
Postnatal and Adult Neurogenesis in Mammals, Including Marsupials



Abstract
:1. Introduction
2. Postnatal Developmental Neurogenesis in Mammals
Postnatal Neurogenesis in the Cerebellum
3. Adult Neurogenesis in Mammals
3.1. Adult Neurogenesis in the SVZ/OB
3.2. Does Adult Neurogenesis Occur in the Piriform Cortex?
3.3. Adult-Born Neurons of the Dentate Gyrus (DG)
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ashwell, K.W.S. The Neurobiology of Australian Marsupials. Brain Evolution in the Other Mammalian Radiation; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2010; pp. 18–119. [Google Scholar]
- Chuah, M.I.; Tennent, R.; Teague, R. Developmental anatomy of the primary olfactory pathway in the opossum Monodelphis domestica. Histol. Histopathol. 1997, 12, 799–806. [Google Scholar] [PubMed]
- Sanderson, K.J.; Wilson, P.M. Neurogenesis in septum, amygdala and hippocampus in the marsupial brush-tailed possum (Trichosurus vulpecula). Rev. Bras. Biol. 1997, 57, 323–335. [Google Scholar]
- Sanderson, K.J.; Weller, W.L. Neurogenesis in a marsupial: The brush-tailed possum (Trichosurus vulpecula). II. Sensorimotor pathways. Brain Behav. Evol. 1990, 35, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Butts, T.; Green, M.J.; Wingate, R.J. Development of the cerebellum: Simple steps to make a ‘little brain’. Development 2014, 141, 4031–4041. [Google Scholar] [CrossRef] [PubMed]
- Altman, J.; Das, G.D. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J. Comp. Neurol. 1965, 124, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Bayer, S.A. Joseph Altman (1925–2016): A life in neurodevelopment. J. Comp. Neurol. 2016, 524, 2933–2943. [Google Scholar] [CrossRef]
- Ramon y Cajal, S. Histologie du Systeme Nerveux de L’homme et des Vertebres. Paris Maloine 1909, 2, 80–106. [Google Scholar]
- Kuhn, H.G.; Dickinson-Anson, H.; Gage, F.H. Neurogenesis in the dentate gyrus of the adult rat: Age-related decrease of neuronal progenitor proliferation. J. Neurosci. 1996, 16, 2027–2033. [Google Scholar] [CrossRef]
- Lois, C.; Alvarez-Buylla, A. Long-distance neuronal migration in the adult mammalian brain. Science 1994, 264, 1145–1148. [Google Scholar] [CrossRef]
- Rao, M.S.; Hattiangady, B.; Shetty, A.K. The window and mechanisms of major age-related decline in the production of new neurons within the dentate gyrus of the hippocampus. Aging Cell 2006, 5, 545–558. [Google Scholar] [CrossRef]
- Conover, J.C.; Shook, B.A. Aging of the subventricular zone neural stem cell niche. Aging Dis. 2011, 2, 49–63. [Google Scholar] [PubMed]
- Duque, A.; Arellano, J.I.; Rakic, P. An assessment of the existence of adult neurogenesis in humans and value of its rodent models for neuropsychiatric diseases. Mol. Psychiatry 2022, 27, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, D.M.; Bordey, A.; Bonfanti, L. Noncanonical Sites of Adult Neurogenesis in the Mammalian Brain. Cold Spring Harb. Perspect. Biol. 2015, 7, a018846. [Google Scholar] [CrossRef] [PubMed]
- Jurkowski, M.P.; Bettio, L.K.; Woo, E.; Patten, A.; Yau, S.Y.; Gil-Mohapel, J. Beyond the Hippocampus and the SVZ: Adult Neurogenesis Throughout the Brain. Front. Cell Neurosci. 2020, 14, 576444. [Google Scholar] [CrossRef] [PubMed]
- Ponti, G.; Peretto, P.; Bonfanti, L. Genesis of neuronal and glial progenitors in the cerebellar cortex of peripuberal and adult rabbits. PLoS ONE 2008, 3, e2366. [Google Scholar] [CrossRef]
- Luo, Z.X.; Yuan, C.X.; Meng, Q.J.; Ji, Q. A Jurassic eutherian mammal and divergence of marsupials and placentals. Nature 2011, 476, 442–445. [Google Scholar] [CrossRef]
- Rose, R.W. Embryonic growth rates of marsupials with a note on monotremes. J. Zool. Lond. 1989, 218, 11–16. [Google Scholar] [CrossRef]
- Hickford, D.; Frankenberg, S.; Renfree, M.B. The tammar wallaby, Macropus eugenii: A model kangaroo for the study of developmental and reproductive biology. Cold Spring Harb. Protoc. 2009, 2009, pdb.emo137. [Google Scholar] [CrossRef]
- Smith, K.K. Early development of the neural plate, neural crest and facial region of marsupials. J. Anat. 2001, 199, 121–131. [Google Scholar] [CrossRef]
- Warner, F.J. The development of the diencephalon in Trichosurus vulpecula. Okajimas Folia Anat. Jpn. 1969, 46, 265–295. [Google Scholar] [CrossRef]
- Harman, A.M.; Eastough, N.J.; Beazley, L.D. Development of the visual cortex in a wallaby—Phylogenetic implications. Brain Behav. Evol. 1995, 45, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Renfree, M.B.; Holt, A.B.; Green, S.W.; Carr, J.P.; Cheek, D.B. Ontogeny of the brain in a marsupial (Macropus eugenii) throughout pouch life. I. Brain growth. Brain Behav. Evol. 1982, 20, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Sauerland, C.; Menzies, B.R.; Glatzle, M.; Seeger, J.; Renfree, M.B.; Fietz, S.A. The Basal Radial Glia Occurs in Marsupials and Underlies the Evolution of an Expanded Neocortex in Therian Mammals. Cereb. Cortex 2018, 28, 145–157. [Google Scholar] [CrossRef]
- Harder, J.D.; Stonerook, M.J.; Pondy, J. Gestation and placentation in two New World opossums: Didelphis virginiana and Monodelphis domestica. J. Exp. Zool. 1993, 266, 463–479. [Google Scholar] [CrossRef] [PubMed]
- Saunders, N.R.; Adam, E.; Reader, M.; Møllgård, K. Monodelphis domestica (grey short-tailed opossum): An accessible model for studies of early neocortical development. Anat. Embryol. 1989, 180, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Cardoso-Moreira, M.; Halbert, J.; Valloton, D.; Velten, B.; Chen, C.; Shao, Y.; Liechti, A.; Ascenção, K.; Rummel, C.; Ovchinnikova, S.; et al. Gene expression across mammalian organ development. Nature 2019, 571, 505–509. [Google Scholar] [CrossRef]
