
Hepatic, Extrahepatic and Extracellular Vesicle Cytochrome P450 2E1 in Alcohol and Acetaminophen-Mediated Adverse Interactions and Potential Treatment Options

 ,
, 

Abstract
:1. Introduction
2. The Role of Hepatic CYP2E1 in Causal Toxicity and Potential Treatment Options
2.1. Acetaminophen (APAP)-Induced Liver Toxicity/Injury
2.2. Alcohol-Induced Liver Toxicity/Injury
2.3. Drug-Drug Interactions
2.4. Alcohol-Drug Interactions
3. Extrahepatic CYP2E1 in Drug Interactions and Potential Treatments
3.1. Kidney
3.2. Monocytes, Astrocytes and Neuronal Cells
3.3. Tumor Cells
4. CYP2E1 in Clinical Drug-Drug and Alcohol-Drug Interactions
4.1. Genotype of CYP2E1 and Related Drug Interactions
4.2. Probe Substrates and Drug Interactions
5. Extracellular Vesicles (EVs) and CYP2E1
5.1. Extracellular Vesicular CYP2E1
5.2. CYP2E1 in Cell-Cell Interactions
6. Authors’ Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ahmad, F.B.; Rossen, L.; Sutton, P. Provisional Drug Overdose Death Counts. Available online: https://www.cdc.gov/nchs/nvss/vsrr/drug-overdose-data.htm (accessed on 17 November 2021).
- Agrawal, S.; Khazaeni, B. Acetaminophen toxicity. In Statpearls [Internet]; Statpearls publishing: Tampa, FA, USA, 2021. [Google Scholar]
- Prescott, L.F. Paracetamol, alcohol and the liver. Br. J. Clin. Pharmacol. 2000, 49, 291–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Moreno, G.; Guardia, J.; Cutando, A. Interaction of paracetamol in chronic alcoholic patients. Importance for odontologists. Med. Oral Patol. Oral. Cir. Bucal 2008, 13, E235–E238. [Google Scholar] [PubMed]
- Wolf, K.K.; Wood Sg Fau-Allard, J.L.; Allard Jl Fau-Hunt, J.A.; Hunt Ja Fau-Gorman, N.; Gorman N Fau-Walton-Strong, B.W.; Walton-Strong Bw Fau-Szakacs, J.G.; Szakacs Jg Fau-Duan, S.X.; Duan Sx Fau-Hao, Q.; Hao Q Fau-Court, M.H.; Court Mh Fau-von Moltke, L.L.; et al. Role of CYP3A and CYP2E1 in alcohol-mediated increases in acetaminophen hepatotoxicity: Comparison of wild-type and Cyp2e1(-/-) mice. Res. Artic. 2007, 35, 1223–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, F.J.; Ueno, T.; Umeno, M.; Song, B.J.; Veech, R.L.; Gelboin, H.V. Microsomal ethanol oxidizing system: Transcriptional and posttranscriptional regulation of cytochrome P450, CYP2E1. Alcohol Alcohol. Suppl. 1991, 1, 97–101. [Google Scholar]
- Yoon, M.; Madden, M.C.; Barton, H.A. Extrahepatic metabolism by CYP2E1 in PBPK modeling of lipophilic volatile organic chemicals: Impacts on metabolic parameter estimation and prediction of dose metrics. J. Toxicol. Environ. Health A 2007, 70, 1527–1541. [Google Scholar] [CrossRef]
- Arinç, E.; Arslan, S.; Bozcaarmutlu, A.; Adali, O. Effects of diabetes on rabbit kidney and lung CYP2E1 and CYP2B4 expression and drug metabolism and potentiation of carcinogenic activity of N-nitrosodimethylamine in kidney and lung. Food Chem. Toxicol. 2007, 45, 107–118. [Google Scholar] [CrossRef]
- Roberts, B.J.; Shoaf, S.E.; Jeong, K.S.; Song, B.J. Induction of CYP2E1 in liver, kidney, brain and intestine during chronic ethanol administration and withdrawal: Evidence that CYP2E1 possesses a rapid phase half-life of 6 hours or less. Biochem. Biophys. Res. Commun. 1994, 205, 1064–1071. [Google Scholar] [CrossRef]
- Hannon-Fletcher, M.P.; Barnett, Y.A. Lymphocyte cytochrome P450 expression: Inducibility studies in male Wistar rats. Br. J. Biomed. Sci. 2008, 65, 1–6. [Google Scholar] [CrossRef]
- Gong, Y.; Rao, P.S.S.; Sinha, N.; Ranjit, S.; Cory, T.J.; Kumar, S. The role of cytochrome P450 2E1 on ethanol-mediated oxidative stress and HIV replication in human monocyte-derived macrophages. Biochem. Biophys. Rep. 2019, 17, 65–70. [Google Scholar] [CrossRef]
- Jin, M.; Arya, P.; Patel, K.; Singh, B.; Silverstein, P.S.; Bhat, H.K.; Kumar, A.; Kumar, S. Effect of alcohol on drug efflux protein and drug metabolic enzymes in U937 macrophages. Alcohol. Clin. Exp. Res. 2011, 35, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Jin, M.; Ande, A.; Sinha, N.; Silverstein, P.S.; Kumar, A. Alcohol consumption effect on antiretroviral therapy and HIV-1 pathogenesis: Role of cytochrome P450 isozymes. Expert Opin. Drug Metab. Toxicol. 2012, 8, 1363–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, M.; Ande, A.; Kumar, A.; Kumar, S. Regulation of cytochrome P450 2e1 expression by ethanol: Role of oxidative stress-mediated pkc/jnk/sp1 pathway. Cell Death Dis. 2013, 4, e554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, F.J. Role of cytochromes P450 in chemical toxicity and oxidative stress: Studies with CYP2E1. Mutat. Res. 2005, 569, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Pentiuk, O.O.; Kachula, S.O.; Herych, O. Cytochrome P4502E1. Polymorphism, physiological function, regulation, and role in pathology. Ukr. Biokhim. Zh. 2004, 76, 16–28. [Google Scholar]
- Haorah, J.; Ramirez, S.H.; Floreani, N.; Gorantla, S.; Morsey, B.; Persidsky, Y. Mechanism of alcohol-induced oxidative stress and neuronal injury. Free Radic. Biol. Med. 2008, 45, 1542–1550. [Google Scholar] [CrossRef] [Green Version]
- Conde de la Rosa, L.; Moshage, H.; Nieto, N. Hepatocyte oxidant stress and alcoholic liver disease. Rev. Esp. Enferm Dig. 2008, 100, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Seitz, H.K.; Mueller, S. The role of Cytochrom P4502E1 in Alcoholic Liver Disease and alcohol mediated carcinogenesis. Z. Gastroenterol. 2019, 57, 37–45. [Google Scholar] [CrossRef]
