes in Alzheimer’s Disease: Novel Clues to Cerebral Amyloid Angiopathy Pathogenesis. Adv. Exp. Med. Biol. 2019, 1147, 147–166. [Google Scholar] [CrossRef] [PubMed]
- Broman, T. Blood-Brain Barrier Damage in Multiple Sclerosis Supravital Test-Observations. Acta Neurol. Scand. 1964, 40, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Gay, D.; Esiri, M. Blood-brain barrier damage in acute multiple sclerosis plaques. An immunocytological study. Brain 1991, 114, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B.V. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gündüz, T.; Tanyel Kiremitçi, T.; Ulusoy, C.; Kürtüncü, M.; Türkoğlu, R. Cerebrospinal fluid analysis of pericytic mediators in clinically isolated syndrome and multiple sclerosis: A preliminary study. J. Exp. Med. 2018, 81, 18–22. [Google Scholar]
- Absinta, M.; Lassmann, H.; Trapp, B.D. Mechanisms underlying progression in multiple sclerosis. Curr. Opin. Neurol. 2020, 33, 277–285. [Google Scholar] [CrossRef]
- Iacobaeus, E.; Sugars, R.V.; Tornqvist Andren, A.; Alm, J.J.; Qian, H.; Frantzen, J.; Newcombe, J.; Alkass, K.; Druid, H.; Bottai, M.; et al. Dynamic Changes in Brain Mesenchymal Perivascular Cells Associate with Multiple Sclerosis Disease Duration, Active Inflammation, and Demyelination. Stem Cells Transl. Med. 2017, 6, 1840–1851. [Google Scholar] [CrossRef]
- Lassmann, H.; Bradl, M. Multiple sclerosis: Experimental models and reality. Acta Neuropathol. 2017, 133, 223–244. [Google Scholar] [CrossRef] [Green Version]
- Matsushima, G.K.; Morell, P. The neurotoxicant, cuprizone, as a model to study demyelination and remyelination in the central nervous system. Brain Pathol. 2001, 11, 107–116. [Google Scholar] [CrossRef]
- Ferrara, G.; Errede, M.; Girolamo, F.; Morando, S.; Ivaldi, F.; Panini, N.; Bendotti, C.; Perris, R.; Furlan, R.; Virgintino, D.; et al. NG2, a common denominator for neuroinflammation, blood-brain barrier alteration, and oligodendrocyte precursor response in EAE, plays a role in dendritic cell activation. Acta Neuropathol. 2016, 132, 23–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Errede, M.; Girolamo, F.; Ferrara, G.; Strippoli, M.; Morando, S.; Boldrin, V.; Rizzi, M.; Uccelli, A.; Perris, R.; Bendotti, C.; et al. Blood-brain barrier alterations in the cerebral cortex in experimental autoimmune encephalomyelitis. J. Neuropathol. Exp. Neurol. 2012, 71, 840–854. [Google Scholar] [CrossRef] [Green Version]
- Maki, T.; Choi, Y.K.; Miyamoto, N.; Shindo, A.; Liang, A.C.; Ahn, B.J.; Mandeville, E.T.; Kaji, S.; Itoh, K.; Seo, J.H.; et al. A-Kinase Anchor Protein 12 Is Required for Oligodendrocyte Differentiation in Adult White Matter. Stem Cells 2018, 36, 751–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grygorowicz, T.; Dabrowska-Bouta, B.; Struzynska, L. Administration of an antagonist of P2X7 receptor to EAE rats prevents a decrease of expression of claudin-5 in cerebral capillaries. Purinergic Signal. 2018, 14, 385–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushik, D.K.; Bhattacharya, A.; Lozinski, B.M.; Wee Yong, V. Pericytes as mediators of infiltration of macrophages in multiple sclerosis. J. Neuroinflamm. 2021, 18, 301. [Google Scholar] [CrossRef]
- Guijarro-Munoz, I.; Compte, M.; Alvarez-Cienfuegos, A.; Alvarez-Vallina, L.; Sanz, L. Lipopolysaccharide activates Toll-like receptor 4 (TLR4)-mediated NF-kappaB signaling pathway and proinflammatory response in human pericytes. J. Biol. Chem. 2014, 289, 2457–2468. [Google Scholar] [CrossRef] [Green Version]
- Jansson, D.; Rustenhoven, J.; Feng, S.; Hurley, D.; Oldfield, R.L.; Bergin, P.S.; Mee, E.W.; Faull, R.L.; Dragunow, M. Erratum To: A role for human brain pericytes in neuroinflammation. J. Neuroinflamm. 2014, 11, 104. [Google Scholar] [CrossRef] [Green Version]
- Pieper, C.; Marek, J.J.; Unterberg, M.; Schwerdtle, T.; Galla, H.J. Brain capillary pericytes contribute to the immune defense in response to cytokines or LPS in vitro. Brain Res. 2014, 1550, 1–8. [Google Scholar] [CrossRef]
- Ulusoy, C.; Sekerdag, E.; Yilmaz, V.; Yilmaz, A.B.; Atak, D.; Vural, A.; Kucukali, C.I.; Karaaslan, Z.; Kurtuncu, M.; Turkoglu, R.; et al. Impact of Autoimmune Demyelinating Brain Disease Sera on Pericyte Survival. Noro Psikiyatr Ars. 2021, 58, 83–86. [Google Scholar] [CrossRef]
- Kovac, A.; Erickson, M.A.; Banks, W.A. Brain microvascular pericytes are immunoactive in culture: Cytokine, chemokine, nitric oxide, and LRP-1 expression in response to lipopolysaccharide. J. Neuroinflamm. 2011, 8, 139. [Google Scholar] [CrossRef] [Green Version]
- Takata, F.; Dohgu, S.; Matsumoto, J.; Machida, T.; Kaneshima, S.; Matsuo, M.; Sakaguchi, S.; Takeshige, Y.; Yamauchi, A.; Kataoka, Y. Metformin Induces up-Regulation of Blood-Brain Barrier Functions by Activating Amp-Activated Protein Kinase in Rat Brain Microvascular Endothelial Cells. Biochem. Biophys. Res. Commun. 2013, 433, 586–590. [Google Scholar] [CrossRef] [PubMed]
