
Early Dietary Exposures Epigenetically Program Mammary Cancer Susceptibility through Igf1-Mediated Expansion of the Mammary Stem Cell Compartment

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
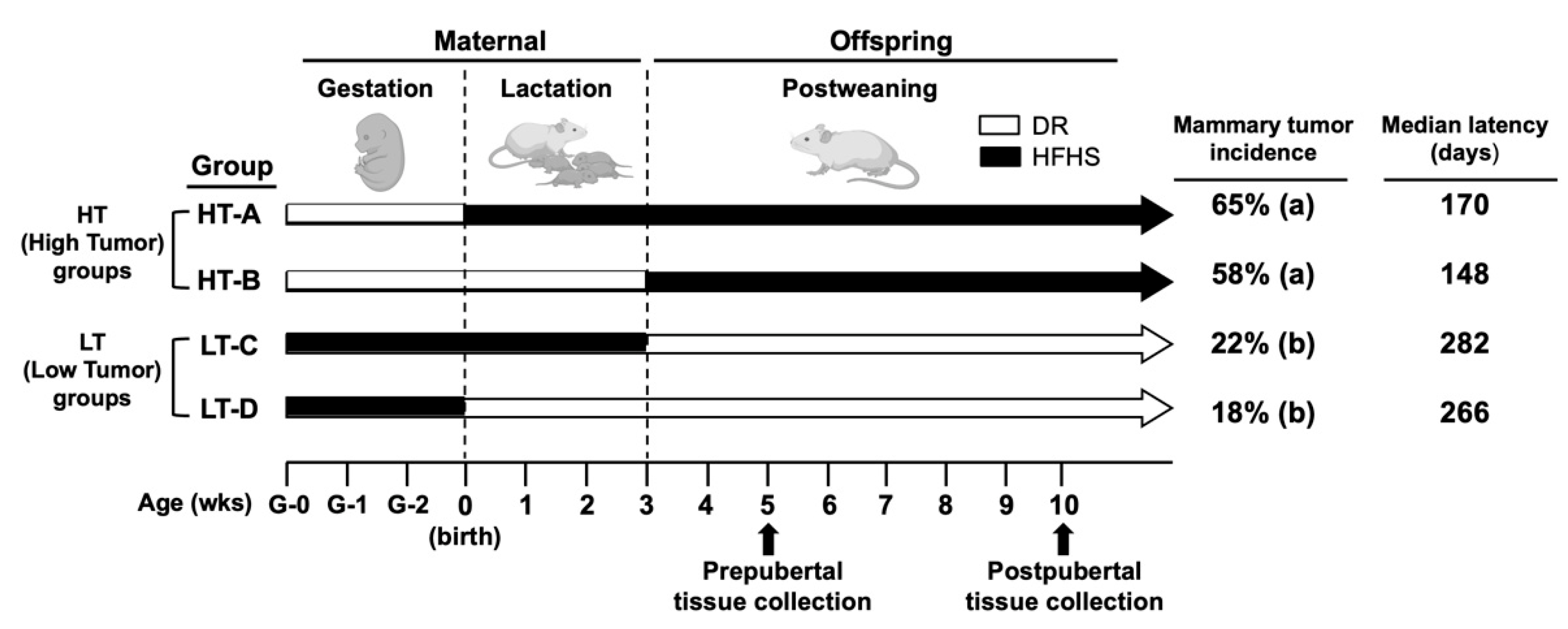
2.1. Animal Model
2.2. Mice and Diets
2.3. Mammary Epithelial Cell Isolation
2.4. Flow Cytometry and Cell Sorting
2.5. Mammosphere Culture and Limiting Dilution Assay
2.6. Conditioned Medium (CM) Assay
2.7. RT-qPCR and Western Blot Analysis
2.8. ELISA Assay
2.9. DNA Methylation Analysis
2.10. siRNA Transfection
2.11. Carcinogen Treatment
2.12. Histological Analyses and Immunohistochemistry
2.13. Statistics
3. Results
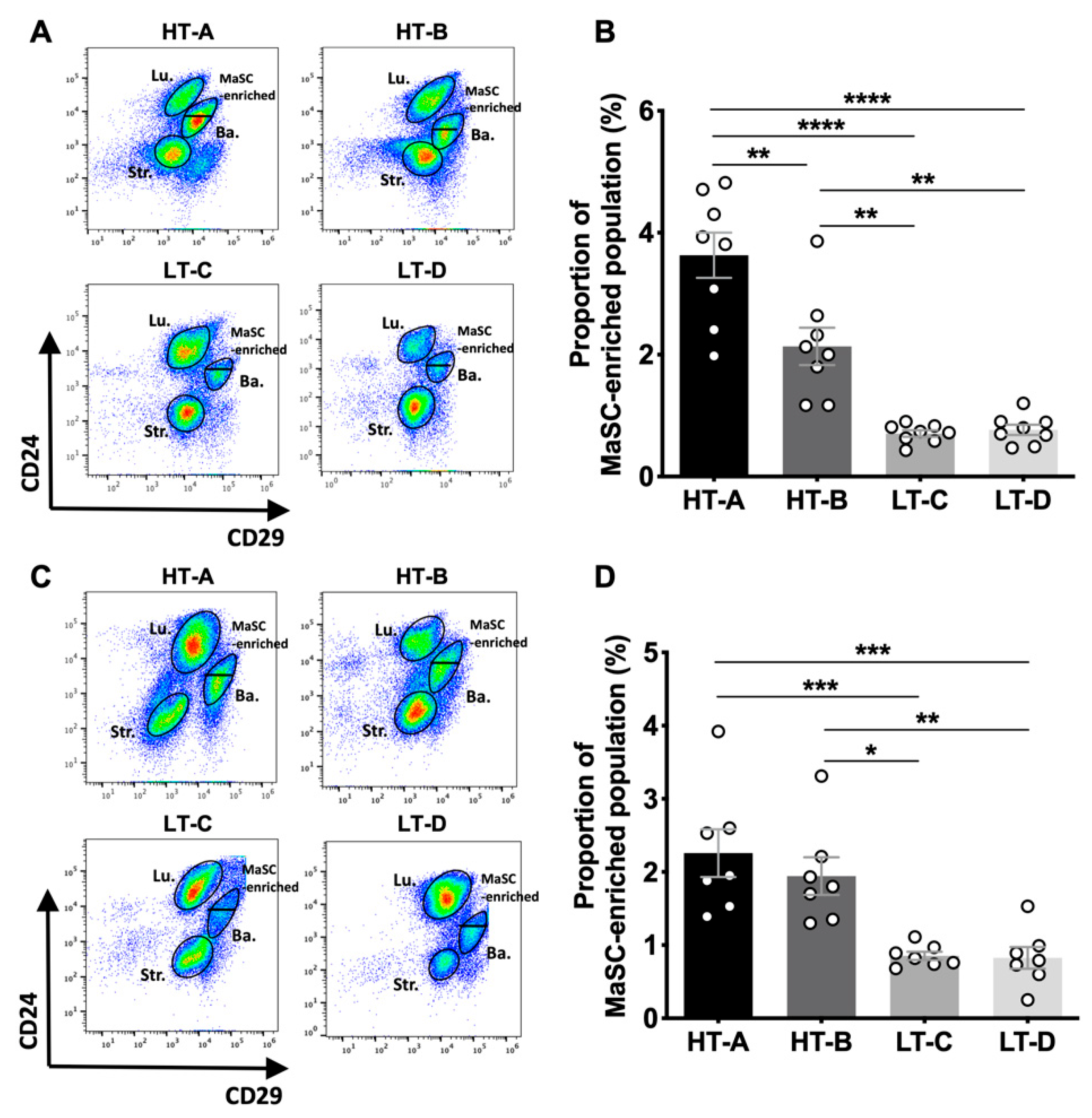
3.1. Mammary Glands from Pre- and Post-Pubertal Female Mice in the HT-A and HT-B Groups Had an Expanded MaSC-Enriched Compartment
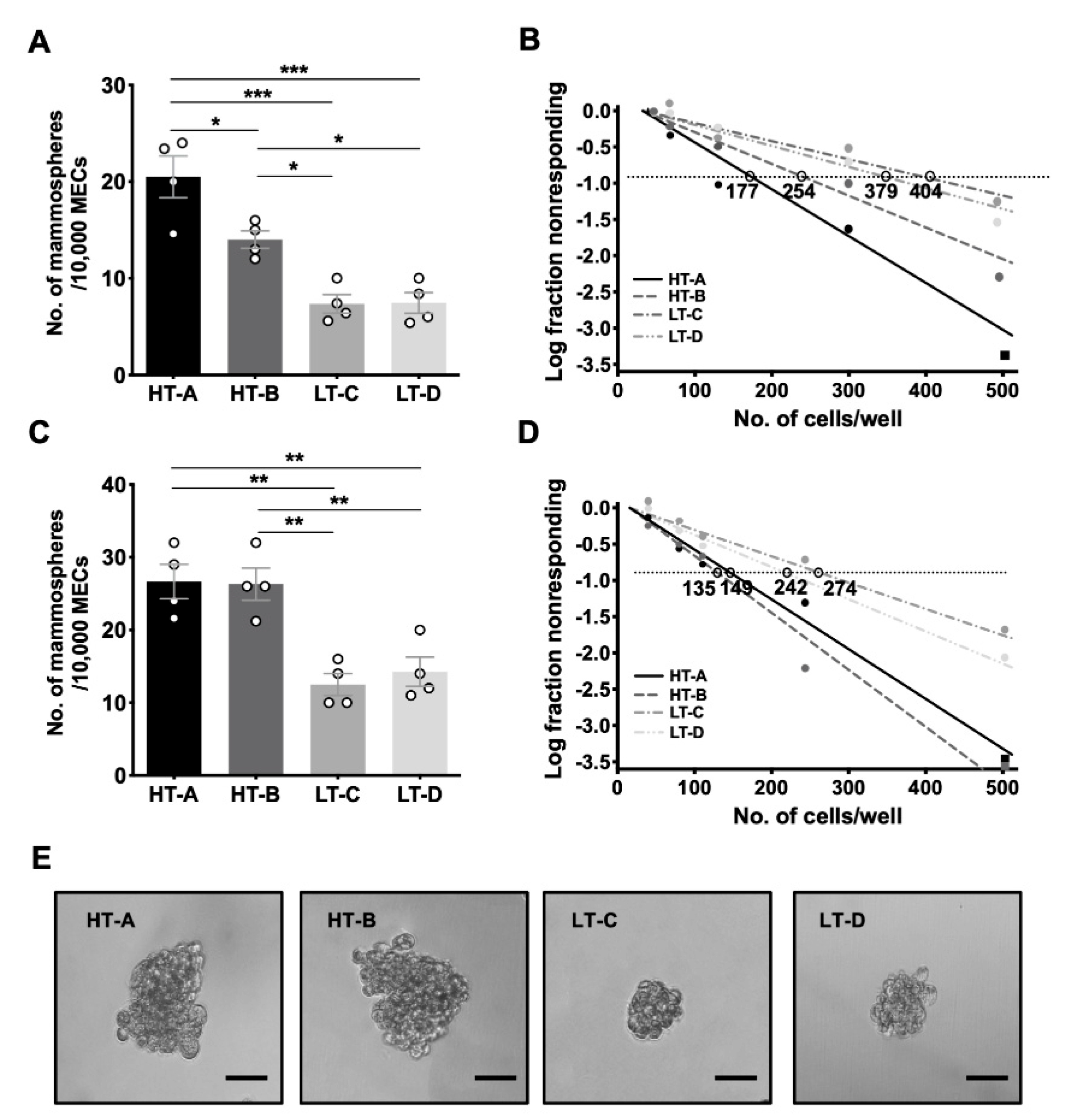
3.2. The Number of Mammosphere-Initiating Cells Was Increased in Mice from the HT Dietary Regimen Groups (HT-A and HT-B)
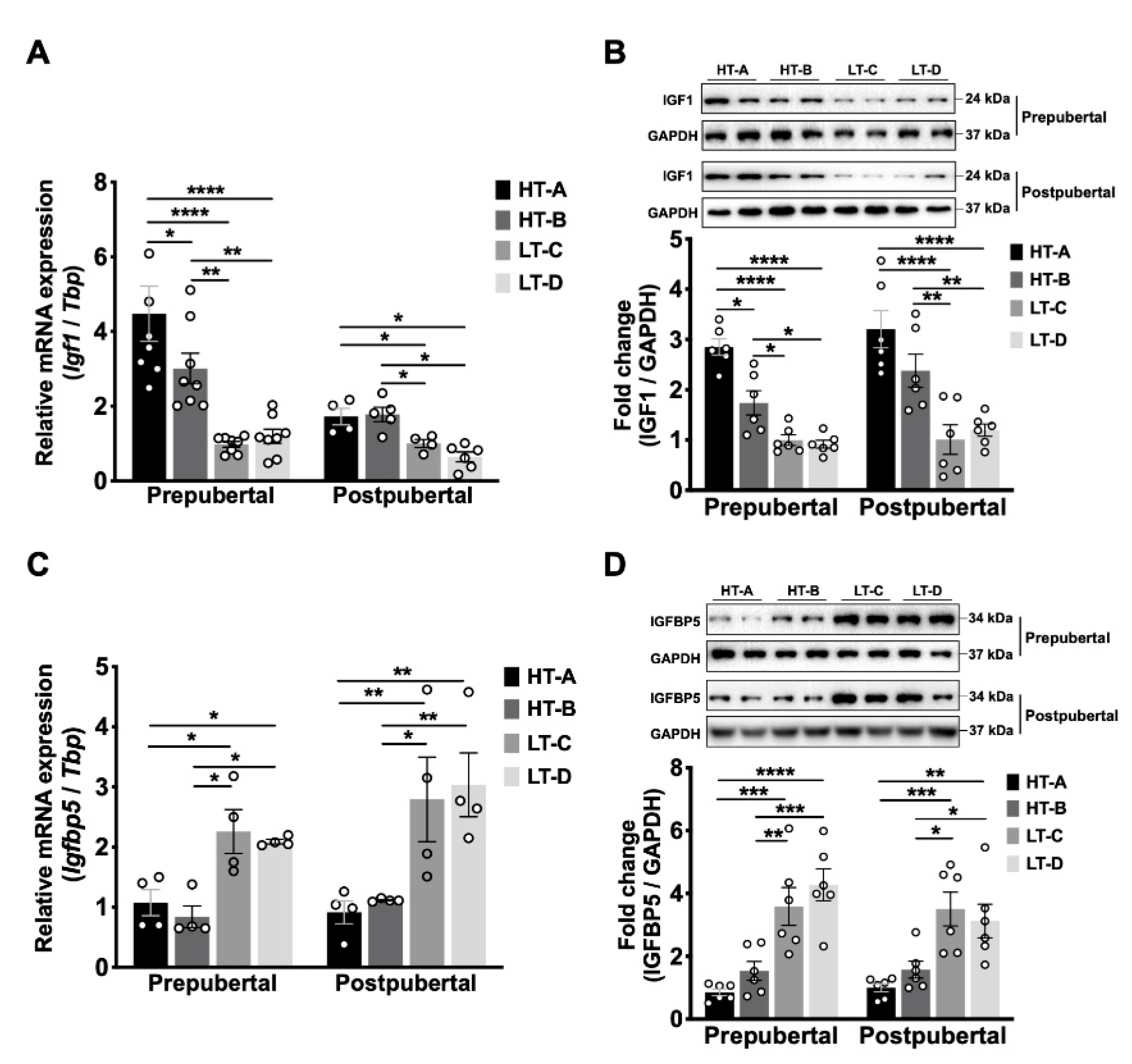
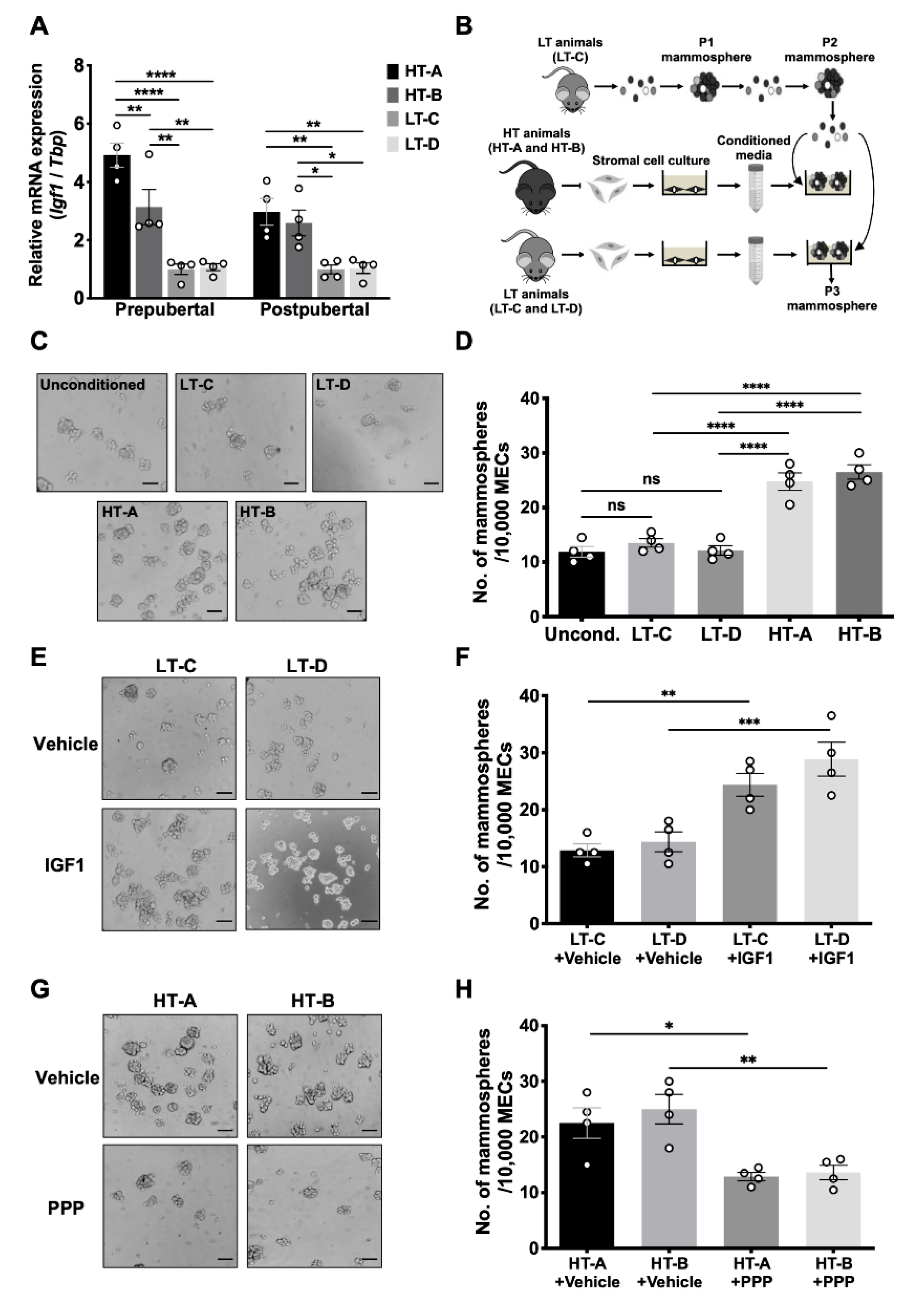
3.3. The HT-A and HT-B Dietary Regimens Increased Mammary Igf1 Levels and Decreased Igfbp5 Levels
3.4. The HT-A and HT-B Dietary Regimens Increased Igf1 Production in Mammary Stromal Cells and Promoted MaSC Self-Renewal
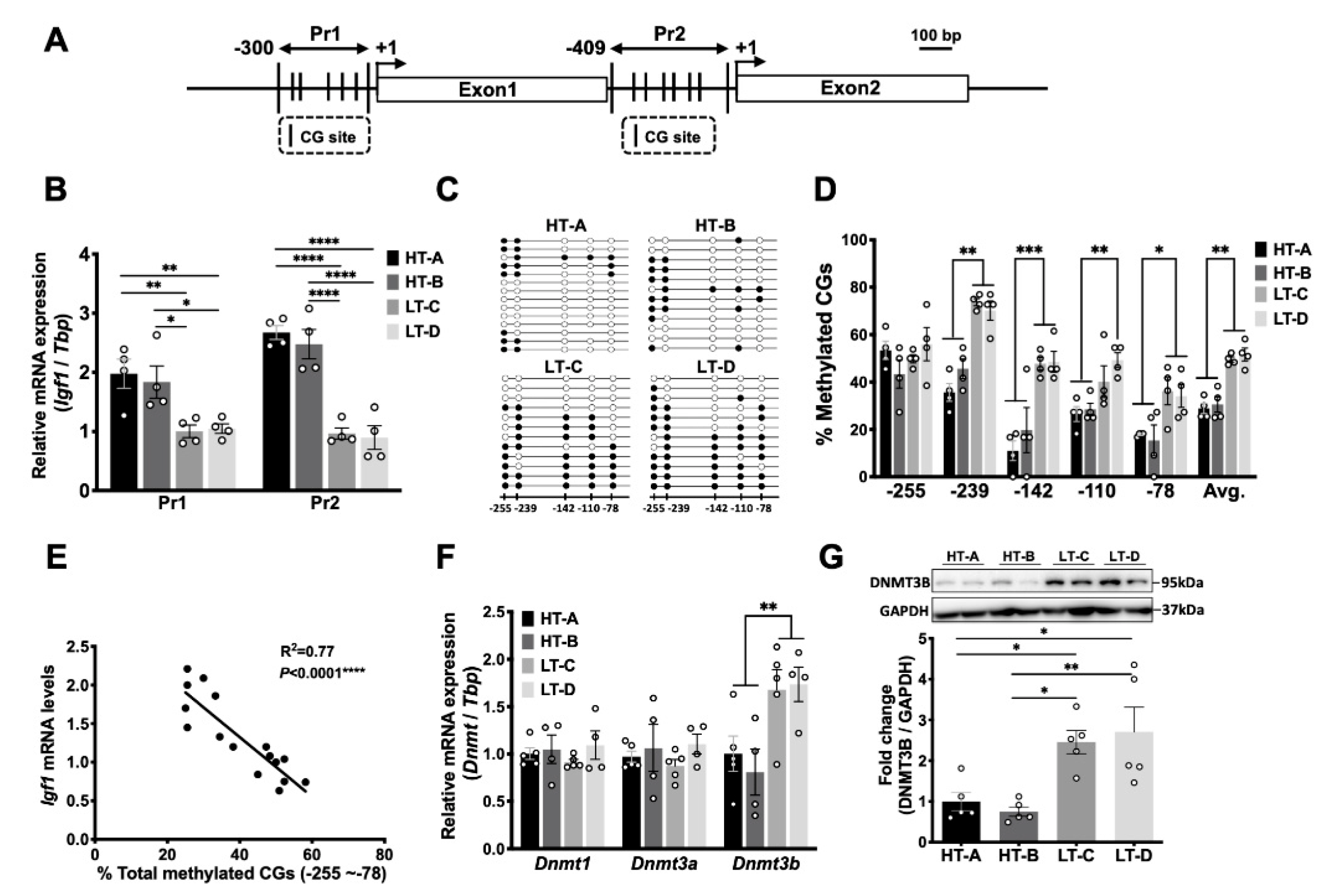
3.5. The HT-A and HT-B Dietary Regimens Decreased DNA Methylation of the Igf1 Pr1 Promoter in Mammary Stromal Cells
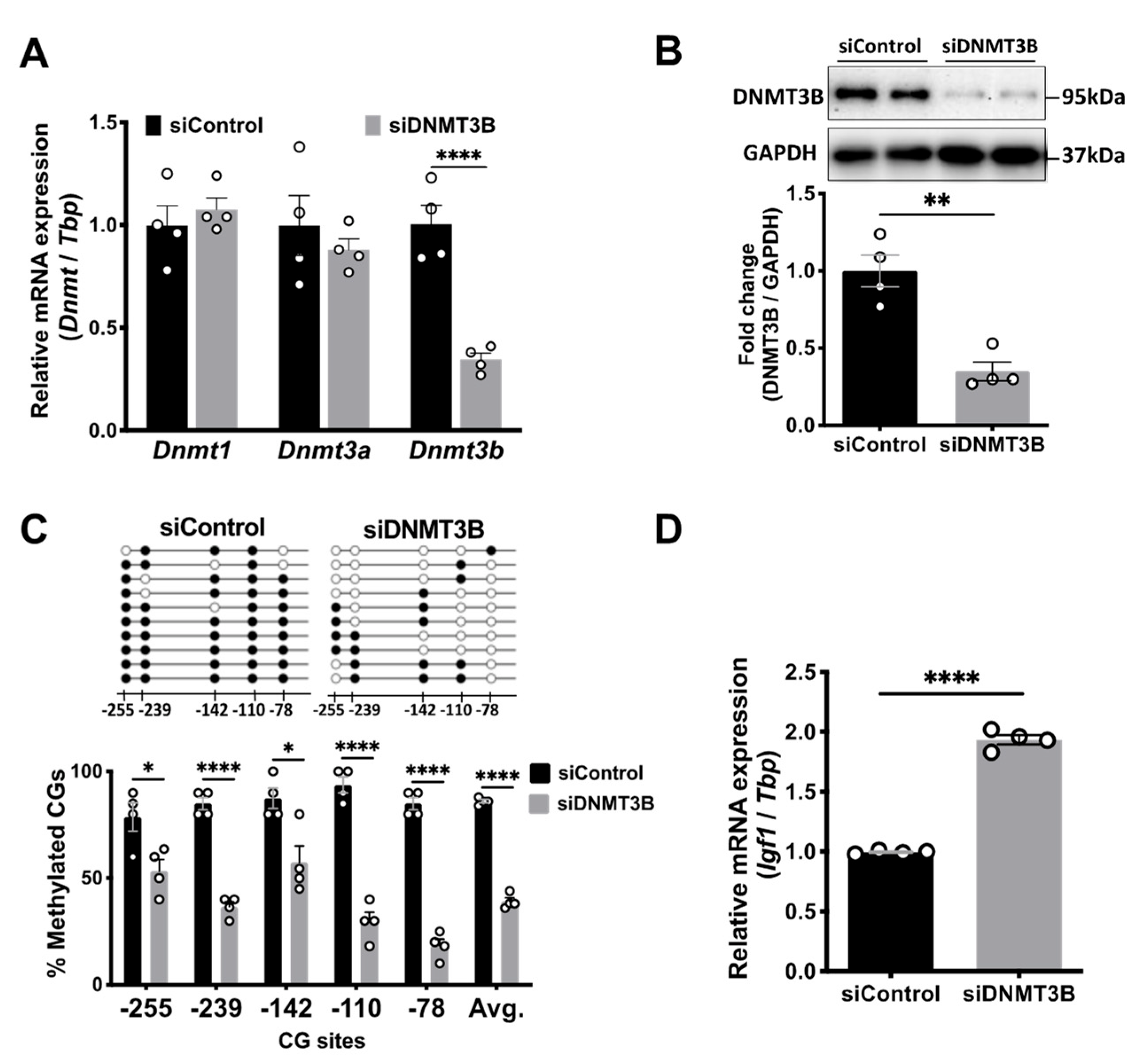
3.6. Dnmt3b Knockdown Decreased Igf1 Pr1 DNA Methylation and Increased Igf1 mRNA Expression
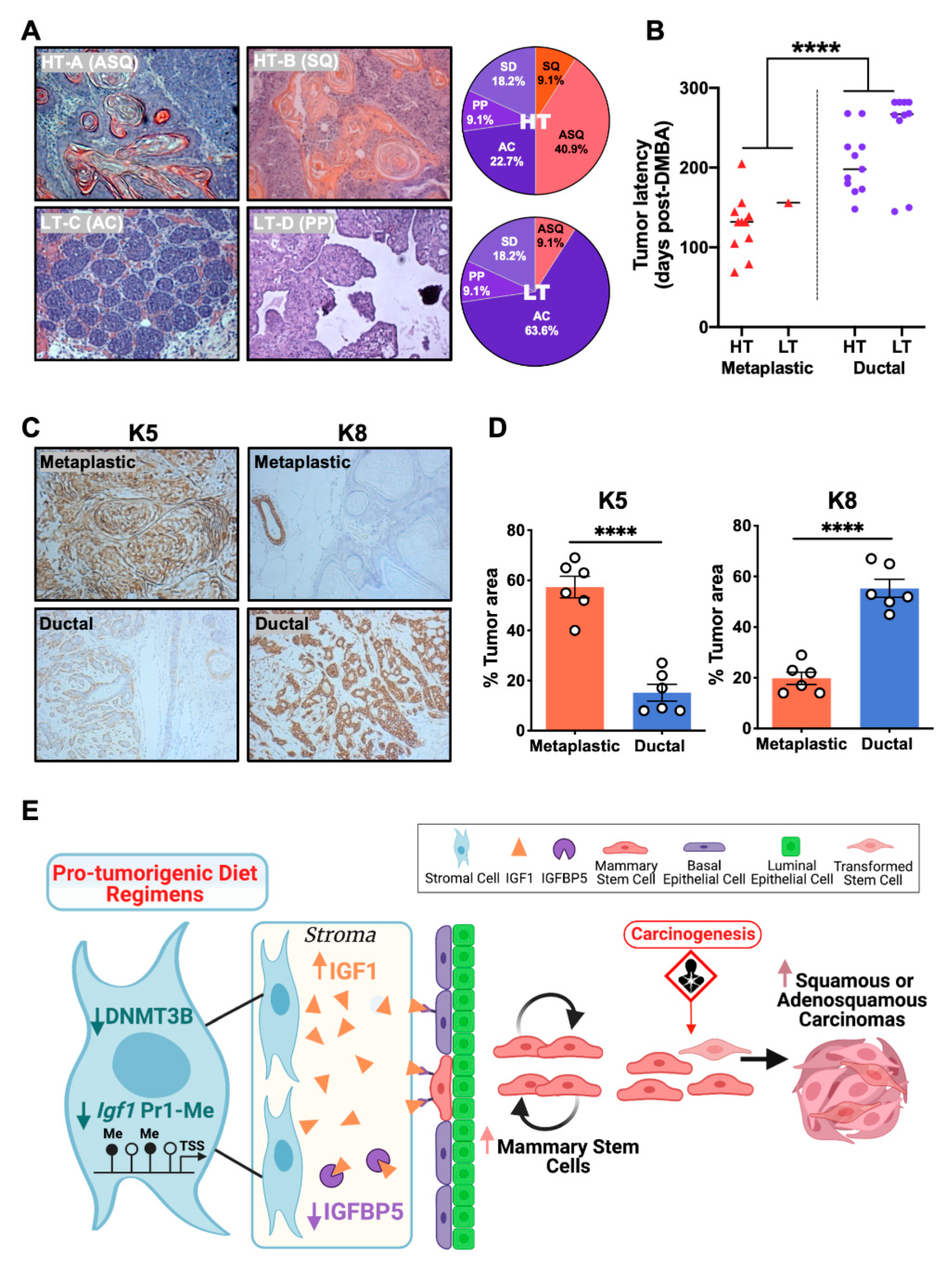
3.7. The HT Groups Developed Increased Proportions of Metaplastic Carcinomas
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barker, D.J. The fetal and infant origins of adult disease. BMJ 1990, 301, 1111. [Google Scholar] [CrossRef] [PubMed]
