
Stem Cell-Based Trophoblast Models to Unravel the Genetic Causes of Human Miscarriages


Abstract
:1. Introduction
2. Genetic Causes of Miscarriage
3. Placental Defects Are the Main Cause of First-Trimester Human Miscarriage
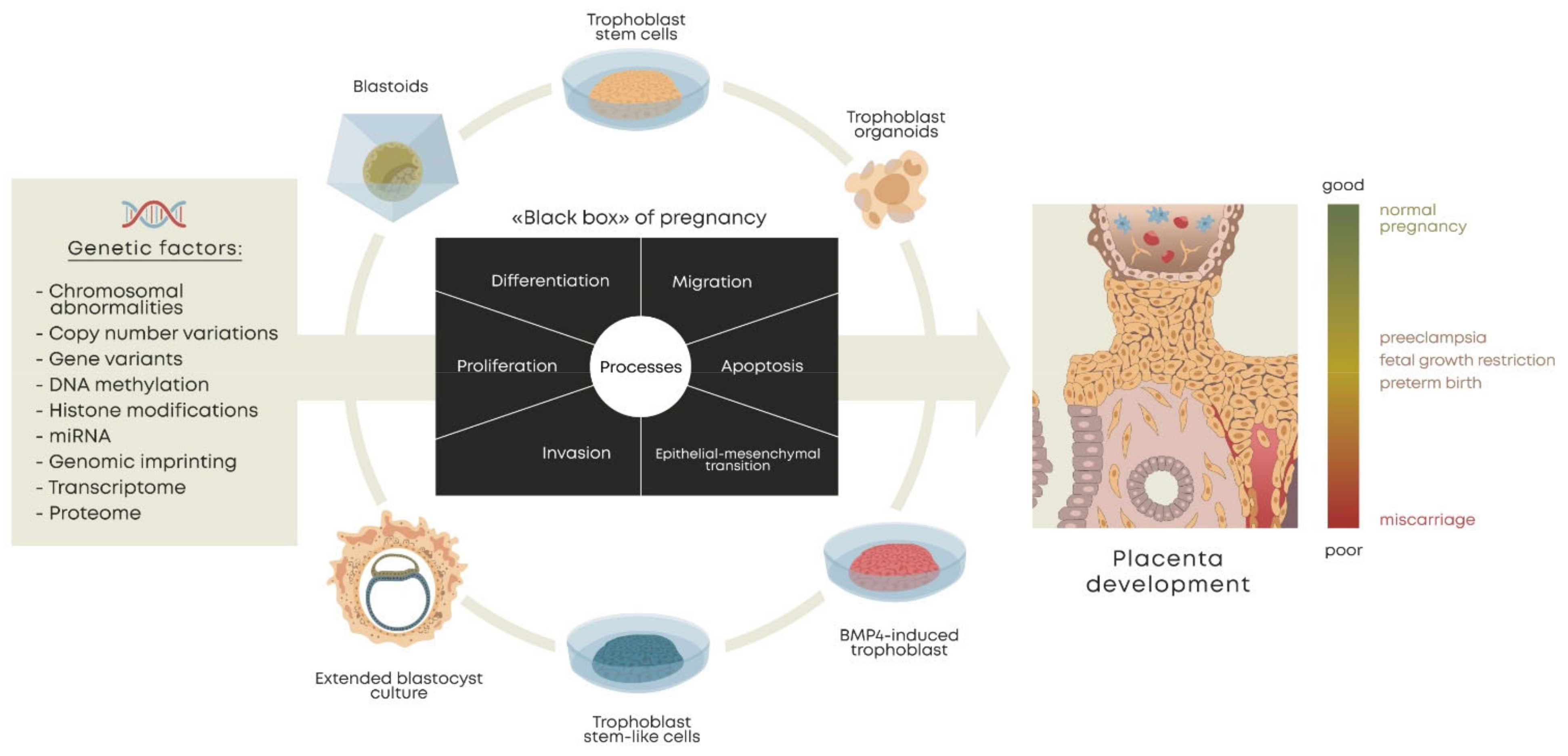
4. Stem Cell-Based Trophoblast Models
5. Methods for Derivation of Cells and Organoids for Trophoblast Modeling
5.1. BMP4-Induced Trophoblast-Like Cells (Terminal Trophoblast-Like Cells)
5.2. “True” Human Trophoblast Stem Cells (hTSCs)
5.3. Trophoblast Stem-Like Cells (TSLC)
5.4. Induced Trophoblast Stem Cells (iTSCs), TF-Mediated Conversion Models
5.5. 3D Culture Models
5.6. Artificial Embryos Generated Using TS and ES Cells Together (Blastoids)
5.7. Extended Blastocyst Culture
6. Availability and Advantages of Different Types of Cellular Material for Miscarriage Studies
7. Stem Cell-Based Studies of Chromosomal Aneuploidy and Mosaicism in Human Trophoblast Development
8. Future Perspectives
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Macklon, N.S.; Geraedts, J.P.M.; Fauser, B.C.J.M. Conception to ongoing pregnancy: The ‘black box’ of early pregnancy loss. Hum. Reprod. Updat. 2002, 8, 333–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, E.C.; Christiansen, O.B.; Kolte, A.M.; Macklon, N. New insights into mechanisms behind miscarriage. BMC Med. 2013, 11, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilcox, A.J.; Harmon, Q.; Doody, K.; Wolf, D.P.; Adashi, E.Y. Preimplantation loss of fertilized human ova: Estimating the unobservable. Hum. Reprod. 2020, 35, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Short, R.V. When a Conception Fails to Become a Pregnancy. Matern. Recognit. Pregnancy 1979, 377–411. [Google Scholar] [CrossRef]
- Fragouli, E.; Alfarawati, S.; Spath, K.; Jaroudi, S.; Sarasa, J.; Enciso, M.; Wells, D. The origin and impact of embryonic aneuploidy. Qual. Life Res. 2013, 132, 1001–1013. [Google Scholar] [CrossRef]
- Franasiak, J.M.; Forman, E.J.; Hong, K.H.; Werner, M.D.; Upham, K.M.; Treff, N.R.; Scott, R.T. Aneuploidy across individual chromosomes at the embryonic level in trophectoderm biopsies: Changes with patient age and chromosome structure. J. Assist. Reprod. Genet. 2014, 31, 1501–1509. [Google Scholar] [CrossRef] [Green Version]
- Honnma, H.; Baba, T.; Sasaki, M.; Hashiba, Y.; Ohno, H.; Fukunaga, T.; Endo, T.; Saito, T.; Asada, Y. Trophectoderm morphology significantly affects the rates of ongoing pregnancy and miscarriage in frozen-thawed single-blastocyst transfer cycle in vitro fertilization. Fertil. Steril. 2012, 98, 361–367. [Google Scholar] [CrossRef]
- Saha, B.; Ganguly, A.; Home, P.; Bhattacharya, B.; Ray, S.; Ghosh, A.; Rumi, M.A.K.; Marsh, C.; French, V.A.; Gunewardena, S.; et al. TEAD4 ensures postimplantation development by promoting trophoblast self-renewal: An implication in early human pregnancy loss. Proc. Natl. Acad. Sci. USA 2020, 117, 17864–17875. [Google Scholar] [CrossRef]
- Del Gobbo, G.F.; Konwar, C.; Robinson, W.P. The significance of the placental genome and methylome in fetal and maternal health. Qual. Life Res. 2019, 139, 1183–1196. [Google Scholar] [CrossRef]
- Menasha, J.; Levy, B.; Hirschhorn, K.; Kardon, N.B. Incidence and spectrum of chromosome abnormalities in spontaneous abortions: New insights from a 12-year study. Genet. Med. 2005, 7, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, M.M.; van Maarle, M.C.; van Wely, M.; Goddijn, M. Genetics of early miscarriage. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 1951–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, M.A.; van Maarle, M.; Hamer, G.; Mastenbroek, S.; Goddijn, M.; van Wely, M. Cytogenetic testing of pregnancy loss tissue: A meta-analysis. Reprod. Biomed. Online 2020, 40, 867–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, B.; Sigurjonsson, S.; Pettersen, B.; Maisenbacher, M.K.; Hall, M.P.; Demko, Z.; Lathi, R.B.; Tao, R.; Aggarwal, V.; Rabinowitz, M. Genomic Imbalance in Products of Conception. Obstet. Gynecol. 2014, 124, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, T.; Dzidic, N.; Strecker, M.N.; Commander, S.; Travis, M.K.; Doherty, C.; Tyson, R.W.; Mendoza, A.E.; Stephenson, M.; Dise, C.A.; et al. Comprehensive genetic analysis of pregnancy loss by chromosomal microarrays: Outcomes, benefits, and challenges. Genet. Med. 2017, 19, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Wu, J.; Wu, Y.; Shi, X.; Xin, X.; Li, S.; Zeng, W.; Deng, D.; Feng, L.; Chen, S.; et al. Analysis of Chromosomal Copy Number in First-Trimester Pregnancy Loss Using Next-Generation Sequencing. Front. Genet. 2020, 11, 545856. [Google Scholar] [CrossRef] [PubMed]
- Greco, E.; Minasi, M.G.; Fiorentino, F. Healthy Babies after Intrauterine Transfer of Mosaic Aneuploid Blastocysts. N. Engl. J. Med. 2015, 373, 2089–2090. [Google Scholar] [CrossRef] [Green Version]
- Munné, S.; Wells, D. Detection of mosaicism at blastocyst stage with the use of high-resolution next-generation sequencing. Fertil. Steril. 2017, 107, 1085–1091. [Google Scholar] [CrossRef] [Green Version]
- Patrizio, P.; Shoham, G.; Shoham, Z.; Leong, M.; Barad, D.H.; Gleicher, N. Worldwide live births following the transfer of chromosomally “Abnormal” embryos after PGT/A: Results of a worldwide web-based survey. J. Assist. Reprod. Genet. 2019, 36, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- Hong, B.; Hao, Y. The outcome of human mosaic aneuploid blastocysts after intrauterine transfer. Medicine 2020, 99, e18768. [Google Scholar] [CrossRef]
- Capalbo, A.; Poli, M.; Rienzi, L.; Girardi, L.; Patassini, C.; Fabiani, M.; Cimadomo, D.; Benini, F.; Farcomeni, A.; Cuzzi, J.; et al. Mosaic human preimplantation embryos and their developmental potential in a prospective, non-selection clinical trial. Am. J. Hum. Genet. 2021, 108, 2238–2247. [Google Scholar] [CrossRef] [PubMed]
- Viotti, M.; McCoy, R.C.; Griffin, D.K.; Spinella, F.; Greco, E.; Madjunkov, M.; Madjunkova, S.; Librach, C.L.; Victor, A.R.; Barnes, F.L.; et al. Let the data do the talking: The need to consider mosaicism during embryo selection. Fertil. Steril. 2021, 116, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Orvieto, R. The reproducibility of trophectoderm biopsies—The chaos behind preimplantation genetic testing for aneuploidy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 254, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Taylor, T.H.; Stankewicz, T.; Katz, S.L.; Patrick, J.L.; Johnson, L.; Griffin, D.K. Preliminary assessment of aneuploidy rates between the polar, mid and mural trophectoderm. Zygote 2019, 28, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Kalousek, D.K.; Howard-Peebles, P.N.; Olson, S.B.; Barrett, I.J.; Dorfmann, A.; Black, S.H.; Schulman, J.D.; Wilson, R.D. Confirmation of CVS mosaicism in term placentae and high frequency of intrauterine growth retardation association with confined placental mosaicism. Prenat. Diagn. 1991, 11, 743–750. [Google Scholar] [CrossRef]
- Gardner, R.M.; Sutherland, G.R.; Shaffer, L.G. Chromosome Abnormalities and Genetic Counseling; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Andersen, A.-M.N.; Wohlfahrt, J.; Christens, P.; Olsen, J.; Melbye, M. Maternal age and fetal loss: Population based register linkage study. BMJ 2000, 320, 1708–1712. [Google Scholar] [CrossRef] [Green Version]
- Bruckner, T.A.; Mortensen, L.H.; Catalano, R.A. Spontaneous Pregnancy Loss in Denmark Following Economic Downturns. Am. J. Epidemiol. 2016, 183, 701–708. [Google Scholar] [CrossRef] [Green Version]
- Quenby, S.; Gallos, I.D.; Dhillon-Smith, R.K.; Podesek, M.; Stephenson, M.D.; Fisher, J.; Brosens, J.J.; Brewin, J.; Ramhorst, R.; Lucas, E.S.; et al. Miscarriage matters: The epidemiological, physical, psychological, and economic costs of early pregnancy loss. Lancet 2021, 397, 1658–1667. [Google Scholar] [CrossRef]
- Rull, K.; Nagirnaja, L.; Laan, M. Genetics of Recurrent Miscarriage: Challenges, Current Knowledge, Future Directions. Front. Genet. 2012, 3, 34. [Google Scholar] [CrossRef] [Green Version]
- Dimitriadis, E.; Menkhorst, E.; Saito, S.; Kutteh, W.H.; Brosens, J.J. Recurrent pregnancy loss. Nat. Rev. Dis. Prim. 2020, 6, 1–19. [Google Scholar] [CrossRef]
- Magnus, M.C.; Wilcox, A.J.; Morken, N.-H.; Weinberg, C.; Håberg, S.E. Role of maternal age and pregnancy history in risk of miscarriage: Prospective register based study. BMJ 2019, 364, l869. [Google Scholar] [CrossRef] [Green Version]