- Keough, M.B.; Rogers, J.A.; Zhang, P.; Jensen, S.K.; Stephenson, E.L.; Chen, T.; Hurlbert, M.G.; Lau, L.W.; Rawji, K.S.; Plemel, J.R.; et al. An inhibitor of chondroitin sulfate proteoglycan synthesis promotes central nervous system remyelination. Nat. Commun. 2016, 7, 11312. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, E.L.; Mishra, M.K.; Moussienko, D.; Laflamme, N.; Rivest, S.; Ling, C.C.; Yong, V.W. Chondroitin sulfate proteoglycans as novel drivers of leucocyte infiltration in multiple sclerosis. Brain 2018, 141, 1094–1110. [Google Scholar] [CrossRef] [PubMed]
- Holley, J.E.; Newcombe, J.; Whatmore, J.L.; Gutowski, N.J. Increased blood vessel density and endothelial cell proliferation in multiple sclerosis cerebral white matter. Neurosci. Lett. 2010, 470, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Ludwin, S. Vascular proliferation and angiogenesis in MS: Clinical and pathogenic implications. J. Neuropathol. Exp. Neurol. 2001, 60, 505. [Google Scholar]
- Papadaki, E.Z.; Simos, P.G.; Mastorodemos, V.C.; Panou, T.; Maris, T.G.; Karantanas, A.H.; Plaitakis, A. Regional MRI perfusion measures predict motor/executive function in patients with clinically isolated syndrome. Behav. Neurol. 2014, 2014, 252419. [Google Scholar] [CrossRef] [PubMed]
- Boroujerdi, A.; Welser-Alves, J.V.; Milner, R. Extensive vascular remodeling in the spinal cord of pre-symptomatic experimental autoimmune encephalomyelitis mice; increased vessel expression of fibronectin and the alpha5beta1 integrin. Exp. Neurol. 2013, 250, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Girolamo, F.; Coppola, C.; Ribatti, D.; Trojano, M. Angiogenesis in multiple sclerosis and experimental autoimmune encephalomyelitis. Acta Neuropathol. Commun. 2014, 2, 84. [Google Scholar] [CrossRef]
- Kirk, S.L.; Karlik, S.J. VEGF and vascular changes in chronic neuroinflammation. J. Autoimmun. 2003, 21, 353–363. [Google Scholar] [CrossRef]
- Macmillan, C.J.; Starkey, R.J.; Easton, A.S. Angiogenesis is regulated by angiopoietins during experimental autoimmune encephalomyelitis and is indirectly related to vascular permeability. J. Neuropathol. Exp. Neurol. 2011, 70, 1107–1123. [Google Scholar] [CrossRef] [Green Version]
- Piraino, P.S.; Yednock, T.A.; Messersmith, E.K.; Pleiss, M.A.; Freedman, S.B.; Hammond, R.R.; Karlik, S.J. Spontaneous remyelination following prolonged inhibition of alpha4 integrin in chronic EAE. J. Neuroimmunol. 2005, 167, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Roscoe, W.A.; Welsh, M.E.; Carter, D.E.; Karlik, S.J. VEGF and angiogenesis in acute and chronic MOG((35-55)) peptide induced EAE. J. Neuroimmunol. 2009, 209, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Seabrook, T.J.; Littlewood-Evans, A.; Brinkmann, V.; Pollinger, B.; Schnell, C.; Hiestand, P.C. Angiogenesis is present in experimental autoimmune encephalomyelitis and pro-angiogenic factors are increased in multiple sclerosis lesions. J. Neuroinflamm. 2010, 7, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobel, R.A.; Blanchette, B.W.; Bhan, A.K.; Colvin, R.B. The immunopathology of experimental allergic encephalomyelitis. II. Endothelial cell Ia increases prior to inflammatory cell infiltration. J. Immunol. 1984, 132, 2402–2407. [Google Scholar]
- Azevedo, P.O.; Sena, I.F.G.; Andreotti, J.P.; Carvalho-Tavares, J.; Alves-Filho, J.C.; Cunha, T.M.; Cunha, F.Q.; Mintz, A.; Birbrair, A. Pericytes modulate myelination in the central nervous system. J. Cell. Physiol. 2018, 233, 5523–5529. [Google Scholar] [CrossRef] [PubMed]
- Iadecola, C. Neurovascular regulation in the normal brain and in Alzheimer’s disease. Nat. Rev. Neurosci. 2004, 5, 347–360. [Google Scholar] [CrossRef]
- Abbott, N.J.; Ronnback, L.; Hansson, E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef]
- Hartmann, D.A.; Berthiaume, A.A.; Grant, R.I.; Harrill, S.A.; Koski, T.; Tieu, T.; McDowell, K.P.; Faino, A.V.; Kelly, A.L.; Shih, A.Y. Brain capillary pericytes exert a substantial but slow influence on blood flow. Nat. Neurosci. 2021, 24, 633–645. [Google Scholar] [CrossRef]
- Proebstl, D.; Voisin, M.B.; Woodfin, A.; Whiteford, J.; D’Acquisto, F.; Jones, G.E.; Rowe, D.; Nourshargh, S. Pericytes support neutrophil subendothelial cell crawling and breaching of venular walls in vivo. J. Exp. Med. 2012, 209, 1219–1234. [Google Scholar] [CrossRef] [Green Version]
- Dohgu, S.; Banks, W.A. Brain pericytes increase the lipopolysaccharide-enhanced transcytosis of HIV-1 free virus across the in vitro blood-brain barrier: Evidence for cytokine-mediated pericyte-endothelial cell crosstalk. Fluids Barriers CNS 2013, 10, 23. [Google Scholar] [CrossRef] [Green Version]