- Brunjes, P.C.; Jazaeri, A.; Sutherland, M.J. Olfactory bulb organization and development in Monodelphis domestica (grey short-tailed opossum). J. Comp. Neurol. 1992, 320, 544–554. [Google Scholar] [CrossRef]
- Tepper, B.; Bartkowska, K.; Okrasa, M.; Ngati, S.; Braszak, M.; Turlejski, K.; Djavadian, R. Downregulation of TrkC Receptors Increases Dendritic Arborization of Purkinje Cells in the Developing Cerebellum of the Opossum, Monodelphis domestica. Front. Neuroanat. 2020, 14, 56. [Google Scholar] [CrossRef]
- Reynolds, M.L.; Cavanagh, M.E.; Dziegielewska, K.M.; Hinds, L.A.; Saunders, N.R.; Tyndale-Biscoe, C.H. Postnatal development of the telencephalon of the tammar wallaby (Macropus eugenii). An accessible model of neocortical differentiation. Anat. Embryol 1985, 173, 81–94. [Google Scholar] [CrossRef]
- Curley, J.P.; Jordan, E.R.; Swaney, W.T.; Izraelit, A.; Kammel, S.; Champagne, F.A. The meaning of weaning: Influence of the weaning period on behavioral development in mice. Dev. Neurosci. 2009, 31, 318–331. [Google Scholar] [CrossRef]
- Shimada, M.; Nakamura, T. Time of neuron origin in mouse hypothalamic nuclei. Exp. Neurol. 1973, 41, 163–173. [Google Scholar] [CrossRef]
- Shimogori, T.; A Lee, D.; Miranda-Angulo, A.; Yang, Y.; Wang, H.; Jiang, L.; Yoshida, A.C.; Kataoka, A.; Mashiko, H.; Avetisyan, M.; et al. A genomic atlas of mouse hypothalamic development. Nat. Neurosci. 2010, 13, 767–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, G.; Marotte, L.R.; Mai, J.K.; Ashwell, K.W. Early development of the hypothalamus of a wallaby (Macropus eugenii). J. Comp. Neurol. 2002, 453, 199–215. [Google Scholar] [CrossRef] [PubMed]
- Rivkees, S.A.; Fox, C.A.; Jacobson, C.D.; Reppert, S.M. Anatomic and functional development of the suprachiasmatic nuclei in the gray short-tailed opossum. J. Neurosci. 1988, 8, 4269–4276. [Google Scholar] [CrossRef] [PubMed]
- Noctor, S.C.; Flint, A.C.; Weissman, T.A.; Dammerman, R.S.; Kriegstein, A.R. Neurons derived from radial glial cells establish radial units in neocortex. Nature 2001, 409, 714–720. [Google Scholar] [CrossRef]
- Rakic, P. Evolution of the neocortex: A perspective from developmental biology. Nat. Rev. Neurosci. 2009, 10, 724–735. [Google Scholar] [CrossRef]
- Katz, M.J.; Lasek, R.J.; Silver, J. Ontophyletics of the nervous system: Development of the corpus callosum and evolution of axon tracts. Proc. Natl. Acad. Sci. USA 1983, 80, 5936–5940. [Google Scholar] [CrossRef]
- Cheung, A.F.; Kondo, S.; Abdel-Mannan, O.; Chodroff, R.A.; Sirey, T.M.; Bluy, L.E.; Webber, N.; DeProto, J.; Karlen, S.J.; Krubitzer, L.; et al. The subventricular zone is the developmental milestone of a 6-layered neocortex: Comparisons in metatherian and eutherian mammals. Cereb. Cortex 2010, 20, 1071–1081. [Google Scholar] [CrossRef]
- Puzzolo, E.; Mallamaci, A. Cortico-cerebral histogenesis in the opossum Monodelphis domestica: Generation of a hexalaminar neocortex in the absence of a basal proliferative compartment. Neural. Dev. 2010, 5, 8. [Google Scholar] [CrossRef]
- Bartkowska, K.; Gajerska, M.; Turlejski, K.; Djavadian, R.L. Expression of TrkC receptors in the developing brain of the Monodelphis opossum and its effect on the development of cortical cells. PLoS ONE 2013, 8, e74346. [Google Scholar] [CrossRef]
- Bartkowska, K.; Tepper, B.; Gawda, A.; Jarosik, M.; Sobolewska, P.; Turlejski, K.; Djavadian, R.L. Inhibition of TrkB- and TrkC-Signaling Pathways Affects Neurogenesis in the Opossum Developing Neocortex. Cereb. Cortex 2019, 29, 3666–3675. [Google Scholar] [CrossRef] [PubMed]
- Polleux, F.; Dehay, C.; Kennedy, H. The timetable of laminar neurogenesis contributes to the specification of cortical areas in mouse isocortex. J. Comp. Neurol. 1997, 385, 95–116. [Google Scholar] [CrossRef]
- Sansom, S.N.; Livesey, F.J. Gradients in the brain: The control of the development of form and function in the cerebral cortex. Cold Spring Harb. Perspect. Biol. 2009, 1, a002519. [Google Scholar] [CrossRef]
- Lorente de N´, R. Studies on the structure of the cerebral cortex II. Continuation of the study of the Ammonic system. J. Psychol. Neurol. 1934, 46, 113–177. [Google Scholar]
- Insausti, R. Comparative anatomy of the entorhinal cortex and hippocampus in mammals. Hippocampus 1993, 3, 19–26. [Google Scholar] [CrossRef]
- Amaral, D.G. A Golgi study of cell types in the hilar region of the hippocampus in the rat. J. Comp. Neurol. 1978, 182, 851–914. [Google Scholar] [CrossRef]
- Angevine, J.B., Jr. Time of neuron origin in the hippocampal region. An autoradiographic study in the mouse. Exp. Neurol. Suppl. 1965, 2, 1–70. [Google Scholar]
- Bond, A.M.; Berg, D.A.; Lee, S.; Garcia-Epelboim, A.S.; Adusumilli, V.S.; Ming, G.L.; Song, H. Differential Timing and Coordination of Neurogenesis and Astrogenesis in Developing Mouse Hippocampal Subregions. Brain Sci. 2020, 10, 909. [Google Scholar] [CrossRef]
- Harman, A.M. Development and cell generation in the hippocampus of a marsupial, the quokka wallaby (Setonix brachyurus). Brain Res. Dev. Brain Res. 1997, 104, 41–54. [Google Scholar] [CrossRef]
- Nagayama, S.; Homma, R.; Imamura, F. Neuronal organization of olfactory bulb circuits. Front. Neural Circuits 2014, 8, 98. [Google Scholar] [CrossRef]
- Treloar, H.B.; Miller, A.M.; Ray, A.; Greer, C.A. Development of the Olfactory System. In The Neurobiology of Olfaction; Menini, A., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2010; Chapter 5. Available online: https://www.ncbi.nlm.nih.gov/books/NBK55972/ (accessed on 1 July 2022).
- Brunjes, P.C.; Frazier, L.L. Maturation and plasticity in the olfactory system of vertebrates. Brain Res. 1986, 396, 1–45. [Google Scholar] [CrossRef]
- Ashwell, K.W.; Marotte, L.R.; Cheng, G. Development of the olfactory system in a wallaby (Macropus eugenii). Brain Behav. Evol. 2008, 71, 216–230. [Google Scholar] [CrossRef]