- Sugiura-Ogasawara, M.; Ozaki, Y.; Katano, K.; Suzumori, N.; Kitaori, T.; Mizutani, E. Abnormal embryonic karyotype is the most frequent cause of recurrent miscarriage. Hum. Reprod. 2012, 27, 2297–2303. [Google Scholar] [CrossRef] [PubMed]
- Feichtinger, M.; Reiner, A.; Hartmann, B.; Philipp, T. Embryoscopy and karyotype findings of repeated miscarriages in recurrent pregnancy loss and spontaneous pregnancy loss. J. Assist. Reprod. Genet. 2018, 35, 1401–1406. [Google Scholar] [CrossRef] [PubMed]
- Nikitina, T.V.; Sazhenova, E.A.; Zhigalina, D.I.; Tolmacheva, E.N.; Sukhanova, N.N.; Lebedev, I.N. Karyotype evaluation of repeated abortions in primary and secondary recurrent pregnancy loss. J. Assist. Reprod. Genet. 2020, 37, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Woolner, A.M.F.; Nagdeve, P.; Raja, E.; Bhattacharya, S.; Bhattacharya, S. Family history and risk of miscarriage: A systematic review and meta-analysis of observational studies. Acta Obstet. Gynecol. Scand. 2020, 99, 1584–1594. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, O.B.; Mathiesen, O.; Lauritsen, J.G.; Grunnet, N. Idiopathic Recurrent Spontaneous Abortion: Evidence of a Familial Predisposition. Acta Obstet. Gynecol. Scand. 1990, 69, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Kolte, A.; Nielsen, H.; Moltke, I.; Degn, B.; Pedersen, B.; Sunde, L.; Nielsen, F.; Christiansen, O. A genome-wide scan in affected sibling pairs with idiopathic recurrent miscarriage suggests genetic linkage. Mol. Hum. Reprod. 2011, 17, 379–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boué, J.; Boué, A.; Lazar, P. Retrospective and prospective epidemiological studies of 1500 karyotyped spontaneous human abortions. Teratology 1975, 12, 11–26. [Google Scholar] [CrossRef]
- Hassold, T.J. A cytogenetic study of repeated spontaneous abortions. Am. J. Hum. Genet. 1980, 32, 723–730. [Google Scholar]
- Kajii, T.; Ferrier, A.; Niikawa, N.; Takahara, H.; Ohama, K.; Avirachan, S. Anatomic and chromosomal anomalies in 639 spontaneous abortuses. Qual. Life Res. 1980, 55, 87–98. [Google Scholar] [CrossRef]
- Eiben, B.; Bartels, I.; Bähr-Porsch, S.; Borgmann, S.; Gatz, G.; Gellert, G.; Goebel, R.; Hammans, W.; Hentemann, M.; Osmers, R. Cytogenetic analysis of 750 spontaneous abortions with the direct-preparation method of chorionic villi and its implications for studying genetic causes of pregnancy wastage. Am. J. Hum. Genet. 1990, 47, 656–663. [Google Scholar]
- Dejmek, J.; Vojtaššák, J.; Malová, J. Cytogenetic analysis of 1508 spontaneous abortions originating from south Slovakia. Eur. J. Obstet. Gynecol. Reprod. Biol. 1992, 46, 129–136. [Google Scholar] [CrossRef]
- Hardy, K.; Hardy, P.J.; Jacobs, P.A.; Lewallen, K.; Hassold, T.J. Temporal changes in chromosome abnormalities in human spontaneous abortions: Results of 40 years of analysis. Am. J. Med. Genet. Part A 2016, 170, 2671–2680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, R.; Hafezi, N.; Amrani, M.; Schweiger, S.; Dewenter, M.K.; Thomas, P.; Lieb, C.; Hasenburg, A.; Skala, C. Genetic findings in miscarriages and their relation to the number of previous miscarriages. Arch. Gynecol. Obstet. 2020, 303, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yuan, D.; Wang, S.; Luo, L.; Zhang, Y.; Ye, J.; Zhu, K. Cytogenetic and genetic investigation of miscarriage cases in Eastern China. J. Matern. Neonatal Med. 2019, 33, 3385–3390. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Su, L.; Xie, X.; He, D.; Chen, X.; Wang, M.; Wang, L.; Zheng, L.; Xu, L. Comprehensive analysis of early pregnancy loss based on cytogenetic findings from a tertiary referral center. Mol. Cytogenet. 2021, 14, 56. [Google Scholar] [CrossRef]
- Lebedev, I.N.; Ostroverkhova, N.V.; Nikitina, T.V.; Sukhanova, N.N.; Nazarenko, S.A. Features of chromosomal abnormalities in spontaneous abortion cell culture failures detected by interphase FISH analysis. Eur. J. Hum. Genet. 2004, 12, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Soler, A.; Morales, C.; Mademont-Soler, I.; Margarit, E.; Borrell, A.; Borobio, V.; Muñoz, M.; Sánchez, A. Overview of Chromosome Abnormalities in First Trimester Miscarriages: A Series of 1,011 Consecutive Chorionic Villi Sample Karyotypes. Cytogenet. Genome Res. 2017, 152, 81–89. [Google Scholar] [CrossRef]
- Maisenbacher, M.K.; Merrion, K.; Levy, B.; Kutteh, W.H. Single nucleotide polymorphism (SNP) array analysis of 63,277 products of conception (POC) samples: A 10-year laboratory experience. Fertil. Steril. 2020, 114, e47. [Google Scholar] [CrossRef]
- Gu, C.; Li, K.; Li, R.; Li, L.; Li, X.; Dai, X.; He, Y. Chromosomal Aneuploidy Associated with Clinical Characteristics of Pregnancy Loss. Front. Genet. 2021, 12, 667697. [Google Scholar] [CrossRef]
- Pereza, N.; Ostojić, S.; Kapović, M.; Peterlin, B. Systematic review and meta-analysis of genetic association studies in idiopathic recurrent spontaneous abortion. Fertil. Steril. 2016, 107, 150–159.e2. [Google Scholar] [CrossRef] [Green Version]
- Kline, J.; Vardarajan, B.; Abhyankar, A.; Kytömaa, S.; Levin, B.; Sobreira, N.; Tang, A.; Thomas-Wilson, A.; Zhang, R.; Jobanputra, V. Embryonic lethal genetic variants and chromosomally normal pregnancy loss. Fertil. Steril. 2021, 116, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.-R.; Hou, S.-Y.; Hu, W.-T.; Wei, C.-Y.; Liu, Y.-K.; Liu, Y.-Y.; Jiang, L.; Xiang, J.-J.; Sun, X.-X.; Lei, C.-X.; et al. Characterization of Copy-Number Variations and Possible Candidate Genes in Recurrent Pregnancy Losses. Genes 2021, 12, 141. [Google Scholar] [CrossRef] [PubMed]
- Colley, E.; Hamilton, S.; Smith, P.; Morgan, N.V.; Coomarasamy, A.; Allen, S. Potential genetic causes of miscarriage in euploid pregnancies: A systematic review. Hum. Reprod. Updat. 2019, 25, 452–472. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Ronderos, P.; Laissue, P. Genetic Variants Contributing to Early Recurrent Pregnancy Loss Etiology Identified by Sequencing Approaches. Reprod. Sci. 2020, 27, 1541–1552. [Google Scholar] [CrossRef] [PubMed]
- Rajcan-Separovic, E. Next generation sequencing in recurrent pregnancy loss-approaches and outcomes. Eur. J. Med Genet. 2019, 63, 103644. [Google Scholar] [CrossRef]
- Boss, A.L.; Chamley, L.W.; James, J. Placental formation in early pregnancy: How is the centre of the placenta made? Hum. Reprod. Updat. 2018, 24, 750–760. [Google Scholar] [CrossRef]
- Knöfler, M.; Haider, S.; Saleh, L.; Pollheimer, J.; Gamage, T.K.J.B.; James, J. Human placenta and trophoblast development: Key molecular mechanisms and model systems. Exp. 2019, 76, 3479–3496. [Google Scholar] [CrossRef] [Green Version]
- Silini, A.R.; Di Pietro, R.; Lang-Olip, I.; Alviano, F.; Banerjee, A.; Basile, M.; Borutinskaite, V.; Eissner, G.; Gellhaus, A.; Giebel, B.; et al. Perinatal Derivatives: Where Do We Stand? A Roadmap of the Human Placenta and Consensus for Tissue and Cell Nomenclature. Front. Bioeng. Biotechnol. 2020, 8, 610544. [Google Scholar] [CrossRef]
- Perez-Garcia, V.; Fineberg, E.; Wilson, R.; Murray, A.; Mazzeo, C.I.; Tudor, C.; Sienerth, A.; White, J.K.; Tuck, E.; Ryder, E.J.; et al. Placentation defects are highly prevalent in embryonic lethal mouse mutants. Nature 2018, 555, 463–468. [Google Scholar] [CrossRef]
- Shukla, V.; Soares, M.J. Modeling Trophoblast Cell-Guided Uterine Spiral Artery Transformation in the Rat. Int. J. Mol. Sci. 2022, 23, 2947. [Google Scholar] [CrossRef]
- Laisk, T.; Soares, A.L.G.; Ferreira, T.; Painter, J.N.; Censin, J.C.; Laber, S.; Bacelis, J.; Chen, C.-Y.; Lepamets, M.; Lin, K.; et al. The genetic architecture of sporadic and multiple consecutive miscarriage. Nat. Commun. 2020, 11, 5980. [Google Scholar] [CrossRef] [PubMed]
- Morgan, T.K. Role of the Placenta in Preterm Birth: A Review. Am. J. Perinatol. 2016, 33, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. The cytotrophoblastic shell and complications of pregnancy. Placenta 2017, 60, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Pathophysiology of placental-derived fetal growth restriction. Am. J. Obstet. Gynecol. 2018, 218, S745–S761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, G.J.; Cindrova-Davies, T.; Yung, H.W.; Jauniaux, E. Hypoxia and Reproductive Health: Oxygen and development of the human placenta. Reproduction 2021, 161, F53–F65. [Google Scholar] [CrossRef]
