
Heparanase Expression Propagates Liver Damage in CCL4-Induced Mouse Model


Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. CCL4 Induced Liver Damage
2.3. Analysis of Plasma
2.4. Morphological and Histological Analyses
2.5. Tunel Assay
2.6. Western Blotting Analysis
2.7. Isolation and Analysis of Hepatocytes
2.8. Isolation and Characterization of Hepatic Stellate Cells (HSC)
3. Results
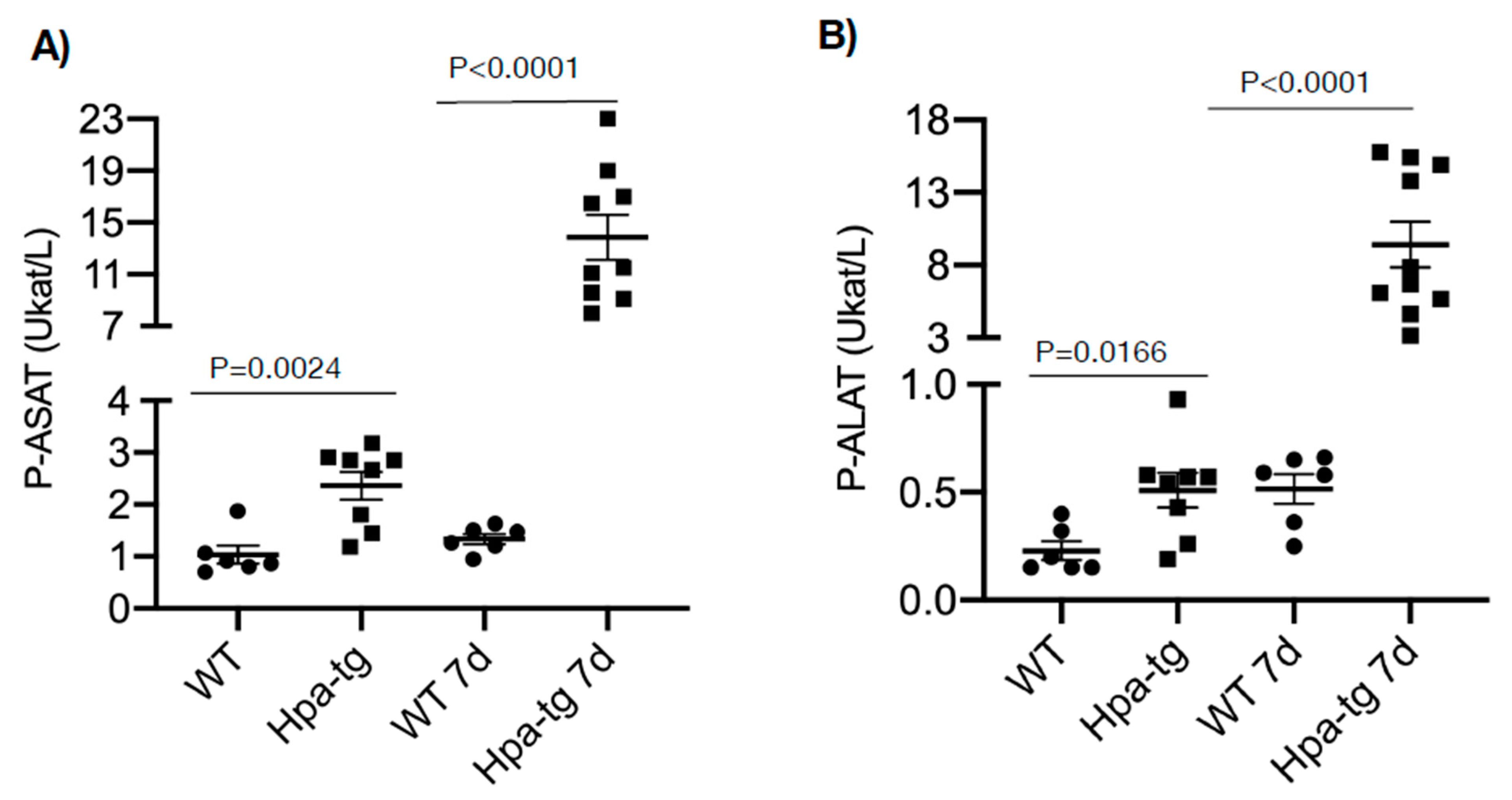
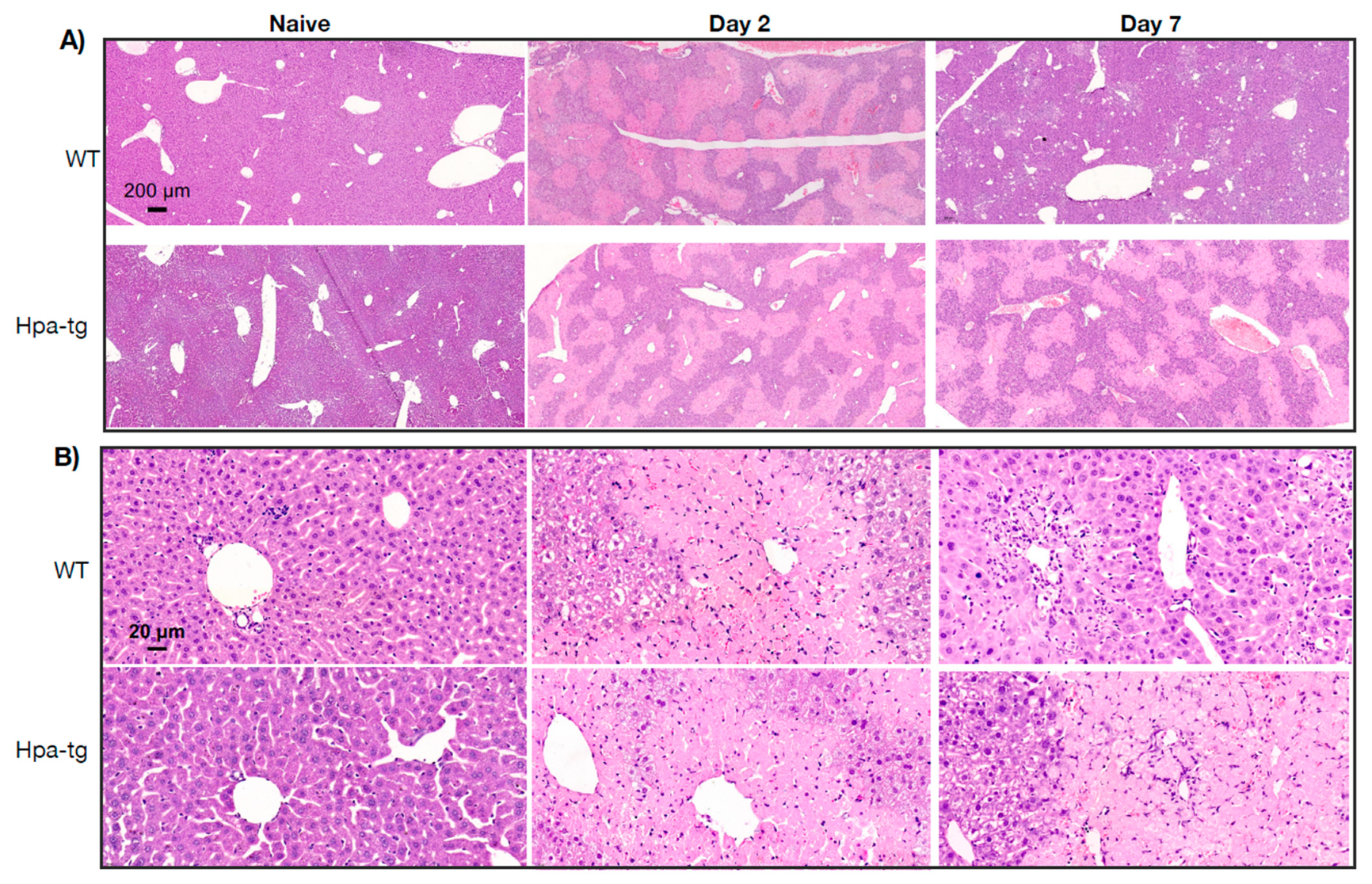
3.1. Enhanced Damage of Hpa-Tg Liver in Response to CCL4
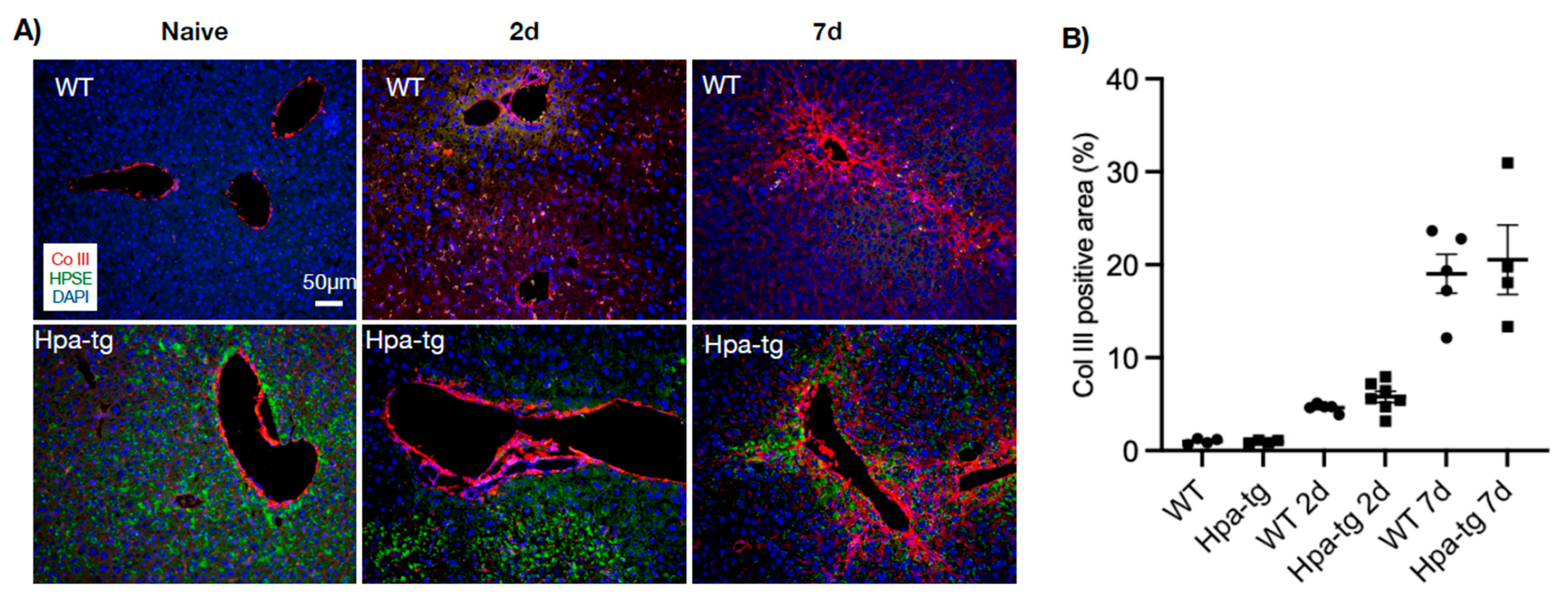
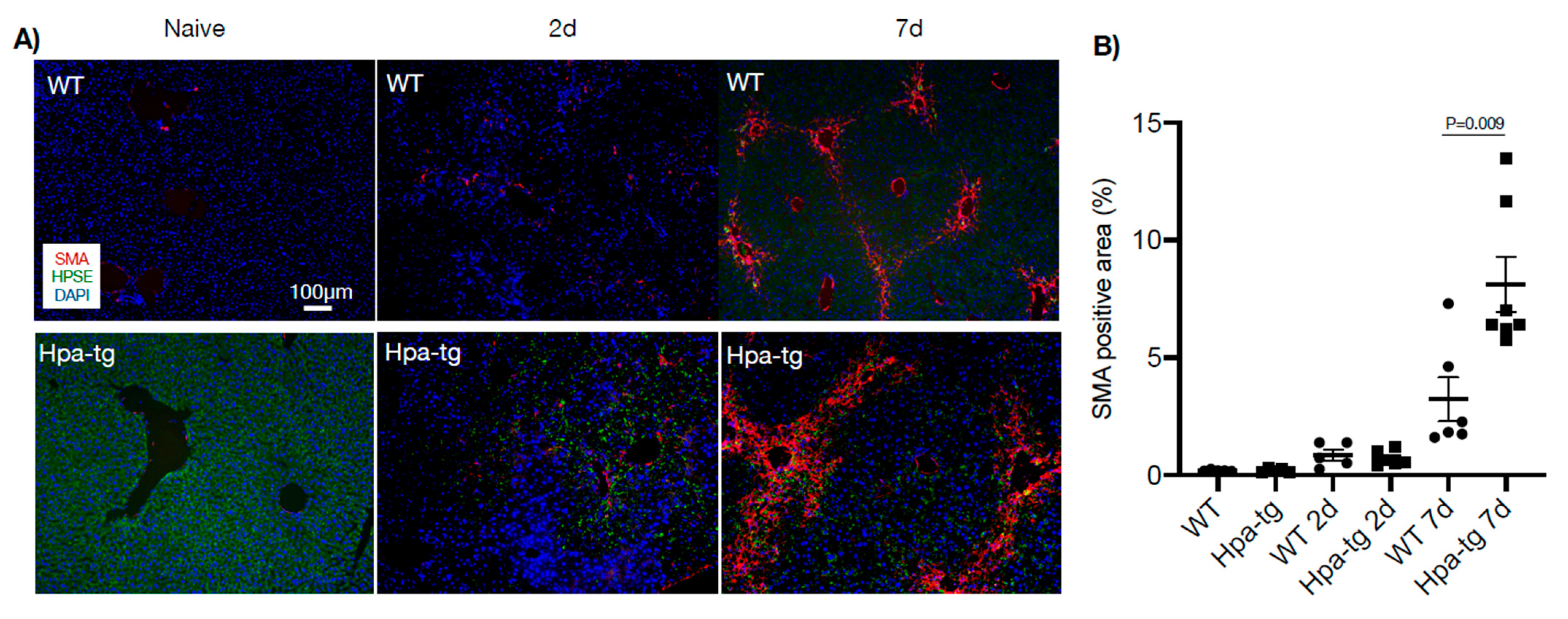
3.2. Higher Levels of Alpha-SMA in Hpa-Tg Liver
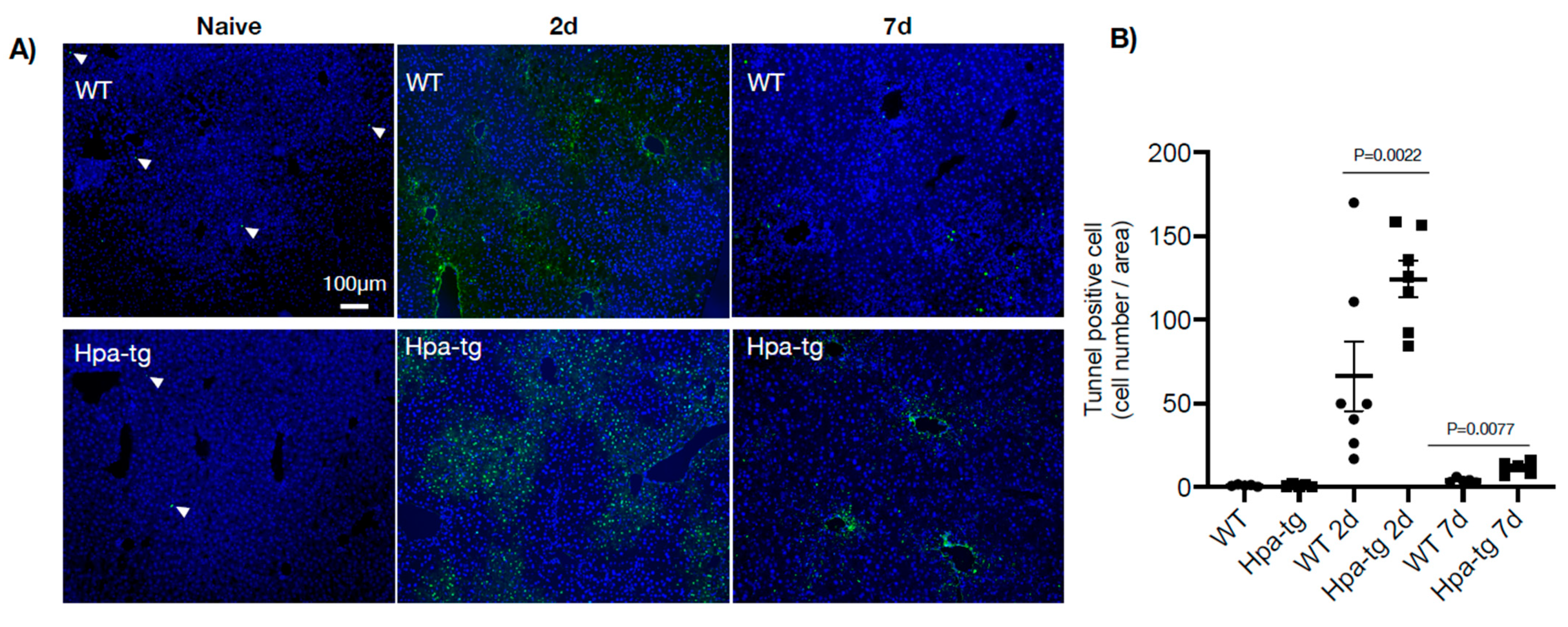
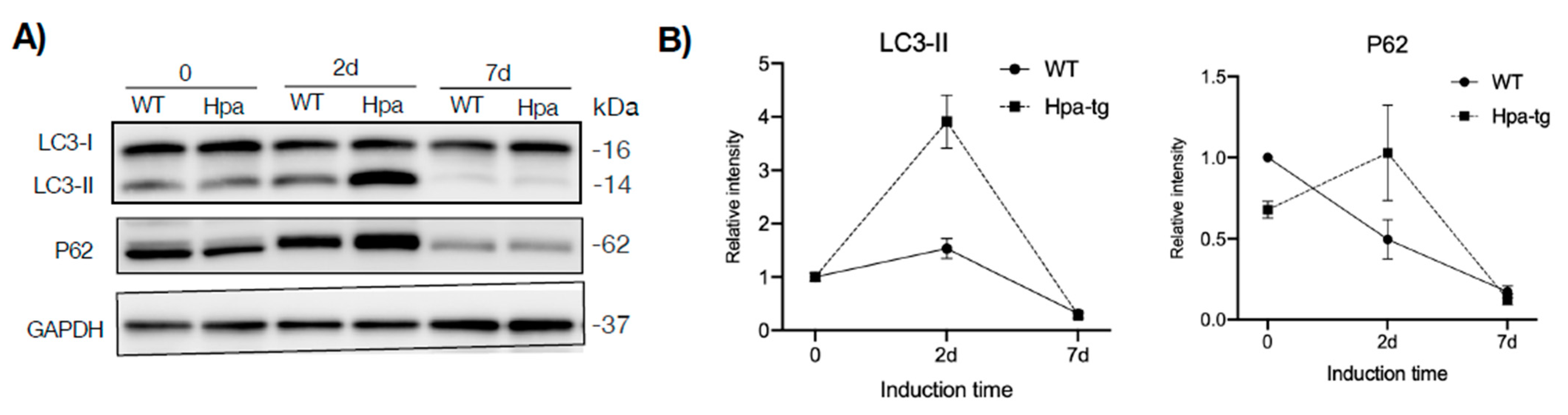
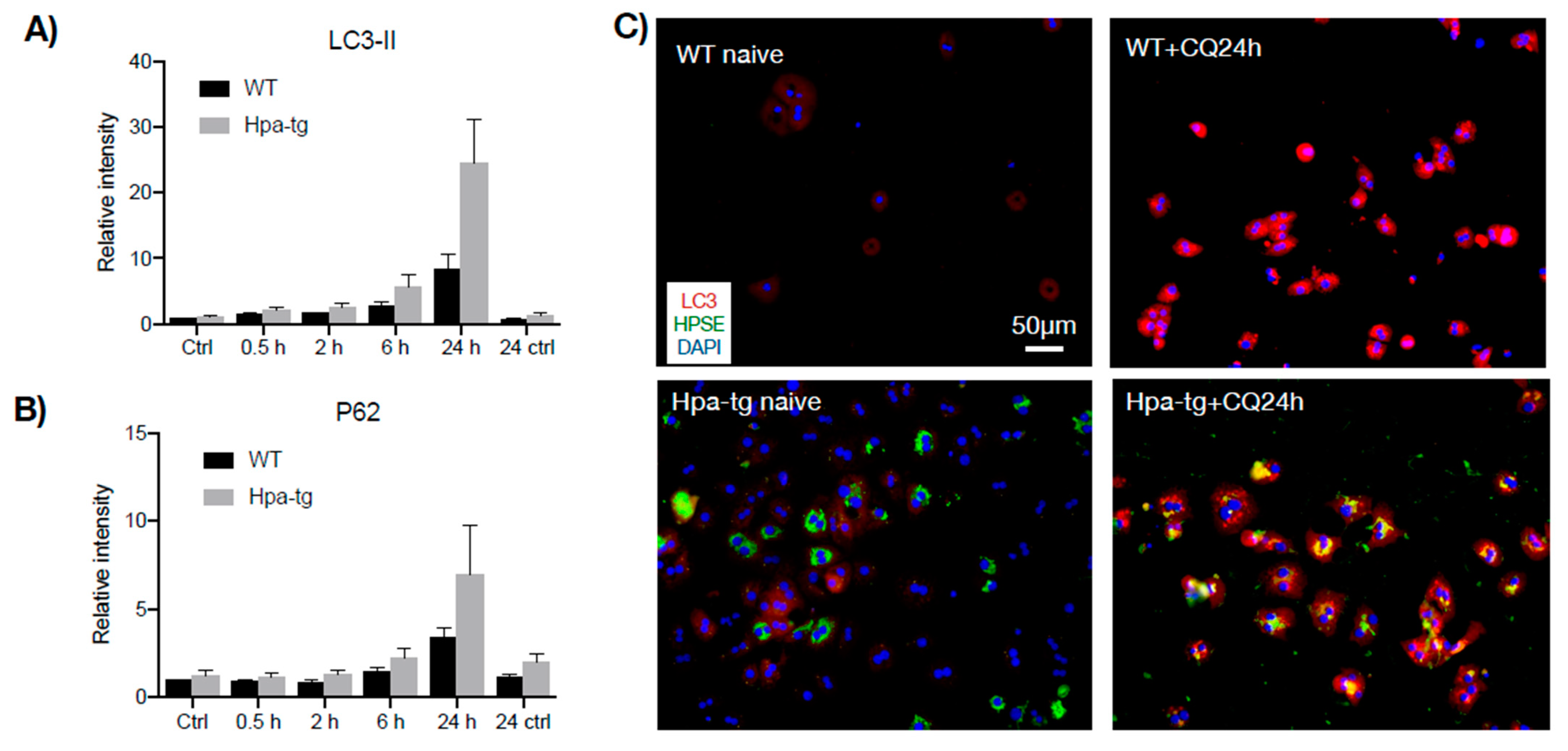
3.3. Higher Autophagic Activity of Hpa-Tg Hepatocytes in Response to CCL4
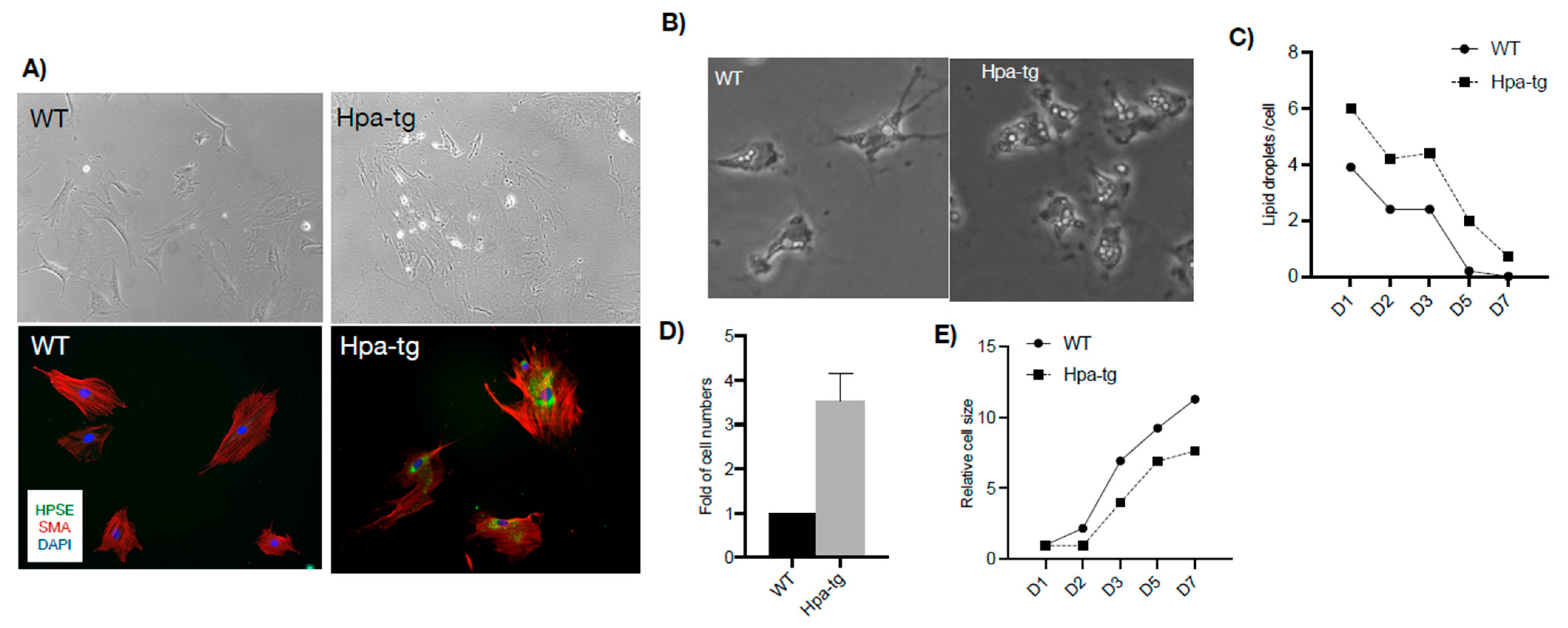
3.4. Hepatic Stellate Cells
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pellicoro, A.; Ramachandran, P.; Iredale, J.P.; Fallowfield, J.A. Liver fibrosis and repair: Immune regulation of wound healing in a solid organ. Nat. Rev. Immunol. 2014, 14, 181–194. [Google Scholar] [CrossRef]