- Linhart, K.; Bartsch, H.; Seitz, H.K. The role of reactive oxygen species (ROS) and cytochrome P-450 2E1 in the generation of carcinogenic etheno-DNA adducts. Redox Biol. 2014, 3, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Bonsaksen, T.; Ekeberg, Ø.; Schou-Bredal, I.; Skogstad, L.; Heir, T.; Grimholt, T.K. Use of Alcohol and Addictive Drugs During the COVID-19 Outbreak in Norway: Associations With Mental Health and Pandemic-Related Problems. Front. Public Health 2021, 9, 667729. [Google Scholar] [CrossRef]
- Ramalho, R.A.-O. Alcohol consumption and alcohol-related problems during the COVID-19 pandemic: A narrative review. Australas. Psychiatry 2020, 28, 524–526. [Google Scholar] [CrossRef]
- Abdelmegeed, M.A.; Moon, K.H.; Chen, C.; Gonzalez, F.J.; Song, B.J. Role of cytochrome P450 2E1 in protein nitration and ubiquitin-mediated degradation during acetaminophen toxicity. Biochem. Pharm. 2010, 79, 57–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, S.G.; Cartun, R.W.; Wyand, D.S.; Khairallah, E.A.; Cohen, S.D. Immunohistochemical localization of acetaminophen in target tissues of the CD-1 mouse: Correspondence of covalent binding with toxicity. Fundam. Appl. Toxicol. 1995, 24, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Enriquez, A.; Leclercq, I.; Farrell, G.C.; Robertson, G. Altered expression of hepatic CYP2E1 and CYP4A in obese, diabetic ob/ob mice, and fa/fa Zucker rats. Biochem. Biophys. Res. Commun. 1999, 255, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Lucas, D.; Farez, C.; Bardou, L.G.; Vaisse, J.; Attali, J.R.; Valensi, P. Cytochrome P450 2E1 activity in diabetic and obese patients as assessed by chlorzoxazone hydroxylation. Fundam. Clin. Pharm. 1998, 12, 553–558. [Google Scholar] [CrossRef]
- Tomankova, V.; Anzenbacher, P.; Anzenbacherova, E. Effects of obesity on liver cytochromes P450 in various animal models. Biomed. Pap. 2017, 161, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Hanley, M.J.; Abernethy, D.R.; Greenblatt, D.J. Effect of obesity on the pharmacokinetics of drugs in humans. Clin. Pharm. 2010, 49, 71–87. [Google Scholar] [CrossRef]
- Rao, P.S.; Kumar, S. Chronic Effects of Ethanol and/or Darunavir/Ritonavir on U937 Monocytic Cells: Regulation of Cytochrome P450 and Antioxidant Enzymes, Oxidative Stress, and Cytotoxicity. Alcohol. Clin. Exp. Res. 2016, 40, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Wu, L.F.; Deng, F.Y. Exosome: An Emerging Source of Biomarkers for Human Diseases. Curr. Mol. Med. 2019, 19, 387–394. [Google Scholar] [CrossRef]
- Deng, F.; Miller, J. A review on protein markers of exosome from different bio-resources and the antibodies used for characterization. J. Histotechnol. 2019, 42, 226–239. [Google Scholar] [CrossRef]
- Sidhom, K.; Obi, P.O.; Saleem, A. A Review of Exosomal Isolation Methods: Is Size Exclusion Chromatography the Best Option? Int. J. Mol. Sci. 2020, 21, 6466. [Google Scholar] [CrossRef]
- Kumar, S.; Sinha, N.; Gerth, K.A.; Rahman, M.A.; Yallapu, M.M.; Midde, N.M. Specific packaging and circulation of cytochromes P450, especially 2E1 isozyme, in human plasma exosomes and their implications in cellular communications. Biochem. Biophys. Res. Commun. 2017, 491, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.E.; Mezey, E.; Hardwick, J.P.; Salem, N., Jr.; Clemens, D.L.; Song, B.J. Increased ethanol-inducible cytochrome P450-2E1 and cytochrome P450 isoforms in exosomes of alcohol-exposed rodents and patients with alcoholism through oxidative and endoplasmic reticulum stress. Hepatol. Commun. 2017, 1, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Kodidela, S.; Sinha, N.; Haque, S.; Shukla, P.K.; Rao, R.; Kumar, S. Plasma exosomes exacerbate alcohol- and acetaminophen-induced toxicity via CYP2E1 pathway. Sci. Rep. 2019, 9, 6571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.A.; Patters, B.J.; Kodidela, S.; Kumar, S. Extracellular Vesicles: Intercellular Mediators in Alcohol-Induced Pathologies. J. Neuroimmune Pharm. 2020, 15, 409–421. [Google Scholar] [CrossRef]
- Assis, J.B.; Cogliati, B.; Esteves, E.; Capurro, M.L.; Fonseca, D.M.; Sá-Nunes, A. Aedes aegypti mosquito saliva ameliorates acetaminophen-induced liver injury in mice. PLoS ONE 2021, 16, e0245788. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Jiang, Z.; Chang, X.; Xue, H.; Yahefu, W.; Zhang, X. 4-Hydroxyphenylacetic Acid Prevents Acute APAP-Induced Liver Injury by Increasing Phase II and Antioxidant Enzymes in Mice. Front. Pharm. 2018, 9, 653. [Google Scholar] [CrossRef]
- James, L.; Sullivan, J.E.; Roberts, D. The proper use of acetaminophen. Paediatr. Child. Health 2011, 16, 544–547. [Google Scholar] [CrossRef] [Green Version]
- Sodeifian, F.; Seyedalhosseini, Z.S.; Kian, N.; Eftekhari, M.; Najari, S.; Mirsaeidi, M.; Farsi, Y.; Nasiri, M.J. Drug-Induced Liver Injury in COVID-19 Patients: A Systematic Review. Front. Med. 2021, 8, 731436. [Google Scholar] [CrossRef]
- Jiang, S.; Vozmediano, V.; Abdel-Rahman, S.M.; Schmidt, S.; James, L.P. Acetaminophen Protein Adducts in Hospitalized Children Receiving Multiple Doses of Acetaminophen. J. Clin. Pharm. 2019, 59, 1291–1299. [Google Scholar] [CrossRef]
- Massart, J.; Begriche, K.; Fromenty, B. Cytochrome P450 2E1 should not be neglected for acetaminophen-induced liver injury in metabolic diseases with altered insulin levels or glucose homeostasis. Clin. Res. Hepatol. Gastroenterol. 2021, 45, 101470. [Google Scholar] [CrossRef]
- Jiang, Z.; Guo, X.; Zhang, K.; Sekaran, G.; Cao, B.; Zhao, Q.; Zhang, S.; Kirby, G.M.; Zhang, X. The Essential Oils and Eucalyptol From Artemisia vulgaris L. Prevent Acetaminophen-Induced Liver Injury by Activating Nrf2-Keap1 and Enhancing APAP Clearance Through Non-Toxic Metabolic Pathway. Front. Pharm. 2019, 10, 782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Sun, J.; Li, D.; Hu, Y.; Yu, X.; Hua, H.; Jing, X.; Chen, F.; Jia, Z.; Xu, J. Shikonin attenuates acetaminophen-induced acute liver injury via inhibition of oxidative stress and inflammation. Biomed. Pharm. 2019, 112, 108704. [Google Scholar] [CrossRef] [PubMed]
