
Actin-Related Protein 4 and Linker Histone Sustain Yeast Replicative Ageing

 ,
,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Yeast Mating Experiments
2.3. Assessment of the Yeast Cell Growth Rate
2.4. Calculation of the Mean Doubling Time
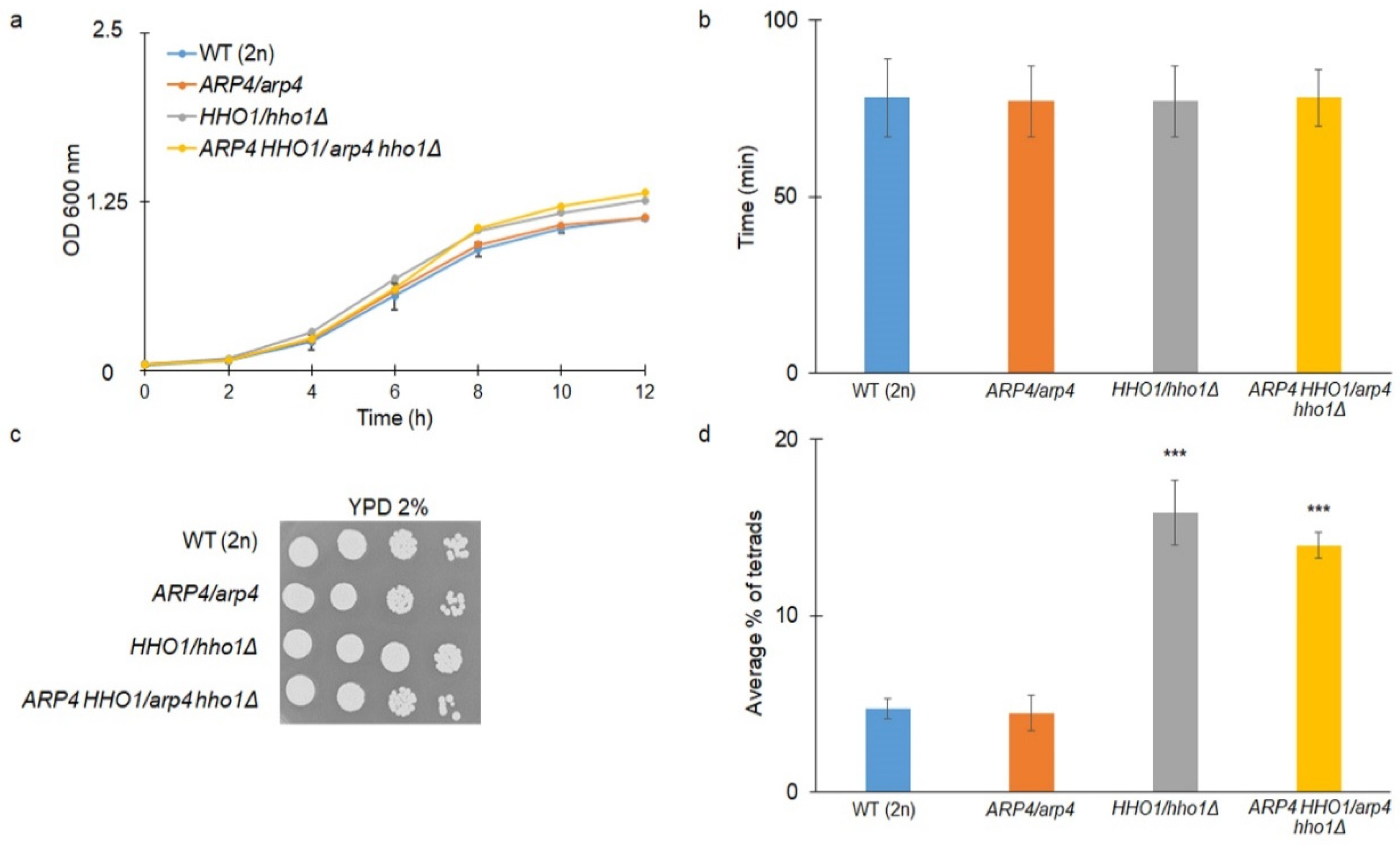
2.5. Sporulation Efficiency Assays
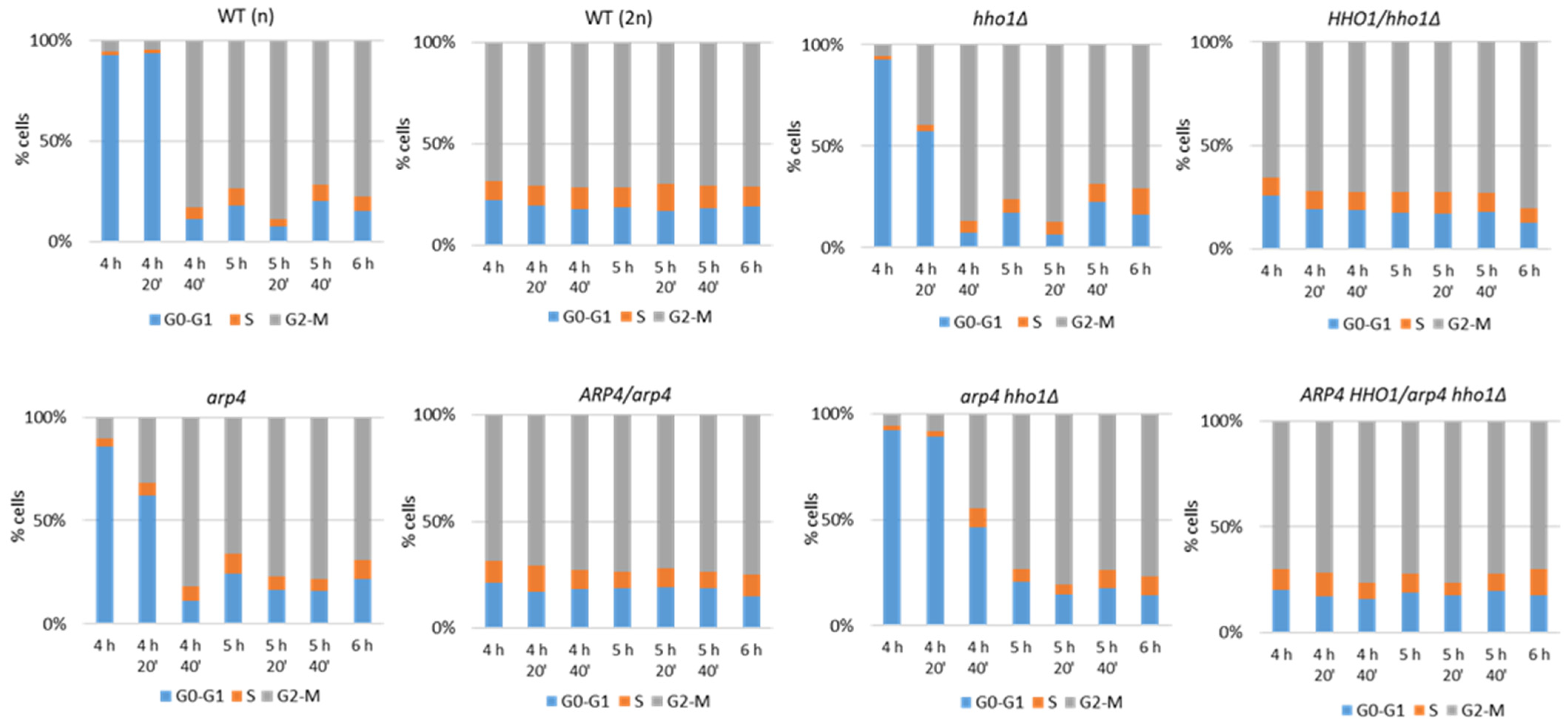
2.6. Fluorescence-Activated Cell Sorting (FACS) for Cell Cycle Analyses
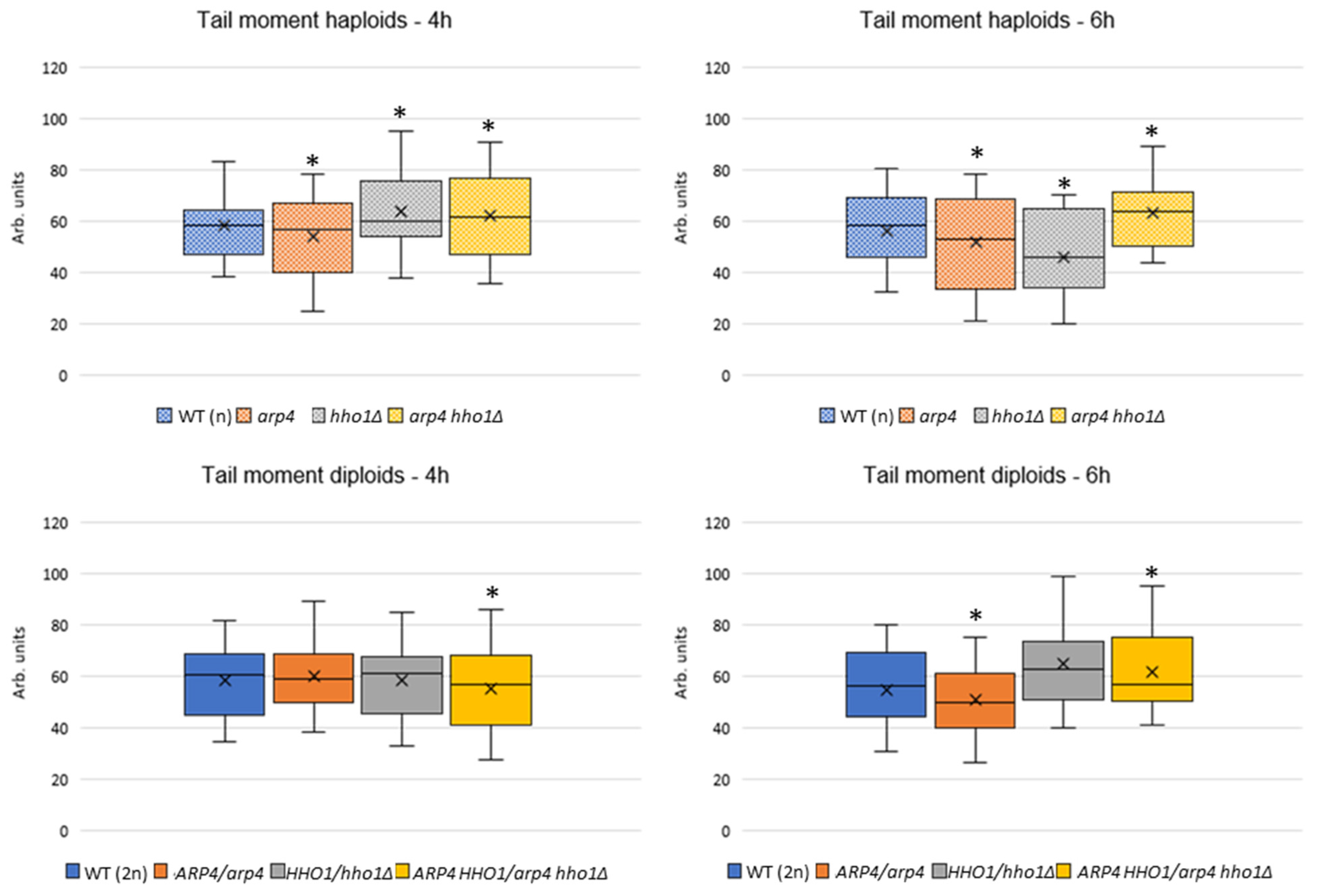
2.7. Chromatin Yeast Comet Assay (ChYCA)
2.8. Determination of the Yeast Budding Lifespan
2.9. Determination of the Yeast Total Lifespan
2.10. Yeast Vacuole Staining
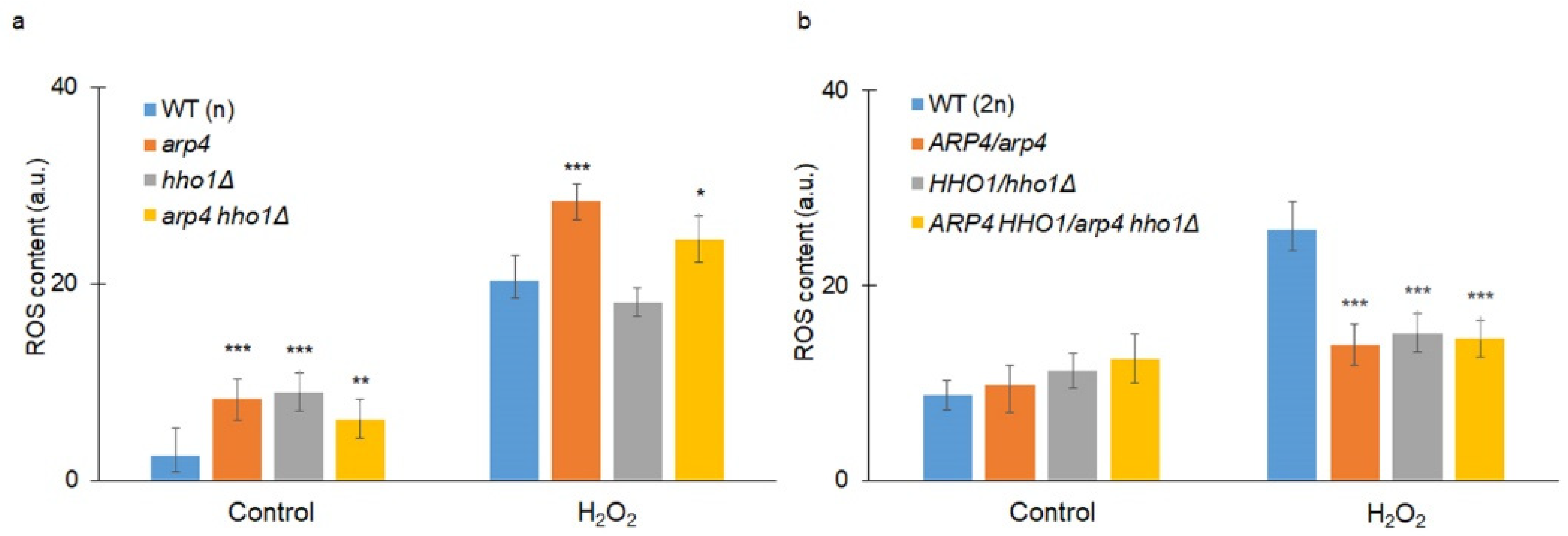
2.11. Measurement of the Amount of Superoxide Anion Generation from the Yeast Cells during Their RLS
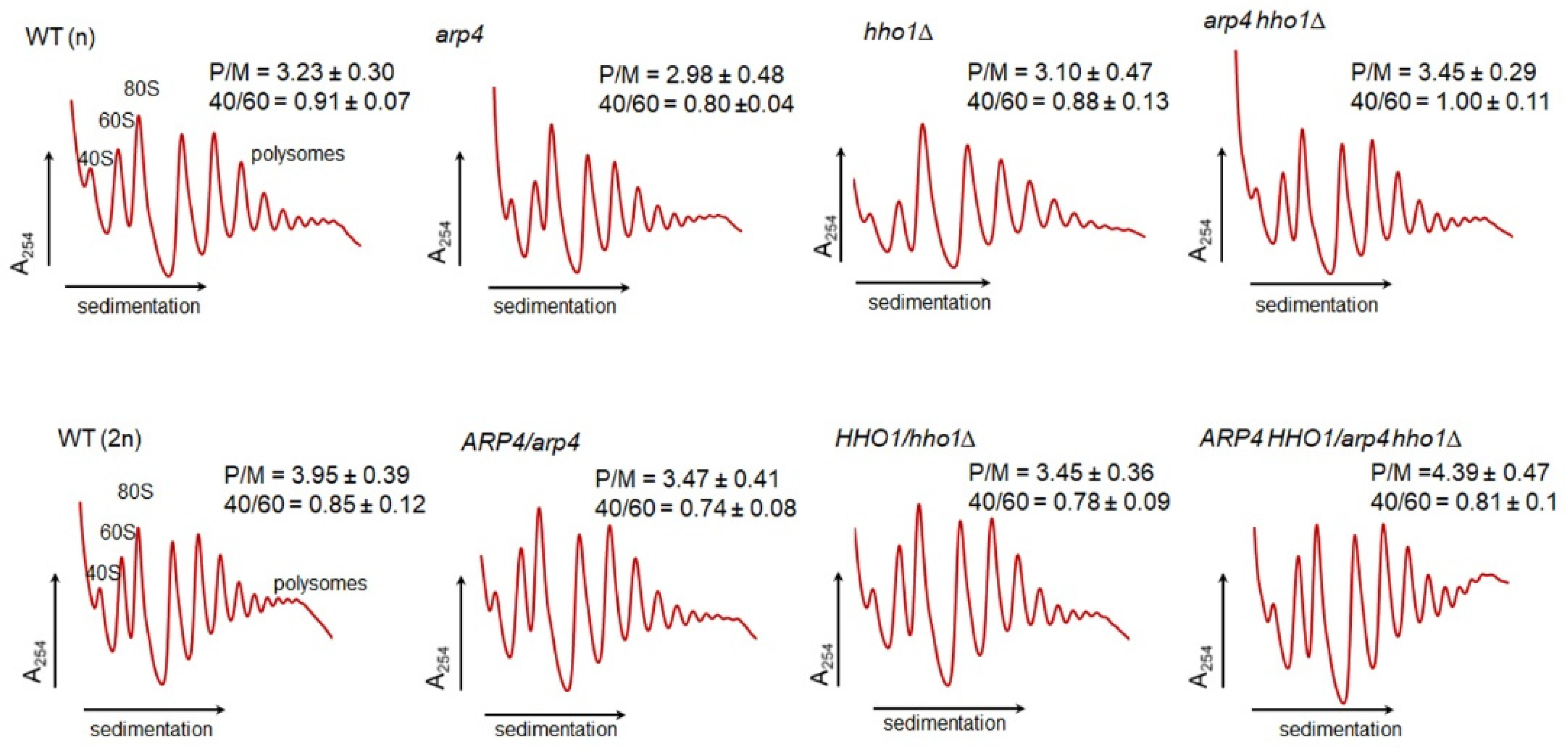
2.12. Polysome Profiles of the Studied Yeast Cells during the RLS
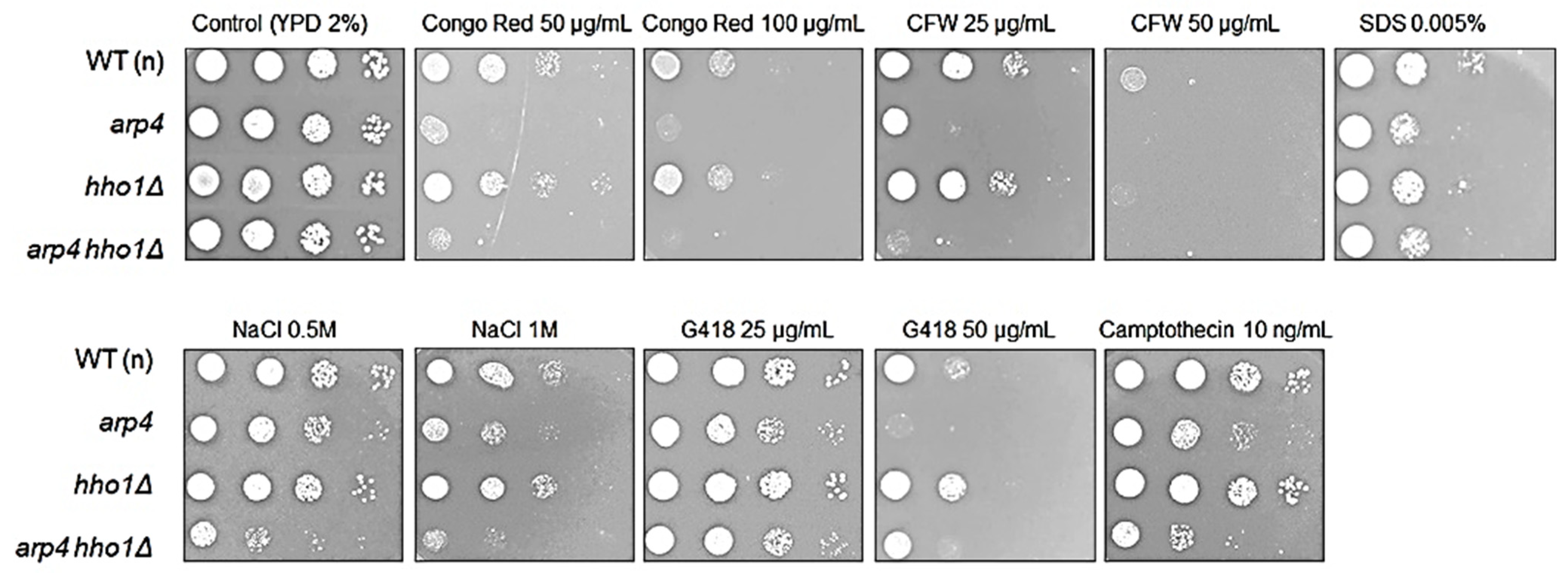
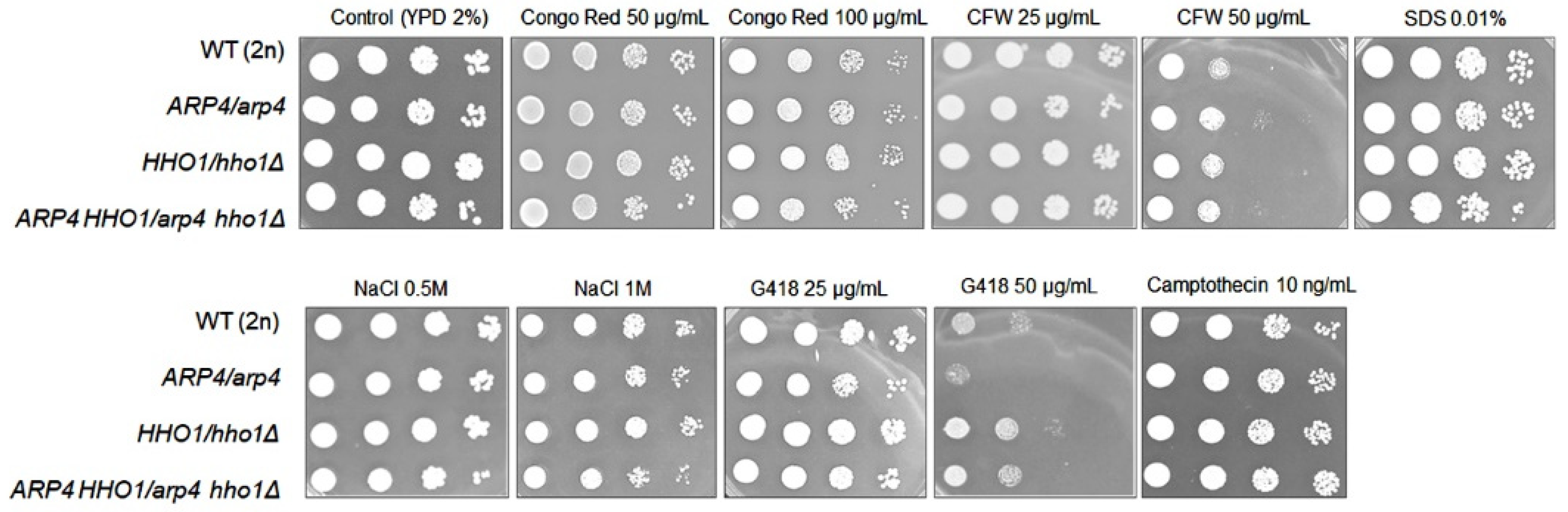
2.13. Phenotypic Analysis—Spot Tests
2.14. Examination of the Yeast Cells’ Chemical Composition by Raman Spectra
2.15. Statistical Analysis
3. Results
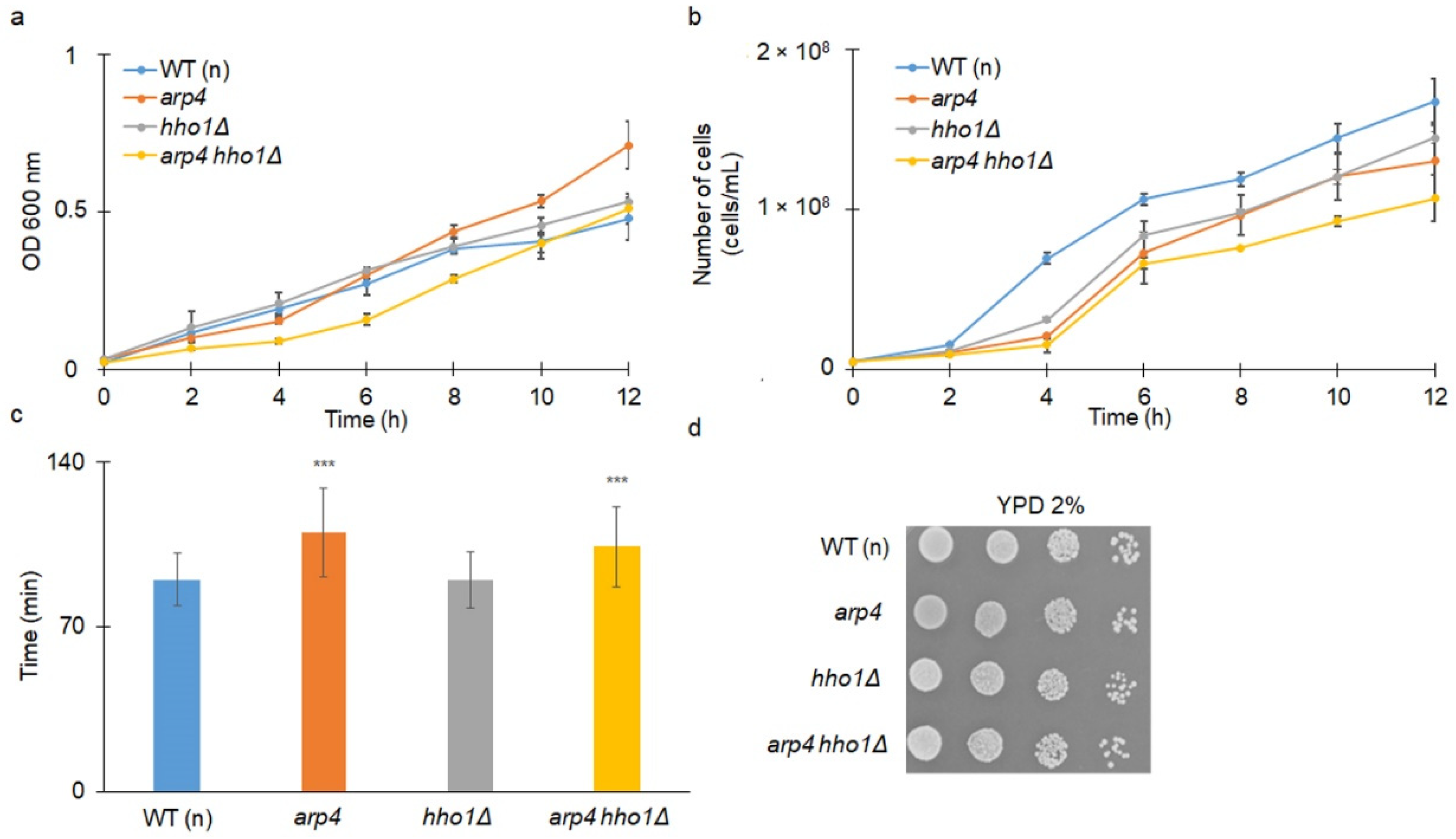
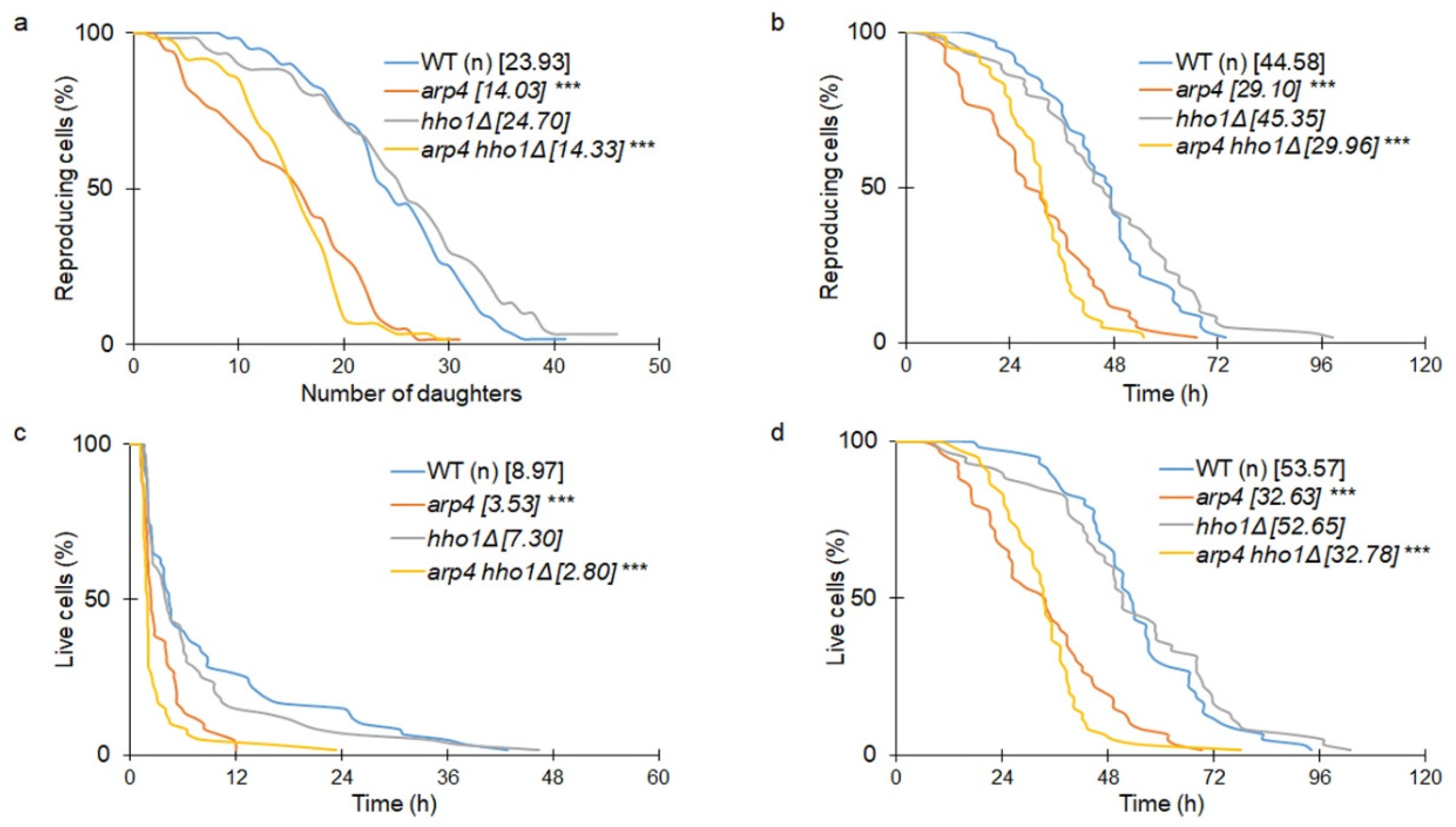
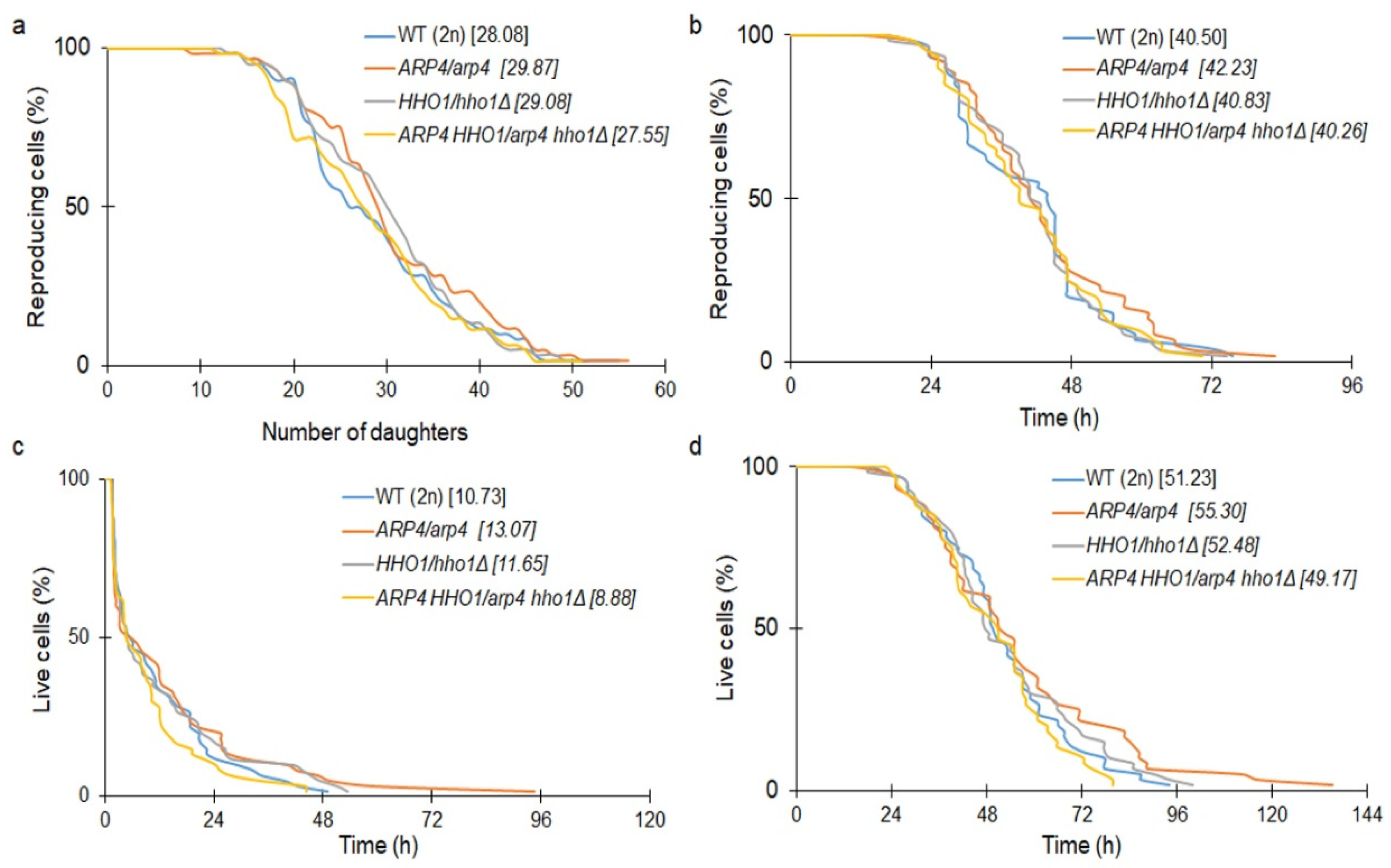
3.1. The Haploid Mutants arp4 and arp4 hho1Δ Exhibit Abnormal Growth during Their RLS Due to Changes in Cell Cycle and Genome Organization
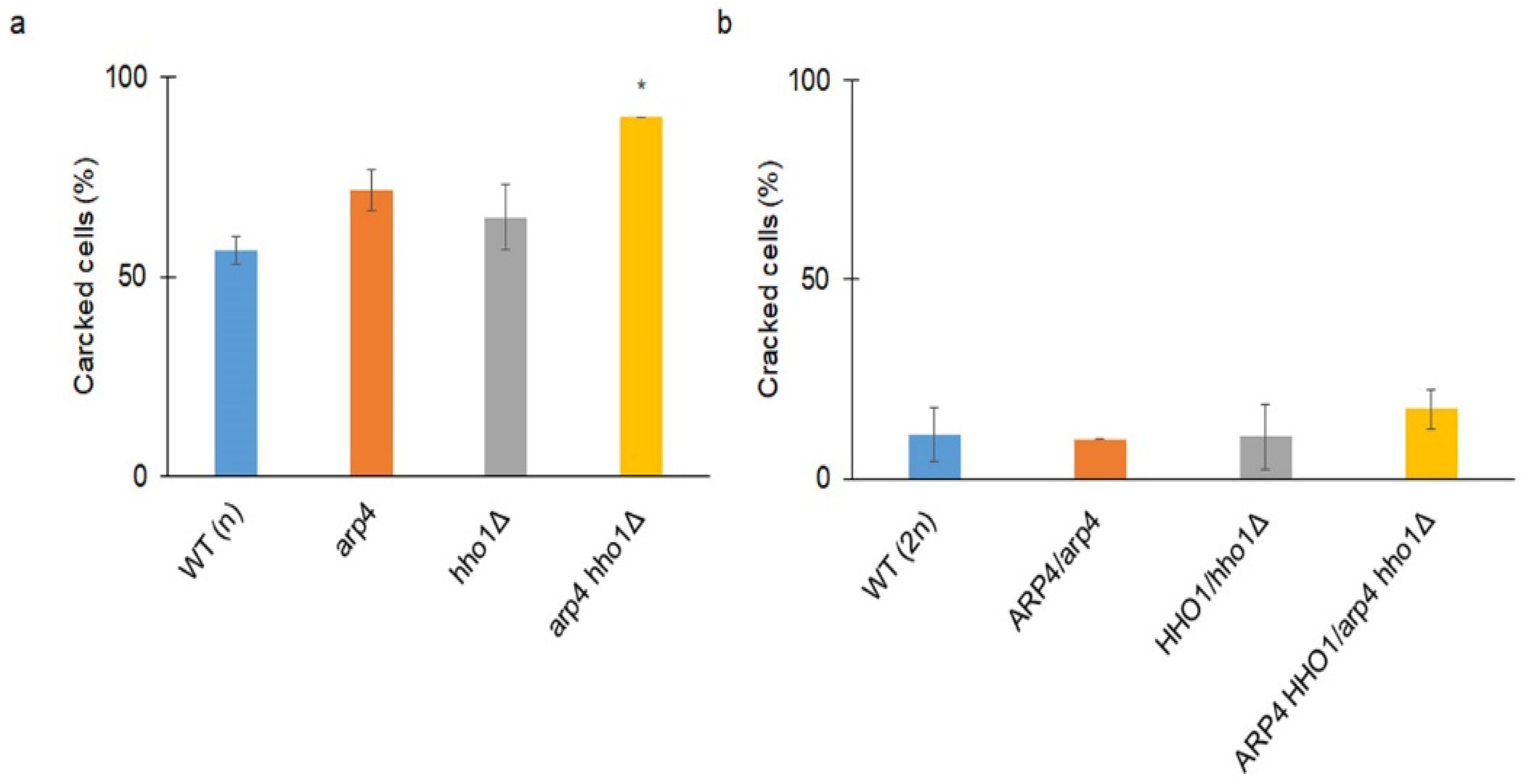
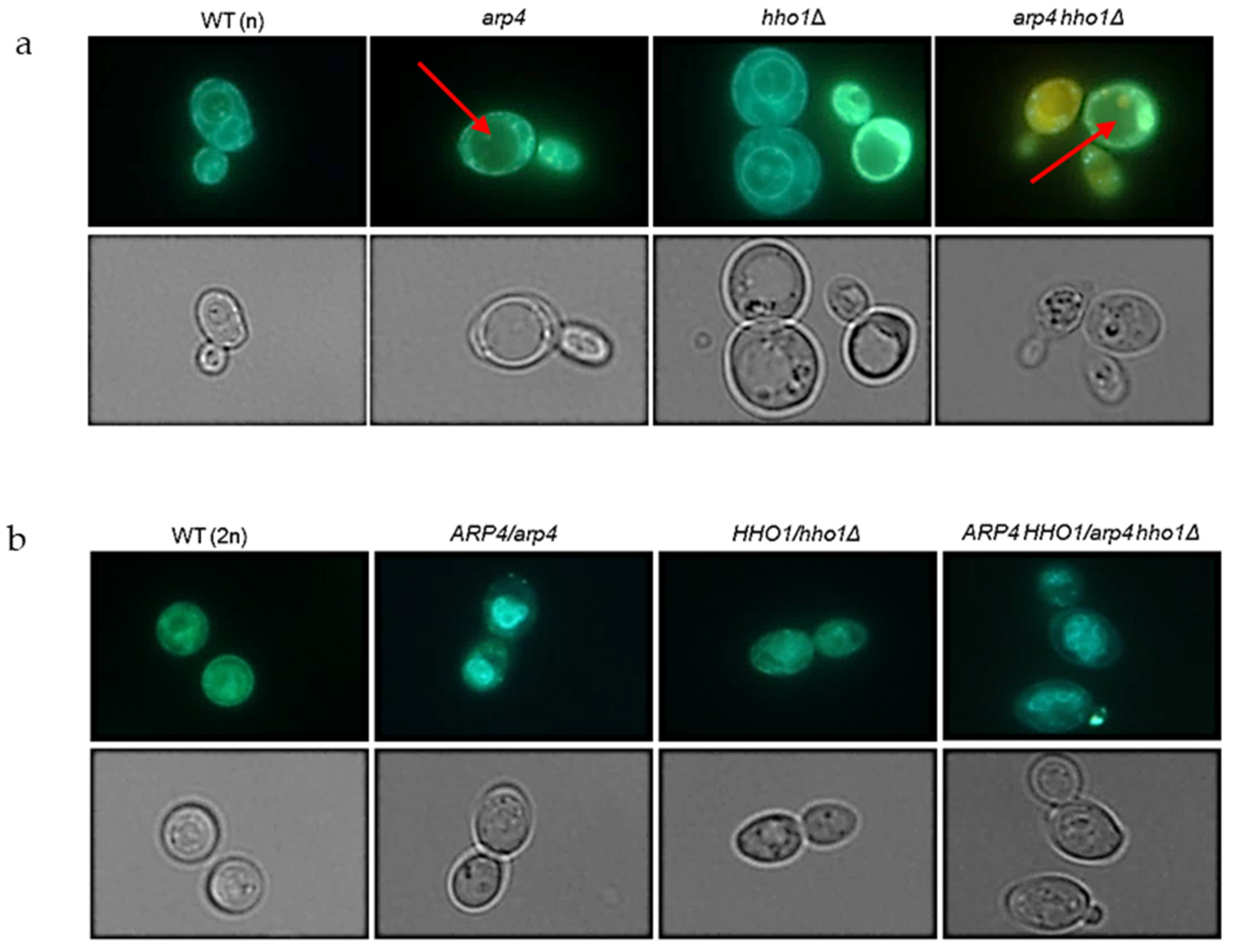
3.2. arp4 and arp4 hho1Δ Display Accelerated Replicative Ageing, Accompanied by Alterations in the Vacuolar, Nuclear and Cell Wall Fitness and Morphology
Translational Fitness
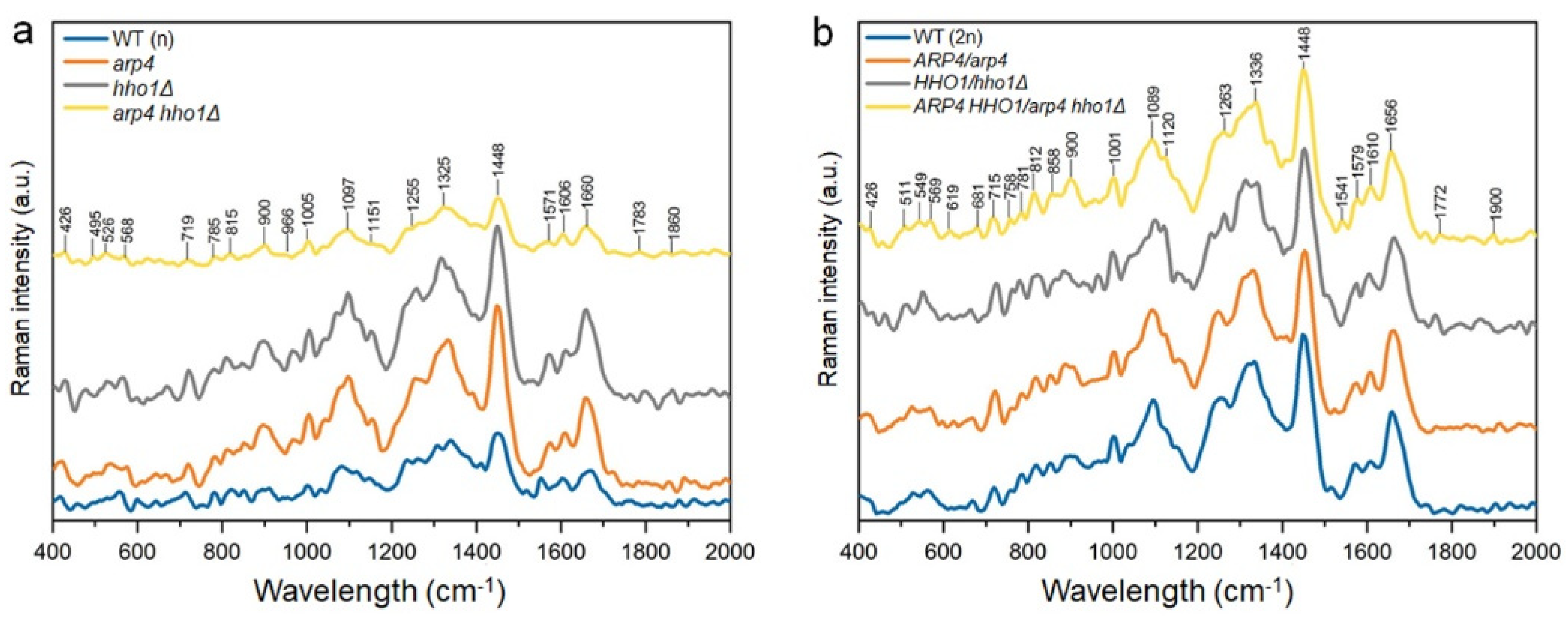
3.3. Changes in the Chemical Composition of the Haploid and Heterozygous Strains
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sahabi, S.; Jafari-Gharabaghlou, D.; Zarghami, N. A new insight into cell biological and biochemical changes through aging. Acta Histochem. 2022, 124, 151841. [Google Scholar] [CrossRef] [PubMed]
- Viña, J.; Borrás, C.; Miquel, J. Theories of ageing. IUBMB Life 2007, 59, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Breitenbach, M.; Laun, P.; Dickinson, J.R.; Klocker, A.; Rinnerthaler, M.; Dawes, I.W.; Aung-Htut, M.T.; Breitenbach-Koller, L.; Caballero, A.; Nyström, T.; et al. The role of mitochondria in the aging processes of yeast. Sub-Cell. Biochem. 2012, 57, 55–78. [Google Scholar] [CrossRef]