- Stark, K.; Eckart, A.; Haidari, S.; Tirniceriu, A.; Lorenz, M.; von Bruhl, M.L.; Gartner, F.; Khandoga, A.G.; Legate, K.R.; Pless, R.; et al. Capillary and arteriolar pericytes attract innate leukocytes exiting through venules and ‘instruct’ them with pattern-recognition and motility programs. Nat. Immunol. 2013, 14, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Leick, M.; Azcutia, V.; Newton, G.; Luscinskas, F.W. Leukocyte recruitment in inflammation: Basic concepts and new mechanistic insights based on new models and microscopic imaging technologies. Cell Tissue Res. 2014, 355, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Tietz, S.M.; Engelhardt, B. Visualizing Impairment of the Endothelial and Glial Barriers of the Neurovascular Unit during Experimental Autoimmune Encephalomyelitis In Vivo. J. Vis. Exp. 2019, 145, e59249. [Google Scholar] [CrossRef] [PubMed]
- Dalkara, T. Pericytes: A Novel Target to Improve Success of Recanalization Therapies. Stroke 2019, 50, 2985–2991. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.I.; Hartmann, D.A.; Underly, R.G.; Berthiaume, A.A.; Bhat, N.R.; Shih, A.Y. Organizational hierarchy and structural diversity of microvascular pericytes in adult mouse cortex. J. Cereb. Blood Flow Metab. 2019, 39, 411–425. [Google Scholar] [CrossRef] [Green Version]
- Dalkara, T.; Alarcon-Martinez, L.; Yemisci, M. Pericytes in Ischemic Stroke. Adv. Exp. Med. Biol. 2019, 1147, 189–213. [Google Scholar] [CrossRef]
- Herman, I.M.; D’Amore, P.A. Microvascular pericytes contain muscle and nonmuscle actins. J. Cell Biol. 1985, 101, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Joyce, N.C.; Haire, M.F.; Palade, G.E. Contractile proteins in pericytes. II. Immunocytochemical evidence for the presence of two isomyosins in graded concentrations. J. Cell Biol. 1985, 100, 1387–1395. [Google Scholar] [CrossRef] [Green Version]
- Bandopadhyay, R.; Orte, C.; Lawrenson, J.G.; Reid, A.R.; De Silva, S.; Allt, G. Contractile proteins in pericytes at the blood-brain and blood-retinal barriers. J. Neurocytol. 2001, 30, 35–44. [Google Scholar] [CrossRef]
- Kelley, C.; D’Amore, P.; Hechtman, H.B.; Shepro, D. Microvascular pericyte contractility in vitro: Comparison with other cells of the vascular wall. J. Cell Biol. 1987, 104, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Dodge, A.B.; Hechtman, H.B.; Shepro, D. Microvascular endothelial-derived autacoids regulate pericyte contractility. Cell Motil. Cytoskelet. 1991, 18, 180–188. [Google Scholar] [CrossRef]
- Puro, D.G. Physiology and pathobiology of the pericyte-containing retinal microvasculature: New developments. Microcirculation 2007, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kotecki, M.; Zeiger, A.S.; Van Vliet, K.J.; Herman, I.M. Calpain- and talin-dependent control of microvascular pericyte contractility and cellular stiffness. Microvasc. Res. 2010, 80, 339–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Klett, F.; Priller, J. Diverse functions of pericytes in cerebral blood flow regulation and ischemia. J. Cereb. Blood Flow Metab. 2015, 35, 883–887. [Google Scholar] [CrossRef] [Green Version]
- Biesecker, K.R.; Srienc, A.I.; Shimoda, A.M.; Agarwal, A.; Bergles, D.E.; Kofuji, P.; Newman, E.A. Glial Cell Calcium Signaling Mediates Capillary Regulation of Blood Flow in the Retina. J. Neurosci. 2016, 36, 9435–9445. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Reynolds, J.P.; Chen, Y.; Gourine, A.V.; Rusakov, D.A.; Attwell, D. Astrocytes mediate neurovascular signaling to capillary pericytes but not to arterioles. Nat. Neurosci. 2016, 19, 1619–1627. [Google Scholar] [CrossRef] [Green Version]
- Krueger, M.; Bechmann, I. CNS pericytes: Concepts, misconceptions, and a way out. Glia 2010, 58, 1–10. [Google Scholar] [CrossRef]
- Hill, R.A.; Tong, L.; Yuan, P.; Murikinati, S.; Gupta, S.; Grutzendler, J. Regional Blood Flow in the Normal and Ischemic Brain Is Controlled by Arteriolar Smooth Muscle Cell Contractility and Not by Capillary Pericytes. Neuron 2015, 87, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Attwell, D.; Buchan, A.M.; Charpak, S.; Lauritzen, M.; Macvicar, B.A.; Newman, E.A. Glial and neuronal control of brain blood flow. Nature 2010, 468, 232–243. [Google Scholar] [CrossRef] [Green Version]
- Serebryakov, V.; Zakharenko, S.; Snetkov, V.; Takeda, K. Effects of prostaglandins E1 and E2 on cultured smooth muscle cells and strips of rat aorta. Prostaglandins 1994, 47, 353–365. [Google Scholar] [CrossRef]
- Crawford, C.; Kennedy-Lydon, T.; Sprott, C.; Desai, T.; Sawbridge, L.; Munday, J.; Unwin, R.J.; Wildman, S.S.; Peppiatt-Wildman, C.M. An intact kidney slice model to investigate vasa recta properties and function in situ. Nephron. Physiol. 2012, 120, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Gluck, C.; Ferrari, K.D.; Binini, N.; Keller, A.; Saab, A.S.; Stobart, J.L.; Weber, B. Distinct signatures of calcium activity in brain mural cells. eLife 2021, 10, e70591. [Google Scholar] [CrossRef] [PubMed]
- El Ali, A.; Theriault, P.; Rivest, S. The role of pericytes in neurovascular unit remodeling in brain disorders. Int. J. Mol. Sci. 2014, 15, 6453–6474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torok, O.; Schreiner, B.; Schaffenrath, J.; Tsai, H.C.; Maheshwari, U.; Stifter, S.A.; Welsh, C.; Amorim, A.; Sridhar, S.; Utz, S.G.; et al. Pericytes regulate vascular immune homeostasis in the CNS. Proc. Natl. Acad. Sci. USA 2021, 118, e2016587118. [Google Scholar] [CrossRef]