- Sotelo, C. Viewing the cerebellum through the eyes of Ramón Y Cajal. Cerebellum 2008, 7, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Sultan, F.; Glickstein, M. The cerebellum: Comparative and animal studies. Cerebellum 2007, 6, 168–176. [Google Scholar] [CrossRef]
- Sánchez-Villagra, M.R.; Sultan, F. The cerebellum at birth in therian mammals, with special reference to rodents. Brain Behav. Evol. 2002, 59, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Altman, J.; Bayer, S.A. Prenatal development of the cerebellar system in the rat. I. Cytogenesis and histogenesis of the deep nuclei and the cortex of the cerebellum. J. Comp. Neurol. 1978, 179, 23–48. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, C. Cellular and genetic regulation of the development of the cerebellar system. Prog. Neurobiol. 2004, 72, 295–339. [Google Scholar] [CrossRef] [PubMed]
- Altman, J.; Bayer, S.A. Embryonic development of the rat cerebellum. III. Regional differences in the time of origin, migration, and settling of Purkinje cells. J. Comp. Neurol. 1985, 231, 42–65. [Google Scholar] [CrossRef]
- Rakic, P.; Sidman, R.L. Histogenesis of cortical layers in human cerebellum, particularly the lamina dissecans. J. Comp. Neurol. 1970, 139, 473–500. [Google Scholar] [CrossRef]
- Hatten, M.E.; Heintz, N. Mechanisms of neural patterning and specification in the developing cerebellum. Annu. Rev. Neurosci. 1995, 18, 85–408. [Google Scholar] [CrossRef]
- Consalez, G.G.; Goldowitz, D.; Casoni, F.; Hawkes, R. Origins, Development, and Compartmentation of the Granule Cells of the Cerebellum. Front. Neural Circuits 2021, 14, 611841. [Google Scholar] [CrossRef] [PubMed]
- Abrahám, H.; Tornóczky, T.; Kosztolányi, G.; Seress, L. Cell formation in the cortical layers of the developing human cerebellum. Int. J. Dev. Neurosci. 2001, 19, 53–62. [Google Scholar] [CrossRef]
- Grabiec, M.; Turlejski, K.; Djavadian, R.L. The partial 5-HT1A receptor agonist buspirone enhances neurogenesis in the opossum (Monodelphis domestica). Eur. Neuropsychopharmacol. 2009, 19, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Harman, A.; Meyer, P.; Ahmat, A. Neurogenesis in the hippocampus of an adult marsupial. Brain Behav. Evol. 2003, 62, 1–12. [Google Scholar] [CrossRef]
- Kaplan, M.S.; Bell, D.H. Mitotic neuroblasts in the 9-day-old and 11-month-old rodent hippocampus. J. Neurosci. 1984, 4, 1429–1441. [Google Scholar] [CrossRef]
- Guidi, S.; Ciani, E.; Severi, S.; Contestabile, A.; Bartesaghi, R. Postnatal neurogenesis in the dentate gyrus of the guinea pig. Hippocampus 2005, 15, 285–301. [Google Scholar] [CrossRef]
- Amrein, I. Adult hippocampal neurogenesis in natural populations of mammals. Cold Spring Harb. Perspect. Biol. 2015, 7, a021295. [Google Scholar] [CrossRef]
- Bekiari, C.; Grivas, I.; Tsingotjidou, A.; Papadopoulos, G.C. Adult neurogenesis and gliogenesis in the dorsal and ventral canine hippocampus. J. Comp. Neurol. 2020, 528, 1216–1230. [Google Scholar] [CrossRef]
- Altman, J. Autoradiographic investigation of cell proliferation in the brains of rats and cats. Anat. Rec. 1963, 145, 573–591. [Google Scholar] [CrossRef]
- Lévy, F.; Batailler, M.; Meurisse, M.; Migaud, M. Adult Neurogenesis in Sheep: Characterization and Contribution to Reproduction and Behavior. Front. Neurosci. 2017, 11, 570. [Google Scholar] [CrossRef]
- Low, V.F.; Faull, R.L.; Bennet, L.; Gunn, A.J.; Curtis, M.A. Neurogenesis and progenitor cell distribution in the subgranular zone and subventricular zone of the adult sheep brain. Neuroscience 2013, 244, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Bartkowska, K.; Djavadian, R.L.; Taylor, J.R.; Turlejski, K. Generation recruitment and death of brain cells throughout the life cycle of Sorex shrews (Lipotyphla). Eur. J. Neurosci. 2008, 27, 1710–1721. [Google Scholar] [CrossRef] [PubMed]
- Bartkowska, K.; Turlejski, K.; Tepper, B.; Rychlik, L.; Vogel, P.; Djavadian, R. Effects of Brain Size on Adult Neurogenesis in Shrews. Int. J. Mol. Sci. 2021, 22, 7664. [Google Scholar] [CrossRef]
- Patzke, N.; Kaswera, C.; Gilissen, E.; Ihunwo, A.O.; Manger, P.R. Adult neurogenesis in a giant otter shrew (Potamogale velox). Neuroscience 2013, 238, 270–279. [Google Scholar] [CrossRef]
- Gould, E.; McEwen, B.S.; Tanapat, P.; Galea, L.A.; Fuchs, E. Neurogenesis in the dentate gyrus of the adult tree shrew is regulated by psychosocial stress and NMDA receptor activation. J. Neurosci. 1997, 17, 2492–2498. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.Q.; Luo, R.; Tu, T.; Yang, C.; Jiang, J.; Zhang, B.; Bi, R.; Tu, E.; Yao, Y.G.; Yan, X.X. Doublecortin-Expressing Neurons in Chinese Tree Shrew Forebrain Exhibit Mixed Rodent and Primate-Like Topographic Characteristics. Front. Neuroanat. 2021, 15, 727883. [Google Scholar] [CrossRef] [PubMed]
- Patzke, N.; LeRoy, A.; Ngubane, N.W.; Bennett, N.C.; Medger, K.; Gravett, N.; Kaswera-Kyamakya, C.; Gilissen, E.; Chawana, R.; Manger, P.R. The distribution of doublecortin-immunopositive cells in the brains of four afrotherian mammals: The Hottentot golden mole (Amblysomus hottentotus), the rock hyrax (Procavia capensis), the eastern rock sengi (Elephantulus myurus) and the four-toed sengi (Petrodromus tetradactylus). Brain Behav. Evol. 2014, 84, 227–241. [Google Scholar] [CrossRef]
- Bartkowska, K.; Turlejski, K.; Grabiec, M.; Ghazaryan, A.; Yavruoyan, E.; Djavadian, R.L. Adult neurogenesis in the hedgehog (Erinaceus concolor) and mole (Talpa europaea). Brain Behav. Evol. 2010, 76, 128–143. [Google Scholar] [CrossRef]
- Chawana, R.; Alagaili, A.; Patzke, N.; Spocter, M.A.; Mohammed, O.B.; Kaswera, C.; Gilissen, E.; Bennett, N.C.; Ihunwo, A.O.; Manger, P.R. Microbats appear to have adult hippocampal neurogenesis, but post-capture stress causes a rapid decline in the number of neurons expressing doublecortin. Neuroscience 2014, 277, 724–733. [Google Scholar] [CrossRef]