- Puche, J.E.; Saiman, Y.; Friedman, S.L. Hepatic stellate cells and liver fibrosis. Compr. Physiol. 2013, 3, 1473–1492. [Google Scholar] [PubMed]
- Ginès, P.; Castera, L.; Lammert, F.; Graupera, I.; Serra-Burriel, M.; Allen, A.M.; Wong, V.W.; Hartmann, P.; Thiele, M.; Caballeria, L.; et al. Population screening for liver fibrosis: Toward early diagnosis and intervention for chronic liver diseases. Hepatology 2021, 75, 219–228. [Google Scholar] [CrossRef]
- Lindahl, U.; Kjellén, L. Pathophysiology of heparan sulphate: Many diseases, few drugs. J. Intern. Med. 2013, 273, 555–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlodavsky, I.; Ilan, N.; Sanderson, R.D. Forty Years of Basic and Translational Heparanase Research. Adv. Exp. Med. Biol. 2020, 1221, 3–59. [Google Scholar] [CrossRef] [Green Version]
- Ikeguchi, M.; Hirooka, Y.; Kaibara, N. Heparanase gene expression and its correlation with spontaneous apoptosis in hepatocytes of cirrhotic liver and carcinoma. Eur. J. Cancer 2003, 39, 86–90. [Google Scholar] [CrossRef]
- Tátrai, P.; Egedi, K.; Somorácz, Á.; van Kuppevelt, T.H.; Dam, G.T.; Lyon, M.; Deakin, J.A.; Kiss, A.; Schaff, Z.; Kovalszky, I. Quantitative and Qualitative Alterations of Heparan Sulfate in Fibrogenic Liver Diseases and Hepatocellular Cancer. J. Histochem. Cytochem. 