- Alonso, E.M.; James, L.P.; Zhang, S.; Squires, R.H. Acetaminophen Adducts Detected in Serum of Pediatric Patients With Acute Liver Failure. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Ferron, P.J.; Gicquel, T.; Mégarbane, B.; Clément, B.; Fromenty, B. Treatments in Covid-19 patients with pre-existing metabolic dysfunction-associated fatty liver disease: A potential threat for drug-induced liver injury? Biochimie 2020, 179, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, J.; Pan, L.; Chen, L.; Wang, Y.; Liu, X.; You, L.; Jia, Y.; Hu, C. Protective effect of 7,3’,4’-flavon-3-ol (fisetin) on acetaminophen-induced hepatotoxicity in vitro and in vivo. Phytomedicine 2019, 58, 152865. [Google Scholar] [CrossRef] [PubMed]
- Ginting, C.N.; Lister, I.N.E.; Girsang, E.; Widowati, W.; Yusepany, D.T.; Azizah, A.M.; Kusuma, H.S.W. Hepatotoxicity prevention in Acetaminophen-induced HepG2 cells by red betel (Piper crocatum Ruiz and Pav) extract from Indonesia via antioxidant, anti-inflammatory, and anti-necrotic. Heliyon 2021, 7, e05620. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Jin, S.W.; Lee, G.H.; Han, E.H.; Hwang, Y.P.; Jeong, H.G. Rutaecarpine Protects against Acetaminophen-Induced Acute Liver Injury in Mice by Activating Antioxidant Enzymes. Antioxidant 2021, 10, 86. [Google Scholar] [CrossRef]
- Ko, J.W.; Shin, J.Y.; Kim, J.W.; Park, S.H.; Shin, N.R.; Lee, I.C.; Shin, I.S.; Moon, C.; Kim, S.H.; Kim, S.H.; et al. Protective effects of diallyl disulfide against acetaminophen-induced nephrotoxicity: A possible role of CYP2E1 and NF-κB. Food. Chem. Toxicol. 2017, 102, 156–165. [Google Scholar] [CrossRef]
- Han, J.H.; Ju, J.H.; Lee, Y.S.; Park, J.H.; Yeo, I.J.; Park, M.H.; Roh, Y.S.; Han, S.B.; Hong, J.T. Astaxanthin alleviated ethanol-induced liver injury by inhibition of oxidative stress and inflammatory responses via blocking of STAT3 activity. Sci. Rep. 2018, 8, 14090. [Google Scholar] [CrossRef]
- Ekstrom, G.; Ingelman-Sundberg, M. Rat liver microsomal NADPH-supported oxidase activity and lipid peroxidation dependent on ethanol-inducible cytochrome P-450 (P-450IIE1). Biochem. Pharm. 1989, 38, 1313–1319. [Google Scholar] [CrossRef]
- Tomasi, M.L.; Ramani, K.; Ryoo, M.; Cossu, C.; Floris, A.; Murray, B.J.; Iglesias-Ara, A.; Spissu, Y.; Mavila, N. SUMOylation regulates cytochrome P450 2E1 expression and activity in alcoholic liver disease. FASEB J. 2018, 32, 3278–3288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, C.; Yu, B.; Jiang, J.D.; Kong, W.J. Gastrodin Protects against Ethanol-Induced Liver Injury and Apoptosis in HepG2 Cells and Animal Models of Alcoholic Liver Disease. Biol. Pharm. Bull. 2018, 41, 670–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagappan, A.; Kim, J.H.; Jung, D.Y.; Jung, M.H. Cryptotanshinone from the Salvia miltiorrhiza Bunge Attenuates Ethanol-Induced Liver Injury by Activation of AMPK/SIRT1 and Nrf2 Signaling Pathways. Int. J. Mol. Sci. 2019, 21, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avila, M.A.; Dufour, J.F.; Gerbes, A.L.; Zoulim, F.; Bataller, R.; Burra, P.; Cortez-Pinto, H.; Gao, B.; Gilmore, I.; Mathurin, P.; et al. Recent advances in alcohol-related liver disease (ALD): Summary of a Gut round table meeting. Gut 2020, 69, 764–780. [Google Scholar] [CrossRef]
- Lu, Y.; Wu, D.; Wang, X.; Ward, S.C.; Cederbaum, A.I. Chronic alcohol-induced liver injury and oxidant stress are decreased in cytochrome P4502E1 knockout mice and restored in humanized cytochrome P4502E1 knock-in mice. Free Radic. Biol. Med. 2010, 49, 1406–1416. [Google Scholar] [CrossRef] [Green Version]
- Gouillon, Z.; Lucas, D.; Li, J.; Hagbjork, A.L.; French, B.A.; Fu, P.; Fang, C.; Ingelman-Sundberg, M.; Donohue, T.M., Jr.; French, S.W. Inhibition of ethanol-induced liver disease in the intragastric feeding rat model by chlormethiazole. Proc. Soc. Exp. Biol. Med. 2000, 224, 302–308. [Google Scholar] [CrossRef]
- Morgan, K.; French, S.W.; Morgan, T.R. Production of a cytochrome P450 2E1 transgenic mouse and initial evaluation of alcoholic liver damage. Hepatology 2002, 36, 122–134. [Google Scholar] [CrossRef]
- Bansal, S.; Anandatheerthavarada, H.K.; Prabu, G.K.; Milne, G.L.; Martin, M.V.; Guengerich, F.P.; Avadhani, N.G. Human cytochrome P450 2E1 mutations that alter mitochondrial targeting efficiency and susceptibility to ethanol-induced toxicity in cellular models. J. Biol. Chem. 2013, 288, 12627–12644. [Google Scholar] [CrossRef] [Green Version]
- Bansal, S.; Liu, C.P.; Sepuri, N.B.; Anandatheerthavarada, H.K.; Selvaraj, V.; Hoek, J.; Milne, G.L.; Guengerich, F.P.; Avadhani, N.G. Mitochondria-targeted cytochrome P450 2E1 induces oxidative damage and augments alcohol-mediated oxidative stress. J. Biol. Chem. 2010, 285, 24609–24619. [Google Scholar] [CrossRef] [Green Version]
- Chacko, B.K.; Srivastava, A.; Johnson, M.S.; Benavides, G.A.; Chang, M.J.; Ye, Y.; Jhala, N.; Murphy, M.P.; Kalyanaraman, B.; Darley-Usmar, V.M. Mitochondria-targeted ubiquinone (MitoQ) decreases ethanol-dependent micro and macro hepatosteatosis. Hepatology 2011, 54, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Chen, C.; Zhang, X. Drug-drug interaction of acetaminophen and roxithromycin with the cocktail of cytochrome P450 and hepatotoxicity in rats. Int. J. Med. Sci. 2020, 17, 414–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Sun, S.; Rui, H.; Li, X. In vitro inhibitory effects of dihydromyricetin on human liver cytochrome P450 enzymes. Pharm. Biol. 2017, 55, 1868–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Rao, P.S.; Earla, R.; Kumar, A. Drug-drug interactions between anti-retroviral therapies and drugs of abuse in HIV systems. Expert Opin. Drug Metab. Toxicol. 2015, 11, 343–355. [Google Scholar] [CrossRef] [Green Version]