- Chawana, R.; Patzke, N.; Bhagwandin, A.; Kaswera-Kyamakya, C.; Gilissen, E.; Bertelsen, M.F.; Hemingway, J.; Manger, P.R. Adult hippocampal neurogenesis in Egyptian fruit bats from three different environments: Are interpretational variations due to the environment or methodology? J. Comp. Neurol. 2020, 528, 2994–3007. [Google Scholar] [CrossRef]
- Eckenhoff, M.F.; Rakic, P. Nature and fate of proliferative cells in the hippocampal dentate gyrus during the life span of the rhesus monkey. J. Neurosci. 1988, 8, 729–747. [Google Scholar] [CrossRef]
- Kornack, D.R.; Rakic, P. Continuation of neurogenesis in the hippocampus of the adult macaque monkey. Proc. Natl. Acad. Sci. USA 1999, 96, 5768–5773. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, P.S.; Perfilieva, E.; Björk-Eriksson, T.; Alborn, A.M.; Nordborg, C.; Peterson, D.A.; Gage, F.H. Neurogenesis in the adult human hippocampus. Nat. Med. 1998, 4, 1313–1317. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Reeves, A.J.; Fallah, M.; Tanapat, P.; Gross, C.G.; Fuchs, E. Hippocampal neurogenesis in adult Old World primates. Proc. Natl. Acad. Sci. USA 1999, 96, 5263–5267. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.-W.; Li, Z.-Y.; Du, J.; Chen, Y.; Chen, B.-Y.; Wang, T.-T.; Huang, Z.; Hou, S.; Wang, Y. Visualization of Rostral Migratory Stream in the Developing Rat Brain by In Vivo Electroporation. Cell Mol. Neurobiol. 2018, 38, 1067–1079. [Google Scholar] [CrossRef]
- Dayer, A.G.; Ford, A.A.; Cleaver, K.M.; Yassaee, M.; Cameron, H.A. Short-term and long-term survival of new neurons in the rat dentate gyrus. J. Comp. Neurol. 2003, 460, 563–572. [Google Scholar] [CrossRef]
- Schlessinger, A.R.; Cowan, W.M.; Gottlieb, D.I. An autoradiographic study of the time of origin and the pattern of granule cell migration in the dentate gyrus of the rat. J. Comp. Neurol. 1975, 159, 149–175. [Google Scholar] [CrossRef]
- Fukushima, N.; Kato, T.; Li, Z.; Yokouchi, K.; Moriizumi, T. Adult neurogenesis and gliogenesis in the rat olfactory nervous system. Chem. Sens. 2005, 30 (Suppl. S1), i113–i114. [Google Scholar] [CrossRef]
- Coviello, S.; Gramuntell, Y.; Castillo-Gomez, E.; Nacher, J. Effects of Dopamine on the Immature Neurons of the Adult Rat Piriform Cortex. Front. Neurosci. 2020, 14, 574234. [Google Scholar] [CrossRef]
- Xu, Y.; Tamamaki, N.; Noda, T.; Kimura, K.; Itokazu, Y.; Matsumoto, N.; Dezawa, M.; Ide, C. Neurogenesis in the ependymal layer of the adult rat 3rd ventricle. Exp. Neurol. 2005, 192, 251–264. [Google Scholar] [CrossRef]
- Pérez-Martín, M.; Cifuentes, M.; Grondona, J.M.; López-Avalos, M.D.; Gómez-Pinedo, U.; García-Verdugo, J.M.; Fernández-Llebrez, P. IGF-I stimulates neurogenesis in the hypothalamus of adult rats. Eur. J. Neurosci. 2010, 31, 1533–1548. [Google Scholar] [CrossRef] [PubMed]
- Dayer, A.G.; Cleaver, K.M.; Abouantoun, T.; Cameron, H.A. New GABAergic interneurons in the adult neocortex and striatum are generated from different precursors. J. Cell Biol. 2005, 168, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Bauer, S.; Hay, M.; Amilhon, B.; Jean, A.; Moyse, E. In vivo neurogenesis in the dorsal vagal complex of the adult rat brain-stem. Neuroscience 2005, 130, 75–90. [Google Scholar] [CrossRef]
- Batista-Brito, R.; Close, J.; Machold, R.; Fishell, G. The distinct temporal origins of olfactory bulb interneuron subtypes. J. Neurosci. 2008, 28, 3966–3975. [Google Scholar] [CrossRef]
- Matsue, K.; Minakawa, S.; Kashiwagi, T.; Toda, K.; Sato, T.; Shioda, S.; Seki, T. Dentate granule progenitor cell properties are rapidly altered soon after birth. Brain Struct. Funct. 2018, 223, 357–369. [Google Scholar] [CrossRef]
- Olaleye, O.O.; Ihunwo, A.O. Adult neurogenesis in the four-striped mice (Rhabdomys pumilio). Neural. Regen. Res. 2014, 9, 1907–1911. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, L.A.; Ng, K.; Zhou, Q.Y.; Ribak, C.E. Subventricular zone-derived, newly generated neurons populate several olfactory and limbic forebrain regions. Epilepsy Behav. 2009, 14 (Suppl. S1), 74–80. [Google Scholar] [CrossRef] [PubMed]
- Salvi, R.; Steigleder, T.; Schlachetzki, J.C.; Waldmann, E.; Schwab, S.; Winner, B.; Winkler, J.; Kohl, Z. Distinct Effects of Chronic Dopaminergic Stimulation on Hippocampal Neurogenesis and Striatal Doublecortin Expression in Adult Mice. Front. Neurosci. 2016, 10, 77. [Google Scholar] [CrossRef]
- Klempin, F.; Kronenberg, G.; Cheung, G.; Kettenmann, H.; Kempermann, G. Properties of doublecortin-(DCX)-expressing cells in the piriform cortex compared to the neurogenic dentate gyrus of adult mice. PLoS ONE 2011, 6, e25760. [Google Scholar] [CrossRef]
- Zhao, M.; Momma, S.; Delfani, K.; Carlen, M.; Cassidy, R.M.; Johansson, C.B.; Brismar, H.; Shupliakov, O.; Frisen, J.; Janson, A.M. Evidence for neurogenesis in the adult mammalian substantia nigra. Proc Natl. Acad. Sci. USA 2003, 100, 7925–7930. [Google Scholar] [CrossRef]
- Jhaveri, D.J.; Tedoldi, A.; Hunt, S.; Sullivan, R.; Watts, N.R.; Power, J.M.; Bartlett, P.F.; Sah, P. Evidence for newly generated interneurons in the basolateral amygdala of adult mice. Mol. Psychiatry 2018, 23, 521–532. [Google Scholar] [CrossRef]
- Magavi, S.S.; Leavitt, B.R.; Macklis, J.D. Induction of neurogenesis in the neocortex of adult mice. Nature 2000, 405, 951–955. [Google Scholar] [CrossRef] [PubMed]