- Sinclair, D.A.; LaPlante, M.D. Lifespan: Why We Age—And Why We Don’t Have to; Atria Books: New York, NY, USA, 2019. [Google Scholar]
- Son, J.M.; Lee, C. Aging: All roads lead to mitochondria. Semin. Cell Dev. Biol. 2021, 116, 160–168. [Google Scholar] [CrossRef]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- López-Otín, C.; Galluzzi, L.; Freije, J.M.P.; Madeo, F.; Kroemer, G. Metabolic Control of Longevity. Cell 2016, 166, 802–821. [Google Scholar] [CrossRef]
- Covarrubias, A.J.; Perrone, R.; Grozio, A.; Verdin, E. NAD+ metabolism and its roles in cellular processes during ageing. Nat. Rev. Mol. Cell Biol. 2021, 22, 119–141. [Google Scholar] [CrossRef]
- Suganuma, T.; Workman, J.L. Chromatin and Metabolism. Annu. Rev. Biochem. 2018, 87, 27–49. [Google Scholar] [CrossRef]
- Mirisola, M.G.; Longo, V.D. Yeast Chronological Lifespan: Longevity Regulatory Genes and Mechanisms. Cells 2022, 11, 1714. [Google Scholar] [CrossRef]
- Kaeberlein, M. Lessons on longevity from budding yeast. Nature 2010, 464, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.S.; Kapahi, P.; Hsueh, W.C.; Kockel, L. TOR signaling never gets old: Aging, longevity and TORC1 activity. Ageing Res. Rev. 2011, 10, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, M.; Powers, R.W.; Steffen, K.K.; Westman, E.A.; Hu, D.; Dang, N.; Kerr, E.O.; Kirkland, K.T.; Fields, S.; Kennedy, B.K. Regulation of yeast replicative life span by TOR and Sch9 in response to nutrients. Science 2005, 310, 1193–1196. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.D.; Fabrizio, P. Chronological aging in Saccharomyces cerevisiae. Sub-Cell. Biochem. 2012, 57, 101–121. [Google Scholar] [CrossRef]
- Baccolo, G.; Stamerra, G.; Coppola, D.P.; Orlandi, I.; Vai, M. Chapter One—Mitochondrial Metabolism and Aging in Yeast. In International Review of Cell and Molecular Biology; López-Otín, C., Galluzzi, L., Eds.; Academic Press: Cambridge, MA, USA, 2018; Volume 340, pp. 1–33. [Google Scholar]
- Taormina, G.; Ferrante, F.; Vieni, S.; Grassi, N.; Russo, A.; Mirisola, M.G. Longevity: Lesson from Model Organisms. Genes 2019, 10, 518. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Kim, E.W.; Croteau, D.L.; Bohr, V.A. Heterochromatin: An epigenetic point of view in aging. Exp. Mol. Med. 2020, 52, 1466–1474. [Google Scholar] [CrossRef] [PubMed]