- Yemisci, M.; Gursoy-Ozdemir, Y.; Vural, A.; Can, A.; Topalkara, K.; Dalkara, T. Pericyte contraction induced by oxidative-nitrative stress impairs capillary reflow despite successful opening of an occluded cerebral artery. Nat. Med. 2009, 15, 1031–1037. [Google Scholar] [CrossRef]
- Kamouchi, M.; Kitazono, T.; Ago, T.; Wakisaka, M.; Ooboshi, H.; Ibayashi, S.; Iida, M. Calcium influx pathways in rat CNS pericytes. Brain Res. Mol. Brain Res. 2004, 126, 114–120. [Google Scholar] [CrossRef]
- Nakamura, K.; Kamouchi, M.; Kitazono, T.; Kuroda, J.; Shono, Y.; Hagiwara, N.; Ago, T.; Ooboshi, H.; Ibayashi, S.; Iida, M. Amiloride inhibits hydrogen peroxide-induced Ca2+ responses in human CNS pericytes. Microvasc. Res. 2009, 77, 327–334. [Google Scholar] [CrossRef]
- Gursoy-Ozdemir, Y.; Can, A.; Dalkara, T. Reperfusion-induced oxidative/nitrative injury to neurovascular unit after focal cerebral ischemia. Stroke 2004, 35, 1449–1453. [Google Scholar] [CrossRef] [Green Version]
- Gursoy-Ozdemir, Y.; Yemisci, M.; Dalkara, T. Microvascular protection is essential for successful neuroprotection in stroke. J. Neurochem. 2012, 123 (Suppl. 2), 2–11. [Google Scholar] [CrossRef] [Green Version]
- Manea, A.; Raicu, M.; Simionescu, M. Expression of functionally phagocyte-type NAD(P)H oxidase in pericytes: Effect of angiotensin II and high glucose. Biol. Cell 2005, 97, 723–734. [Google Scholar] [CrossRef]
- Kuroda, J.; Ago, T.; Nishimura, A.; Nakamura, K.; Matsuo, R.; Wakisaka, Y.; Kamouchi, M.; Kitazono, T. Nox4 is a major source of superoxide production in human brain pericytes. J. Vasc. Res. 2014, 51, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, A.; Ago, T.; Kuroda, J.; Arimura, K.; Tachibana, M.; Nakamura, K.; Wakisaka, Y.; Sadoshima, J.; Iihara, K.; Kitazono, T. Detrimental role of pericyte Nox4 in the acute phase of brain ischemia. J. Cereb. Blood Flow Metab. 2016, 36, 1143–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallet, P.; Charnay, Y.; Steger, K.; Ogier-Denis, E.; Kovari, E.; Herrmann, F.; Michel, J.P.; Szanto, I. Neuronal expression of the NADPH oxidase NOX4, and its regulation in mouse experimental brain ischemia. Neuroscience 2005, 132, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.H. Role of oxidants in ischemic brain damage. Stroke 1996, 27, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Ames, A., 3rd; Wright, R.L.; Kowada, M.; Thurston, J.M.; Majno, G. Cerebral ischemia. II. The no-reflow phenomenon. Am. J. Pathol. 1968, 52, 437–453. [Google Scholar] [PubMed]
- Crowell, R.M.; Olsson, Y. Impaired microvascular filling after focal cerebral ischemia in the monkey: Modification by treatment. Neurology 1972, 22, 500–504. [Google Scholar] [CrossRef] [PubMed]
- del Zoppo, G.J. Virchow’s triad: The vascular basis of cerebral injury. Rev. Neurol. Dis. 2008, 5 (Suppl. 1), S12–S21. [Google Scholar]
- Dziennis, S.; Qin, J.; Shi, L.; Wang, R.K. Macro-to-micro cortical vascular imaging underlies regional differences in ischemic brain. Sci. Rep. 2015, 5, 10051. [Google Scholar] [CrossRef] [Green Version]
- Little, J.R.; Kerr, F.W.; Sundt, T.M., Jr. Microcirculatory Obstruction in Focal Cerebral Ischemia: An Electron Microscopic Investigation in Monkeys. Stroke 1976, 7, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Belayev, L.; Pinard, E.; Nallet, H.; Seylaz, J.; Liu, Y.; Riyamongkol, P.; Zhao, W.; Busto, R.; Ginsberg, M.D. Albumin therapy of transient focal cerebral ischemia: In vivo analysis of dynamic microvascular responses. Stroke 2002, 33, 1077–1084. [Google Scholar] [CrossRef] [Green Version]
- Hallenbeck, J.M.; Dutka, A.J.; Tanishima, T.; Kochanek, P.M.; Kumaroo, K.K.; Thompson, C.B.; Obrenovitch, T.P.; Contreras, T.J. Polymorphonuclear leukocyte accumulation in brain regions with low blood flow during the early postischemic period. Stroke 1986, 17, 246–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, E.; del Zoppo, G.J.; Chambers, J.D.; Copeland, B.R.; Arfors, K.E. Inhibition of polymorphonuclear leukocyte adherence suppresses no-reflow after focal cerebral ischemia in baboons. Stroke 1992, 23, 712–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, M.; Vowinkel, T.; Stokes, K.Y.; Arumugam, T.V.; Yilmaz, G.; Nanda, A.; Granger, D.N. CD40/CD40 ligand signaling in mouse cerebral microvasculature after focal ischemia/reperfusion. Circulation 2005, 111, 1690–1696. [Google Scholar] [CrossRef] [PubMed]
- Sienel, R.I.; Kataoka, H.; Kim, S.W.; Seker, F.B.; Plesnila, N. Adhesion of Leukocytes to Cerebral Venules Precedes Neuronal Cell Death and Is Sufficient to Trigger Tissue Damage After Cerebral Ischemia. Front. Neurol. 2021, 12, 807658. [Google Scholar] [CrossRef] [PubMed]
- South, K.; Saleh, O.; Lemarchand, E.; Coutts, G.; Smith, C.J.; Schiessl, I.; Allan, S.M. Robust thrombolytic and anti-inflammatory action of a constitutively active ADAMTS13 variant in murine stroke models. Blood 2022, 139, 1575–1587. [Google Scholar] [CrossRef]