- Knop, M.R.; Geng, T.T.; Gorny, A.W.; Ding, R.; Li, C.; Ley, S.H.; Huang, T. Birth Weight and Risk of Type 2 Diabetes Mellitus, Cardiovascular Disease, and Hypertension in Adults: A Meta-Analysis of 7 646 267 Participants From 135 Studies. J. Am. Heart Assoc. 2018, 7, e008870. [Google Scholar] [CrossRef] [PubMed]
- Elias, S.G.; Peeters, P.H.; Grobbee, D.E.; van Noord, P.A. Breast cancer risk after caloric restriction during the 1944–1945 Dutch famine. J. Natl. Cancer Inst. 2004, 96, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Painter, R.C.; De Rooij, S.R.; Bossuyt, P.M.; Osmond, C.; Barker, D.J.; Bleker, O.P.; Roseboom, T.J. A possible link between prenatal exposure to famine and breast cancer: A preliminary study. Am. J. Hum. Biol. 2006, 18, 853–856. [Google Scholar] [CrossRef] [PubMed]
- Russo, J. The Windows of Susceptibility to Breast Cancer. In The Pathobiology of Breast Cancer; Russo, J., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–20. [Google Scholar]
- de Oliveira Andrade, F.; Fontelles, C.C.; Rosim, M.P.; de Oliveira, T.F.; de Melo Loureiro, A.P.; Mancini-Filho, J.; Rogero, M.M.; Moreno, F.S.; de Assis, S.; Barbisan, L.F.; et al. Exposure to lard-based high-fat diet during fetal and lactation periods modifies breast cancer susceptibility in adulthood in rats. J. Nutr. Biochem. 2014, 25, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Aupperlee, M.D.; Zhao, Y.; Tan, Y.S.; Zhu, Y.; Langohr, I.M.; Kirk, E.L.; Pirone, J.R.; Troester, M.A.; Schwartz, R.C.; Haslam, S.Z. Puberty-specific promotion of mammary tumorigenesis by a high animal fat diet. Breast Cancer Res. 2015, 17, 138. [Google Scholar] [CrossRef]