- Purohit, J.S.; Chaturvedi, M.M. Chromatin and Aging. In Topics in Biomedical Gerontology; Rath, P.C., Sharma, R., Prasad, S., Eds.; Springer: Singapore, 2017; pp. 205–241. [Google Scholar]
- Vasileva, B.; Staneva, D.; Krasteva, N.; Miloshev, G.; Georgieva, M. Changes in Chromatin Organization Eradicate Cellular Stress Resilience to UVA/B Light and Induce Premature Aging. Cells 2021, 10, 1755. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, M.; Staneva, D.; Uzunova, K.; Efremov, T.; Balashev, K.; Harata, M.; Miloshev, G. The linker histone in Saccharomyces cerevisiae interacts with actin-related protein 4 and both regulate chromatin structure and cellular morphology. Int. J. Biochem. Cell Biol. 2015, 59, 182–192. [Google Scholar] [CrossRef]
- Uzunova, K.; Georgieva, M.; Miloshev, G. Saccharomyces cerevisiae linker histone—Hho1p maintains chromatin loop organization during ageing. Oxidative Med. Cell. Longev. 2013, 2013, 437146. [Google Scholar] [CrossRef]
- Georgieva, M.; Roguev, A.; Balashev, K.; Zlatanova, J.; Miloshev, G. Hho1p, the linker histone of Saccharomyces cerevisiae, is important for the proper chromatin organization in vivo. Biochim. Biophys. Acta BBA Gene Regul. Mech. 2012, 1819, 366–374. [Google Scholar] [CrossRef]
- Georgieva, M.; Harata, M.; Miloshev, G. The nuclear actin-related protein Act3p/Arp4 influences yeast cell shape and bulk chromatin organization. J. Cell. Biochem. 2008, 104, 59–67. [Google Scholar] [CrossRef]
- Harata, M.; Zhang, Y.; Stillman, D.J.; Matsui, D.; Oma, Y.; Nishimori, K.; Mochizuki, R. Correlation between chromatin association and transcriptional regulation for the Act3p/Arp4 nuclear actin-related protein of Saccharomyces cerevisiae. Nucleic Acids Res. 2002, 30, 1743–1750. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, M.; Staneva, D.; Uzunova, K.; Miloshev, G. The Deletion of the Gene for the Linker Histone in ARP 4 Mutant Yeast Cells is not Deleterious. Biotechnol. Biotechnol. Equip. 2012, 26, 134–139. [Google Scholar] [CrossRef]
- Molon, M.; Szajwaj, M.; Tchorzewski, M.; Skoczowski, A.; Niewiadomska, E.; Zadrag-Tecza, R. The rate of metabolism as a factor determining longevity of the Saccharomyces cerevisiae yeast. Age 2016, 38, 11. [Google Scholar] [CrossRef] [PubMed]