- Powers, W.J.; Rabinstein, A.A.; Ackerson, T.; Adeoye, O.M.; Bambakidis, N.C.; Becker, K.; Biller, J.; Brown, M.; Demaerschalk, B.M.; Hoh, B.; et al. Guidelines for the Early Management of Patients with Acute Ischemic Stroke: A Guideline for Healthcare Professionals from the American Heart Association/American Stroke Association. Stroke 2018, 49, e46–e110. [Google Scholar] [CrossRef]
- Tseng, Y.J.; Hu, R.F.; Lee, S.T.; Lin, Y.L.; Hsu, C.L.; Lin, S.W.; Liou, C.W.; Lee, J.D.; Peng, T.I.; Lee, T.H. Risk Factors Associated with Outcomes of Recombinant Tissue Plasminogen Activator Therapy in Patients with Acute Ischemic Stroke. Int. J. Environ. Res. Public Health 2020, 17, 618. [Google Scholar] [CrossRef] [Green Version]
- Robichon, E.; Maier, B.; Mazighi, M. Endovascular therapy for acute ischemic stroke: The importance of blood pressure control, sedation modality and anti-thrombotic management to improve functional outcomes. Rev. Neurol. 2022, 178, 175–184. [Google Scholar] [CrossRef]
- Gaudin, A.; Yemisci, M.; Eroglu, H.; Lepetre-Mouelhi, S.; Turkoglu, O.F.; Donmez-Demir, B.; Caban, S.; Sargon, M.F.; Garcia-Argote, S.; Pieters, G.; et al. Squalenoyl adenosine nanoparticles provide neuroprotection after stroke and spinal cord injury. Nat. Nanotechnol. 2014, 9, 1054–1062. [Google Scholar] [CrossRef]
- Taskiran-Sag, A.; Yemisci, M.; Gursoy-Ozdemir, Y.; Erdener, S.E.; Karatas, H.; Yuce, D.; Dalkara, T. Improving Microcirculatory Reperfusion Reduces Parenchymal Oxygen Radical Formation and Provides Neuroprotection. Stroke 2018, 49, 1267–1275. [Google Scholar] [CrossRef]
- Zheng, Y.Q.; Liu, J.X.; Wang, J.N.; Xu, L. Effects of crocin on reperfusion-induced oxidative/nitrative injury to cerebral microvessels after global cerebral ischemia. Brain Res. 2006, 1138, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Dalkara, T.; Arsava, E.M. Can restoring incomplete microcirculatory reperfusion improve stroke outcome after thrombolysis? J. Cereb. Blood Flow Metab. 2012, 32, 2091–2099. [Google Scholar] [CrossRef] [PubMed]
- Arsava, E.M.; Arat, A.; Topcuoglu, M.A.; Peker, A.; Yemisci, M.; Dalkara, T. Angiographic Microcirculatory Obstructions Distal to Occlusion Signify Poor Outcome after Endovascular Treatment for Acute Ischemic Stroke. Transl. Stroke Res. 2018, 9, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Simard, J.M.; Kent, T.A.; Chen, M.; Tarasov, K.V.; Gerzanich, V. Brain oedema in focal ischaemia: Molecular pathophysiology and theoretical implications. Lancet Neurol. 2007, 6, 258–268. [Google Scholar] [CrossRef] [Green Version]
- Underly, R.G.; Levy, M.; Hartmann, D.A.; Grant, R.I.; Watson, A.N.; Shih, A.Y. Pericytes as Inducers of Rapid, Matrix Metalloproteinase-9-Dependent Capillary Damage during Ischemia. J. Neurosci. 2017, 37, 129–140. [Google Scholar] [CrossRef]
- Bhatia, R.; Warrier, A.R.; Sreenivas, V.; Bali, P.; Sisodia, P.; Gupta, A.; Singh, N.; Padma Srivastava, M.V.; Prasad, K. Role of Blood Biomarkers in Differentiating Ischemic Stroke and Intracerebral Hemorrhage. Neurol. India 2020, 68, 824–829. [Google Scholar] [CrossRef]
- Gaude, E.; Nogueira, B.; Ladreda Mochales, M.; Graham, S.; Smith, S.; Shaw, L.; Graziadio, S.; Ladreda Mochales, G.; Sloan, P.; Bernstock, J.D.; et al. A Novel Combination of Blood Biomarkers and Clinical Stroke Scales Facilitates Detection of Large Vessel Occlusion Ischemic Strokes. Diagnostics 2021, 11, 1137. [Google Scholar] [CrossRef]
- Foschi, M.; Padroni, M.; Abu-Rumeileh, S.; Abdelhak, A.; Russo, M.; D’Anna, L.; Guarino, M. Diagnostic and Prognostic Blood Biomarkers in Transient Ischemic Attack and Minor Ischemic Stroke: An Up-To-Date Narrative Review. J. Stroke Cerebrovasc. Dis. 2022, 31, 106292. [Google Scholar] [CrossRef]
- Hirtz, C.; Chevalier, F.; Centeno, D.; Egea, J.C.; Rossignol, M.; Sommerer, N.; de Periere, D. Complexity of the human whole saliva proteome. J. Physiol. Biochem. 2005, 61, 469–480. [Google Scholar] [CrossRef] [Green Version]
- Al-Rawi, N.H.; Atiyah, K.M. Salivary neuron specific enolase: An indicator for neuronal damage in patients with ischemic stroke and stroke-prone patients. Clin. Chem. Lab. Med. 2009, 47, 1519–1524. [Google Scholar] [CrossRef] [Green Version]
- Di Pietro, V.; Porto, E.; Ragusa, M.; Barbagallo, C.; Davies, D.; Forcione, M.; Logan, A.; Di Pietro, C.; Purrello, M.; Grey, M.; et al. Salivary MicroRNAs: Diagnostic Markers of Mild Traumatic Brain Injury in Contact-Sport. Front. Mol. Neurosci. 2018, 11, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczor-Urbanowicz, K.E.; Kim, Y.; Li, F.; Galeev, T.; Kitchen, R.R.; Gerstein, M.; Koyano, K.; Jeong, S.H.; Wang, X.; Elashoff, D.; et al. Novel approaches for bioinformatic analysis of salivary RNA sequencing data for development. Bioinformatics 2018, 34, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindblad, C.; Nelson, D.W.; Zeiler, F.A.; Ercole, A.; Ghatan, P.H.; von Horn, H.; Risling, M.; Svensson, M.; Agoston, D.V.; Bellander, B.M.; et al. Influence of Blood-Brain Barrier Integrity on Brain Protein Biomarker Clearance in Severe Traumatic Brain Injury: A Longitudinal Prospective Study. J. Neurotrauma 2020, 37, 1381–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renner, O.; Tsimpas, A.; Kostin, S.; Valable, S.; Petit, E.; Schaper, W.; Marti, H.H. Time- and cell type-specific induction of platelet-derived growth factor receptor-beta during cerebral ischemia. Brain Res. Mol. Brain Res. 2003, 113, 44–51. [Google Scholar] [CrossRef]