- Abbott, K.L.; Flannery, P.C.; Gill, K.S.; Boothe, D.M.; Dhanasekaran, M.; Mani, S.; Pondugula, S.R. Adverse pharmacokinetic interactions between illicit substances and clinical drugs. Drug Metab. Rev. 2020, 52, 44–65. [Google Scholar] [CrossRef] [PubMed]
- Shakeel, F.; Fang, F.; Kidwell, K.M.; Marcath, L.A.; Hertz, D.L. Comparison of eight screening tools to detect interactions between herbal supplements and oncology agents. J. Oncol. Pharm. Pr. 2020, 26, 1843–1849. [Google Scholar] [CrossRef]
- Shah, R.R. Mechanistic basis of adverse drug reactions: The perils of inappropriate dose schedules. Expert Opin. Drug Saf. 2005, 4, 103–128. [Google Scholar] [CrossRef]
- Dresser, G.K.; Spence, J.D.; Bailey, D.G. Pharmacokinetic-pharmacodynamic consequences and clinical relevance of cytochrome P450 3A4 inhibition. Clin. Pharm. 2000, 38, 41–57. [Google Scholar] [CrossRef]
- Torres, S.; Baulies, A.; Insausti-Urkia, N.; Alarcón-Vila, C.; Fucho, R.; Solsona-Vilarrasa, E.; Núñez, S.; Robles, D.; Ribas, V.; Wakefield, L.; et al. Endoplasmic Reticulum Stress-Induced Upregulation of STARD1 Promotes Acetaminophen-Induced Acute Liver Failure. Gastroenterology 2019, 157, 552–568. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Feng, P.; Gao, X.; Wang, B.; Gou, C.; Bian, R. In vitro inhibitory effects of cepharanthine on human liver cytochrome P450 enzymes. Pharm. Biol. 2020, 58, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Jiang, W.; Zhou, J.; Xue, X.; Yin, C. Anemarsaponin BII inhibits the activity of CYP3A4, 2D6, and 2E1 with human liver microsomes. Pharm. Biol. 2020, 58, 1064–1069. [Google Scholar] [CrossRef]
- Su, Q.; Kuang, W.; Hao, W.; Liang, J.; Wu, L.; Tang, C.; Wang, Y.; Liu, T. Antituberculosis Drugs (Rifampicin and Isoniazid) Induce Liver Injury by Regulating NLRP3 Inflammasomes. Mediat. Inflamm. 2021, 2021, 8086253. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Cederbaum, A.I. Alcohol Upregulation of CYP2A5: Role of Reactive Oxygen Species. React. Oxyg. Species 2016, 1, 117–130. [Google Scholar] [CrossRef]
- Lin, Q.; Kang, X.; Li, X.; Wang, T.; Liu, F.; Jia, J.; Jin, Z.; Xue, Y. NF-κB-mediated regulation of rat CYP2E1 by two independent signaling pathways. PLoS ONE 2019, 14, e0225531. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. Inhibition of CYP2E1 with natural agents may be a feasible strategy for minimizing the hepatotoxicity of ethanol. Med. Hypotheses 2001, 56, 8–11. [Google Scholar] [CrossRef]
- Jiang, Z.; Jiang, X.; Li, C.; Xue, H.; Zhang, X. Development of an IgY Antibody-Based Immunoassay for the Screening of the CYP2E1 Inhibitor/Enhancer from Herbal Medicines. Front. Pharm. 2016, 7, 502. [Google Scholar] [CrossRef] [Green Version]
- Leung, T.M.; Lu, Y. Alcoholic Liver Disease: From CYP2E1 to CYP2A5. Curr. Mol. Pharm. 2017, 10, 172–178. [Google Scholar] [CrossRef]
- Inoue, D.; Usui, R.; Nitta, K.; Koike, M. A case of acetaminophen-induced acute tubulointerstitial nephritis in adult. CEN Case Rep. 2017, 6, 185–188. [Google Scholar] [CrossRef] [Green Version]
- Akakpo, J.Y.; Ramachandran, A.; Orhan, H.; Curry, S.C.; Rumack, B.H.; Jaeschke, H. 4-methylpyrazole protects against acetaminophen-induced acute kidney injury. Toxicol. Appl. Pharm. 2020, 409, 115317. [Google Scholar] [CrossRef]
- Hua, H.; Ge, X.; Wu, M.; Zhu, C.; Chen, L.; Yang, G.; Zhang, Y.; Huang, S.; Zhang, A.; Jia, Z. Rotenone Protects Against Acetaminophen-Induced Kidney Injury by Attenuating Oxidative Stress and Inflammation. Kidney Blood Press. Res. 2018, 43, 1297–1309. [Google Scholar] [CrossRef]
- Cheng, S.; Wu, T.; Li, Y.; Huang, J.; Cai, T. Romidepsin (FK228) in a Mouse Model of Lipopolysaccharide-Induced Acute Kidney Injury is Associated with Down-Regulation of the CYP2E1 Gene. Med. Sci. Monit. 2020, 26, e918528. [Google Scholar] [CrossRef]
- Un, H.; Ugan, R.A.; Kose, D.; Bayir, Y.; Cadirci, E.; Selli, J.; Halici, Z. A novel effect of Aprepitant: Protection for cisplatin-induced nephrotoxicity and hepatotoxicity. Eur. J. Pharm. 2020, 880, 173168. [Google Scholar] [CrossRef] [PubMed]
- Toselli, F.; Dodd, P.R.; Gillam, E.M. Emerging roles for brain drug-metabolizing cytochrome P450 enzymes in neuropsychiatric conditions and responses to drugs. Drug Metab. Rev. 2016, 48, 379–404. [Google Scholar] [CrossRef] [PubMed]
- McMillan, D.M.; Tyndale, R.F. CYP-mediated drug metabolism in the brain impacts drug response. Pharmacol. Ther. 2018, 184, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Kuban, W.; Daniel, W.A. Cytochrome P450 expression and regulation in the brain. Drug Metab. Rev. 2021, 53, 1–29. [Google Scholar] [CrossRef]
- García-Suástegui, W.A.; Ramos-Chávez, L.A.; Rubio-Osornio, M.; Calvillo-Velasco, M.; Atzin-Méndez, J.A.; Guevara, J.; Silva-Adaya, D. The Role of CYP2E1 in the Drug Metabolism or Bioactivation in the Brain. Oxid. Med. Cell. Longev. 2017, 2017, 4680732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Zhu, H.; Kindy, M.S.; Taheri, S. Cytochrome P450 CYP2E1 Suppression Ameliorates Cerebral Ischemia Reperfusion Injury. Antioxidant 2021, 10, 52. [Google Scholar] [CrossRef]
- Bonacini, M. Alcohol use among patients with HIV infection. Ann. Hepatol. 2011, 10, 502–507. [Google Scholar] [CrossRef]
- NIAAA. Alcohol Alert No. 57 September 2002. Available online: https://pubs.niaaa.nih.gov/publications/aa57.htm (accessed on 5 April 2021).