- Lambertz, I.U.; Luo, L.; Berton, T.R.; Schwartz, S.L.; Hursting, S.D.; Conti, C.J.; Fuchs-Young, R. Early Exposure to a High Fat/High Sugar Diet Increases the Mammary Stem Cell Compartment and Mammary Tumor Risk in Female Mice. Cancer Prev. Res. 2017, 10, 553–562. [Google Scholar] [CrossRef]
- Prins, G.S. Developmental estrogenization: Prostate gland reprogramming leads to increased disease risk with aging. Differentiation 2021, 118, 72–81. [Google Scholar] [CrossRef]
- Helmin, K.A.; Morales-Nebreda, L.; Torres Acosta, M.A.; Anekalla, K.R.; Chen, S.-Y.; Abdala-Valencia, H.; Politanska, Y.; Cheresh, P.; Akbarpour, M.; Steinert, E.M.; et al. Maintenance DNA methylation is essential for regulatory T cell development and stability of suppressive function. J. Clin. Investig. 2020, 130, 6571–6587. [Google Scholar] [CrossRef]
- Atlasi, Y.; Stunnenberg, H.G. The interplay of epigenetic marks during stem cell differentiation and development. Nat. Rev. Genet. 2017, 18, 643–658. [Google Scholar] [CrossRef]
- de Almeida, B.P.; Apolonio, J.D.; Binnie, A.; Castelo-Branco, P. Roadmap of DNA methylation in breast cancer identifies novel prognostic biomarkers. BMC Cancer 2019, 19, 219. [Google Scholar] [CrossRef] [PubMed]
- de Assis, S.; Warri, A.; Cruz, M.I.; Laja, O.; Tian, Y.; Zhang, B.; Wang, Y.; Huang, T.H.; Hilakivi-Clarke, L. High-fat or ethinyl-oestradiol intake during pregnancy increases mammary cancer risk in several generations of offspring. Nat. Commun. 2012, 3, 1053. [Google Scholar] [CrossRef] [PubMed]
- Govindarajah, V.; Leung, Y.K.; Ying, J.; Gear, R.; Bornschein, R.L.; Medvedovic, M.; Ho, S.M. In utero exposure of rats to high-fat diets perturbs gene expression profiles and cancer susceptibility of prepubertal mammary glands. J. Nutr. Biochem. 2016, 29, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Fu, N.Y.; Nolan, E.; Lindeman, G.J.; Visvader, J.E. Stem Cells and the Differentiation Hierarchy in Mammary Gland Development. Physiol. Rev. 2020, 100, 489–523. [Google Scholar] [CrossRef] [PubMed]