- Hutanu, A.; Iancu, M.; Maier, S.; Balasa, R.; Dobreanu, M. Plasma Biomarkers as Potential Predictors of Functional Dependence in Daily Life Activities after Ischemic Stroke: A Single Center Study. Ann. Indian Acad. Neurol. 2020, 23, 496–503. [Google Scholar] [CrossRef]
- Kim, K.J.; Jeong, S.W.; Ryu, W.S.; Kim, D.E.; Saver, J.L.; Kim, J.S.; Kwon, S.U.; Group, T.-S. Platelet-Derived Growth Factor Is Associated with Progression of Symptomatic Intracranial Atherosclerotic Stenosis. J. Clin. Neurol. 2021, 17, 70–76. [Google Scholar] [CrossRef]
- Cui, Y.; Zhao, Y.; Chen, S.Y.; Sheng, B.Y.; Wang, L.H.; Meng, W.H.; Chen, H.S. Association of Serum Biomarkers with Post-Thrombolytic Symptomatic Intracranial Hemorrhage in Stroke: A Comprehensive Protein Microarray Analysis from INTRECIS Study. Front. Neurol. 2022, 13, 751912. [Google Scholar] [CrossRef]
- Gerhardt, H.; Betsholtz, C. Endothelial-pericyte interactions in angiogenesis. Cell Tissue Res. 2003, 314, 15–23. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Z.G.; Chopp, M. The neurovascular unit and combination treatment strategies for stroke. Trends Pharmacol. Sci. 2012, 33, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.; Chang, M.S.; Koh, S.H.; Choi, Y.K. Repair Mechanisms of the Neurovascular Unit after Ischemic Stroke with a Focus on VEGF. Int. J. Mol. Sci. 2021, 22, 8543. [Google Scholar] [CrossRef]
- Kamouchi, M.; Ago, T.; Kitazono, T. Brain pericytes: Emerging concepts and functional roles in brain homeostasis. Cell Mol. Neurobiol. 2011, 31, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Machida, T.; Dohgu, S.; Takata, F.; Matsumoto, J.; Kimura, I.; Koga, M.; Nakamoto, K.; Yamauchi, A.; Kataoka, Y. Role of thrombin-PAR1-PKCtheta/delta axis in brain pericytes in thrombin-induced MMP-9 production and blood-brain barrier dysfunction in vitro. Neuroscience 2017, 350, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Dias, D.O.; Kalkitsas, J.; Kelahmetoglu, Y.; Estrada, C.P.; Tatarishvili, J.; Holl, D.; Jansson, L.; Banitalebi, S.; Amiry-Moghaddam, M.; Ernst, A.; et al. Pericyte-derived fibrotic scarring is conserved across diverse central nervous system lesions. Nat. Commun. 2021, 12, 5501. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Liu, H.; Zhao, J.; Chen, L.Y.; Chen, J.; Lu, Z.; Hu, X. Pericytes in Brain Injury and Repair After Ischemic Stroke. Transl. Stroke Res. 2017, 8, 107–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, M.; Ninomiya, I.; Kanazawa, M. Angiogenesis and neuronal remodeling after ischemic stroke. Neural Regen. Res. 2020, 15, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Prakash, R.; Li, W.; Qu, Z.; Johnson, M.A.; Fagan, S.C.; Ergul, A. Vascularization pattern after ischemic stroke is different in control versus diabetic rats: Relevance to stroke recovery. Stroke 2013, 44, 2875–2882. [Google Scholar] [CrossRef]
- Ergul, A.; Abdelsaid, M.; Fouda, A.Y.; Fagan, S.C. Cerebral neovascularization in diabetes: Implications for stroke recovery and beyond. J. Cereb. Blood Flow Metab. 2014, 34, 553–563. [Google Scholar] [CrossRef] [Green Version]
- Ergul, A.; Valenzuela, J.P.; Fouda, A.Y.; Fagan, S.C. Cellular connections, microenvironment and brain angiogenesis in diabetes: Lost communication signals in the post-stroke period. Brain Res. 2015, 1623, 81–96. [Google Scholar] [CrossRef] [Green Version]
- Mishiro, K.; Imai, T.; Sugitani, S.; Kitashoji, A.; Suzuki, Y.; Takagi, T.; Chen, H.; Oumi, Y.; Tsuruma, K.; Shimazawa, M.; et al. Diabetes mellitus aggravates hemorrhagic transformation after ischemic stroke via mitochondrial defects leading to endothelial apoptosis. PLoS ONE 2014, 9, e103818. [Google Scholar] [CrossRef]
- Prakash, R.; Somanath, P.R.; El-Remessy, A.B.; Kelly-Cobbs, A.; Stern, J.E.; Dore-Duffy, P.; Johnson, M.; Fagan, S.C.; Ergul, A. Enhanced cerebral but not peripheral angiogenesis in the Goto-Kakizaki model of type 2 diabetes involves VEGF and peroxynitrite signaling. Diabetes 2012, 61, 1533–1542. [Google Scholar] [CrossRef] [Green Version]