- Krol, K.; Brozda, I.; Skoneczny, M.; Bretne, M.; Skoneczna, A. A Genomic Screen Revealing the Importance of Vesicular Trafficking Pathways in Genome Maintenance and Protection against Genotoxic Stress in Diploid Saccharomyces cerevisiae Cells. PLoS ONE 2015, 10, e0124186. [Google Scholar] [CrossRef]
- Molon, M.; Zebrowski, J. Phylogenetic relationship and Fourier-transform infrared spectroscopy-derived lipid determinants of lifespan parameters in the Saccharomyces cerevisiae yeast. FEMS Yeast Res. 2017, 17, fox031. [Google Scholar] [CrossRef]
- Minois, N.; Frajnt, M.; Wilson, C.; Vaupel, J.W. Advances in measuring lifespan in the yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2005, 102, 402–406. [Google Scholar] [CrossRef]
- Stepien, K.; Wojdyla, D.; Nowak, K.; Molon, M. Impact of curcumin on replicative and chronological aging in the Saccharomyces cerevisiae yeast. Biogerontology 2020, 21, 109–123. [Google Scholar] [CrossRef]
- Miloshev, G.; Staneva, D.; Uzunova, K.; Vasileva, B.; Draganova-Filipova, M.; Zagorchev, P.; Georgieva, M. Linker histones and chromatin remodelling complexes maintain genome stability and control cellular ageing. Mech. Ageing Dev. 2019, 177, 55–65. [Google Scholar] [CrossRef]
- Bryant, J.M.; Govin, J.; Zhang, L.Y.; Donahue, G.; Pugh, B.F.; Berger, S.L. The Linker Histone Plays a Dual Role during Gametogenesis in Saccharomyces cerevisiae. Mol. Cell. Biol. 2012, 32, 2771–2783. [Google Scholar] [CrossRef]
- Delobel, P.; Tesnière, C. A simple FCM method to avoid misinterpretation in Saccharomyces cerevisiae cell cycle assessment between G0 and sub-G1. PLoS ONE 2014, 9, e84645. [Google Scholar] [CrossRef] [Green Version]
- Smeal, T.; Claus, J.; Kennedy, B.; Cole, F.; Guarente, L. Loss of transcriptional silencing causes sterility in old mother cells of S. cerevisiae. Cell 1996, 84, 633–642. [Google Scholar] [CrossRef]
- Almeida, R.; Fernández-Justel, J.M.; Santa-María, C.; Cadoret, J.-C.; Cano-Aroca, L.; Lombraña, R.; Herranz, G.; Agresti, A.; Gómez, M. Chromatin conformation regulates the coordination between DNA replication and transcription. Nat. Commun. 2018, 9, 1590. [Google Scholar] [CrossRef] [PubMed]
- Ogiwara, H.; Ui, A.; Kawashima, S.; Kugou, K.; Onoda, F.; Iwahashi, H.; Harata, M.; Ohta, K.; Enomoto, T.; Seki, M. Actin-related protein Arp4 functions in kinetochore assembly. Nucleic Acids Res. 2007, 35, 3109–3117. [Google Scholar] [CrossRef] [PubMed]