2010, 58, 429–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvis, M.L.E.; Jia, J.; Zhang, X.; Jastrebova, N.; Spillmann, D.; Gottfridsson, E.; Van Kuppevelt, T.H.; Zcharia, E.; Vlodavsky, I.; Lindahl, U.; et al. Transgenic or tumor-induced expression of heparanase upregulates sulfation of heparan sulfate. Nat. Chem. Biol. 2007, 3, 773–778. [Google Scholar] [CrossRef]
- Zcharia, E.; Metzger, S.; Chajek-Shaul, T.; Aingorn, H.; Elkin, M.; Friedmann, Y.; Weinstein, T.; Li, J.-P.; Lindahl, U.; Vlodavsky, I. Transgenic expression of mammalian heparanase uncovers physiological functions of heparan sulfate in tissue morphogenesis, vascularization, and feeding behavior. FASEB J. 2004, 18, 252–263. [Google Scholar] [CrossRef]
- Faccioli, L.A.; Dias, M.L.; Paranhos, B.A.; Goldenberg, R.C.D.S. Liver cirrhosis: An overview of experimental models in rodents. Life Sci. 2022, 301, 120615. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Goldblatt, P.J.; Hinton, D.E.; Lipsky, M.M.; Chacko, J.; Trump, B.F. Mouse liver cell culture. I. Hepatocyte isolation. Vitro 1981, 17, 913–925. [Google Scholar] [CrossRef]