- Kaufmann, P.; Black, S.; Huppertz, B. Endovascular Trophoblast Invasion: Implications for the Pathogenesis of Intrauterine Growth Retardation and Preeclampsia. Biol. Reprod. 2003, 69, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Brosens, I.; Pijnenborg, R.; Vercruysse, L.; Romero, R. The “Great Obstetrical Syndromes” are associated with disorders of deep placentation. Am. J. Obstet. Gynecol. 2011, 204, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Velicky, P.; Knöfler, M.; Pollheimer, J. Function and control of human invasive trophoblast subtypes: Intrinsic vs. maternal control. Cell Adhes. Migr. 2016, 10, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Funai, E.F.; Friedlander, Y.; Paltiel, O.; Tiram, E.; Xue, X.; Deutsch, L.; Harlap, S. Long-Term Mortality After Preeclampsia. Epidemiology 2005, 16, 206–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gluckman, P.D.; Hanson, M.; Cooper, C.; Thornburg, K. Effect of In Utero and Early-Life Conditions on Adult Health and Disease. N. Engl. J. Med. 2008, 359, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Ursini, G.; Punzi, G.; Langworthy, B.W.; Chen, Q.; Xia, K.; Cornea, E.A.; Goldman, B.D.; Styner, M.A.; Knickmeyer, R.C.; Gilmore, J.H.; et al. Placental genomic risk scores and early neurodevelopmental outcomes. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.P.; Wainstock, T.; Sheiner, E.; Sergienko, R.; Landau, D.; Walfisch, A. Maternal recurrent pregnancy loss is associated with an increased risk for long-term neurological morbidity in offspring. Dev. Med. Child Neurol. 2018, 61, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, F.; Miao, M.; Yu, Y.; Ji, H.; Liu, H.; Huang, R.; Obel, C.; Zhang, J.; Li, J. Maternal spontaneous abortion and the risk of attention-deficit/hyperactivity disorder in offspring: A population-based cohort study. Hum. Reprod. 2020, 35, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Yu, Y.; Miao, M.; Qian, X.; Yuan, W.; Lin, Y.; Liang, H.; Li, J. Risk of intellectual disability and maternal history of spontaneous abortion: A nationwide cohort study. Dev. Med. Child Neurol. 2021, 63, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Soncin, F.; Natale, D.; Parast, M.M. Signaling pathways in mouse and human trophoblast differentiation: A comparative review. Cell. Mol. Life Sci. 2014, 72, 1291–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soncin, F.; Khater, M.; To, C.; Pizzo, D.; Farah, O.; Wakeland, A.; Rajan, K.A.N.; Nelson, K.K.; Chang, C.-W.; Moretto-Zita, M.; et al. Comparative analysis of mouse and human placentae across gestation reveals species-specific regulators of placental development. Development 2018, 145. [Google Scholar] [CrossRef] [Green Version]
- Shahbazi, M.N.; Siggia, E.D.; Zernicka-Goetz, M. Self-organization of stem cells into embryos: A window on early mammalian development. Science 2019, 364, 948–951. [Google Scholar] [CrossRef]
- Hemberger, M.; Hanna, C.W.; Dean, W. Mechanisms of early placental development in mouse and humans. Nat. Rev. Genet. 2019, 21, 27–43. [Google Scholar] [CrossRef]
- Shibata, S.; Kobayashi, E.H.; Kobayashi, N.; Oike, A.; Okae, H.; Arima, T. Unique features and emerging in vitro models of human placental development. Reprod. Med. Biol. 2020, 19, 301–313. [Google Scholar] [CrossRef]
- Io, S.; Kondoh, E.; Chigusa, Y.; Kawasaki, K.; Mandai, M.; Yamada, A.S. New era of trophoblast research: Integrating morphological and molecular approaches. Hum. Reprod. Updat. 2020, 26, 611–633. [Google Scholar] [CrossRef]
- Lee, B.-K.; Kim, J. Integrating High-Throughput Approaches and in vitro Human Trophoblast Models to Decipher Mechanisms Underlying Early Human Placenta Development. Front. Cell Dev. Biol. 2021, 9, 673065. [Google Scholar] [CrossRef]
- Kunath, T.; Yamanaka, Y.; Detmar, J.; MacPhee, D.; Caniggia, I.; Rossant, J.; Jurisicova, A. Developmental differences in the expression of FGF receptors between human and mouse embryos. Placenta 2014, 35, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Alici-Garipcan, A.; Özçimen, B.; Süder, I.; Ülker, V.; Onder, T.; Özören, N. NLRP7 plays a functional role in regulating BMP4 signaling during differentiation of patient-derived trophoblasts. Cell Death Dis. 2020, 11, 658. [Google Scholar] [CrossRef] [PubMed]
- Graham, C.H.; Hawley, T.S.; Hawley, R.C.; MacDougall, J.R.; Kerbel, R.S.; Khoo, N.; Lala, P.K. Establishment and Characterization of First Trimester Human Trophoblast Cells with Extended Lifespan. Exp. Cell Res. 1993, 206, 204–211. [Google Scholar] [CrossRef]
- Feng, H.C.; Choy, M.Y.; Deng, W.; Wong, H.L.; Lau, W.M.; Cheung, A.N.Y.; Ngan, H.Y.S.; Tsao, S.W. Establishment and Characterization of a Human First-Trimester Extravillous Trophoblast Cell Line (TEV-1). J. Soc. Gynecol. Investig. 2005, 12, e21–e32. [Google Scholar] [CrossRef] [PubMed]
- Hiden, U.; Wadsack, C.; Prutsch, N.; Gauster, M.; Weiss, U.; Frank, H.-G.; Schmitz, U.; Fast-Hirsch, C.; Hengstschläger, M.; Pötgens, A.; et al. The first trimester human trophoblast cell line ACH-3P: A novel tool to study autocrine/paracrine regulatory loops of human trophoblast subpopulations—TNF-α stimulates MMP15 expression. BMC Dev. Biol. 2007, 7, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, T.; Handschuh, K.; Tsatsaris, V.; Evain-Brion, D. Involvement of PPARγ in Human Trophoblast Invasion. Placenta 2007, 28, S76–S81. [Google Scholar] [CrossRef]
- Mischler, A.; Karakis, V.; Mahinthakumar, J.; Carberry, C.K.; Miguel, A.S.; Rager, J.E.; Fry, R.C.; Rao, B.M. Two distinct trophectoderm lineage stem cells from human pluripotent stem cells. J. Biol. Chem. 2021, 296, 100386. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, T.; Ma, L.; Zhang, Y.; Zhao, Y.; Lye, K.; Xiao, L.; Chen, C.; Wang, Z.; Ma, Y.; et al. Efficient derivation of human trophoblast stem cells from primed pluripotent stem cells. Sci. Adv. 2021, 7, eabf4416. [Google Scholar] [CrossRef]
- Lee, C.Q.; Gardner, L.; Turco, M.; Zhao, N.; Murray, M.J.; Coleman, N.; Rossant, J.; Hemberger, M.; Moffett, A. What Is Trophoblast? A Combination of Criteria Define Human First-Trimester Trophoblast. Stem Cell Rep. 2016, 6, 257–272. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.-H.; Chen, X.; Li, D.S.; Li, R.; Addicks, G.C.; Glennon, C.; Zwaka, T.P.; Thomson, J.A. BMP4 initiates human embryonic stem cell differentiation to trophoblast. Nat. Biotechnol. 2002, 20, 1261–1264. [Google Scholar] [CrossRef] [PubMed]
- Amita, M.; Adachi, K.; Alexenko, A.P.; Sinha, S.; Schust, D.J.; Schulz, L.C.; Roberts, R.M.; Ezashi, T. Complete and unidirectional conversion of human embryonic stem cells to trophoblast by BMP4. Proc. Natl. Acad. Sci. USA 2013, 110, E1212–E1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Moretto-Zita, M.; Soncin, F.; Wakeland, A.; Wolfe, L.; Leon-Garcia, S.; Pandian, R.; Pizzo, D.; Cui, L.; Nazor, K.; et al. BMP4-directed trophoblast differentiation of human embryonic stem cells is mediated through a ΔNp63+ cytotrophoblast stem cell state. Development 2013, 140, 3965–3976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wang, K.; Chandramouli, G.V.; Knott, J.G.; Leach, R. Trophoblast lineage cells derived from human induced pluripotent stem cells. Biochem. Biophys. Res. Commun. 2013, 436, 677–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horii, M.; Li, Y.; Wakeland, A.K.; Pizzo, D.P.; Nelson, K.K.; Sabatini, K.; Laurent, L.C.; Liu, Y.; Parast, M.M. Human pluripotent stem cells as a model of trophoblast differentiation in both normal development and disease. Proc. Natl. Acad. Sci. USA 2016, 113, 201604747. [Google Scholar] [CrossRef] [Green Version]
- Guo, G.; Stirparo, G.G.; Strawbridge, S.E.; Spindlow, D.; Yang, J.; Clarke, J.; Dattani, A.; Yanagida, A.; Li, M.A.; Myers, S.; et al. Human naive epiblast cells possess unrestricted lineage potential. Cell Stem Cell 2021, 28, 1040–1056.e6. [Google Scholar] [CrossRef]
- Io, S.; Kabata, M.; Iemura, Y.; Semi, K.; Morone, N.; Minagawa, A.; Wang, B.; Okamoto, I.; Nakamura, T.; Kojima, Y.; et al. Capturing human trophoblast development with naive pluripotent stem cells in vitro. Cell Stem Cell 2021, 28, 1023–1039.e13. [Google Scholar] [CrossRef]
- Bernardo, A.S.; Faial, T.; Gardner, L.; Niakan, K.K.; Ortmann, D.; Senner, C.E.; Callery, E.M.; Trotter, M.W.; Hemberger, M.; Smith, J.C.; et al. BRACHYURY and CDX2 Mediate BMP-Induced Differentiation of Human and Mouse Pluripotent Stem Cells into Embryonic and Extraembryonic Lineages. Cell Stem Cell 2011, 9, 144–155. [Google Scholar] [CrossRef] [Green Version]
- Tietze, E.; Barbosa, A.R.; Euclydes, V.; Cho, H.J.; Lee, Y.K.; Feltrin, A.; van de Leemput, J.; Di Carlo, P.; Sawada, T.; Benjamin, K.J.; et al. Single-cell analysis of human trophoblast stem cell specification reveals activation of fetal cytotropho-blast expression programs including coronavirus associated host factors and hu-man endogenous retroviruses. bioRxiv 2020. [Google Scholar] [CrossRef]