- Tharmapalan, P.; Mahendralingam, M.; Berman, H.K.; Khokha, R. Mammary stem cells and progenitors: Targeting the roots of breast cancer for prevention. EMBO J. 2019, 38, e100852. [Google Scholar] [CrossRef]
- Morel, A.P.; Ginestier, C.; Pommier, R.M.; Cabaud, O.; Ruiz, E.; Wicinski, J.; Devouassoux-Shisheboran, M.; Combaret, V.; Finetti, P.; Chassot, C.; et al. A stemness-related ZEB1-MSRB3 axis governs cellular pliancy and breast cancer genome stability. Nat. Med. 2017, 23, 568–578. [Google Scholar] [CrossRef]
- Molyneux, G.; Geyer, F.C.; Magnay, F.A.; McCarthy, A.; Kendrick, H.; Natrajan, R.; MacKay, A.; Grigoriadis, A.; Tutt, A.; Ashworth, A.; et al. BRCA1 basal-like breast cancers originate from luminal epithelial progenitors and not from basal stem cells. Cell Stem Cell 2010, 7, 403–417. [Google Scholar] [CrossRef]
- Bu, W.; Liu, Z.; Jiang, W.; Nagi, C.; Huang, S.; Edwards, D.P.; Jo, E.; Mo, Q.; Creighton, C.J.; Hilsenbeck, S.G.; et al. Mammary Precancerous Stem and Non-Stem Cells Evolve into Cancers of Distinct Subtypes. Cancer Res. 2019, 79, 61–71. [Google Scholar] [CrossRef]
- Hollern, D.P.; Swiatnicki, M.R.; Andrechek, E.R. Histological subtypes of mouse mammary tumors reveal conserved relationships to human cancers. PLoS Genet. 2018, 14, e1007135. [Google Scholar] [CrossRef]
- Hennigs, A.; Riedel, F.; Gondos, A.; Sinn, P.; Schirmacher, P.; Marme, F.; Jager, D.; Kauczor, H.U.; Stieber, A.; Lindel, K.; et al. Prognosis of breast cancer molecular subtypes in routine clinical care: A large prospective cohort study. BMC Cancer 2016, 16, 734. [Google Scholar] [CrossRef]