- Mederacke, I.; Dapito, D.H.; Affò, S.; Uchinami, H.; Schwabe, R.F. High-yield and high-purity isolation of hepatic stellate cells from normal and fibrotic mouse livers. Nat. Protoc. 2015, 10, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Khomich, O.; Ivanov, A.V.; Bartosch, B. Metabolic Hallmarks of Hepatic Stellate Cells in Liver Fibrosis. Cells 2019, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Nah, E.-H.; Cho, S.; Kim, S.; Chu, J.; Kwon, E.; Cho, H.-I. Prevalence of liver fibrosis and associated risk factors in the Korean general population: A retrospective cross-sectional study. BMJ Open 2021, 11, e046529. [Google Scholar] [CrossRef]
- Ravichandra, A.; Schwabe, R.F. Mouse Models of Liver Fibrosis. Methods Mol. Biol. 2021, 2299, 339–356. [Google Scholar] [CrossRef]
- Secchi, M.F.; Crescenzi, M.; Masola, V.; Russo, F.P.; Floreani, A.; Onisto, M. Heparanase and macrophage interplay in the onset of liver fibrosis. Sci. Rep. 2017, 7, 14956. [Google Scholar] [CrossRef] [Green Version]
- Goldshmidt, O.; Yeikilis, R.; Mawasi, N.; Paizi, M.; Gan, N.; Ilan, N.; Pappo, O.; Vlodavsky, I.; Spira, G. Heparanase expression during normal liver development and following partial hepatectomy. J. Pathol. 2004, 203, 594–602. [Google Scholar] [CrossRef]