- Tang, H.; Sun, Q.; Wang, T.; Liao, Y.; Wang, G.; Zhao, F.; Jin, Y. Upregulation of CYP2E1 expression causes oxidative damage induced by 2-chloroethanol in primary cultured rat astrocytes. Neurotoxicology 2019, 75, 233–244. [Google Scholar] [CrossRef]
- Malik, D.E.; David, R.M.; Gooderham, N.J. Ethanol potentiates the genotoxicity of the food-derived mammary carcinogen PhIP in human estrogen receptor-positive mammary cells: Mechanistic support for lifestyle factors (cooked red meat and ethanol) associated with mammary cancer. Arch. Toxicol. 2018, 92, 1639–1655. [Google Scholar] [CrossRef] [Green Version]
- Harnagea-Theophilus, E.; Gadd Sl Fau-Knight-Trent, A.H.; Knight-Trent Ah Fau-DeGeorge, G.L.; DeGeorge Gl Fau-Miller, M.R.; Miller, M.R. Acetaminophen-induced proliferation of breast cancer cells involves estrogen receptors. Toxicol. Appl. Pharm. 1999, 155, 273–279. [Google Scholar] [CrossRef]
- Leung, T.; Rajendran, R.; Singh, S.; Garva, R.; Krstic-Demonacos, M.; Demonacos, C. Cytochrome P450 2E1 (CYP2E1) regulates the response to oxidative stress and migration of breast cancer cells. Breast Cancer Res. 2013, 15, R107. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Zhang, F.; Chen, D.; Su, K.; Zhang, L.; Jiang, R. In vitro inhibitory effects of ganoderic acid A on human liver cytochrome P450 enzymes. Pharm. Biol. 2020, 58, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Vuilleumier, N.; Rossier, M.F.; Chiappe, A.; Degoumois, F.; Dayer, P.; Mermillod, B.; Nicod, L.; Desmeules, J.; Hochstrasser, D. CYP2E1 genotype and isoniazid-induced hepatotoxicity in patients treated for latent tuberculosis. Eur. J. Clin. Pharm. 2006, 62, 423–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, G.N.; Magno, L.A.; Santana, C.V.; Konstantinovas, C.; Saito, S.T.; Machado, M.; Di Pietro, G.; Bastos-Rodrigues, L.; Miranda, D.M.; De Marco, L.A.; et al. Genetic interaction between NAT2, GSTM1, GSTT1, CYP2E1, and environmental factors is associated with adverse reactions to anti-tuberculosis drugs. Mol. Diagn. Ther. 2012, 16, 241–250. [Google Scholar] [CrossRef]
- Kang, T.S.; Woo, S.W.; Park, H.J.; Lee, Y.; Roh, J. Comparison of genetic polymorphisms of CYP2E1, ADH2, and ALDH2 genes involved in alcohol metabolism in Koreans and four other ethnic groups. J. Clin. Pharm. Ther. 2009, 34, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.H.; Zhang, H.; Lai, M.P.; Chen, S.L.; Wu, M.; Shen, N. Single-nucleotide polymorphisms and haplotype of CYP2E1 gene associated with systemic lupus erythematosus in Chinese population. Arthritis Res. Ther. 2011, 13, R11. [Google Scholar] [CrossRef] [Green Version]
- Guengerich, F.P. Cytochrome P450 2E1 and its roles in disease. Chem. Biol. Interact. 2020, 322, 109056. [Google Scholar] [CrossRef]
- Ueshima, Y.; Tsutsumi, M.; Takase, S.; Matsuda, Y.; Kawahara, H. Acetaminophen metabolism in patients with different cytochrome P-4502E1 genotypes. Alcohol. Clin. Exp. Res. 1996, 20, 25A–28A. [Google Scholar] [CrossRef]
- Tsutsumi, M.; Takada, A.; Wang, J.S. Genetic polymorphisms of cytochrome P4502E1 related to the development of alcoholic liver disease. Gastroenterology 1994, 107, 1430–1435. [Google Scholar] [CrossRef]
- Bedada, S.K.; Neerati, P. Resveratrol Pretreatment Affects CYP2E1 Activity of Chlorzoxazone in Healthy Human Volunteers. Phytother. Res. 2016, 30, 463–468. [Google Scholar] [CrossRef]
- Bedada, S.K.; Boga, P.K. Effect of piperine on CYP2E1 enzyme activity of chlorzoxazone in healthy volunteers. Xenobiotica 2017, 47, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Bedada, S.K.; Neerati, P. The effect of quercetin on the pharmacokinetics of chlorzoxazone, a CYP2E1 substrate, in healthy subjects. Eur. J. Clin. Pharm. 2018, 74, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, I.; Desager, J.P.; Horsmans, Y. Inhibition of chlorzoxazone metabolism, a clinical probe for CYP2E1, by a single ingestion of watercress. Clin. Pharmacol. Ther. 1998, 64, 144–149. [Google Scholar] [CrossRef]
- Sun, X.; He, L.; Bi, H.; Huang, M.; Xiang, E.; Li, X.; Wang, H.; Guo, Y. Prenatal ethanol exposure induces dynamic changes of expression and activity of hepatic cytochrome P450 isoforms in male rat offspring. Reprod. Toxicol. 2022, 109, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Wang, Y.; He, Y.; Pan, J.; Jin, Y.; Zheng, L.; Huang, Y.; Li, Y.; Liu, W. Aidi injection altered the activity of CYP2D4, CYP1A2, CYP2C19, CYP3A2, CYP2E1 and CYP2C11 in normal and diethylnitrosamine-induced hepatocellular carcinoma in rats. J. Ethnopharmacol. 2022, 286, 114930. [Google Scholar] [CrossRef]
- Bolla, L.; Srivastava, P.; Ravichandiran, V.; Nanjappan, S.K. Cytochrome P450 and P-gp Mediated Herb-Drug Interactions and Molecular Docking Studies of Garcinol. Membranes 2021, 11, 992. [Google Scholar] [CrossRef]