- Slaga, T.J. SENCAR mouse skin tumorigenesis model versus other strains and stocks of mice. Environ. Health Perspect. 1986, 68, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Kumaki, Y.; Oda, M.; Okano, M. QUMA: Quantification tool for methylation analysis. Nucleic. Acids Res. 2008, 36, W170–W175. [Google Scholar] [CrossRef] [PubMed]
- Qing, W.G.; Conti, C.J.; LaBate, M.; Johnston, D.; Slaga, T.J.; MacLeod, M.C. Induction of mammary cancer and lymphoma by multiple, low oral doses of 7,12-dimethylbenz[a]anthracene in SENCAR mice. Carcinogenesis 1997, 18, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Cardiff, R.D.; Anver, M.R.; Gusterson, B.A.; Hennighausen, L.; Jensen, R.A.; Merino, M.J.; Rehm, S.; Russo, J.; Tavassoli, F.A.; Wakefield, L.M.; et al. The mammary pathology of genetically engineered mice: The consensus report and recommendations from the Annapolis meeting. Oncogene 2000, 19, 968–988. [Google Scholar] [CrossRef] [PubMed]
- Rota, L.M.; Lazzarino, D.A.; Ziegler, A.N.; LeRoith, D.; Wood, T.L. Determining mammosphere-forming potential: Application of the limiting dilution analysis. J. Mammary Gland Biol. Neoplasia 2012, 17, 119–123. [Google Scholar] [CrossRef]
- Hu, Y.; Smyth, G.K. ELDA: Extreme limiting dilution analysis for comparing depleted and enriched populations in stem cell and other assays. J. Immunol. Methods 2009, 347, 70–78. [Google Scholar] [CrossRef]
- Shackleton, M.; Vaillant, F.; Simpson, K.J.; Stingl, J.; Smyth, G.K.; Asselin-Labat, M.L.; Wu, L.; Lindeman, G.J.; Visvader, J.E. Generation of a functional mammary gland from a single stem cell. Nature 2006, 439, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Stingl, J.; Eirew, P.; Ricketson, I.; Shackleton, M.; Vaillant, F.; Choi, D.; Li, H.I.; Eaves, C.J. Purification and unique properties of mammary epithelial stem cells. Nature 2006, 439, 993–997. [Google Scholar] [CrossRef]