- Ohayon, O.; Mawasi, N.; Pevzner, A.; Tryvitz, A.; Gildor, T.; Pines, M.; Rojkind, M.; Paizi, M.; Spira, G. Halofuginone upregulates the expression of heparanase in thioacetamide-induced liver fibrosis in rats. Lab. Investig. 2008, 88, 627–633. [Google Scholar] [CrossRef]
- El-Assal, O.N.; Yamanoi, A.; Ono, T.; Kohno, H.; Nagasue, N. The clinicopathological significance of heparanase and basic fibroblast growth factor expressions in hepatocellular carcinoma. Clin. Cancer Res. 2001, 7, 1299–1305. [Google Scholar]
- Xiao, Y. Heparanase expression in hepatocellular carcinoma and the cirrhotic liver. Hepatol. Res. 2003, 26, 192–198. [Google Scholar] [CrossRef]
- Masola, V.; Zaza, G.; Secchi, M.F.; Gambaro, G.; Lupo, A.; Onisto, M. Heparanase is a key player in renal fibrosis by regulating TGF-beta expression and activity. Biochim. Biophys. Acta 2014, 1843, 2122–2128. [Google Scholar] [CrossRef]
- Kinaneh, S.; Hijaze, W.; Mansour-Wattad, L.; Hammoud, R.; Zaidani, H.; Kabala, A.; Hamoud, S. Heparanase Inhibition Prevents Liver Steatosis in E0 Mice. J. Clin. Med. 2022, 11, 1672. [Google Scholar] [CrossRef]
- Blaner, W.S.; O’Byrne, S.M.; Wongsiriroj, N.; Kluwe, J.; D’Ambrosio, D.M.; Jiang, H.; Schwabe, R.F.; Hillman, E.M.; Piantedosi, R.; Libien, J. Hepatic stellate cell lipid droplets: A specialized lipid droplet for retinoid storage. Biochim. Biophys. Acta 2009, 1791, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef]