- Horii, M.; Bui, T.; Touma, O.; Cho, H.Y.; Parast, M.M. An Improved Two-Step Protocol for Trophoblast Differentiation of Human Pluripotent Stem Cells. Curr. Protoc. Stem Cell Biol. 2019, 50, e96. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R.M.; Ezashi, T.; Sheridan, M.A.; Yang, Y. Specification of trophoblast from embryonic stem cells exposed to BMP4. Biol. Reprod. 2018, 99, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Ahern, D.T.; Bansal, P.; Faustino, I.; Kondaveeti, Y.; Glatt-Deeley, H.R.; Banda, E.C.; Pinter, S.F. Monosomy X in isogenic human iPSC-derived trophoblast model impacts expression modules preserved in human placenta. bioRxiv 2021. [Google Scholar] [CrossRef]
- Yabe, S.; Alexenko, A.P.; Amita, M.; Yang, Y.; Schust, D.J.; Sadovsky, Y.; Ezashi, T.; Roberts, R.M. Comparison of syncytiotrophoblast generated from human embryonic stem cells and from term placentas. Proc. Natl. Acad. Sci. USA 2016, 113, E2598–E2607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karvas, R.M.; McInturf, S.; Zhou, J.; Ezashi, T.; Schust, D.J.; Roberts, R.M.; Schulz, L.C. Use of a human embryonic stem cell model to discover GABRP, WFDC2, VTCN1 and ACTC1 as markers of early first trimester human trophoblast. Mol. Hum. Reprod. 2020, 26, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, N.; Kojima, J.; Fukuda, A.; Oda, M.; Kawasaki, T.; Ito, H.; Kuji, N.; Isaka, K.; Nishi, H.; Umezawa, A.; et al. Transcriptomic features of trophoblast lineage cells derived from human induced pluripotent stem cells treated with BMP 4. Placenta 2019, 89, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Ma, L.; Wang, Z.; Yu, Y.; Lye, S.J.; Shan, Y.; Wei, Y. Deciphering a distinct regulatory network of TEAD4, CDX2 and GATA3 in humans for trophoblast transition from embryonic stem cells. Biochim. Biophys. Acta 2020, 1867, 118736. [Google Scholar] [CrossRef] [PubMed]
- Shpiz, A.; Kalma, Y.; Frumkin, T.; Telias, M.; Carmon, A.; Amit, A.; Ben-Yosef, D. Human embryonic stem cells carrying an unbalanced translocation demonstrate impaired differentiation into trophoblasts: An in vitro model of human implantation failure. Mol. Hum. Reprod. 2014, 21, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Pidoux, G.; Gerbaud, P.; Cocquebert, M.; Segond, N.; Badet, J.; Fournier, T.; Guibourdenche, J.; Evain-Brion, D. Review: Human trophoblast fusion and differentiation: Lessons from trisomy 21 placenta. Placenta 2012, 33, S81–S86. [Google Scholar] [CrossRef]
- Canki, N.; Warburton, D.; Byrne, J. Morphological characteristics of mono-somy X in spontaneous abortions. Ann. Genet. 1988, 31, 4–13. [Google Scholar]
- Hook, E.B.; Warburton, D. Turner syndrome revisited: Review of new data supports the hypothesis that all viable 45,X cases are cryptic mosaics with a rescue cell line, implying an origin by mitotic loss. Qual. Life Res. 2014, 133, 417–424. [Google Scholar] [CrossRef]
- Shpiz, A.; Ben-Yosef, D.; Kalma, Y. Impaired function of trophoblast cells derived from translocated hESCs may explain pregnancy loss in women with balanced translocation (11;22). J. Assist. Reprod. Genet. 2016, 33, 1493–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rungsiwiwut, R.; Numchaisrika, P.; Ahnonkitpanit, V.; Virutamasen, P.; Pruksananonda, K. Triploid human embryonic stem cells derived from tripronuclear zygotes displayed pluripotency and trophoblast differentiation ability similar to the diploid human embryonic stem cells. J. Reprod. Dev. 2016, 62, 167–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baharvand, H.; Ashtiani, S.K.; Taee, A.; Massumi, M.; Valojerdi, M.R.; Eftekhari-Yazdi, P.; Moradi, S.Z.; Farrokhi, A. Generation of new human embryonic stem cell lines with diploid and triploid karyotypes. Dev. Growth Differ. 2006, 48, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, M.A.; Yang, Y.; Jain, A.; Lyons, A.S.; Yang, P.; Brahmasani, S.R.; Dai, A.; Tian, Y.; Ellersieck, M.R.; Tuteja, G.; et al. Early onset preeclampsia in a model for human placental trophoblast. Proc. Natl. Acad. Sci. USA 2019, 116, 4336–4345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horii, M.; Morey, R.; Bui, T.; Touma, O.; Nelson, K.K.; Cho, H.-Y.; Rishik, H.; Laurent, L.C.; Parast, M.M. Modeling preeclampsia using human induced pluripotent stem cells. Sci. Rep. 2021, 11, 5877. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E. Placental Oxidative Stress: From Miscarriage to Preeclampsia. J. Soc. Gynecol. Investig. 2004, 11, 342–352. [Google Scholar] [CrossRef]
- Gunne-Braden, A.; Sullivan, A.; Gharibi, B.; Sheriff, R.S.; Maity, A.; Wang, Y.-F.; Edwards, A.; Jiang, M.; Howell, M.; Goldstone, R.; et al. GATA3 Mediates a Fast, Irreversible Commitment to BMP4-Driven Differentiation in Human Embryonic Stem Cells. Cell Stem Cell 2020, 26, 693–706.e9. [Google Scholar] [CrossRef]
- Yang, M.; Rito, T.; Metzger, J.; Naftaly, J.; Soman, R.; Hu, J.; Albertini, D.F.; Barad, D.H.; Brivanlou, A.H.; Gleicher, N. Depletion of aneuploid cells in human embryos and gastruloids. Nat. Cell Biol. 2021, 23, 314–321. [Google Scholar] [CrossRef]
- Etoc, F.; Metzger, J.; Ruzo, A.; Kirst, C.; Yoney, A.; Ozair, M.Z.; Brivanlou, A.H.; Siggia, E.D. A Balance between Secreted Inhibitors and Edge Sensing Controls Gastruloid Self-Organization. Dev. Cell 2016, 39, 302–315. [Google Scholar] [CrossRef] [Green Version]
- Warmflash, A.; Sorre, B.; Etoc, F.; Siggia, E.D.; Brivanlou, A.H. A method to recapitulate early embryonic spatial patterning in human embryonic stem cells. Nat. Methods 2014, 11, 847–854. [Google Scholar] [CrossRef] [Green Version]
- Minn, K.T.; Fu, Y.C.; He, S.; Dietmann, S.; George, S.C.; Anastasio, M.A.; Morris, S.A.; Solnica-Krezel, L. High-resolution transcriptional and morphogenetic profiling of cells from micropatterned human ESC gastruloid cultures. eLife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Okae, H.; Toh, H.; Sato, T.; Hiura, H.; Takahashi, S.; Shirane, K.; Kabayama, Y.; Suyama, M.; Sasaki, H.; Arima, T. Derivation of Human Trophoblast Stem Cells. Cell Stem Cell 2018, 22, 50–63.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, C.; Beltcheva, M.; Gontarz, P.; Zhang, B.; Popli, P.; Fischer, L.A.; Khan, S.A.; Park, K.-M.; Yoon, E.-J.; Xing, X.; et al. Derivation of trophoblast stem cells from naïve human pluripotent stem cells. eLife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Tong, X.; Hao, Y.; Li, C.; Zhang, Y.; Pan, Y.; Dai, Y.; Liu, L.; Zhang, T.; Zhang, S. Abnormal Cullin1 neddylation-mediated p21 accumulation participates in the pathogenesis of recurrent spontaneous abortion by regulating trophoblast cell proliferation and differentiation. Mol. Hum. Reprod. 2020, 26, 327–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishioka, N.; Yamamoto, S.; Kiyonari, H.; Sato, H.; Sawada, A.; Ota, M.; Nakao, K.; Sasaki, H. Tead4 is required for specification of trophectoderm in pre-implantation mouse embryos. Mech. Dev. 2008, 125, 270–283. [Google Scholar] [CrossRef]
- Morey, R.; Farah, O.; Kallol, S.; Requena, D.F.; Meads, M.; Moretto-Zita, M.; Soncin, F.; Laurent, L.C.; Parast, M.M. Transcriptomic Drivers of Differentiation, Maturation, and Polyploidy in Human Extravillous Trophoblast. Front. Cell Dev. Biol. 2021, 9, 702046. [Google Scholar] [CrossRef]
- Cinkornpumin, J.K.; Kwon, S.Y.; Guo, Y.; Hossain, I.; Sirois, J.; Russett, C.S.; Tseng, H.-W.; Okae, H.; Arima, T.; Duchaine, T.F.; et al. Naive Human Embryonic Stem Cells Can Give Rise to Cells with a Trophoblast-like Transcriptome and Methylome. Stem Cell Rep. 2020, 15, 198–213. [Google Scholar] [CrossRef]
- Guo, G.; von Meyenn, F.; Rostovskaya, M.; Clarke, J.; Dietmann, S.; Baker, D.; Sahakyan, A.; Myers, S.; Bertone, P.; Reik, W.; et al. Epigenetic resetting of human pluripotency. Development 2017, 144, 2748–2763. [Google Scholar] [CrossRef] [Green Version]
- Hamada, H.; Okae, H.; Toh, H.; Chiba, H.; Hiura, H.; Shirane, K.; Sato, T.; Suyama, M.; Yaegashi, N.; Sasaki, H.; et al. Allele-Specific Methylome and Transcriptome Analysis Reveals Widespread Imprinting in the Human Placenta. Am. J. Hum. Genet. 2016, 99, 1045–1058. [Google Scholar] [CrossRef] [Green Version]
- Thamban, T.; Agarwaal, V.; Khosla, S. Role of genomic imprinting in mammalian development. J. Biosci. 2020, 45, 1–21. [Google Scholar] [CrossRef]