- Dontu, G.; Abdallah, W.M.; Foley, J.M.; Jackson, K.W.; Clarke, M.F.; Kawamura, M.J.; Wicha, M.S. In vitro propagation and transcriptional profiling of human mammary stem/progenitor cells. Genes Dev. 2003, 17, 1253–1270. [Google Scholar] [CrossRef]
- Shaw, F.L.; Harrison, H.; Spence, K.; Ablett, M.P.; Simões, B.M.; Farnie, G.; Clarke, R.B. A detailed mammosphere assay protocol for the quantification of breast stem cell activity. J. Mammary Gland Biol. Neoplasia 2012, 17, 111–117. [Google Scholar] [CrossRef]
- Murphy, N.; Knuppel, A.; Papadimitriou, N.; Martin, R.M.; Tsilidis, K.K.; Smith-Byrne, K.; Fensom, G.; Perez-Cornago, A.; Travis, R.C.; Key, T.J.; et al. Insulin-like growth factor-1, insulin-like growth factor-binding protein-3, and breast cancer risk: Observational and Mendelian randomization analyses with approximately 430 000 women. Ann. Oncol. 2020, 31, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Santos, A.; Konganti, K.; Hillhouse, A.; Lambertz, I.U.; Zheng, Y.; Gunaratna, R.T.; Threadgill, D.W.; Fuchs-Young, R.S. Overexpression of IGF-1 During Early Development Expands the Number of Mammary Stem Cells and Primes them for Transformation. Stem Cells 2022, 40, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Beattie, J.; Hawsawi, Y.; Alkharobi, H.; El-Gendy, R. IGFBP-2 and -5: Important regulators of normal and neoplastic mammary gland physiology. J. Cell Commun. Signal. 2015, 9, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Sitar, T.; Popowicz, G.M.; Siwanowicz, I.; Huber, R.; Holak, T.A. Structural basis for the inhibition of insulin-like growth factors by insulin-like growth factor-binding proteins. Proc. Natl. Acad. Sci. USA 2006, 103, 13028–13033. [Google Scholar] [CrossRef]