- Trousil, S.; Lee, P.; Edwards, R.J.; Maslen, L.; Lozan-Kuehne, J.P.; Ramaswami, R.; Aboagye, E.O.; Clarke, S.; Liddle, C.; Sharma, R. Altered cytochrome 2E1 and 3A P450-dependent drug metabolism in advanced ovarian cancer correlates to tumour-associated inflammation. Br. J. Pharm. 2019, 176, 3712–3722. [Google Scholar] [CrossRef]
- Escudier, B.; Dorval, T.; Chaput, N.; André, F.; Caby, M.P.; Novault, S.; Flament, C.; Leboulaire, C.; Borg, C.; Amigorena, S.; et al. Vaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived-exosomes: Results of thefirst phase I clinical trial. J. Transl. Med. 2005, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Hall, S.D.; Maya, J.F.; Li, L.; Asghar, A.; Gorski, J.C. Diabetes mellitus increases the in vivo activity of cytochrome P450 2E1 in humans. Br. J. Clin. Pharm. 2003, 55, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Loizou, G.D.; Cocker, J. The effects of alcohol and diallyl sulphide on CYP2E1 activity in humans: A phenotyping study using chlorzoxazone. Hum. Exp. Toxicol. 2001, 20, 321–327. [Google Scholar] [CrossRef]
- Oneta, C.M.; Lieber, C.S.; Li, J.; Rüttimann, S.; Schmid, B.; Lattmann, J.; Rosman, A.S.; Seitz, H.K. Dynamics of cytochrome P4502E1 activity in man: Induction by ethanol and disappearance during withdrawal phase. J. Hepatol. 2002, 36, 47–52. [Google Scholar] [CrossRef]
- Polonikov, A.V.; Ivanov, V.P.; Solodilova, M.A. CYP2E1 gene promoter polymorphism -1293G>C increases the risk of essential hypertension in men with alcohol abuse. Bull. Exp. Biol. Med. 2013, 155, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.S.; Chern, H.D.; Su, W.J.; Wu, J.C.; Chang, S.C.; Chiang, C.H.; Chang, F.Y.; Lee, S.D. Cytochrome P450 2E1 genotype and the susceptibility to antituberculosis drug-induced hepatitis. Hepatology 2003, 37, 924–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eap, C.B.; Schnyder, C.; Besson, J.; Savary, L.; Buclin, T. Inhibition of CYP2E1 by chlormethiazole as measured by chlorzoxazone pharmacokinetics in patients with alcoholism and in healthy volunteers. Clin. Pharmacol. Ther. 1998, 64, 52–57. [Google Scholar] [CrossRef]
- Hohmann, N.; Blank, A.; Burhenne, J.; Suzuki, Y.; Mikus, G.; Haefeli, W.E. Simultaneous phenotyping of CYP2E1 and CYP3A using oral chlorzoxazone and midazolam microdoses. Br. J. Clin. Pharm. 2019, 85, 2310–2320. [Google Scholar] [CrossRef]
- Palmer, J.L.; Scott, R.J.; Gibson, A.; Dickins, M.; Pleasance, S. An interaction between the cytochrome P450 probe substrates chlorzoxazone (CYP2E1) and midazolam (CYP3A). Br. J. Clin. Pharm. 2001, 52, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Frye, R.F.; Branch, R.A. Effect of chronic disulfiram administration on the activities of CYP1A2, CYP2C19, CYP2D6, CYP2E1, and N-acetyltransferase in healthy human subjects. Br. J. Clin. Pharm. 2002, 53, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Manyike, P.T.; Kharasch, E.D.; Kalhorn, T.F.; Slattery, J.T. Contribution of CYP2E1 and CYP3A to acetaminophen reactive metabolite formation. Clin. Pharmacol. Ther. 2000, 67, 275–282. [Google Scholar] [CrossRef]
- Mitra, A.K.; Thummel, K.E.; Kalhorn, T.F.; Kharasch, E.D.; Unadkat, J.D.; Slattery, J.T. Metabolism of dapsone to its hydroxylamine by CYP2E1 in vitro and in vivo. Clin. Pharmacol. Ther. 1995, 58, 556–566. [Google Scholar] [CrossRef]
- Frye, R.F.; Tammara, B.; Cowart, T.D.; Bramer, S.L. Effect of disulfiram-mediated CYP2E1 inhibition on the disposition of vesnarinone. J. Clin. Pharm. 1999, 39, 1177–1183. [Google Scholar]
- Desager, J.P.; Golnez, J.L.; De Buck, C.; Horsmans, Y. Watercress has no Importance for the elimination of ethanol by CYP2E1 inhibition. Pharm. Toxicol. 2002, 91, 103–105. [Google Scholar] [CrossRef] [PubMed]
- Makarova, J.; Turchinovich, A.; Shkurnikov, M.; Tonevitsky, A. Extracellular miRNAs and Cell-Cell Communication: Problems and Prospects. Trends Biochem. Sci. 2021, 46, 640–651. [Google Scholar] [CrossRef] [PubMed]
- Caruso Bavisotto, C.; Scalia, F.; Marino Gammazza, A.; Carlisi, D.; Bucchieri, F.; Conway de Macario, E.; Macario, A.J.L.; Cappello, F.; Campanella, C. Extracellular Vesicle-Mediated Cell⁻Cell Communication in the Nervous System: Focus on Neurological Diseases. Int. J. Mol. Sci. 2019, 20, 434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerth, K.; Kodidela, S.; Mahon, M.; Haque, S.; Verma, N.; Kumar, S. Circulating Extracellular Vesicles Containing Xenobiotic Metabolizing CYP Enzymes and Their Potential Roles in Extrahepatic Cells Via Cell-Cell Interactions. Int. J. Mol. Sci. 2019, 20, 6178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbar, N.; Azzimato, V.; Choudhury, R.P.; Aouadi, M. Extracellular vesicles in metabolic disease. Diabetologia 2019, 62, 2179–2187. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Zhou, L.; Zhi, K.; Raji, B.; Pernell, S.; Tadrous, E.; Kodidela, S.; Nookala, A.; Kochat, H.; Kumar, S. Challenges in Biomaterial-Based Drug Delivery Approach for the Treatment of Neurodegenerative Diseases: Opportunities for Extracellular Vesicles. Int. J. Mol. Sci. 2020, 22, 138. [Google Scholar] [CrossRef]