- Zhou, L.; Pu, Y.; Zhou, Y.; Wang, B.; Chen, Y.; Bai, Y.; He, S. Genome wide methylation analysis to uncover genes related to recurrent pregnancy loss. Genes Genom. 2021, 43, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Sazhenova, E.A.; Nikitina, T.V.; Vasilyev, S.A.; Tolmacheva, E.N.; Vasilyeva, O.Y.; Markov, A.V.; Yuryev, S.Y.; Skryabin, N.A.; Zarubin, A.A.; Kolesnikov, N.A.; et al. NLRP7 variants in spontaneous abortions with multilocus imprinting disturbances from women with recurrent pregnancy loss. J. Assist. Reprod. Genet. 2021, 38, 2893–2908. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.; Edsgärd, D.; Reinius, B.; Deng, Q.; Panula, S.P.; Codeluppi, S.; Reyes, A.P.; Linnarsson, S.; Sandberg, R.; Lanner, F. Single-Cell RNA-Seq Reveals Lineage and X Chromosome Dynamics in Human Preimplantation Embryos. Cell 2016, 165, 1012–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Nowak-Imialek, M.; Chen, X.; Chen, D.; Herrmann, D.; Ruan, D.; Chen, A.C.H.; Eckersley-Maslin, M.A.; Ahmad, S.; Lee, Y.L.; et al. Establishment of Porcine and Human Expanded Potential Stem Cells. Nat. Cell Biol. 2019, 21, 687–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Kurosawa, O.; Iwata, H. Establishment of human trophoblast stem cells from human induced pluripotent stem cell-derived cystic cells under micromesh culture. Stem Cell Res. Ther. 2019, 10, 245. [Google Scholar] [CrossRef]
- Castel, G.; Meistermann, D.; Bretin, B.; Firmin, J.; Blin, J.; Loubersac, S.; Bruneau, A.; Chevolleau, S.; Kilens, S.; Chariau, C.; et al. Induction of Human Trophoblast Stem Cells from Somatic Cells and Pluripotent Stem Cells. Cell Rep. 2020, 33, 108419. [Google Scholar] [CrossRef]
- Liu, X.; Ouyang, J.F.; Rossello, F.J.; Tan, J.P.; Davidson, K.C.; Valdes, D.S.; Schröder, J.; Sun, Y.B.Y.; Chen, J.; Knaupp, A.S.; et al. Reprogramming roadmap reveals route to human induced trophoblast stem cells. Nature 2020, 586, 101–107. [Google Scholar] [CrossRef]
- Benchetrit, H.; Herman, S.; van Wietmarschen, N.; Wu, T.; Makedonski, K.; Maoz, N.; Tov, N.Y.; Stave, D.; Lasry, R.; Zayat, V.; et al. Extensive Nuclear Reprogramming Underlies Lineage Conversion into Functional Trophoblast Stem-like Cells. Cell Stem Cell 2015, 17, 543–556. [Google Scholar] [CrossRef] [Green Version]
- Kubaczka, C.; Senner, C.; Cierlitza, M.; Araúzo-Bravo, M.J.; Kuckenberg, P.; Peitz, M.; Hemberger, M.; Schorle, H. Direct Induction of Trophoblast Stem Cells from Murine Fibroblasts. Cell Stem Cell 2015, 17, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Bai, T.; Peng, C.-Y.; Aneas, I.; Sakabe, N.; Requena, D.F.; Billstrand, C.; Nobrega, M.; Ober, C.; Parast, M.; Kessler, J.A. Establishment of human induced trophoblast stem-like cells from term villous cytotrophoblasts. Stem Cell Res. 2021, 56, 102507. [Google Scholar] [CrossRef]
- Haider, S.; Meinhardt, G.; Saleh, L.; Kunihs, V.; Gamperl, M.; Kaindl, U.; Ellinger, A.; Burkard, T.R.; Fiala, C.; Pollheimer, J.; et al. Self-Renewing Trophoblast Organoids Recapitulate the Developmental Program of the Early Human Placenta. Stem Cell Rep. 2018, 11, 537–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turco, M.Y.; Gardner, L.; Kay, R.G.; Hamilton, R.; Prater, M.; Hollinshead, M.S.; McWhinnie, A.; Esposito, L.; Fernando, R.; Skelton, H.; et al. Trophoblast organoids as a model for maternal–fetal interactions during human placentation. Nature 2018, 564, 263–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheridan, M.A.; Fernando, R.C.; Gardner, L.; Hollinshead, M.S.; Burton, G.J.; Moffett, A.; Turco, M.Y. Establishment and differentiation of long-term trophoblast organoid cultures from the human placenta. Nat. Protoc. 2020, 15, 3441–3463. [Google Scholar] [CrossRef] [PubMed]
- Karvas, R.M.; Khan, S.A.; Verma, S.; Yin, Y.; Kulkarni, D.; Dong, C.; Park, K.-M.; Chew, B.; Sane, E.; Fischer, L.A.; et al. Stem-cell-derived trophoblast organoids model human placental development and susceptibility to emerging pathogens. Cell Stem Cell 2022, 29, 810–825.e8. [Google Scholar] [CrossRef]
- Shannon, M.J.; Baltayeva, J.; Castellana, B.; Wächter, J.; McNeill, G.L.; Yoon, J.S.; Treissman, J.; Le, H.T.; Lavoie, P.M.; Beristain, A.G. Cell trajectory modeling identifies a primitive trophoblast state defined by BCAM enrichment. Development 2022, 149, dev199840. [Google Scholar] [CrossRef]
- Meinhardt, G.; Haider, S.; Kunihs, V.; Saleh, L.; Pollheimer, J.; Fiala, C.; Hetey, S.; Feher, Z.; Szilagyi, A.; Than, N.G.; et al. Pivotal role of the transcriptional co-activator YAP in trophoblast stemness of the developing human placenta. Proc. Natl. Acad. Sci. USA 2020, 117, 13562–13570. [Google Scholar] [CrossRef]
- Rivron, N.C.; Frias-Aldeguer, J.; Vrij, E.; Boisset, J.-C.; Korving, J.; Vivié, J.; Truckenmüller, R.; Van Oudenaarden, A.; Van Blitterswijk, C.A.; Geijsen, N. Blastocyst-like structures generated solely from stem cells. Nature 2018, 557, 106–111. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, T.; Chen, N.; Gao, D.; Shi, B.; Kong, S.; West, R.; Yuan, Y.; Zhi, M.; Wei, Q.; et al. Implantation initiation of self-assembled embryo-like structures generated using three types of mouse blastocyst-derived stem cells. Nat. Commun. 2019, 10, 496. [Google Scholar] [CrossRef]
- Sozen, B.; Cox, A.; De Jonghe, J.; Bao, M.; Hollfelder, F.; Glover, D.M.; Zernicka-Goetz, M. Self-Organization of Mouse Stem Cells into an Extended Potential Blastoid. Dev. Cell 2019, 51, 698–712.e8. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Zhong, C.; Yu, Y.; Liu, H.; Sakurai, M.; Yu, L.; Min, Z.; Shi, L.; Wei, Y.; Takahashi, Y.; et al. Generation of Blastocyst-like Structures from Mouse Embryonic and Adult Cell Cultures. Cell 2019, 179, 687–702.e18. [Google Scholar] [CrossRef]
- Yu, L.; Wei, Y.; Duan, J.; Schmitz, D.A.; Sakurai, M.; Wang, L.; Wang, K.; Zhao, S.; Hon, G.C.; Wu, J. Blastocyst-like structures generated from human pluripotent stem cells. Nature 2021, 591, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tan, J.P.; Schröder, J.; Aberkane, A.; Ouyang, J.F.; Mohenska, M.; Lim, S.M.; Sun, Y.B.Y.; Chen, J.; Sun, G.; et al. Modelling human blastocysts by reprogramming fibroblasts into iBlastoids. Nature 2021, 591, 627–632. [Google Scholar] [CrossRef]
- Yanagida, A.; Spindlow, D.; Nichols, J.; Dattani, A.; Smith, A.; Guo, G. Naive stem cell blastocyst model captures human embryo lineage segregation. Cell Stem Cell 2021, 28, 1016–1022.e4. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Min, Z.; Alsolami, S.; Ma, Z.; Zhang, E.; Chen, W.; Zhong, K.; Pei, W.; Kang, X.; Zhang, P.; et al. Generation of human blastocyst-like structures from pluripotent stem cells. Cell Discov. 2021, 7, 81. [Google Scholar] [CrossRef] [PubMed]
- Kagawa, H.; Javali, A.; Khoei, H.H.; Sommer, T.M.; Sestini, G.; Novatchkova, M.; Reimer, Y.S.O.; Castel, G.; Bruneau, A.; Maenhoudt, N.; et al. Human blastoids model blastocyst development and implantation. Nature 2021, 601, 600–605. [Google Scholar] [CrossRef]
- Sozen, B.; Jorgensen, V.; Weatherbee, B.A.T.; Chen, S.; Zhu, M.; Zernicka-Goetz, M. Reconstructing aspects of human embryogenesis with pluripotent stem cells. Nat. Commun. 2021, 12, 5550. [Google Scholar] [CrossRef]
- Rossant, J.; Tam, P.P. Opportunities and challenges with stem cell-based embryo models. Stem Cell Rep. 2021, 16, 1031–1038. [Google Scholar] [CrossRef]
- Deglincerti, A.; Croft, G.; Pietila, L.N.; Zernicka-Goetz, M.; Siggia, E.D.; Brivanlou, A.H. Self-organization of the in vitro attached human embryo. Nature 2016, 533, 251–254. [Google Scholar] [CrossRef]
- Shahbazi, M.N.; Jedrusik, A.; Vuoristo, S.; Recher, G.; Hupalowska, A.; Bolton, V.; Fogarty, N.; Campbell, A.; DeVito, L.G.; Ilic, D.; et al. Self-organization of the human embryo in the absence of maternal tissues. Nat. Cell Biol. 2016, 18, 700–708. [Google Scholar] [CrossRef]
- Xiang, L.; Yin, Y.; Zheng, Y.; Ma, Y.; Li, Y.; Zhao, Z.; Guo, J.; Ai, Z.; Niu, Y.; Duan, K.; et al. A developmental landscape of 3D-cultured human pre-gastrulation embryos. Nature 2019, 577, 537–542. [Google Scholar] [CrossRef]
- Zhou, F.; Wang, R.; Yuan, P.; Ren, Y.; Mao, Y.; Li, R.; Lian, Y.; Li, J.; Wen, L.; Yan, L.; et al. Reconstituting the transcriptome and DNA methylome landscapes of human implantation. Nature 2019, 572, 660–664. [Google Scholar] [CrossRef] [PubMed]
- West, R.C.; Ming, H.; Logsdon, D.M.; Sun, J.; Rajput, S.K.; Kile, R.A.; Schoolcraft, W.B.; Roberts, R.M.; Krisher, R.L.; Jiang, Z.; et al. Dynamics of trophoblast differentiation in peri-implantation–stage human embryos. Proc. Natl. Acad. Sci. USA 2019, 116, 22635–22644. [Google Scholar] [CrossRef]