- Mastoridou, E.M.; Goussia, A.C.; Glantzounis, G.K.; Kanavaros, P.; Charchanti, A.V. Autophagy and Exosomes: Cross-Regulated Pathways Playing Major Roles in Hepatic Stellate Cells Activation and Liver Fibrosis. Front. Physiol. 2022, 12, 801340. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.; Wang, D.; Xu, Y.; Dong, R.; Yang, Y.; Lv, Q.; Chen, X.; Zhang, Z. The Upstream Pathway of mTOR-Mediated Autophagy in Liver Diseases. Cells 2019, 8, 1597. [Google Scholar] [CrossRef]
- Ilan, N.; Shteingauz, A.; Vlodavsky, I. Function from within: Autophagy induction by HPSE/heparanase—New possibilities for intervention. Autophagy 2015, 11, 2387–2389. [Google Scholar] [CrossRef] [Green Version]
- Sanderson, R.D.; Elkin, M.; Rapraeger, A.C.; Ilan, N.; Vlodavsky, I. Heparanase regulation of cancer, autophagy and inflammation: New mechanisms and targets for therapy. FEBS J. 2016, 284, 42–55. [Google Scholar] [CrossRef]
- Shteingauz, A.; Boyango, I.; Naroditsky, I.; Hammond, E.; Gruber, M.; Doweck, I.; Ilan, N.; Vlodavsky, I. Heparanase Enhances Tumor Growth and Chemoresistance by Promoting Autophagy. Cancer Res. 2015, 75, 3946–3957. [Google Scholar] [CrossRef] [Green Version]