- Zhi, K.; Kumar, A.; Raji, B.; Kochat, H.; Kumar, S. Formulation, manufacturing and regulatory strategies for extracellular vesicles-based drug products for targeted therapy of central nervous system diseases. Expert Rev. Precis. Med. Drug Dev. 2020, 5, 469–481. [Google Scholar] [CrossRef]
- Haque, S.; Sinha, N.; Ranjit, S.; Midde, N.M.; Kashanchi, F.; Kumar, S. Monocyte-derived exosomes upon exposure to cigarette smoke condensate alter their characteristics and show protective effect against cytotoxicity and HIV-1 replication. Sci. Rep. 2017, 7, 16120. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.E.; Song, B.J.; Akbar, M.; Baek, M.C. Extracellular vesicles as potential biomarkers for alcohol- and drug-induced liver injury and their therapeutic applications. Pharmacol. Ther. 2018, 187, 180–194. [Google Scholar] [CrossRef]
- Cho, Y.E.; Seo, W.; Kim, D.K.; Moon, P.G.; Kim, S.H.; Lee, B.H.; Song, B.J.; Baek, M.C. Exogenous exosomes from mice with acetaminophen-induced liver injury promote toxicity in the recipient hepatocytes and mice. Sci. Rep. 2018, 8, 16070. [Google Scholar] [CrossRef] [Green Version]
- Thietart, S.; Rautou, P.E. Extracellular vesicles as biomarkers in liver diseases: A clinician’s point of view. J. Hepatol. 2020, 73, 1507–1525. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Gong, Y.; Kumar, S. In vitro evaluation of structural analogs of diallyl sulfide as novel CYP2E1 inhibitors for their protective effect against xenobiotic-induced toxicity and HIV replication. Toxicol. Lett. 2018, 292, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Kodidela, S.; Ranjit, S.; Sinha, N.; McArthur, C.; Kumar, A.; Kumar, S. Cytokine profiling of exosomes derived from the plasma of HIV-infected alcohol drinkers and cigarette smokers. PLoS ONE 2018, 13, e0201144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodidela, S.; Wang, Y.; Patters, B.J.; Gong, Y.; Sinha, N.; Ranjit, S.; Gerth, K.; Haque, S.; Cory, T.; McArthur, C.; et al. Proteomic Profiling of Exosomes Derived from Plasma of HIV-Infected Alcohol Drinkers and Cigarette Smokers. J. Neuroimmune Pharm. 2020, 15, 501–519. [Google Scholar] [CrossRef]
- Diesinger, T.; Buko, V.; Lautwein, A.; Dvorsky, R.; Belonovskaya, E.; Lukivskaya, O.; Naruta, E.; Kirko, S.; Andreev, V.; Buckert, D.; et al. Drug targeting CYP2E1 for the treatment of early-stage alcoholic steatohepatitis. PLoS ONE 2020, 15, e0235990. [Google Scholar] [CrossRef]
- Rao, P.S.; Midde, N.M.; Miller, D.D.; Chauhan, S.; Kumar, A.; Kumar, S. Diallyl Sulfide: Potential Use in Novel Therapeutic Interventions in Alcohol, Drugs, and Disease Mediated Cellular Toxicity by Targeting Cytochrome P450 2E1. Curr. Drug Metab. 2015, 16, 486–503. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.S.; Chhabra, S.K.; Hong, J.Y.; Smith, T.J. Mechanisms of inhibition of chemical toxicity and carcinogenesis by diallyl sulfide (DAS) and related compounds from garlic. J. Nutr. 2001, 131, 1041s–1045s. [Google Scholar] [CrossRef] [Green Version]
- Brady, J.F.; Li, D.C.; Ishizaki, H.; Yang, C.S. Effect of diallyl sulfide on rat liver microsomal nitrosamine metabolism and other monooxygenase activities. Cancer Res. 1988, 48, 5937–5940. [Google Scholar]
- Rahman, M.A.; Midde, N.M.; Wu, X.; Li, W.; Kumar, S. Kinetic characterizations of diallyl sulfide analogs for their novel role as CYP2E1 enzyme inhibitors. Pharm. Res. Perspect. 2017, 5, e00362. [Google Scholar] [CrossRef] [Green Version]
- Kodidela, S.; Gerth, K.; Sinha, N.; Kumar, A.; Kumar, P.; Kumar, S. Circulatory Astrocyte and Neuronal EVs as Potential Biomarkers of Neurological Dysfunction in HIV-Infected Subjects and Alcohol/Tobacco Users. Diagnostic 2020, 10, 349. [Google Scholar] [CrossRef]
- Bergh, A.F.; Strobel, H.W. Reconstitution of the brain mixed function oxidase system: Purification of NADPH-cytochrome P450 reductase and partial purification of cytochrome P450 from whole rat brain. J. Neurochem. 1992, 59, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Riedl, A.G.; Watts, P.M.; Edwards, R.J.; Boobis, A.R.; Jenner, P.; Marsden, C.D. Selective localisation of P450 enzymes and NADPH-P450 oxidoreductase in rat basal ganglia using anti-peptide antisera. Brain Res. 1996, 743, 324–328. [Google Scholar] [CrossRef]