- Ergul, A.; Elgebaly, M.M.; Middlemore, M.L.; Li, W.; Elewa, H.; Switzer, J.A.; Hall, C.; Kozak, A.; Fagan, S.C. Increased hemorrhagic transformation and altered infarct size and localization after experimental stroke in a rat model type 2 diabetes. BMC Neurol. 2007, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, C.; Chen, S.; Ruan, Y.; Liu, Y.; Cheng, H.; Zeng, Y.; Chen, Y.; Cheng, Q.; Huang, G.; He, W.; et al. The Stress Hyperglycemia Ratio is Associated with Hemorrhagic Transformation in Patients with Acute Ischemic Stroke. Clin. Interv. Aging 2021, 16, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Masawa, N.; Sakata, N.; Takatama, M. Pathologic evidence of microvascular rarefaction in the brain of renal hypertensive rats. J. Stroke Cerebrovasc. Dis. 2003, 12, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Ostergaard, L.; Engedal, T.S.; Moreton, F.; Hansen, M.B.; Wardlaw, J.M.; Dalkara, T.; Markus, H.S.; Muir, K.W. Cerebral small vessel disease: Capillary pathways to stroke and cognitive decline. J. Cereb. Blood Flow Metab. 2016, 36, 302–325. [Google Scholar] [CrossRef]
- He, L.; Mae, M.A.; Muhl, L.; Sun, Y.; Pietila, R.; Nahar, K.; Liébanas, E.V.; Fagerlund, M.J.; Oldner, A.; Liu, J.; et al. Pericyte-specific vascular expression of SARS-CoV-2 receptor ACE2—Implications for microvascular inflammation and hypercoagulopathy in COVID-19. bioRxiv 2020. [Google Scholar] [CrossRef]
- Bocci, M.; Oudenaarden, C.; Saenz-Sarda, X.; Simren, J.; Eden, A.; Sjolund, J.; Moller, C.; Gisslen, M.; Zetterberg, H.; Englund, E.; et al. Infection of Brain Pericytes Underlying Neuropathology of COVID-19 Patients. Int. J. Mol. Sci. 2021, 22, 11622. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.; Jin, H.; Wang, M.; Hu, Y.; Chen, S.; He, Q.; Chang, J.; Hong, C.; Zhou, Y.; Wang, D.; et al. Neurologic Manifestations of Hospitalized Patients with Coronavirus Disease 2019 in Wuhan, China. JAMA Neurol. 2020, 77, 683–690. [Google Scholar] [CrossRef] [Green Version]
- Klok, F.A.; Kruip, M.; van der Meer, N.J.M.; Arbous, M.S.; Gommers, D.; Kant, K.M.; Kaptein, F.H.J.; van Paassen, J.; Stals, M.A.M.; Huisman, M.V.; et al. Confirmation of the high cumulative incidence of thrombotic complications in critically ill ICU patients with COVID-19, An updated analysis. Thromb. Res. 2020, 191, 148–150. [Google Scholar] [CrossRef]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, V.S.; Zetter, M.A.; Guerra, E.C.; Hernandez-Araiza, I.; Karuzin, N.; Hernandez-Perez, O.R.; Eiden, L.E.; Zhang, L. ACE2 expression in rat brain: Implications for COVID-19 associated neurological manifestations. Exp. Neurol. 2021, 345, 113837. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sievert, D.; Clark, A.E.; Lee, S.; Federman, H.; Gastfriend, B.D.; Shusta, E.V.; Palecek, S.P.; Carlin, A.F.; Gleeson, J.G. A human three-dimensional neural-perivascular ‘assembloid’ promotes astrocytic development and enables modeling of SARS-CoV-2 neuropathology. Nat. Med. 2021, 27, 1600–1606. [Google Scholar] [CrossRef] [PubMed]
- Matschke, J.; Lutgehetmann, M.; Hagel, C.; Sperhake, J.P.; Schroder, A.S.; Edler, C.; Mushumba, H.; Fitzek, A.; Allweiss, L.; Dandri, M.; et al. Neuropathology of patients with COVID-19 in Germany: A post-mortem case series. Lancet Neurol. 2020, 19, 919–929. [Google Scholar] [CrossRef]
- Daisley, H., Jr.; Rampersad, A.; Daisley, M.; Ramdin, A.; Acco, O.; Narinesingh, F.; Humphrey, O. COVID-19, a closer look at the pathology in two autopsied cases. Is the pericyte at the center of the pathological process in COVID-19? Autops. Case Rep. 2021, 11, e2021262. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Perl, D.P.; Nair, G.; Li, W.; Maric, D.; Murray, H.; Dodd, S.J.; Koretsky, A.P.; Watts, J.A.; Cheung, V.; et al. Microvascular Injury in the Brains of Patients with COVID-19. N. Engl. J. Med. 2021, 384, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Thakur, K.T.; Miller, E.H.; Glendinning, M.D.; Al-Dalahmah, O.; Banu, M.A.; Boehme, A.K.; Boubour, A.L.; Bruce, S.S.; Chong, A.M.; Claassen, J.; et al. COVID-19 neuropathology at Columbia University Irving Medical Center/New York Presbyterian Hospital. Brain 2021, 144, 2696–2708. [Google Scholar] [CrossRef]
- Khaddaj-Mallat, R.; Aldib, N.; Bernard, M.; Paquette, A.S.; Ferreira, A.; Lecordier, S.; Saghatelyan, A.; Flamand, L.; El Ali, A. SARS-CoV-2 deregulates the vascular and immune functions of brain pericytes via Spike protein. Neurobiol. Dis. 2021, 161, 105561. [Google Scholar] [CrossRef]
- Hirunpattarasilp, C.; James, G.; Freitas, F.; Sethi, H.; Kittler, J.T.; Huo, J.; Owens, R.J.; Attwell, D. SARS-CoV-2 binding to ACE2 triggers pericyte-mediated angiotensin-evoked cerebral capillary constriction. bioRxiv 2021. [Google Scholar] [CrossRef]
- Eyring, K.W.; Geschwind, D.H. Three decades of ASD genetics: Building a foundation for neurobiological understanding and treatment. Hum. Mol. Genet. 2021, 30, R236–R244. [Google Scholar] [CrossRef]
- Ouellette, J.; Toussay, X.; Comin, C.H.; Costa, L.D.F.; Ho, M.; Lacalle-Aurioles, M.; Freitas-Andrade, M.; Liu, Q.Y.; Leclerc, S.; Pan, Y.; et al. Vascular contributions to 16p11.2 deletion autism syndrome modeled in mice. Nat. Neurosci. 2020, 23, 1090–1101. [Google Scholar] [CrossRef]