- Batailler, M.; Droguerre, M.; Baroncini, M.; Fontaine, C.; Prevot, V.; Migaud, M. DCX-expressing cells in the vicinity of the hypothalamic neurogenic niche: A comparative study between mouse, sheep, and human tissues. J. Comp. Neurol. 2014, 522, 1966–1985. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Bittman, E.L. Olfactory bulb cells generated in adult male golden hamsters are specifically activated by exposure to estrous females. Horm. Behav. 2002, 4, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohr, M.A.; Sisk, C.L. Pubertally born neurons and glia are functionally integrated into limbic and hypothalamic circuits of the male Syrian hamster. Proc. Natl. Acad. Sci. USA 2013, 110, 4792–4797. [Google Scholar] [CrossRef]
- Fowler, C.D.; Liu, Y.; Ouimet, C.; Wang, Z. The effects of social environment on adult neurogenesis in the female prairie vole. J. Neurobiol. 2002, 51, 115–128. [Google Scholar] [CrossRef]
- Fowler, C.D.; Johnson, F.; Wang, Z. Estrogen regulation of cell proliferation and distribution of estrogen receptor-alpha in the brains of adult female prairie and meadow voles. J. Comp. Neurol. 2005, 489, 166–179. [Google Scholar] [CrossRef]
- Castro, A.E.; Young, L.J.; Camacho, F.J.; Paredes, R.G.; Diaz, N.F.; Portillo, W. Effects of Mating and Social Exposure on Cell Proliferation in the Adult Male Prairie Vole (Microtus ochrogaster). Neural. Plast. 2020, 2020, 8869669. [Google Scholar] [CrossRef]
- Jara, N.; Cifuentes, M.; Martínez, F.; Salazar, K.; Nualart, F. Cytoarchitecture, Proliferative Activity and Neuroblast Migration in the Subventricular Zone and Lateral Ventricle Extension of the Adult Guinea Pig Brain. Stem Cells 2016, 34, 2574–2586. [Google Scholar] [CrossRef]
- Altman, J.; Das, G.D. Postnatal neurogenesis in the guinea-pig. Nature 1967, 214, 1098–1101. [Google Scholar] [CrossRef]
- He, X.; Zhang, X.M.; Wu, J.; Fu, J.; Mou, L.; Lu, D.H.; Cai, Y.; Luo, X.G.; Pan, A.; Yan, X.X. Olfactory experience modulates immature neuron development in postnatal and adult guinea pig piriform cortex. Neuroscience 2014, 259, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Amrein, I.; Becker, A.S.; Engler, S.; Huang, S.H.; Müller, J.; Slomianka, L.; Oosthuizen, M.K. Adult neurogenesis and its anatomical context in the hippocampus of three mole-rat species. Front. Neuroanat. 2014, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Peragine, D.E.; Simpson, J.A.; Mooney, S.J.; Lovern, M.B.; Holmes, M.M. Social regulation of adult neurogenesis in a eusocial mammal. Neuroscience 2014, 268, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Oosthuizen, M.K. Exploratory behaviour, memory and neurogenesis in the social Damaraland mole-rat (Fukomys damarensis). J. Exp. Biol. 2020, 223, jeb221093. [Google Scholar] [CrossRef]
- Oosthuizen, M.K.; Amrein, I. Trading new neurons for status: Adult hippocampal neurogenesis in eusocial Damaraland mole-rats. Neuroscience 2016, 324, 227–237. [Google Scholar] [CrossRef]
- Luzzati, F.; De Marchis, S.; Fasolo, A.; Peretto, P. Neurogenesis in the caudate nucleus of the adult rabbit. J. Neurosci. 2006, 26, 609–621. [Google Scholar] [CrossRef]
- Siwak-Tapp, C.T.; Head, E.; Muggenburg, B.A.; Milgram, N.W.; Cotman, C.W. Neurogenesis decreases with age in the canine hippocampus and correlates with cognitive function. Neurobiol. Learn. Mem. 2007, 88, 249–259. [Google Scholar] [CrossRef]
- Kamiya, S.; Sawada, K. Immunohistochemical characterization of postnatal changes in cerebellar cortical cytoarchitectures in ferrets. Anat. Rec. 2021, 304, 413–424. [Google Scholar] [CrossRef]
- Chawana, R.; Patzke, N.; Alagaili, A.N.; Bennett, N.C.; Mohammed, O.B.; Kaswera-Kyamakya, C.; Gilissen, E.; Ihunwo, A.O.; Pettigrew, J.D.; Manger, P.R. The Distribution of Ki-67 and Doublecortin Immunopositive Cells in the Brains of Three Microchiropteran Species, Hipposideros fuliginosus, Triaenops persicus, and Asellia tridens. Anat. Rec. 2016, 299, 1548–1560. [Google Scholar] [CrossRef]
- Chawana, R.; Patzke, N.; Kaswera, C.; Gilissen, E.; Ihunwo, A.O.; Manger, P.R. Adult neurogenesis in eight Megachiropteran species. Neuroscience 2013, 244, 159–172. [Google Scholar] [CrossRef]
- Akter, M.; Kaneko, N.; Herranz-Pérez, V.; Nakamura, S.; Oishi, H.; García-Verdugo, J.M.; Sawamoto, K. Dynamic Changes in the Neurogenic Potential in the Ventricular-Subventricular Zone of Common Marmoset during Postnatal Brain Development. Cereb. Cortex 2020, 30, 4092–4109. [Google Scholar] [CrossRef] [PubMed]
- Leuner, B.; Kozorovitskiy, Y.; Gross, C.G.; Gould, E. Diminished adult neurogenesis in the marmoset brain precedes old age. Proc. Natl. Acad. Sci. USA 2007, 104, 17169–17173. [Google Scholar] [CrossRef] [PubMed]
- Marlatt, M.W.; Philippens, I.; Manders, E.; Czéh, B.; Joels, M.; Krugers, H.; Lucassen, P.J. Distinct structural plasticity in the hippocampus and amygdala of the middle-aged common marmoset (Callithrix jacchus). Exp. Neurol. 2011, 230, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Bédard, A.; Lévesque, M.; Bernier, P.J.; Parent, A. The rostral migratory stream in adult squirrel monkeys: Contribution of new neurons to the olfactory tubercle and involvement of the antiapoptotic protein Bcl-2. Eur. J. Neurosci. 2002, 16, 1917–1924. [Google Scholar] [CrossRef] [PubMed]
- Lyons, D.M.; Buckmaster, P.S.; Lee, A.G.; Wu, C.; Mitra, R.; Duffey, L.M.; Buckmaster, C.L.; Her, S.; Patel, P.D.; Schatzberget, A.F. Stress coping stimulates hippocampal neurogenesis in adult monkeys. Proc. Natl. Acad. Sci. USA 2010, 107, 14823–14827. [Google Scholar] [CrossRef]
- Bernier, P.J.; Bedard, A.; Vinet, J.; Levesque, M.; Parent, A. Newly generated neurons in the amygdala and adjoining cortex of adult primates. Proc. Natl. Acad. Sci. USA 2002, 99, 11464–11469. [Google Scholar] [CrossRef]
- Zhang, X.M.; Cai, Y.; Chu, Y.; Chen, E.Y.; Feng, J.C.; Luo, X.G.; Xiong, K.; Struble, R.G.; Clough, R.W.; Patrylo, P.R.; et al. Doublecortin-expressing cells persist in the associative cerebral cortex and amygdala in aged nonhuman primates. Front. Neuroanat. 2009, 3, 17. [Google Scholar] [CrossRef]