- Aufschnaiter, A.; Büttner, S. The vacuolar shapes of ageing: From function to morphology. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 957–970. [Google Scholar] [CrossRef]
- Molon, M.; Molestak, E.; Kula-Maximenko, M.; Grela, P.; Tchorzewski, M. Ribosomal Protein uL11 as a Regulator of Metabolic Circuits Related to Aging and Cell Cycle. Cells 2020, 9, 1745. [Google Scholar] [CrossRef] [PubMed]
- Wawiórka, L.; Molestak, E.; Szajwaj, M.; Michalec-Wawiórka, B.; Mołoń, M.; Borkiewicz, L.; Grela, P.; Boguszewska, A.; Tchórzewski, M. Multiplication of Ribosomal P-Stalk Proteins Contributes to the Fidelity of Translation. Mol. Cell. Biol. 2017, 37, e00060-17. [Google Scholar] [CrossRef]
- Eleutherio, E.; Brasil, A.A.; França, M.B.; de Almeida, D.S.G.; Rona, G.B.; Magalhães, R.S.S. Oxidative stress and aging: Learning from yeast lessons. Fungal Biol. 2018, 122, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Tyler, J.K. Epigenetics and aging. Sci. Adv. 2016, 2, e1600584. [Google Scholar] [CrossRef] [PubMed]
- Erjavec, N.; Larsson, L.; Grantham, J.; Nystrom, T. Accelerated aging and failure to segregate damaged proteins in Sir2 mutants can be suppressed by overproducing the protein aggregation-remodeling factor Hsp104p. Genes Dev. 2007, 21, 2410–2421. [Google Scholar] [CrossRef]
- Dang, W.W.; Steffen, K.K.; Perry, R.; Dorsey, J.A.; Johnson, F.B.; Shilatifard, A.; Kaeberlein, M.; Kennedy, B.K.; Berger, S.L. Histone H4 lysine 16 acetylation regulates cellular lifespan. Nature 2009, 459, 802–807. [Google Scholar] [CrossRef] [Green Version]
- Kaeberlein, M.; McVey, M.; Guarente, L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev. 1999, 13, 2570–2580. [Google Scholar] [CrossRef] [PubMed]
- Imai, S.; Johnson, F.B.; Marciniak, R.A.; McVey, M.; Park, P.U.; Guarente, L. Sir2: An NAD-dependent histone deacetylase that connects chromatin silencing, metabolism, and aging. Cold Spring Harb. Symp. Quant. Biol. 2000, 65, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Frenk, S.; Houseley, J. Gene expression hallmarks of cellular ageing. Biogerontology 2018, 19, 547–566. [Google Scholar] [CrossRef] [PubMed]