- Richert, M.M.; Wood, T.L. The insulin-like growth factors (IGF) and IGF type I receptor during postnatal growth of the murine mammary gland: Sites of messenger ribonucleic acid expression and potential functions. Endocrinology 1999, 140, 454–461. [Google Scholar] [CrossRef]
- Oberbauer, A.M. The Regulation of IGF-1 Gene Transcription and Splicing during Development and Aging. Front. Endocrinol. 2013, 4, 39. [Google Scholar] [CrossRef]
- Rowzee, A.M.; Lazzarino, D.A.; Rota, L.; Sun, Z.; Wood, T.L. IGF ligand and receptor regulation of mammary development. J. Mammary Gland. Biol. Neoplasia 2008, 13, 361–370. [Google Scholar] [CrossRef]
- Reddy, T.P.; Rosato, R.R.; Li, X.; Moulder, S.; Piwnica-Worms, H.; Chang, J.C. A comprehensive overview of metaplastic breast cancer: Clinical features and molecular aberrations. Breast Cancer Res. 2020, 22, 121. [Google Scholar] [CrossRef]
- Reis-Filho, J.S.; Milanezi, F.; Steele, D.; Savage, K.; Simpson, P.T.; Nesland, J.M.; Pereira, E.M.; Lakhani, S.R.; Schmitt, F.C. Metaplastic breast carcinomas are basal-like tumours. Histopathology 2006, 49, 10–21. [Google Scholar] [CrossRef]
- Keller, P.J.; Arendt, L.M.; Skibinski, A.; Logvinenko, T.; Klebba, I.; Dong, S.; Smith, A.E.; Prat, A.; Perou, C.M.; Gilmore, H.; et al. Defining the cellular precursors to human breast cancer. Proc. Natl. Acad. Sci. USA 2012, 109, 2772–2777. [Google Scholar] [CrossRef]
- Rotwein, P. Diversification of the insulin-like growth factor 1 gene in mammals. PLoS ONE 2017, 12, e0189642. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Zhou, Q.-J.; Xiong, Y.; Li, B.; Li, X.-T. Preeclampsia is associated with hypermethylation of IGF-1 promoter mediated by DNMT1. Am. J. Transl. Res. 2018, 10, 16–39. [Google Scholar] [PubMed]
- Ouni, M.; Gunes, Y.; Belot, M.P.; Castell, A.L.; Fradin, D.; Bougnères, P. The Igf1 P2 promoter is an epigenetic QTL for circulating Igf1 and human growth. Clin. Epigenet. 2015, 7, 22. [Google Scholar] [CrossRef]