- Gould, E.; Reeves, A.J.; Graziano, M.S.; Gross, C.G. Neurogenesis in the neocortex of adult primates. Science 1999, 286, 548–552. [Google Scholar] [CrossRef]
- Sorrells, S.F.; Paredes, M.F.; Cebrian-Silla, A.; Sandoval, K.; Qi, D.; Kelley, K.W.; James, D.; Mayer, S.; Chang, J.; Auguste, K.I.; et al. Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults. Nature 2018, 555, 377–381. [Google Scholar] [CrossRef]
- Sorrells, S.F.; Paredes, M.F.; Zhang, Z.; Kang, G.; Pastor-Alonso, O.; Biagiotti, S.; Page, C.E.; Sandoval, K.; Knox, A.; Connolly, A.; et al. Positive Controls in Adults and Children Support That Very Few, If Any, New Neurons Are Born in the Adult Human Hippocampus. J. Neurosci. 2021, 41, 2554–2565. [Google Scholar] [CrossRef]
- Bédard, A.; Parent, A. Evidence of newly generated neurons in the human olfactory bulb. Brain Res. Dev. Brain Res. 2004, 151, 159–168. [Google Scholar] [CrossRef]
- Spalding, K.L.; Bergmann, O.; Alkass, K.; Bernard, S.; Salehpour, M.; Huttner, H.B.; Boström, E.; Westerlund, I.; Vial, C.; Buchholz, B.A.; et al. Dynamics of hippocampal neurogenesis in adult humans. Cell 2013, 153, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Boldrini, M.; Fulmore, C.A.; Tartt, A.N.; Simeon, L.R.; Pavlova, I.; Poposka, V.; Rosoklija, G.B.; Stankov, A.; Arango, V.; Dwork, A.J.; et al. Human Hippocampal Neurogenesis Persists throughout Aging. Cell Stem Cell 2018, 22, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Jiménez, E.P.; Terreros-Roncal, J.; Flor-García, M.; Rábano, A.; Llorens-Martín, M. Evidences for Adult Hippocampal Neurogenesis in Humans. J. Neurosci. 2021, 41, 2541–2553. [Google Scholar] [CrossRef] [PubMed]
- Ernst, A.; Alkass, K.; Bernard, S.; Salehpour, M.; Perl, S.; Tisdale, J.; Possnert, G.; Druid, H.; Frisén, J. Neurogenesis in the striatum of the adult human brain. Cell. 2014, 156, 1072–1083. [Google Scholar] [CrossRef]
- Sorrells, S.; Paredes, M.F.; Velmeshev, D.; Herranz-Pérez, V.; Sandoval, K.; Mayer, S.; Chang, E.F.; Insausti, R.; Kriegstein, A.R.; Rubenstein, J.L.; et al. Immature excitatory neurons develop during adolescence in the human amygdala. Nat. Commun. 2019, 10, 2748. [Google Scholar] [CrossRef]
- Ache, B.W.; Young, J.M. Olfaction: Diverse species, conserved principles. Neuron 2005, 48, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Luskin, M.B. Restricted proliferation and migration of postnatally generated neurons derived from the forebrain subventricular zone. Neuron 1993, 11, 173–189. [Google Scholar] [CrossRef]
- Carleton, A.; Petreanu, L.T.; Lansford, R.; Alvarez-Buylla, A.; Lledo, P.M. Becoming a new neuron in the adult olfactory bulb. Nat. Neurosci. 2003, 6, 507–518. [Google Scholar] [CrossRef]
- Rochefort, C.; Gheusi, G.; Vincent, J.D.; Lledo, P.M. Enriched odor exposure increases the number of newborn neurons in the adult olfactory bulb and improves odor memory. J. Neurosci. 2002, 22, 2679–2689. [Google Scholar] [CrossRef]
- Sakamoto, M.; Imayoshi, I.; Ohtsuka, T.; Yamaguchi, M.; Mori, K.; Kageyama, R. Continuous neurogenesis in the adult forebrain is required for innate olfactory responses. Proc. Natl. Acad. Sci. USA 2011, 108, 8479–8484. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Ogawa, Y.; Yoshihara, S. A Subtype of Olfactory Bulb Interneurons Is Required for Odor Detection and Discrimination Behaviors. J. Neurosci. 2016, 36, 8210–8227. [Google Scholar] [CrossRef] [PubMed]
- Grelat, A.; Benoit, L.; Wagner, S.; Moigneu, C.; Lledo, P.M.; Alonso, M. Adult-born neurons boost odor-reward association. Proc. Natl. Acad. Sci. USA 2018, 115, 2514–2519. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Yoshihara, S.; Tsuboi, A. The Functional Role of Olfactory Bulb Granule Cell Subtypes Derived From Embryonic and Postnatal Neurogenesis. Front. Mol. Neurosci. 2018, 11, 229. [Google Scholar] [CrossRef] [Green Version]
- Li, W.L.; Chu, M.W.; Wu, A.; Suzuki, Y.; Imayoshi, I.; Komiyama, T. Adult-born neurons facilitate olfactory bulb pattern separation during task engagement. Elife 2018, 7, e33006. [Google Scholar] [CrossRef]
- Shani-Narkiss, H.; Vinograd, A.; Landau, I.D.; Tasaka, G.; Yayon, N.; Terletsky, S.; Groysman, M.; Maor, I.; Sompolinsky, H.; Mizrahi, A. Young adult-born neurons improve odor coding by mitral cells. Nat. Commun. 2020, 11, 5867. [Google Scholar] [CrossRef]
- Sakamoto, M.; Kageyama, R.; Imayoshi, I. The functional significance of newly born neurons integrated into olfactory bulb circuits. Front. Neurosci. 2014, 8, 121. [Google Scholar] [CrossRef]
- Forest, J.; Chalençon, L.; Midroit, M.; Terrier, C.; Caillé, I.; Sacquet, J.; Benetollo, C.; Martin, K.; Richard, M.; Didier, A.; et al. Role of Adult-Born Versus Preexisting Neurons Born at P0 in Olfactory Perception in a Complex Olfactory Environment in Mice. Cereb. Cortex 2020, 30, 534–549. [Google Scholar] [CrossRef]
- Quiñones-Hinojosa, A.; Sanai, N.; Soriano-Navarro, M.; Gonzalez-Perez, O.; Mirzadeh, Z.; Gil-Perotin, S.; Romero-Rodriguez, R.; Berger, M.S.; García-Verdugo, J.M.; Alvarez-Buylla, A. Cellular composition and cytoarchitecture of the adult human subventricular zone: A niche of neural stem cells. J. Comp. Neurol. 2006, 494, 415–434. [Google Scholar] [CrossRef]
- Weickert, C.S.; Webster, M.J.; Colvin, S.M.; Herman, M.M.; Hyde, T.M.; Weinberger, D.R.; Kleinman, J.E. Localization of epidermal growth factor receptors and putative neuroblasts in human subependymal zone. J. Comp. Neurol. 2000, 423, 359–372. [Google Scholar] [CrossRef]
- Curtis, M.A.; Faull, R.L.; Eriksson, P.S. The effect of neurodegenerative diseases on the subventricular zone. Nat. Rev. Neurosci. 2007, 8, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Sanai, N.; Nguyen, T.; Ihrie, R.A. Corridors of migrating neurons in the human brain and their decline during infancy. Nature 2011, 478, 382–386. [Google Scholar] [CrossRef]