- Yiu, G.; McCord, A.; Wise, A.; Jindal, R.; Hardee, J.; Kuo, A.; Shimogawa, M.Y.; Cahoon, L.; Wu, M.; Kloke, J.; et al. Pathways change in expression during replicative aging in Saccharomyces cerevisiae. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2008, 63, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Stepien, K.; Skoneczna, A.; Kula-Maximenko, M.; Jurczyk, L.; Molon, M. Depletion of the Origin Recognition Complex Subunits Delays Aging in Budding Yeast. Cells 2022, 11, 1252. [Google Scholar] [CrossRef]
- Javaheri, A.; Wysocki, R.; Jobin-Robitaille, O.; Altaf, M.; Cote, J.; Kron, S.J. Yeast G1 DNA damage checkpoint regulation by H2A phosphorylation is independent of chromatin remodeling. Proc. Natl. Acad. Sci. USA 2006, 103, 13771–13776. [Google Scholar] [CrossRef]
- Harata, M.; Oma, Y.; Mizuno, S.; Jiang, Y.W.; Stillman, D.J.; Wintersberger, U. The nuclear actin-related protein of Saccharomyces cerevisiae, Act3p/Arp4, interacts with core histones. Mol. Biol. Cell 1999, 10, 2595–2605. [Google Scholar] [CrossRef]
- Galarneau, L.; Nourani, A.; Boudreault, A.A.; Zhang, Y.; Heliot, L.; Allard, S.; Savard, J.; Lane, W.S.; Stillman, D.J.; Cote, J. Multiple links between the NuA4 histone acetyltransferase complex and epigenetic control of transcription. Mol. Cell 2000, 5, 927–937. [Google Scholar] [CrossRef]
- Rouse, J.; Jackson, S.P. Interfaces between the detection, signaling, and repair of DNA damage. Science 2002, 297, 547–551. [Google Scholar] [CrossRef]
- Zhou, B.B.S.; Elledge, S.J. The DNA damage response: Putting checkpoints in perspective. Nature 2000, 408, 433–439. [Google Scholar] [CrossRef]
- Thiriet, C.; Hayes, J.J. Chromatin in need of a fix: Phosphorylation of H2AX connects chromatin to DNA repair. Mol. Cell 2005, 18, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Kruhlak, M.J.; Celeste, A.; Dellaire, G.; Fernandez-Capetillo, O.; Muller, W.G.; McNally, J.G.; Bazett-Jones, D.P.; Nussenzweig, A. Changes in chromatin structure and mobility in living cells at sites of DNA double-strand breaks. J. Cell Biol. 2006, 172, 823–834. [Google Scholar] [CrossRef]
- Kirkwood, T.B.L. Evolution of ageing. Mech. Ageing Dev. 2002, 123, 737–745. [Google Scholar] [CrossRef]
- Zimmermann, A.; Hofer, S.; Pendl, T.; Kainz, K.; Madeo, F.; Carmona-Gutierrez, D. Yeast as a tool to identify anti-aging compounds. FEMS Yeast Res. 2018, 18, foy020. [Google Scholar] [CrossRef] [PubMed]
- Antikainen, H.; Driscoll, M.; Haspel, G.; Dobrowolski, R. TOR-mediated regulation of metabolism in aging. Aging Cell 2017, 16, 1219–1233. [Google Scholar] [CrossRef]