- Amanzada, A.; Malik, I.A.; Nischwitz, M.; Sultan, S.; Naz, N.; Ramadori, G. Myeloperoxidase and elastase are only expressed by neutrophils in normal and in inflammed liver. Histochem. Cell Biol. 2011, 135, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Wójcik, M.; Ramadori, P.; Blaschke, M.; Sultan, S.; Khan, S.; Malik, I.A.; Naz, N.; Martius, G.; Ramadori, G.; Schultze, F.C. Immunodetection of cyclooxygenase-2 (COX-2) is restricted to tissue macrophages in normal rat liver and to recruited mononuclear phagocytes in liver injury and cholangiocarcinoma. Histochem. Cell Biol. 2011, 137, 217–233. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Tsuchiya, A.; Seino, S.; Kawata, Y.; Kojima, Y.; Ikarashi, S.; Lewis, P.J.S.; Lu, W.-Y.; Kikuta, J.; Kawai, H.; et al. Mesenchymal Stem Cells and Induced Bone Marrow-Derived Macrophages Synergistically Improve Liver Fibrosis in Mice. Stem Cells Transl. Med. 2018, 8, 271–284. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Host | Working Dilution | Source and Code |
|---|---|---|---|
| Atg3 | Rabbit | 1:1000 (WB) | CST#3415 |
| Atg7 | Rabbit | 1:1000 (WB) | CST#8558 |
| Beclin-1 | Rabbit | 1:1000 (WB) | CST#3495 |