- Santopolo, G.; Magnusson, J.P.; Lindvall, O.; Kokaia, Z.; Frisén, J. Blocking Notch-Signaling Increases Neurogenesis in the Striatum after Stroke. Cells 2020, 9, 1732. [Google Scholar] [CrossRef] [PubMed]
- Tepper, B.; Koguc-Sobolewska, P.; Jaslan, K.; Turlejski, K.; Bartkowska, K.; Djavadian, R. Impaired olfactory neurogenesis affects the performance of olfactory-guided behavior in aged female opossums. Sci. Rep. 2021, 11, 4418. [Google Scholar] [CrossRef]
- Cádiz-Moretti, B.; Abellán-Álvaro, M.; Pardo-Bellver, C.; Martínez-García, F.; Lanuza, E. Afferent and Efferent Connections of the Cortex-Amygdala Transition Zone in Mice. Front. Neuroanat. 2016, 10, 125. [Google Scholar] [CrossRef] [PubMed]
- Nacher, J.; Crespo, C.; McEwen, B.S. Doublecortin expression in the adult rat telencephalon. Eur. J. Neurosci. 2001, 14, 629–644. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xie, M.-X.; Li, J.-M.; Hu, X.; Patrylo, P.R.; Luo, X.-G.; Cai, Y.; Li, Z.; Yan, X.-X. Prenatal genesis of layer II doublecortin expressing neurons in neonatal and young adult guinea pig cerebral cortex. Front. Neuroanat. 2015, 9, 109. [Google Scholar] [CrossRef] [PubMed]
- Cameron, H.A.; McKay, R.D. Adult neurogenesis produces a large pool of new granule cells in the dentate gyrus. J. Comp. Neurol. 2001, 435, 406–417. [Google Scholar] [CrossRef]
- Snyder, J.S.; Kee, N.; Wojtowicz, J.M. Effects of adult neurogenesis on synaptic plasticity in the rat dentate gyrus. J. Neurophysiol. 2001, 85, 2423–2431. [Google Scholar] [CrossRef]
- van Praag, H.; Schinder, A.F.; Christie, B.R.; Toni, N.; Palmer, T.D.; Gage, F.H. Functional neurogenesis in the adult hippocampus. Nature 2002, 415, 1030–1034. [Google Scholar] [CrossRef]
- Toni, N.; Laplagne, D.A.; Zhao, C.; Lombardi, G.; Ribak, C.E.; Gage, F.H.; Schinder, A.F. Neurons born in the adult dentate gyrus form functional synapses with target cells. Nat. Neurosci. 2008, 11, 901–907. [Google Scholar] [CrossRef]
- Knoth, R.; Singec, I.; Ditter, M.; Pantazis, G.; Capetian, P.; Meyer, R.P.; Horvat, V.; Volk, B.; Kempermann, G. Murine features of neurogenesis in the human hippocampus across the lifespan from 0 to 100 years. PLoS ONE 2010, 5, e8809. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, H.G.; Toda, T.; Gage, F.H. Adult Hippocampal Neurogenesis: A Coming-of-Age Story. J. Neurosci. 2018, 38, 10401–10410. [Google Scholar] [CrossRef]
- Kempermann, G.; Gage, F.H.; Aigner, L.; Song, H.; Curtis, M.A.; Thuret, S.; Kuhn, H.G.; Jessberger, S.; Frankland, P.W.; Cameron, H.A.; et al. Human Adult Neurogenesis: Evidence and Remaining Questions. Cell Stem Cell 2018, 23, 25–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paredes, M.F.; Sorrells, S.F.; Cebrian-Silla, A.; Sandoval, K.; Qi, D.; Kelley, K.W.; James, D.; Mayer, S.; Chang, J.; Auguste, K.I.; et al. Does Adult Neurogenesis Persist in the Human Hippocampus? Cell Stem Cell 2018, 23, 780–781. [Google Scholar] [CrossRef]
- Alam, M.J.; Kitamura, T.; Saitoh, Y.; Ohkawa, N.; Kondo, T.; Inokuchi, K. Adult Neurogenesis Conserves Hippocampal Memory Capacity. J. Neurosci. 2018, 38, 6854–6863. [Google Scholar] [CrossRef] [PubMed]
- Epp, J.R.; Chow, C.; Galea, L.A. Hippocampus-dependent learning influences hippocampal neurogenesis. Front. Neurosci. 2013, 7, 57. [Google Scholar] [CrossRef]
- Suárez-Pereira, I.; Carrión, Á.M. Updating stored memory requires adult hippocampal neurogenesis. Sci. Rep. 2015, 5, 13993. [Google Scholar] [CrossRef]
- Abrous, D.N.; Wojtowicz, J.M. Interaction between Neurogenesis and Hippocampal Memory System: New Vistas. Cold Spring Harb. Perspect. Biol. 2015, 7, a018952. [Google Scholar] [CrossRef]
- Tashiro, A.; Makino, H.; Gage, F.H. Experience-specific functional modification of the dentate gyrus through adult neurogenesis: A critical period during an immature stage. J. Neurosci. 2007, 27, 3252–3259. [Google Scholar] [CrossRef]
- Nilsson, M.; Perfilieva, E.; Johansson, U.; Orwar, O.; Eriksson, P.S. Enriched environment increases neurogenesis in the adult rat dentate gyrus and improves spatial memory. J. Neurobiol. 1999, 39, 569–578. [Google Scholar] [CrossRef]
- Dupret, D.; Fabre, A.; Dobrossy, M.; Panatier, A.; Rodríguez, J.J.; Lamarque, S.; Lemaire, V.; Oliet, S.H.R.; Piazza, P.-V.; Abrous, D.N. Spatial learning depends on both the addition and removal of new hippocampal neurons. PLoS Biol. 2007, 5, e214. [Google Scholar] [CrossRef] [PubMed]
- Tronel, S.; Fabre, A.; Charrier, V.; Oliet, S.H.; Gage, F.H.; Abrous, D.N. Spatial learning sculpts the dendritic arbor of adult-born hippocampal neurons. Proc. Natl. Acad. Sci. USA 2010, 107, 7963–7968. [Google Scholar] [CrossRef] [PubMed]
- Snyder, J.S.; Hong, N.S.; McDonald, R.J.; Wojtowicz, J.M. A role for adult neurogenesis in spatial long-term memory. Neuroscience 2005, 130, 843–852. [Google Scholar] [CrossRef]
- Leuner, B.; Gould, E.; Shors, T.J. Is there a link between adult neurogenesis and learning? Hippocampus 2006, 16, 216–224. [Google Scholar] [CrossRef]
- Groves, J.O.; Leslie, I.; Huang, G.-J.; McHugh, S.B.; E Taylor, A.; Mott, R.; Munafo, M.; Bannerman, D.M.; Flint, J. Ablating adult neurogenesis in the rat has no effect on spatial processing: Evidence from a novel pharmacogenetic model. PLoS Genet. 2013, 9, e1003718. [Google Scholar] [CrossRef]
- Tepper, B.; Aniszewska, A.; Bartkowska, K.; Grochocka, L.; Turlejski, K.; Djavadian, R. Aged Opossums Show Alterations in Spatial Learning Behavior and Reduced Neurogenesis in the Dentate Gyrus. Front. Neurosci. 2019, 13, 1210. [Google Scholar] [CrossRef] [Green Version]



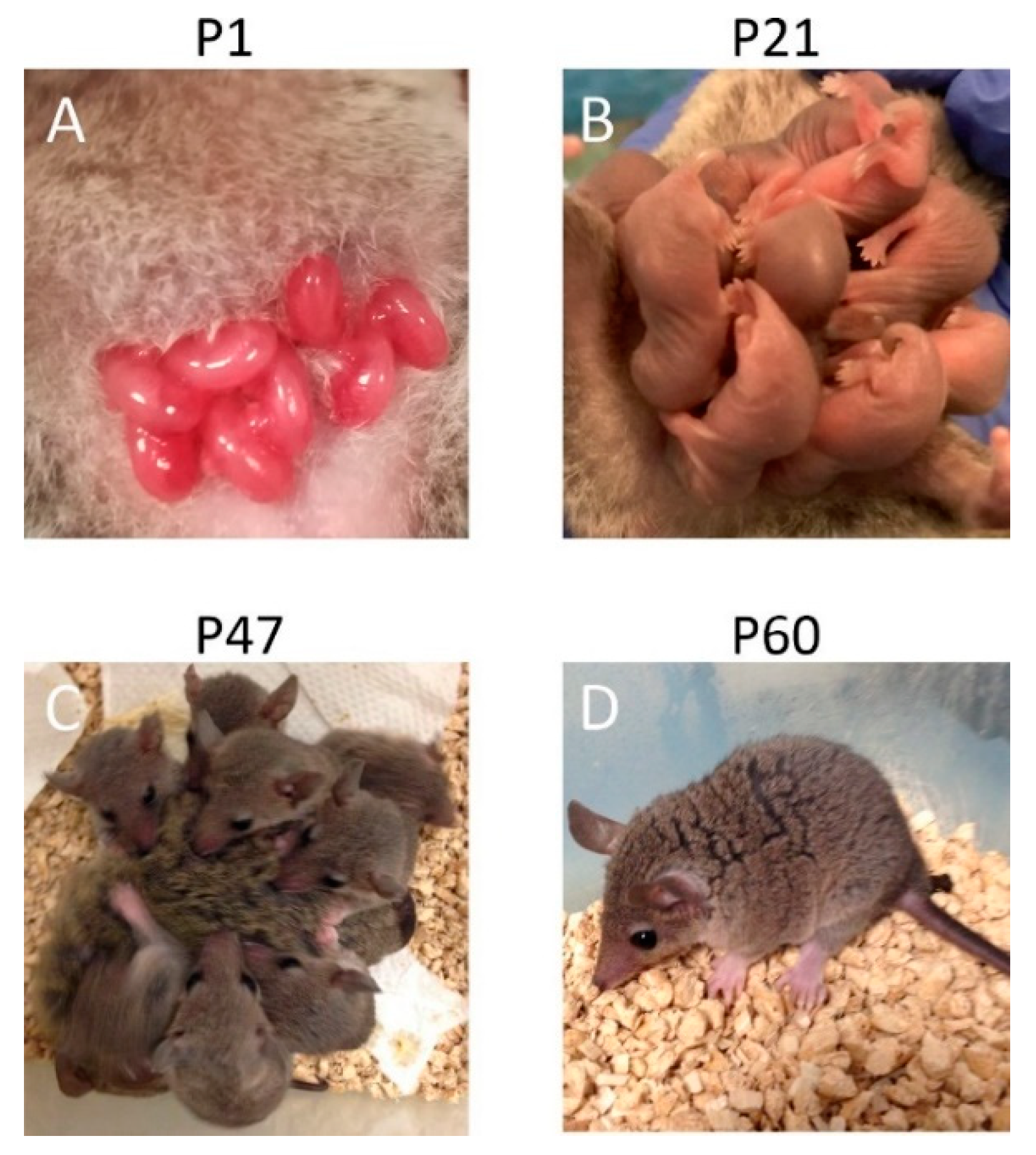{kind=link}