- Bitto, A.; Wang, A.M.; Bennett, C.F.; Kaeberlein, M. Biochemical Genetic Pathways that Modulate Aging in Multiple Species. Cold Spring Harb. Perspect. Med. 2015, 5, a025114. [Google Scholar] [CrossRef]
- Harman, D. Aging—A theory based on free-radical and radiation-chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef]
- Helsen, J.; Voordeckers, K.; Vanderwaeren, L.; Santermans, T.; Tsontaki, M.; Verstrepen, K.J.; Jelier, R. Gene Loss Predictably Drives Evolutionary Adaptation. Mol. Biol. Evol. 2020, 37, 2989–3002. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Appears in Text | Genotype | Source |
|---|---|---|---|
| Haploids | |||
| DY2864 | WT (n) | MATa his4-912Δ-ADE2 his4-912Δ lys2-128Δ can1 trp1 ura3 ACT3 | [24] |
| DY4285 | arp4 | MATa his4-912Δ-ADE2 lys2-128Δ can1 leu2 trp1 ura3 act3-ts26 | [24] |
| DY2864 hho1Δ | hho1Δ | MATa his4-912Δ-ADE2 his4-912Δ lys2-128Δ can1 trp1 ura3 ACT3 ypl127C::K.L.URA3 | [20] |
| DY4285 hho1Δ | arp4 hho1Δ | MATa his4-912Δ-ADE2 lys2-128Δ can1 leu2 trp1 ura3 act3-ts26 ypl127C::K.L.URA3 | [25] |
| Diploids | |||
| BY4742/DY2864 | WT (2n) | MATα/a ADE2/his4-912Δ-ADE2 his3-Δ1/his4-912Δ leu2Δ0/LEU2 lys2Δ0/lys2-128Δ CAN1/can1 TRP1/trp1 ura3-Δ0/ura3 ACT3/ACT3 | this study |
| BY4742/DY4285 | ARP4/arp4 | MATα/a ADE2/his4-912Δ-ADE2 lys2Δ0/lys2-128Δ CAN1/can1 leu2Δ0/leu2 TRP1/trp1 ura3-Δ0/ura3 ACT3/act3-ts26 (ARP4/arp4-ts26) | this study |
| BY4742/DY2864 hho1Δ | HHO1/hho1Δ | MATα/a ADE2/his4-912Δ-ADE2 his3-Δ1/his4-912Δ leu2Δ0/LEU2 lys2Δ0/lys2-128Δ CAN1/can1 TRP1/trp1 ura3-Δ0/ura3 ACT3/ACT3 YPL127C/ypl127C::K.L.URA3 (HHO1/hho1Δ) | this study |
| BY4742/DY4285 hho1Δ | ARP4 HHO1/arp4 hho1Δ | MATα/a ADE2/his4-912Δ-ADE2 lys2Δ0/lys2-128Δ CAN1/can1 leu2Δ0/leu2 TRP1/trp1 ura3-Δ0/ura3 ACT3/act3-ts26 YPL127C/ypl127C::K.L.URA3 (HHO1//hho1Δ) | this study |
| Diploids | Haploids | |||||||
|---|---|---|---|---|---|---|---|---|
| WT (2n) | ARP4/arp4 | HHO1/hho1Δ | ARP4 HHO1/arp4 hho1Δ | WT (n) | arp4 | hho1Δ | arp4 hho1Δ | Vibrations |
| Peaks Position (Raman Shift, cm−1) | ||||||||
| 1660 | 1660 | 1664 | 1656 | 1664 | 1660 | 1660 | 1660 | Lipids, Proteins (2, 4–6) |
| 1610 | 1606 | 1606 | 1610 | - | 1610 | 1610 | 1606 | Mitochondria, Proteins (5, 7, 8) |
| 1571 | 1575 | 1575 | 1579 | 1579 | 1575 | 1572 | 1572 | RNA/DNA (9, 11) |
| 1514 | 1525 | - | 1540 | 1521 | - | 1533 | - | Amide II (11) |
| 1448 | 1452 | 1452 | 1448 | 1448 | 1448 | 1448 | 1452 | Proteins (5, 6) |