| LC3I/II | Rabbit | 1:1000 (WB) | CST#12741 |
| P62 | Rabbit | 1:1000 (WB) | CST#23214 |
| MMP9 | Rabbit | 1:1000 (WB) | Abcam#76003 |
| GAPDH | Rabbit | 1:2000 (WB) | CST#2118 |
| Collagen III | Rabbit | 1:100 (IHC) | Abcam #7778 |
| HPSE | Mouse | 1:200 (IHC) | In-house monoclonal |
| SMA | Rabbit | 1:100 (IHC) | CST #19245 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, X.; Jia, J.; Zhang, T.; Zhang, X.; Vlodavsky, I.; Li, J.-p. Heparanase Expression Propagates Liver Damage in CCL4-Induced Mouse Model. Cells 2022, 11, 2035. https://doi.org/10.3390/cells11132035
Cheng X, Jia J, Zhang T, Zhang X, Vlodavsky I, Li J-p. Heparanase Expression Propagates Liver Damage in CCL4-Induced Mouse Model. Cells. 2022; 11(13):2035. https://doi.org/10.3390/cells11132035
Chicago/Turabian StyleCheng, Xiaowen, Juan Jia, Tianji Zhang, Xiao Zhang, Israel Vlodavsky, and Jin-ping Li. 2022. "Heparanase Expression Propagates Liver Damage in CCL4-Induced Mouse Model" Cells 11, no. 13: 2035. https://doi.org/10.3390/cells11132035
APA StyleCheng, X., Jia, J., Zhang, T., Zhang, X., Vlodavsky, I., & Li, J. -p. (2022). Heparanase Expression Propagates Liver Damage in CCL4-Induced Mouse Model. Cells, 11(13), 2035. https://doi.org/10.3390/cells11132035