- Molè, M.A.; Coorens, T.H.H.; Shahbazi, M.N.; Weberling, A.; Weatherbee, B.A.T.; Gantner, C.W.; Sancho-Serra, C.; Richardson, L.; Drinkwater, A.; Syed, N.; et al. A single cell characterisation of human embryogenesis identifies pluripotency transitions and putative anterior hypoblast centre. Nat. Commun. 2021, 12, 3679. [Google Scholar] [CrossRef] [PubMed]
- Popovic, M.; Dhaenens, L.; Taelman, J.; Dheedene, A.; Bialecka, M.; De Sutter, P.; Lopes, S.M.C.D.S.; Menten, B.; Heindryckx, B. Extended in vitro culture of human embryos demonstrates the complex nature of diagnosing chromosomal mosaicism from a single trophectoderm biopsy. Hum. Reprod. 2019, 34, 758–769. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, M.N.; Wang, T.; Tao, X.; Weatherbee, B.A.T.; Sun, L.; Zhan, Y.; Keller, L.; Smith, G.D.; Pellicer, A.; Scott, R.T., Jr.; et al. Developmental potential of aneuploid human embryos cultured beyond implantation. Nat. Commun. 2020, 11, 3987. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y.; Zhao, N.; Liu, P.; Huang, J. Variations in chromosomal aneuploidy rates in IVF blastocysts and early spontaneous abortion chorionic villi. J. Assist. Reprod. Genet. 2020, 37, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Redline, R.W.; Zaragoza, M.; Hassold, T. Prevalence of developmental and inflammatory lesions in nonmolar first-trimester spontaneous abortions. Hum. Pathol. 1999, 30, 93–100. [Google Scholar] [CrossRef]
- Roost, M.S.; Slieker, R.C.; Bialecka, M.; Van Iperen, L.; Fernandes, M.M.G.; He, N.; Suchiman, H.E.D.; Szuhai, K.; Carlotti, F.; De Koning, E.J.P.; et al. DNA methylation and transcriptional trajectories during human development and reprogramming of isogenic pluripotent stem cells. Nat. Commun. 2017, 8, 908. [Google Scholar] [CrossRef]
- Wang, L.-J.; Chen, C.-P.; Lee, Y.-S.; Ng, P.-S.; Chang, G.-D.; Pao, Y.-H.; Lo, H.-F.; Peng, C.-H.; Cheong, M.-L.; Chen, H. Functional antagonism between ΔNp63α and GCM1 regulates human trophoblast stemness and differentiation. Nat. Commun. 2022, 13, 1626. [Google Scholar] [CrossRef]
- Takahashi, S.; Okae, H.; Kobayashi, N.; Kitamura, A.; Kumada, K.; Yaegashi, N.; Arima, T. Loss of p57 KIP2 expression confers resistance to contact inhibition in human androgenetic trophoblast stem cells. Proc. Natl. Acad. Sci. USA 2019, 116, 26606–26613. [Google Scholar] [CrossRef] [Green Version]
- Hornbachner, R.; Lackner, A.; Papuchova, H.; Haider, S.; Knöfler, M.; Mechtler, K.; Latos, P.A. MSX2 safeguards syncytiotrophoblast fate of human trophoblast stem cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2105130118. [Google Scholar] [CrossRef] [PubMed]
- Goddijn, M.; Leschot, N. Genetic aspects of miscarriage. Best Pr. Res. Clin. Obstet. Gynaecol. 2000, 14, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, N.; Ogawa, K.; Sasaki, A.; Mitsui, M.; Wada, S.; Sago, H. Maternal age, history of miscarriage, and embryonic/fetal size are associated with cytogenetic results of spontaneous early miscarriages. J. Assist. Reprod. Genet. 2019, 36, 749–757. [Google Scholar] [CrossRef]
- Rubio, C.; Simón, C.; Vidal, F.; Rodrigo, L.; Pehlivan, T.; Remohi, J.; Pellicer, A. Chromosomal abnormalities and embryo development in recurrent miscarriage couples. Hum. Reprod. 2003, 18, 182–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanneste, E.; Voet, T.; Le Caignec, C.; Ampe, M.; Konings, P.; Melotte, C.; Debrock, S.; Amyere, M.; Vikkula, M.; Schuit, F.; et al. Chromosome instability is common in human cleavage-stage embryos. Nat. Med. 2009, 15, 577–583. [Google Scholar] [CrossRef]
- Minasi, M.G.; Colasante, A.; Riccio, T.; Ruberti, A.; Casciani, V.; Scarselli, F.; Spinella, F.; Fiorentino, F.; Varricchio, M.T.; Greco, E. Correlation between aneuploidy, standard morphology evaluation and morphokinetic development in 1730 biopsied blastocysts: A consecutive case series study. Hum. Reprod. 2016, 31, 2245–2254. [Google Scholar] [CrossRef] [Green Version]
- McCoy, R.C.; Demko, Z.P.; Ryan, A.; Banjevic, M.; Hill, M.; Sigurjonsson, S.; Rabinowitz, M.; Petrov, D.A. Evidence of Selection against Complex Mitotic-Origin Aneuploidy during Preimplantation Development. PLoS Genet. 2015, 11, e1005601. [Google Scholar] [CrossRef] [Green Version]
- Fragouli, E.; Munne, S.; Wells, D. The cytogenetic constitution of human blastocysts: Insights from comprehensive chromosome screening strategies. Hum. Reprod. Updat. 2018, 25, 15–33. [Google Scholar] [CrossRef]
- Kort, J.D.; McCoy, R.C.; Demko, Z.; Lathi, R.B. Are blastocyst aneuploidy rates different between fertile and infertile populations? J. Assist. Reprod. Genet. 2017, 35, 403–408. [Google Scholar] [CrossRef]
- FitzPatrick, D.R. Transcriptional consequences of autosomal trisomy: Primary gene dosage with complex downstream effects. Trends Genet. 2005, 21, 249–253. [Google Scholar] [CrossRef]
- Sheltzer, J.M.; Amon, A. The aneuploidy paradox: Costs and benefits of an incorrect karyotype. Trends Genet. 2011, 27, 446–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.-J.; Nelliat, A.R.; Choudhury, M.I.; Kucharavy, A.; Bradford, W.D.; Cook, M.E.; Kim, J.; Mair, D.B.; Sun, S.X.; Schatz, M.C.; et al. Hypo-osmotic-like stress underlies general cellular defects of aneuploidy. Nature 2019, 570, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Ribas, I.; Gimeno, P.D.; Sebastian-Leon, P.; Mercader, A.; Quiñonero, A.; Ballesteros, A.; Pellicer, A.; Dominguez, F. Transcriptomic behavior of genes associated with chromosome 21 aneuploidies in early embryo development. Fertil. Steril. 2019, 111, 991–1001.e2. [Google Scholar] [CrossRef]
- Licciardi, F.; Lhakhang, T.; Kramer, Y.G.; Zhang, Y.; Heguy, A.; Tsirigos, A. Human blastocysts of normal and abnormal karyotypes display distinct transcriptome profiles. Sci. Rep. 2018, 8, 14906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruhn, J.R.; Zielinska, A.P.; Shukla, V.; Blanshard, R.; Capalbo, A.; Cimadomo, D.; Nikiforov, D.; Chan, A.C.-H.; Newnham, L.J.; Vogel, I.; et al. Chromosome errors in human eggs shape natural fertility over reproductive life span. Science 2019, 365, 1466–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tšuiko, O.; Vanneste, M.; Melotte, C.; Ding, J.; Debrock, S.; Masset, H.; Peters, M.; Salumets, A.; De Leener, A.; Pirard, C.; et al. Haplotyping-based preimplantation genetic testing reveals parent-of-origin specific mechanisms of aneuploidy formation. npj Genom. Med. 2021, 6, 81. [Google Scholar] [CrossRef] [PubMed]
- Weizman, N.F.; Wyse, B.A.; Antes, R.; Ibarrientos, Z.; Sangaralingam, M.; Motamedi, G.; Kuznyetsov, V.; Madjunkova, S.; Librach, C.L. Towards Improving Embryo Prioritization: Parallel Next Generation Sequencing of DNA and RNA from a Single Trophectoderm Biopsy. Sci. Rep. 2019, 9, 2853. [Google Scholar] [CrossRef]
- Groff, A.F.; Resetkova, N.; DiDomenico, F.; Sakkas, D.; Penzias, A.; Rinn, J.L.; Eggan, K. RNA-seq as a tool for evaluating human embryo competence. Genome Res. 2019, 29, 1705–1718. [Google Scholar] [CrossRef] [Green Version]
- Katz-Jaffe, M.; McReynolds, S.; De Klerk, K.; Henry, L.; Schweitz, M.; Swain, J.; Schoolcraft, W. Extremely low incidence of mosaicism in human blastocysts mimics occurrence in natural and IVF clincial pregnancies. Fertil. Steril. 2017, 108, e87–e88. [Google Scholar] [CrossRef] [Green Version]
- Ruttanajit, T.; Chanchamroen, S.; Cram, D.S.; Sawakwongpra, K.; Suksalak, W.; Leng, X.; Fan, J.; Wang, L.; Yao, Y.; Quangkananurug, W. Detection and quan-titation of chromosomal mosaicism in human blastocysts using copy number variation sequencing. Prenat. Diagn. 2016, 36, 154–162. [Google Scholar] [CrossRef]
- Taylor, T.H.; Gitlin, S.A.; Patrick, J.L.; Crain, J.L.; Wilson, J.M.; Griffin, D.K. The origin, mechanisms, incidence and clinical consequences of chromosomal mosaicism in humans. Hum. Reprod. Updat. 2014, 20, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Van Echten-Arends, J.; Mastenbroek, S.; Sikkema-Raddatz, B.; Korevaar, J.C.; Heineman, M.J.; van der Veen, F.; Repping, S. Chromosomal mosaicism in human preimplantation embryos: A systematic review. Hum. Reprod. Updat. 2011, 17, 620–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCoy, R.C. Mosaicism in Preimplantation Human Embryos: When Chromosomal Abnormalities Are the Norm. Trends Genet. 2017, 33, 448–463. [Google Scholar] [CrossRef] [PubMed]
- Starostik, M.R.; Sosina, O.A.; McCoy, R.C. Single-cell analysis of human embryos reveals diverse patterns of aneuploidy and mosaicism. Genome Res. 2020, 30, 814–825. [Google Scholar] [CrossRef]
- Coorens, T.H.H.; Oliver, T.R.W.; Sanghvi, R.; Sovio, U.; Cook, E.; Vento-Tormo, R.; Haniffa, M.; Young, M.D.; Rahbari, R.; Sebire, N.; et al. Inherent mosaicism and extensive mutation of human placentas. Nature 2021, 592, 80–85. [Google Scholar] [CrossRef]