- Virgintino, D.; Girolamo, F.; Rizzi, M.; Ahmedli, N.; Sadowska, G.B.; Stopa, E.G.; Zhang, J.; Stonestreet, B.S. Ischemia/Reperfusion-induced neovascularization in the cerebral cortex of the ovine fetus. J. Neuropathol. Exp. Neurol. 2014, 73, 495–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziewulska, D.; Lewandowska, E. Pericytes as a new target for pathological processes in CADASIL. Neuropathology 2012, 32, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Genove, G.; Mollick, T.; Johansson, K. Photoreceptor degeneration, structural remodeling and glial activation: A morphological study on a genetic mouse model for pericyte deficiency. Neuroscience 2014, 279, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.C.; Vest, R.T.; Kern, F.; Lee, D.P.; Agam, M.; Maat, C.A.; Losada, P.M.; Chen, M.B.; Schaum, N.; Khoury, N.; et al. A human brain vascular atlas reveals diverse mediators of Alzheimer’s risk. Nature 2022, 603, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Jansson, D.; Rustenhoven, J.; Feng, S.; Hurley, D.; Oldfield, R.L.; Bergin, P.S.; Mee, E.W.; Faull, R.L.; Dragunow, M. A role for human brain pericytes in neuroinflammation. J. Neuroinflamm. 2014, 11, 104. [Google Scholar] [CrossRef] [Green Version]
- Persidsky, Y.; Hill, J.; Zhang, M.; Dykstra, H.; Winfield, M.; Reichenbach, N.L.; Potula, R.; Mukherjee, A.; Ramirez, S.H.; Rom, S. Dysfunction of brain pericytes in chronic neuroinflammation. J. Cereb. Blood Flow Metab. 2016, 36, 794–807. [Google Scholar] [CrossRef] [Green Version]
- Takata, F.; Dohgu, S.; Matsumoto, J.; Takahashi, H.; Machida, T.; Wakigawa, T.; Harada, E.; Miyaji, H.; Koga, M.; Nishioku, T.; et al. Brain pericytes among cells constituting the blood-brain barrier are highly sensitive to tumor necrosis factor-alpha, releasing matrix metalloproteinase-9 and migrating in vitro. J. Neuroinflamm. 2011, 8, 106. [Google Scholar] [CrossRef] [Green Version]
- Michinaga, S.; Koyama, Y. Dual Roles of Astrocyte-Derived Factors in Regulation of Blood-Brain Barrier Function after Brain Damage. Int. J. Mol. Sci. 2019, 20, 571. [Google Scholar] [CrossRef] [Green Version]





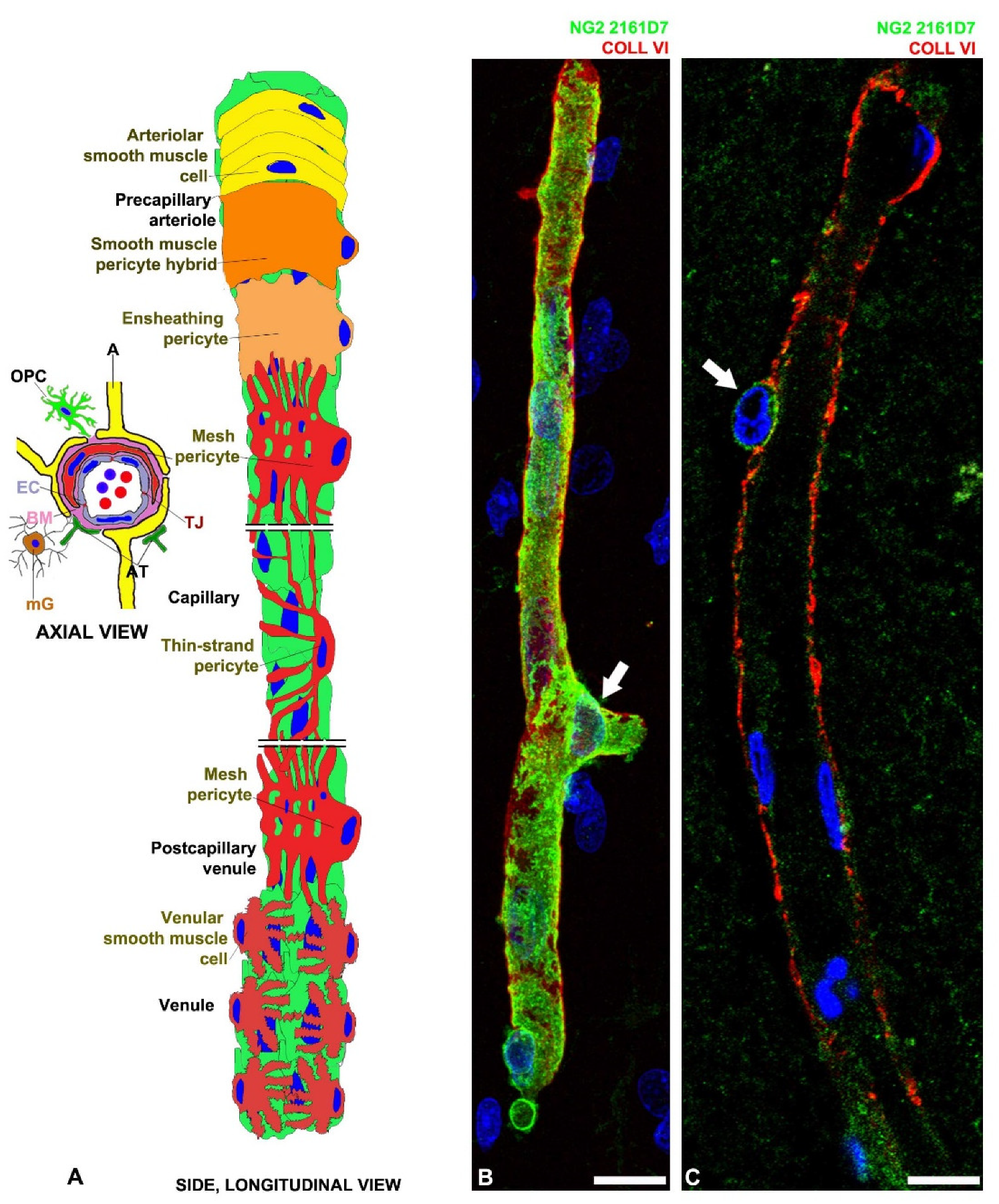{kind=link}
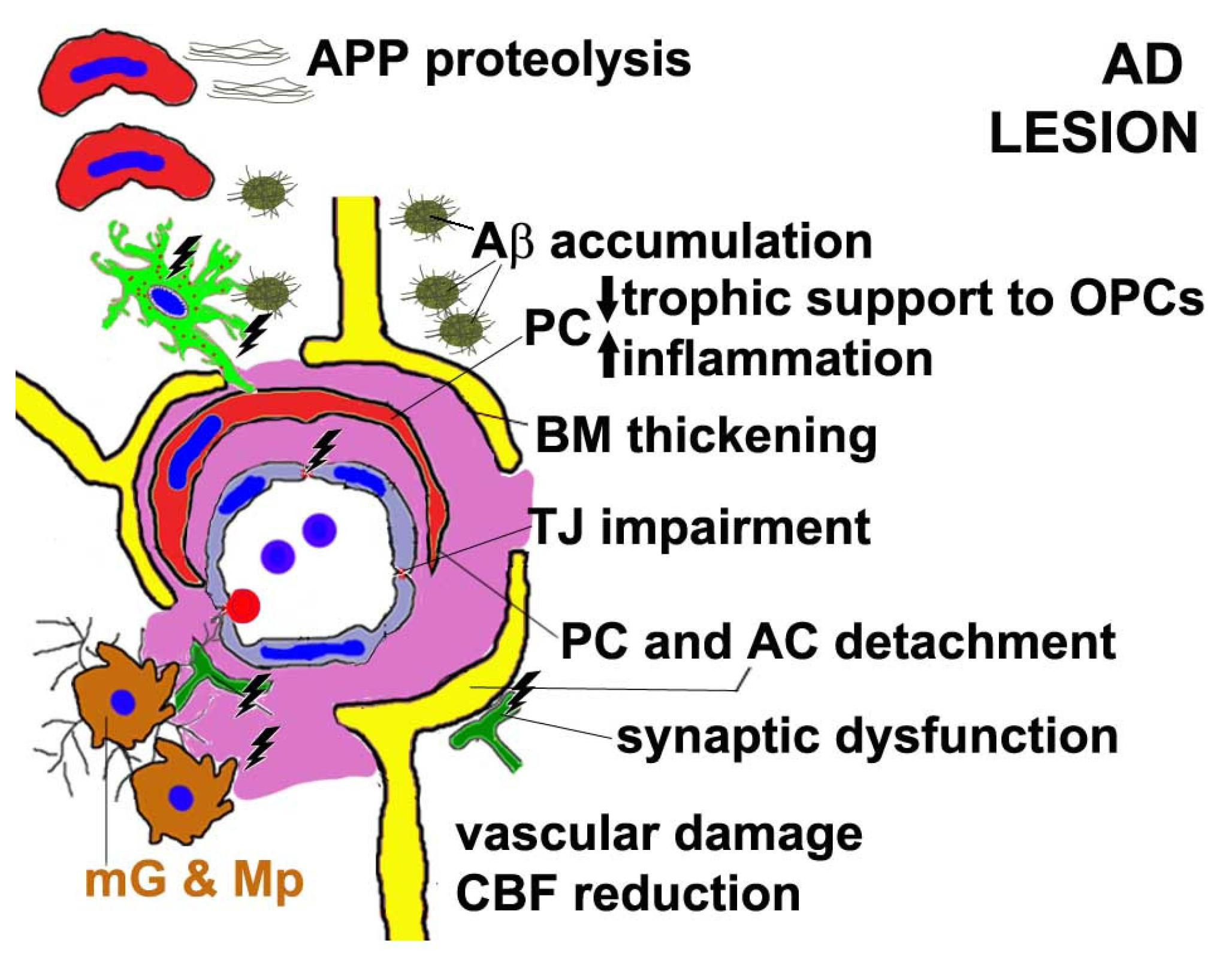{kind=link}
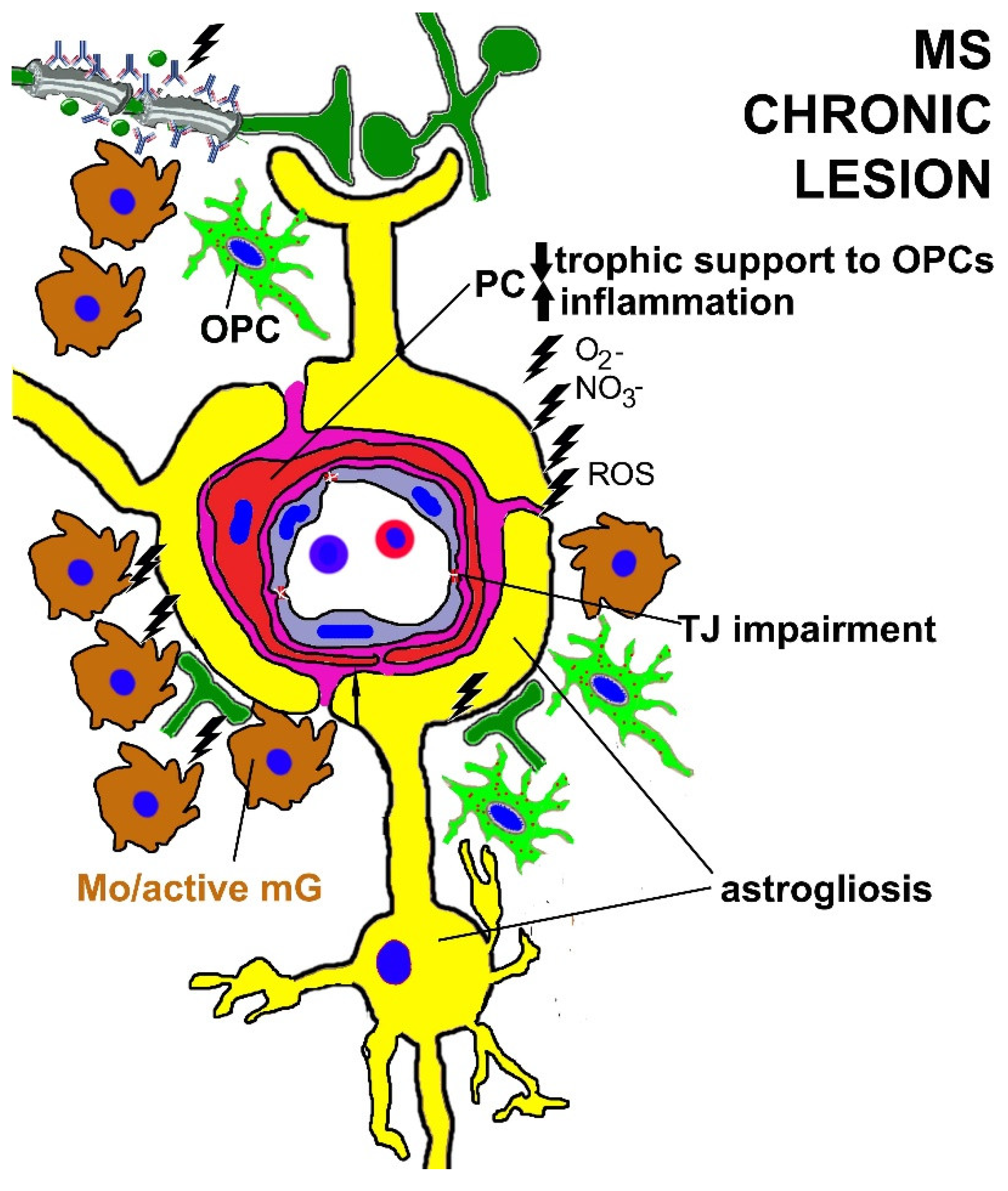{kind=link}
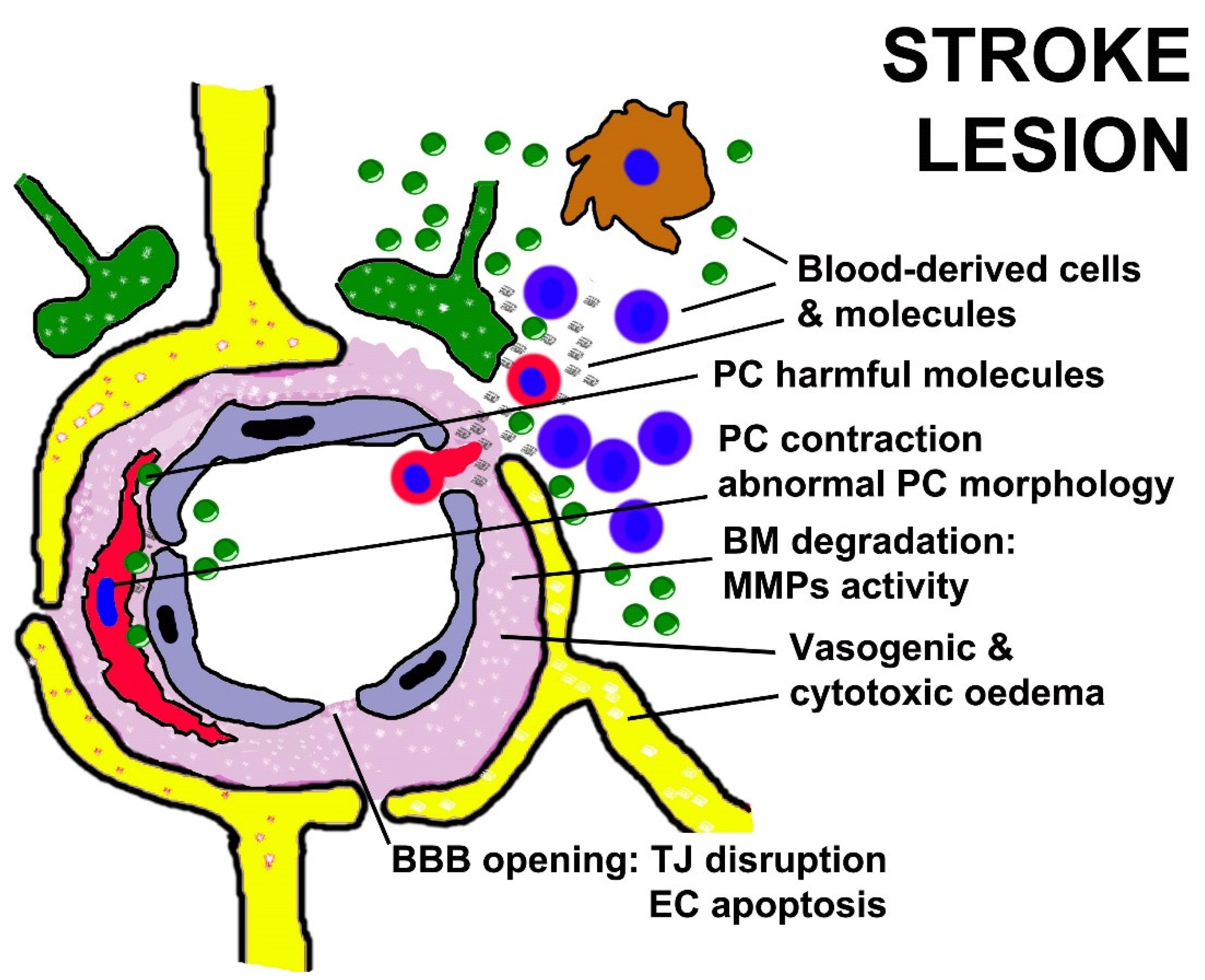{kind=link}
| PC Marker | Development | Healthy Adulthood |
|---|---|---|
| Neuron-glial antigen 2 (NG2) | [19,63] | Only activated PCs of CNS [64,65] |
| Platelet derived growth factor receptor beta (PDGFRβ) | [66,67] | [68] |
| Alanyl aminopeptidase (CD13) | [69] | [70,71] |
| Melanoma Cell Adhesion Molecule (CD146) | [71,72] | [73] |
| Endosialin (CD248) | [66,74] | Not expressed |
| Fluoro-Nissl dye NeuroTrace 500/525 | [75] | [76] |
| Disease | PC Dysfunction(s) in the Clinical Condition | PC Dysfunction(s) in Models |
|---|---|---|
| Autism spectrum disorder | Increased density [182]; genetic risk after PC gene modifications [370] | Putative NVU dysfunction [371] |
| Prenatal life hypoxia | Reduced PC coverage [189,190]; increased angiogenesis and bFGF [372] | |
| White matter degeneration | [373] | Reduced PC coverage [154] |
| Altered neuronal circuitry | [120] | Ablation of PC induce poor interaction between glia and neural precursors [184,374]. |
| AD | Hypoperfusion, PC degeneration and BBB breakdown [117,162,193,194,206,207,209,375]; PC inability to support oligodendrogenesis [233] | Decreased density of PDGFRβ+ PCs in 5xFAD mice [211] and CD13+ PCs in pericyte-deficient APPsw/0; Pdgfrβ+/− [213]; dysfunction and apoptosis of desmin+ PCs in APPswe/PS1 mice [225] |
| MS | PC inability to support oligodendrogenesis [232]; increased density of PDGFRβ+ CD146+ PC in active MS lesions [239]; increased angiogenesis [255,256,257] | PC inability to support oligodendrogenesis in different models [69,144,145,149,150,154,232,244,253]; PDGFRβ+ PC triggers inflammation [245,246,247,248,249,250,251,376,377,378]; increased angiogenesis [258,259,261,262,263,264,265] |
| Stroke | Microvascular PC contractions aggravate ischemia [121]; PCs release harmful molecules [297] | In different models, PCs release harmful molecules [184,225,248,254,352]; decreased PC density, EC apoptosis, and BBB disruption [347,348,350,351]; abnormal PC morphology [354] |
| NeuroCOVID-19 | SARS-CoV-2 infects ACE2+ PCs [356,363,365] | PC vasoconstriction [368,369] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Girolamo, F.; Errede, M.; Bizzoca, A.; Virgintino, D.; Ribatti, D. Central Nervous System Pericytes Contribute to Health and Disease. Cells 2022, 11, 1707. https://doi.org/10.3390/cells11101707
Girolamo F, Errede M, Bizzoca A, Virgintino D, Ribatti D. Central Nervous System Pericytes Contribute to Health and Disease. Cells. 2022; 11(10):1707. https://doi.org/10.3390/cells11101707
Chicago/Turabian StyleGirolamo, Francesco, Mariella Errede, Antonella Bizzoca, Daniela Virgintino, and Domenico Ribatti. 2022. "Central Nervous System Pericytes Contribute to Health and Disease" Cells 11, no. 10: 1707. https://doi.org/10.3390/cells11101707
APA StyleGirolamo, F., Errede, M., Bizzoca, A., Virgintino, D., & Ribatti, D. (2022). Central Nervous System Pericytes Contribute to Health and Disease. Cells, 11(10), 1707. https://doi.org/10.3390/cells11101707