- Nilsson, E.; Ben Maamar, M.; Skinner, M.K. Environmental impacts on sperm and oocyte epigenetics affect embryo cell epigenetics and transcription to promote the epigenetic inheritance of pathology and phenotypic variation. Reprod. Fertil. Dev. 2021, 33, 102–107. [Google Scholar] [CrossRef]
- Hao, J.; Zhang, Y.; Yan, X.; Yan, F.; Sun, Y.; Zeng, J.; Waigel, S.; Yin, Y.; Fraig, M.M.; Egilmez, N.K.; et al. Circulating Adipose Fatty Acid Binding Protein Is a New Link Underlying Obesity-Associated Breast/Mammary Tumor Development. Cell Metab. 2018, 28, 689–705. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Luo, L.; Lambertz, I.U.; Conti, C.J.; Fuchs-Young, R. Early Dietary Exposures Epigenetically Program Mammary Cancer Susceptibility through Igf1-Mediated Expansion of the Mammary Stem Cell Compartment. Cells 2022, 11, 2558. https://doi.org/10.3390/cells11162558
Zheng Y, Luo L, Lambertz IU, Conti CJ, Fuchs-Young R. Early Dietary Exposures Epigenetically Program Mammary Cancer Susceptibility through Igf1-Mediated Expansion of the Mammary Stem Cell Compartment. Cells. 2022; 11(16):2558. https://doi.org/10.3390/cells11162558
Chicago/Turabian StyleZheng, Yuanning, Linjie Luo, Isabel U. Lambertz, Claudio J. Conti, and Robin Fuchs-Young. 2022. "Early Dietary Exposures Epigenetically Program Mammary Cancer Susceptibility through Igf1-Mediated Expansion of the Mammary Stem Cell Compartment" Cells 11, no. 16: 2558. https://doi.org/10.3390/cells11162558
APA StyleZheng, Y., Luo, L., Lambertz, I. U., Conti, C. J., & Fuchs-Young, R. (2022). Early Dietary Exposures Epigenetically Program Mammary Cancer Susceptibility through Igf1-Mediated Expansion of the Mammary Stem Cell Compartment. Cells, 11(16), 2558. https://doi.org/10.3390/cells11162558