- Daughtry, B.L.; Rosenkrantz, J.L.; Lazar, N.H.; Fei, S.S.; Redmayne, N.; Torkenczy, K.A.; Adey, A.; Yan, M.; Gao, L.; Park, B.; et al. Single-cell sequencing of primate preimplantation embryos reveals chromosome elimination via cellular fragmentation and blastomere exclusion. Genome Res. 2019, 29, 367–382. [Google Scholar] [CrossRef] [Green Version]
- Orvieto, R.; Shimon, C.; Rienstein, S.; Jonish-Grossman, A.; Shani, H.; Aizer, A. Do human embryos have the ability of self-correction? Reprod. Biol. Endocrinol. 2020, 18, 1–6. [Google Scholar] [CrossRef]
- Victor, A.R.; Griffin, D.K.; Brake, A.J.; Tyndall, J.C.; Murphy, A.E.; Lepkowsky, L.T.; Lal, A.; Zouves, C.G.; Barnes, F.L.; McCoy, R.; et al. Assessment of aneuploidy concordance between clinical trophectoderm biopsy and blastocyst. Hum. Reprod. 2018, 34, 181–192. [Google Scholar] [CrossRef]
- Zhigalina, D.; Skryabin, N.; Kanbekova, O.; Artyukhova, V.; Svetlakov, A.; Lebedev, I. Correction of the embryonic karyotype at the preimplantation stage of human development. Med. Genet. 2020, 19, 49–50. [Google Scholar]
- Bolton, H.; Graham, S.; Van Der Aa, N.; Kumar, P.; Theunis, K.; Gallardo, E.F.; Voet, T.; Zernicka-Goetz, M. Mouse model of chromosome mosaicism reveals lineage-specific depletion of aneuploid cells and normal developmental potential. Nat. Commun. 2016, 7, 11165. [Google Scholar] [CrossRef] [Green Version]
- Singla, S.; Iwamoto-Stohl, L.K.; Zhu, M.; Zernicka-Goetz, M. Autophagy-mediated apoptosis eliminates aneuploid cells in a mouse model of chromosome mosaicism. Nat. Commun. 2020, 11, 2072–2084. [Google Scholar] [CrossRef] [PubMed]
- Heyer, B.S.; MacAuley, A.; Behrendtsen, O.; Werb, Z. Hypersensitivity to DNA damage leads to increased apoptosis during early mouse development. Genes Dev. 2000, 14, 2072–2084. [Google Scholar] [CrossRef] [PubMed]
- Laurent, A.; Blasi, F. Differential DNA damage signalling and apoptotic threshold correlate with mouse epiblast-specific hypersensitivity to radiation. Development 2015, 142, dev.125708-85. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Barbaric, I. Fitness selection in human pluripotent stem cells and interspecies chimeras: Implications for human development and regenerative medicine. Dev. Biol. 2021, 476, 209–217. [Google Scholar] [CrossRef]
- Home, P.; Kumar, R.P.; Ganguly, A.; Saha, B.; Milano-Foster, J.; Bhattacharya, B.; Ray, S.; Gunewardena, S.; Paul, A.; Camper, S.A.; et al. Genetic redundancy of GATA factors in extraembryonic trophoblast lineage ensures progression of both pre and postimplantation mammalian development. Development 2017, 144, 876–888. [Google Scholar] [CrossRef] [Green Version]
- Lebedev, I.N.; Zhigalina, D.I. From contemplation to classification of chromosomal mosaicism in human preimplantation embryos. J. Assist. Reprod. Genet. 2021, 38, 2833–2848. [Google Scholar] [CrossRef]
- Coticchio, G.; Barrie, A.; Lagalla, C.; Borini, A.; Fishel, S.; Griffin, D.; Campbell, A. Plasticity of the human preimplantation embryo: Developmental dogmas, variations on themes and self-correction. Hum. Reprod. Updat. 2021, 27, 848–865. [Google Scholar] [CrossRef]
- Kalousek, D.; Dill, F.J. Chromosomal Mosaicism Confined to the Placenta in Human Conceptions. Science 1983, 221, 665–667. [Google Scholar] [CrossRef]
- Kalousek, D.K.; Vekemans, M.; Du Montcel, S.T.; Mendizabai, H.; Aymé, S.; Lévy, A.; Philip, N. Confined placental mosaicism. J. Med. Genet. 1996, 33, 529–533. [Google Scholar] [CrossRef]
- Grati, F.R.; Malvestiti, F.; Branca, L.; Agrati, C.; Maggi, F.; Simoni, G. Chromosomal mosaicism in the fetoplacental unit. Best Pr. Res. Clin. Obstet. Gynaecol. 2017, 42, 39–52. [Google Scholar] [CrossRef]
- Amor, D.J.; Neo, W.T.; Waters, E.; Heussler, H.; Pertile, M.; Halliday, J. Health and developmental outcome of children following prenatal diagnosis of confined placental mosaicism. Prenat. Diagn. 2006, 26, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Baffero, G.M.; Somigliana, E.; Crovetto, F.; Paffoni, A.; Persico, N.; Guerneri, S.; Lalatta, F.; Fogliani, R.; Fedele, L. Confined placental mosaicism at chorionic villous sampling: Risk factors and pregnancy outcome. Prenat. Diagn. 2012, 32, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Robinson, W.P.; Barrett, I.J.; Bernard, L.; Telenius, A.; Bernasconi, F.; Wilson, R.D.; Best, R.G.; Howard-Peebles, P.N.; Langlois, S.; Kalousek, D.K. Meiotic origin of trisomy in confined placental mosaicism is correlated with presence of fetal uniparental disomy, high levels of trisomy in trophoblast, and increased risk of fetal intrauterine growth restriction. Am. J. Hum. Genet. 1997, 60, 917–927. [Google Scholar]
- Toutain, J.; Goutte-Gattat, D.; Horovitz, J.; Saura, R. Confined placental mosaicism revisited: Impact on pregnancy characteristics and outcome. PLoS ONE 2018, 13, e0195905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Gobbo, G.F.; Yin, Y.; Choufani, S.; Butcher, E.A.; Wei, J.; Rajcan-Separovic, E.; Bos, H.; von Dadelszen, P.; Weksberg, R.; Robinson, W.P.; et al. Genomic imbalances in the placenta are associated with poor fetal growth. Mol. Med. 2021, 27, 3. [Google Scholar] [CrossRef]
- Eggenhuizen, G.M.; Go, A.; Koster, M.P.H.; Baart, E.B.; Galjaard, R.J. Confined placental mosaicism and the association with pregnancy outcome and fetal growth: A review of the literature. Hum. Reprod. Updat. 2021, 27, 885–903. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Duncan, A.W.; Taylor, M.H.; Hickey, R.D.; Newell, A.E.H.; Lenzi, M.L.; Olson, S.B.; Finegold, M.J.; Grompe, M. The ploidy conveyor of mature hepatocytes as a source of genetic variation. Nature 2010, 467, 707–710. [Google Scholar] [CrossRef] [Green Version]
- Weier, J.F.; Weier, H.-U.G.; Jung, C.J.; Gormley, M.; Zhou, Y.; Chu, L.W.; Genbacev, O.; Wright, A.A.; Fisher, S.J. Human cytotrophoblasts acquire aneuploidies as they differentiate to an invasive phenotype. Dev. Biol. 2005, 279, 420–432. [Google Scholar] [CrossRef] [Green Version]
- Kasak, L.; Rull, K.; Vaas, P.; Teesalu, P.; Laan, M. Extensive load of somatic CNVs in the human placenta. Sci. Rep. 2015, 5, 8342. [Google Scholar] [CrossRef] [Green Version]
- Esteki, M.Z.; Viltrop, T.; Tsuiko, O.; Tiirats, A.; Koel, M.; Nõukas, M.; Žilina, O.; Teearu, K.; Marjonen, H.; Kahila, H.; et al. In vitro fertilization does not increase the incidence of de novo copy number alterations in fetal and placental lineages. Nat. Med. 2019, 25, 1699–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grati, F.R.; Gallazzi, G.; Branca, L.; Maggi, F.; Simoni, G.; Yaron, Y. An evidence-based scoring system for prioritizing mosaic aneuploid embryos following preimplantation genetic screening. Reprod. Biomed. Online 2018, 36, 442–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donato, X.-C.; Brechard, M.-P.; François-Renard, P.; Hairion, D.; Quarello, E.; Hoffet, M.; Katsogiannou, M.; Desbriere, R. Pregnancy course and outcomes in mosaic trisomy 16 confined to the placenta: A case series. Prenat. Diagn. 2018, 38, 924–927. [Google Scholar] [CrossRef] [PubMed]
- Tolmacheva, E.N.; Vasilyev, S.A.; Nikitina, T.V.; Lytkina, E.S.; Sazhenova, E.A.; Zhigalina, D.I.; Vasilyeva, O.Y.; Markov, A.V.; Demeneva, V.V.; Tashireva, L.A.; et al. Identification of differentially methylated genes in first-trimester placentas with trisomy. Sci. Rep. 2022, 12, 1166. [Google Scholar] [CrossRef]
- Van der Meij, K.R.; Sistermans, E.A.; Macville, M.V.; Stevens, S.J.; Bax, C.J.; Bekker, M.N.; Bilardo, C.M.; Boon, E.M.; Boter, M.; Diderich, K.E.; et al. TRIDENT-2: National Implementation of Genome-wide Non-invasive Prenatal Testing as a First-Tier Screening Test in the Netherlands. Am. J. Hum. Genet. 2019, 105, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Bogaert, K.V.D.; Lannoo, L.; Brison, N.; Gatinois, V.; Baetens, M.; Blaumeiser, B.; Boemer, F.; Bourlard, L.; Bours, V.; De Leener, A.; et al. Outcome of publicly funded nationwide first-tier noninvasive prenatal screening. Genet. Med. 2021, 23, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Benn, P. Trisomy 16 and trisomy 16 Mosaicism: A review. Am. J. Med. Genet. 1998, 79, 121–133. [Google Scholar] [CrossRef]