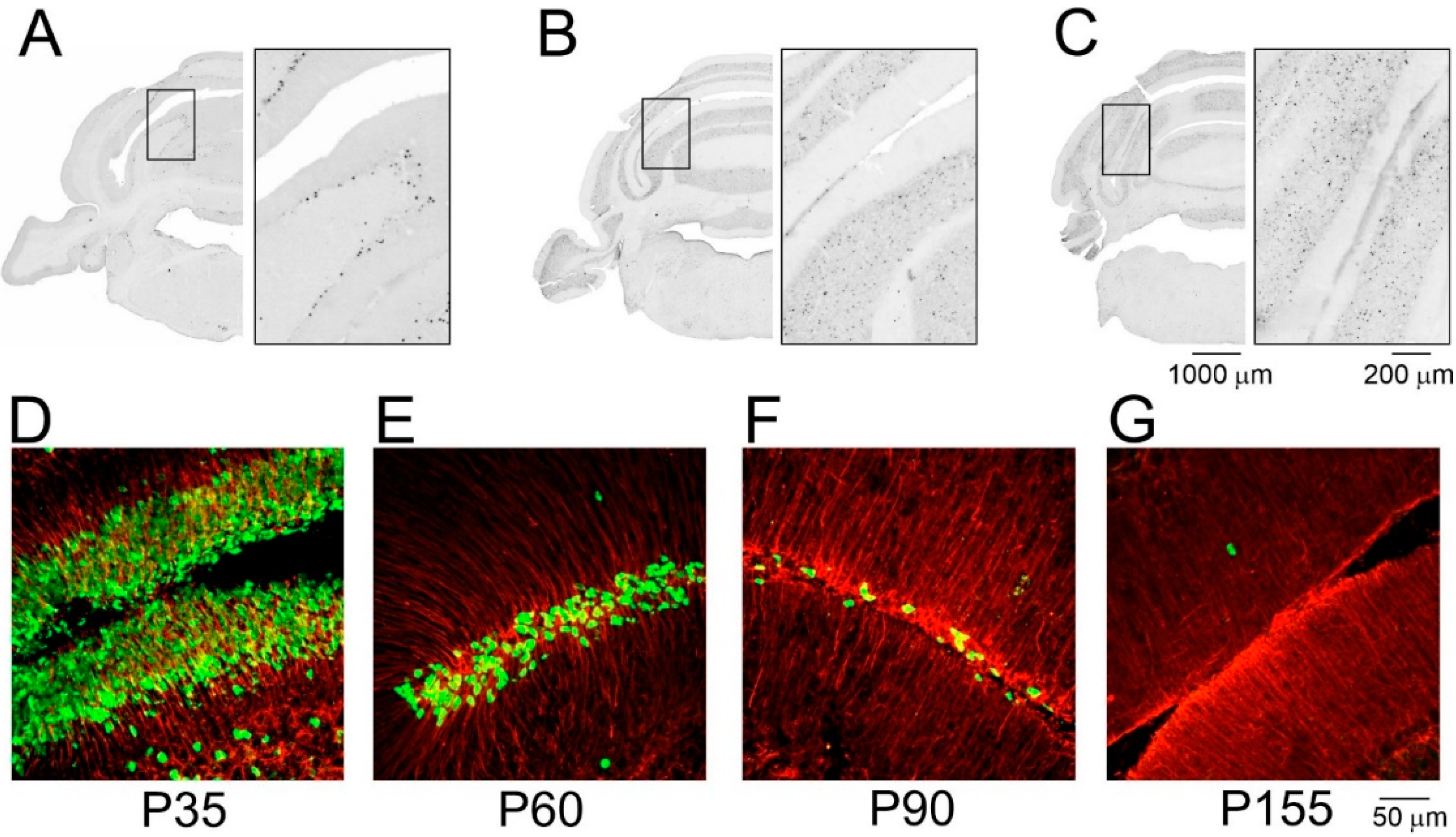{kind=link}
| Order/Family | Species | Gestation | Eyes Opening | Lifespan | Postnatal Developmental Neurogenesis | Adult Neurogenesis | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OB | DG | CER | OB/SVZ | DG | CER | PIR | Other | |||||
| Didelphimorphia/Didelphidae | Gray short-tailed opossum | 14–15 days | P35–P37 | 2.5 yrs | up to P28 [2,28] | P1-P155 [29] | 5 mth-2 yrs [65] | 6.5–21.5 mth [65] | up to P270 | |||
| Dasyuromorphia/Dasyuridae | Fat-tailed dunnart | 13–16 days | P45 | 1–2 yrs | 4–24 mth [66] | |||||||
| Didrotodontia/Phalangeridae | Brush-tailed possum | 18 days | P110 | 13 yrs | P5–P82 [3] | up to 3 mth [4] | ||||||
| Diprotodontia/Macropodidae | Tammar wallaby | 25–28 days | P140 | 11–14 yrs | up to P25 [54] | |||||||
| Quokka wallaby | 28 days | P110 | 8–15 yrs | P20-P85 [50] | ||||||||
| Euliptyphla/Erinaceidae | White-breasted hedgehog | 35 days | P21 | 3–5 yrs | AD [80] | AD [80] | AD [80] | |||||
| Eulipotyhla/Talpidae | European mole | 30 days | P22 | 2–3 yrs | AD [80] | AD [80] | AD [80] | |||||
| Eulipotphla/Soricidae | Hottentot golden mole | AD [79] | AD [79] | AD [79] | CTX [79] | |||||||
| Giant other shrew | 22 days | P20–P24 | 5 yrs | AD [76] | AD [76] | AD [76] | OT, EPN [76] | |||||
| Pygmy shrew, Common shrew | 22–24 days | P20–P24 | 1 yr | 1 yr [74] | 5 mth [74] | |||||||
| Greater white-toothed shrew, Eurasian water shrew, African giant shrew, Asian house shrew | 20–31 days | P20–P24 | 1.5–3 yrs | AD [75] | AD [75] | |||||||
| Rodentia/Muridae | Rats (Long Evans, Sprague-Dawley, Wistar, Fischer 344, Brown Norway wild rats) | 21–23 days | P13–P15 | 2 yrs | P0-P10 [87] | P6-P15 [6,88] up to P14 [89] | up to P21 [63] | AD [90] | 6–21 mth [9,67] 8–9 wk [88] | AD [91] | HTH 2 mth [92,93] CTX, STR 9–10 wk [94] BrSt AD [95] | |
| Mice (C57BL/6, CD1, BALB/c, ICR, A/J, FVB, C3H/HeJ, 129/SvJ, DBA/1, DBA/2) | 19–21 days | P10–P13 | 1–1.5 yrs | P0-P20 [53,96] | up to P20 [48,49,97] | up to P15 [55] | 3–4 mth [98] 2 mth [99] | 3–4 mth [98,100] | AD [101] | SN 2–20 mth [102] BLA 2–4 mth [103] CTX, STR 2–4 mth [98,104] HTH 3–4 mth [105] | ||
| Rodentia/Cricetidae | Syrian hamster | 16 days | P12–P14 | 2–3 yrs | 2.5 mth [106] | P28-P49 [107] | BLA, HTH P28-P49 [107] | |||||
| Meadow vole, Prairie vole | 21 days | P14 | 3–16 mth | 3–5 mth [108,109,110] | 3–5 mth [108,109,110] | CTX, CP, BLA, THT 3–5 mth [108,109] | ||||||
| Rodentia/Caviidae | Guinea pig | 65–68 days | Born with open eyes | 4–5 yrs | up to P30 [68] | 6, 12 mth [111] | 1 yr [68,112] | 12–14 mth [113] | ||||
| Rodentia/Bathyergidae | Highveld mole-rat, Cape mole-rat, Naked mole-rat, Damaraland mole-rat | 70 days | P14 | 6–15 yrs | 1 yr [114] 2–9 yrs [115] AD [116,117] | 2–9 yrs [115] | BLA, 2–9 yrs [115] | |||||
| Rodentia/Leporidae | New Zeland white rabbit | 31 days | P7 | 9 yrs | P10 [6] | 1–3 yrs [6] | CN, AD [118] | |||||
| Hyracoidea/Procaviidae | Rock hyrax | 200 days | Born with open eyes | 8–12 yrs | AD [79] | AD [79] | AD [79] | CTX, AD [79] | ||||
| Macrosce-lidea/Macroscelididae | Eastern rock sengi, Four-toed sengi | 40–60 days | Born with open eyes | 4–6 yrs | AD [79] | AD [79] | AD [79] | CTX, AD [79] | ||||
| Artiodactyla/Bovidae | Ilede-France sheep | 147 days | Born with open eyes | 10–12 yrs | AD [73] | AD [73] | HTH, 18–24 mth [105] | |||||
| Carnivora/Felidae | Domestic cat | 64 days | P7–P10 | 12–18 yrs | 18–24 mth [71] | CTX, EC 18–24 mth [71] | ||||||
| Carnivora/Canidae | Domestic dog | 61 days | P10–P14 | 10–13 yrs | 2–6 yrs [70,119] | |||||||
| Carnivora/Mustelidae | Ferret | 42 days | P32 | 5–12 yrs | P4, P21 [120] | |||||||
| Scandentia/Tupaiidae | Tree shrew | 46 days | P21 | 12 yrs | 2 mth-6 yrs [78] | AD [77] | 2 mth -6 yrs [78] | BLA 2 mth-6 yrs [78] | ||||
| Chiroptera | Microchiroptera bats | 44–180 days | P1–P2 | 20 yrs | AD [121] | AD [81] | AD [121] | AC, BLA [121] | ||||
| Megachiroptera bats | 4–6 m | P1–P2 | 20 yrs | AD [122] | AD [82,122] | AD [122] | AD [122] | BrSt, Tectum, AD [122] | ||||
| Primates/(New World monkey) Callitrichidae | Common marmoset | 151 days | Born with open eyes | 12 yrs | up to P30 [123] | 1.5–7 yrs [124] 4 yrs [123,125] | CTX, CC, AMG 4 yrs [125] | |||||
| Primates/(New World monkey) Cebidae | Squirrel monkey | 160–170 days | Born with open eyes | 21 yrs | 4–6 yrs [126] | 7–10 yrs [127] | 3-6 yrs [128] | CTX, BLA 3-6 yrs [128] | ||||
| Primates/(Old World monkey) Cercopithecidae | Rhesus monkey | 165 days | Born with open eyes | 30 yrs | 12, 21, 31 yrs [129] | CTX, BLA 12, 21, 31 yrs [129]] | ||||||
| Macaque monkey | 162 days | E125 | 25 yrs | 5–23 yrs [84] | 6-12 yrs [128] | CTX AD [130], BLA6-12 yrs [128] | ||||||
| Primates/Hominidae | Humans | 280 days | E195 | 75–80 yrs | 3 wk-1 yr [131,132] up to 5 mth [132] | 16-69 yrs [133] | 23–72 yrs [85] AD [134,135,136] 14–79 yrs [135] | STR 3–79 yrs [137] AMG 24–67 yrs [138] | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartkowska, K.; Tepper, B.; Turlejski, K.; Djavadian, R. Postnatal and Adult Neurogenesis in Mammals, Including Marsupials. Cells 2022, 11, 2735. https://doi.org/10.3390/cells11172735
Bartkowska K, Tepper B, Turlejski K, Djavadian R. Postnatal and Adult Neurogenesis in Mammals, Including Marsupials. Cells. 2022; 11(17):2735. https://doi.org/10.3390/cells11172735
Chicago/Turabian StyleBartkowska, Katarzyna, Beata Tepper, Krzysztof Turlejski, and Ruzanna Djavadian. 2022. "Postnatal and Adult Neurogenesis in Mammals, Including Marsupials" Cells 11, no. 17: 2735. https://doi.org/10.3390/cells11172735
APA StyleBartkowska, K., Tepper, B., Turlejski, K., & Djavadian, R. (2022). Postnatal and Adult Neurogenesis in Mammals, Including Marsupials. Cells, 11(17), 2735. https://doi.org/10.3390/cells11172735