- Zhuang, X.; Xiang, X.; Grizzle, W.; Sun, D.; Zhang, S.; Axtell, R.C.; Ju, S.; Mu, J.; Zhang, L.; Steinman, L.; et al. Treatment of brain inflammatory diseases by delivering exosome encapsulated anti-inflammatory drugs from the nasal region to the brain. Mol. Ther. 2011, 19, 1769–1779. [Google Scholar] [CrossRef] [PubMed]
- Nojima, H.; Freeman, C.M.; Schuster, R.M.; Japtok, L.; Kleuser, B.; Edwards, M.J.; Gulbins, E.; Lentsch, A.B. Hepatocyte exosomes mediate liver repair and regeneration via sphingosine-1-phosphate. J. Hepatol. 2016, 64, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Zhou, L.; Lv, D.; Zhu, X.; Tang, H. Exosome-mediated communication in the tumor microenvironment contributes to hepatocellular carcinoma development and progression. J. Hematol. Oncol. 2019, 12, 53. [Google Scholar] [CrossRef]
- Liberti, L.; Breckenridge, A.; Hoekman, J.; Leufkens, H.; Lumpkin, M.; McAuslane, N.; Stolk, P.; Zhi, K.; Rägo, L. Accelerating access to new medicines: Current status of facilitated regulatory pathways used by emerging regulatory authorities. J. Public Health Policy 2016, 37, 315–333. [Google Scholar] [CrossRef]
- Liberti, L.; Breckenridge, A.; Hoekman, J.; Leufkens, H.; Lumpkin, M.; McAuslane, N.; Stolk, P.; Zhi, K.; Rägo, L. Practical aspects of developing, implementing and using facilitated regulatory pathways in the emerging markets. In Proceedings of the Proceedings of the Poster Drug Information Association Annual Meeting, Philadelphia, PA, USA, 28 June 2016. [Google Scholar]
- Zhi, K.; Raji, B.; Nookala, A.R.; Khan, M.M.; Nguyen, X.H.; Sakshi, S.; Pourmotabbed, T.; Yallapu, M.M.; Kochat, H.; Tadrous, E.; et al. PLGA Nanoparticle-Based Formulations to Cross the Blood-Brain Barrier for Drug Delivery: From R&D to cGMP. Pharmaceutics 2021, 13, 500. [Google Scholar] [CrossRef]
- Zhi, K.; Lebo, D.B. A preformulation strategy for the selection of controlled-release components to simulate a subcutaneous implant. Boletín Latinoam. Y Del Caribe De Plantas Med. Y Aromáticas 2020, 19, 344–356. [Google Scholar] [CrossRef]
- Polarine, J.; Chai, R.; Kochat, H.; Pulliam, P.J.; Zhi, K.; Brooks, K. In-Situ Disinfectant Validation Case Study. Am. Pharm. Rev. Microbiol. 2021, 1–6. [Google Scholar] [CrossRef]
- Harting, M.T.; Srivastava, A.K.; Zhaorigetu, S.; Bair, H.; Prabhakara, K.S.; Toledano Furman, N.E.; Vykoukal, J.V.; Ruppert, K.A.; Cox, C.S., Jr.; Olson, S.D. Inflammation-Stimulated Mesenchymal Stromal Cell-Derived Extracellular Vesicles Attenuate Inflammation. Stem Cells 2018, 36, 79–90. [Google Scholar] [CrossRef] [Green Version]


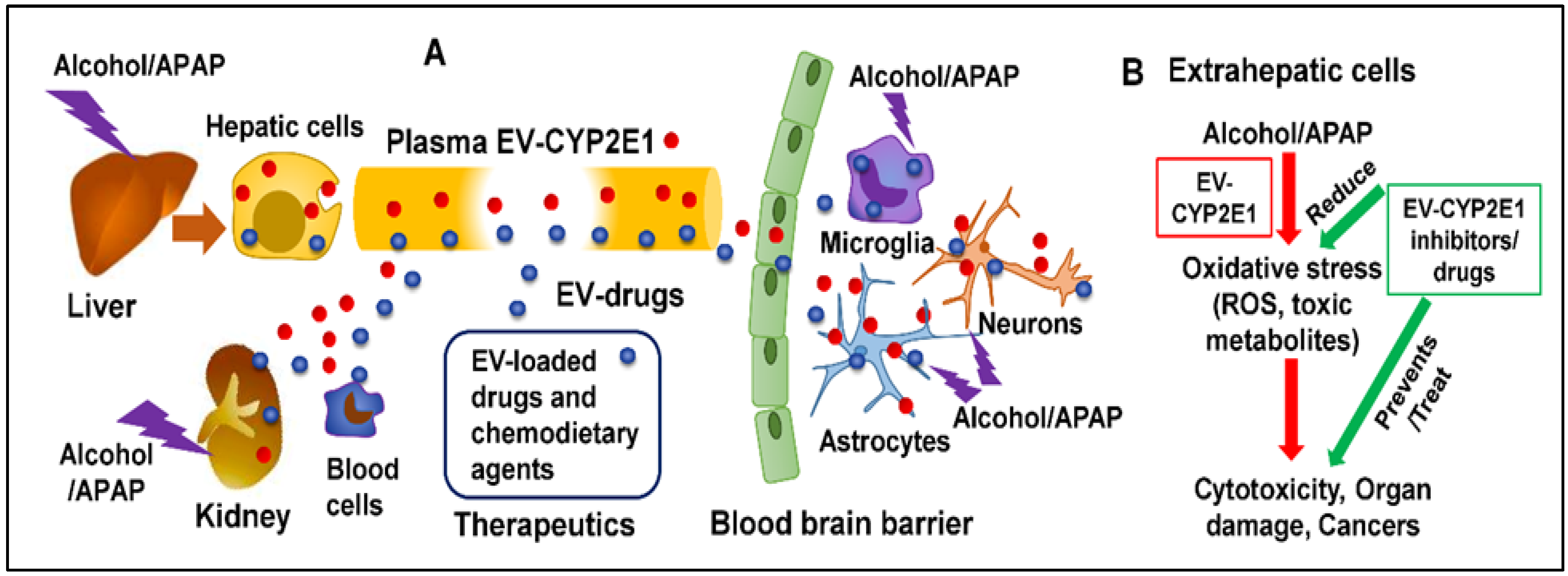{kind=link}
| Reference | Condition/Drug 1 | Effect | Drug 2: Induced Toxicity |
|---|---|---|---|
| [111] | Diabetes | Induction | Chlorzoxazone |
| [112] | Ethanol | Induction | Chlorzoxazone |
| [113] | Ethanol | Induction | Chlorzoxazone |
| [114] | Ethanol | Induction | N/A |
| [115] | Isoniazid | Induction | Chlorzoxazone |
| [116] | Chlormethiazole | Inhibition | Chlorzoxazone |
| [117] | Chlorzoxazone | Inhibition | Midazolam |
| [118] | Chlorzoxazone | Inhibition | Midazolam |
| [119] | Diallyl Sulphide | Inhibition | Chlorzoxazone |
| [113] | Disulfiram | Inhibition | Acetaminophen |
| [120] | Disulfiram | Inhibition | Acetaminophen |
| [121] | Disulfiram | Inhibition | Dapson |
| [122] | Disulfiram | Inhibition | Vesnarinone |
| [123] | Piperine | Inhibition | Chlorzoxazone |
| [104] | Quercetin | Inhibition | Chlorzoxazone |
| [105] | Resveratrol | Inhibition | Chlorzoxazone |
| [103] | Watercress | Inhibition | Chlorzoxazone |
| [106] | Watercress | Inhibition | Ethanol |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, S.; Singla, B.; Singh, A.K.; Thomas-Gooch, S.M.; Zhi, K.; Singh, U.P. Hepatic, Extrahepatic and Extracellular Vesicle Cytochrome P450 2E1 in Alcohol and Acetaminophen-Mediated Adverse Interactions and Potential Treatment Options. Cells 2022, 11, 2620. https://doi.org/10.3390/cells11172620
Kumar S, Singla B, Singh AK, Thomas-Gooch SM, Zhi K, Singh UP. Hepatic, Extrahepatic and Extracellular Vesicle Cytochrome P450 2E1 in Alcohol and Acetaminophen-Mediated Adverse Interactions and Potential Treatment Options. Cells. 2022; 11(17):2620. https://doi.org/10.3390/cells11172620
Chicago/Turabian StyleKumar, Santosh, Bhupesh Singla, Ajay K. Singh, Stacey M. Thomas-Gooch, Kaining Zhi, and Udai P. Singh. 2022. "Hepatic, Extrahepatic and Extracellular Vesicle Cytochrome P450 2E1 in Alcohol and Acetaminophen-Mediated Adverse Interactions and Potential Treatment Options" Cells 11, no. 17: 2620. https://doi.org/10.3390/cells11172620
APA StyleKumar, S., Singla, B., Singh, A. K., Thomas-Gooch, S. M., Zhi, K., & Singh, U. P. (2022). Hepatic, Extrahepatic and Extracellular Vesicle Cytochrome P450 2E1 in Alcohol and Acetaminophen-Mediated Adverse Interactions and Potential Treatment Options. Cells, 11(17), 2620. https://doi.org/10.3390/cells11172620