- Maxwell, S.M.; Colls, P.; Hodes-Wertz, B.; McCulloh, D.H.; McCaffrey, C.; Wells, D.; Munné, S.; Grifo, J.A. Why do euploid embryos miscarry? A case-control study comparing the rate of aneuploidy within presumed euploid embryos that resulted in miscarriage or live birth using next-generation sequencing. Fertil. Steril. 2016, 106, 1414–1419.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Victor, A.R.; Tyndall, J.C.; Brake, A.J.; Lepkowsky, L.T.; Murphy, A.E.; Griffin, D.K.; McCoy, R.C.; Barnes, F.L.; Zouves, C.G.; Viotti, M. One hundred mosaic embryos transferred prospectively in a single clinic: Exploring when and why they result in healthy pregnancies. Fertil. Steril. 2019, 111, 280–293. [Google Scholar] [CrossRef]
- Munné, S.; Spinella, F.; Grifo, J.; Zhang, J.; Beltran, M.P.; Fragouli, E.; Fiorentino, F. Clinical outcomes after the transfer of blastocysts characterized as mosaic by high resolution Next Generation Sequencing- further insights. Eur. J. Med Genet. 2020, 63, 103741. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Chen, J.J.; Nabu, S.; Yeung, Q.S.Y.; Li, Y.; Tan, J.H.; Suksalak, W.; Chanchamroen, S.; Quangkananurug, W.; Wong, P.S.; et al. The Pregnancy Outcome of Mosaic Embryo Transfer: A Prospective Multicenter Study and Meta-Analysis. Genes 2020, 11, 973. [Google Scholar] [CrossRef] [PubMed]
- Bower, O.J.; McCarthy, A.; Lea, R.A.; Alanis-Lobato, G.; Zohren, J.; Gerri, C.; Turner, J.M.; Niakan, K.K. Generating CRISPR-Cas9-Mediated Null Mutations and Screening Targeting Efficiency in Human Pluripotent Stem Cells. Curr. Protoc. 2021, 1, e232. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, N.; McCarthy, A.; Snijders, K.E.; Powell, B.E.; Kubikova, N.; Blakeley, P.; Lea, R.; Elder, K.; Wamaitha, S.; Kim, D.; et al. Genome editing reveals a role for OCT4 in human embryogenesis. Nature 2017, 550, 67–73. [Google Scholar] [CrossRef]
- Zuccaro, M.V.; Xu, J.; Mitchell, C.; Marin, D.; Zimmerman, R.; Rana, B.; Weinstein, E.; King, R.T.; Palmerola, K.L.; Smith, M.E.; et al. Allele-Specific Chromosome Removal after Cas9 Cleavage in Human Embryos. Cell 2020, 183, 1650–1664.e15. [Google Scholar] [CrossRef] [PubMed]
- Alanis-Lobato, G.; Zohren, J.; McCarthy, A.; Fogarty, N.M.E.; Kubikova, N.; Hardman, E.; Greco, M.; Wells, D.; Turner, J.M.A.; Niakan, K.K. Frequent loss of heterozygosity in CRISPR-Cas9–edited early human embryos. Proc. Natl. Acad. Sci. USA 2021, 118, e2004832117. [Google Scholar] [CrossRef]
- Bershteyn, M.; Hayashi, Y.; Desachy, G.; Hsiao, E.C.; Sami, S.; Tsang, K.M.; Weiss, L.A.; Kriegstein, A.R.; Yamanaka, S.; Wynshaw-Boris, A. Cell-autonomous correction of ring chromosomes in human induced pluripotent stem cells. Nature 2014, 507, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Hirota, T.; Ohta, H.; Powell, B.E.; Mahadevaiah, S.K.; Ojarikre, O.A.; Saitou, M.; Turner, J.M.A. Fertile offspring from sterile sex chromosome trisomic mice. Science 2017, 357, 932–935. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Fan, X.; Wang, R.; Lu, X.; Dang, Y.-L.; Wang, H.; Lin, H.-Y.; Zhu, C.; Ge, H.; Cross, J.C.; et al. Single-cell RNA-seq reveals the diversity of trophoblast subtypes and patterns of differentiation in the human placenta. Cell Res. 2018, 28, 819–832. [Google Scholar] [CrossRef] [Green Version]
- Vento-Tormo, R.; Efremova, M.; Botting, R.A.; Turco, M.Y.; Vento-Tormo, M.; Meyer, K.B.; Park, J.-E.; Stephenson, E.; Polański, K.; Goncalves, A.; et al. Single-cell reconstruction of the early maternal–fetal interface in humans. Nature 2018, 563, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Turco, M.Y.; Gardner, L.; Hughes, J.; Cindrova-Davies, T.; Gomez, M.J.; Farrell, L.; Hollinshead, M.; Marsh, S.G.E.; Brosens, J.J.; Critchley, H.O.; et al. Long-term, hormone-responsive organoid cultures of human endometrium in a chemically defined medium. Nat. Cell Biol. 2017, 19, 568–577. [Google Scholar] [CrossRef]
- Abbas, Y.; Turco, M.Y.; Burton, G.J.; Moffett, A. Investigation of human trophoblast invasion in vitro. Hum. Reprod. Update 2020, 26, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, H.C.; Dhakal, P.; Behura, S.K.; Schust, D.J.; Spencer, T.E. Self-renewing endometrial epithelial organoids of the human uterus. Proc. Natl. Acad. Sci. USA 2019, 116, 23132–23142. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Megli, C.; Coyne, C.B. Innate immune signaling in trophoblast and decidua organoids defines differential antiviral defenses at the maternal-fetal in-terface. bioRxiv 2021. [Google Scholar] [CrossRef]


{kind=link}
| Stem Cell-Based Model | Cell Source | Advantages | Limitations | Key Achievements of Model Application for Genetics of Miscarriage or Trophoblast Dysfunction Pathology |
|---|---|---|---|---|
| BMP4-induced trophoblast-like cells | ESCs [93,94]; iPSCs [95]; PSCs [96] | Availability of source material; possibility of genetic modification; variety of genetic backgrounds | Lacking self-renewal; expression does not fully correspond to trophoblasts | Trisomy 21: delay in the induction of the trophoblast lineage, lower fusion index, increased hCG secretion [96] |
| Monosomy X: altered levels of placental genes; skewed trophoblast cell type composition [103] | ||||
| Translocation (11;22): impaired expression of trophoblastic genes; lower invasiveness [108,112] | ||||
| Preeclampsia: reduced trophoblast invasion under high O2 conditions [115,116] | ||||
| Recurrent complete hydatidiform mole: precocious downregulation of pluripotency factors and activation of trophoblast lineage markers [84] | ||||
| Lineage-specific behavior of aneuploid cells in “gastruloids” with prevalent survival of extra-embryonic tissue [119] | ||||
| “True” human trophoblast stem cells (hTSCs) | Blastocysts; 1st trimester placentae [123]; term placenta [170] | Patient-specific; preserved characteristics of trophoblastic cells, unlimited self-renewing capacity | Derivation from the primary tissues of some abortions may be problematic | Association of idiopathic RPL with altered expression of TEAD4; imbalance of self-renewal vs. differentiation in idiopathic RPL placentae [8] |
| Complete hydatidiform mole exhibits resistance to contact inhibition [171] | ||||
| Neddylation inhibition hindered EVT differentiation [125] | ||||
| Depletion of MSX2 resulted in precocious STB differentiation [172] | ||||
| Decreased CKMT11 expression associated with PE [170] | ||||
| Trophoblast stem-like cells (TSLCs) | Naïve PSCs [97,98,124,128]; primed PSCs [89,90,100,136]; EPSCs [135] | Availability of source material; possibility of genetic modification; variety of genetic backgrounds | Possible inconsistencies of epigenetic specificity | No data currently available |
| Induced trophoblast stem cells (iTSCs) | Somatic cells [137,138]; term placenta vCTBs [141] | |||
| 3D culture models | 1st trimester CTBs [142,143,144] | More authentic tissue environments; self-renewing ability | Limited self-renewal in some studies | Treatment with YAP/TAZ inhibitor reduced organoid growth and expression of cyclin A [147] |
| hTSCs [8] | Inefficient organoid formation for RPL cases with reduced expression of TEAD4 [8] | |||
| Transcriptomic landscape for CTB commitment to EVT or SCT; BCAM is a primitive progenitor marker [146] | ||||
| Naïve PSCs [145] | Dynamics of X chromosome inactivation [145] | |||
| Blastoids | Naïve PSCs [152,154,156] | A large number of units in one experiment; availability of source material; possibility of genetic modification; variety of genetic backgrounds | Expression discrepancy with blastocysts in some models | No data currently available |
| hEPSCs [155,157] | ||||
| iBlastoids | Fibroblasts [153] | |||
| Extended blastocyst culture | Blastocysts [159,160] | Patient-specific; preserved native characteristics of cells; autonomy from maternal contribution | Limited access to source | Lethality of autosomal monosomies and multiple aberrations at the peri-implantation period; embryos with duplications develop longer than with deletions [165,166] |
| 3D extended blastocyst culture | Blastocysts [161] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikitina, T.V.; Lebedev, I.N. Stem Cell-Based Trophoblast Models to Unravel the Genetic Causes of Human Miscarriages. Cells 2022, 11, 1923. https://doi.org/10.3390/cells11121923
Nikitina TV, Lebedev IN. Stem Cell-Based Trophoblast Models to Unravel the Genetic Causes of Human Miscarriages. Cells. 2022; 11(12):1923. https://doi.org/10.3390/cells11121923
Chicago/Turabian StyleNikitina, Tatiana V., and Igor N. Lebedev. 2022. "Stem Cell-Based Trophoblast Models to Unravel the Genetic Causes of Human Miscarriages" Cells 11, no. 12: 1923. https://doi.org/10.3390/cells11121923
APA StyleNikitina, T. V., & Lebedev, I. N. (2022). Stem Cell-Based Trophoblast Models to Unravel the Genetic Causes of Human Miscarriages. Cells, 11(12), 1923. https://doi.org/10.3390/cells11121923