| - | 1394 | 1378 | 1371 | - | 1390 | - | 1394 | RNA/DNA (9, 11) |
| 1332 | 1332 | 1340 | 1336 | 1340 | 1332 | 1336 | 1324 | RNA (3, 9, 10) |
| 1255 | 1247 | 1263 | 1263 | 1274 | 1259 | 1266 | Lipids (1, 4) | |
| 1097 | 1093 | 1101 | 1089 | 1082 | 1097 | 1097 | 1097 | Polysaccharides, RNA/DNA (3,5,6) |
| 1001 | 1001 | 1001 | 1001 | 1005 | 1005 | 1005 | 1005 | Proteins, Phenylalanine, RNA (4, 6, 8) |
| 966 | 950 | 966 | 958 | 962 | 969 | 966 | 939 | Polysaccharides (1–3) |
| 900 | 889 | 897 | 897 | 912 | 897 | 897 | 900 | Polysaccharides (1, 2, 6) |
| 854 | 854 | - | 858 | 854 | 850 | 846 | 854 | Proteins (5, 6, 8) |
| 819 | 819 | 823 | 815 | 823 | 811 | 807 | 819 | RNA, Proteins (11) |
| 784 | 784 | 781 | 784 | 785 | 781 | 781 | 785 | RNA/DNA (9, 11) |
| 757 | 761 | 761 | 757 | - | - | - | - | Proteins, Cyt c (11, 12) |
| 719 | 723 | 723 | 715 | 711 | 719 | 719 | 719 | RNA (3, 9, 10) |
| 669 | 669 | 665 | 680 | 669 | 669 | 669 | 649 | RNA/DNA (3, 9, 11) |
| - | 622 | 611 | 615 | 626 | 611 | 623 | 623 | |
| 564 | 568 | 549 | 541 | 557 | 572 | 564 | 568 | |
| 526 | 526 | 514 | 510 | 530 | 537 | 526 | 526 | RNA, Polysaccharides (3, 10) |
| - | 418 | 429 | 426 | 422 | 422 | 430 | 426 | Polysaccharides (1, 2, 6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mołoń, M.; Stępień, K.; Kielar, P.; Vasileva, B.; Lozanska, B.; Staneva, D.; Ivanov, P.; Kula-Maximenko, M.; Molestak, E.; Tchórzewski, M.; et al. Actin-Related Protein 4 and Linker Histone Sustain Yeast Replicative Ageing. Cells 2022, 11, 2754. https://doi.org/10.3390/cells11172754
Mołoń M, Stępień K, Kielar P, Vasileva B, Lozanska B, Staneva D, Ivanov P, Kula-Maximenko M, Molestak E, Tchórzewski M, et al. Actin-Related Protein 4 and Linker Histone Sustain Yeast Replicative Ageing. Cells. 2022; 11(17):2754. https://doi.org/10.3390/cells11172754
Chicago/Turabian StyleMołoń, Mateusz, Karolina Stępień, Patrycja Kielar, Bela Vasileva, Bonka Lozanska, Dessislava Staneva, Penyo Ivanov, Monika Kula-Maximenko, Eliza Molestak, Marek Tchórzewski, and et al. 2022. "Actin-Related Protein 4 and Linker Histone Sustain Yeast Replicative Ageing" Cells 11, no. 17: 2754. https://doi.org/10.3390/cells11172754
APA StyleMołoń, M., Stępień, K., Kielar, P., Vasileva, B., Lozanska, B., Staneva, D., Ivanov, P., Kula-Maximenko, M., Molestak, E., Tchórzewski, M., Miloshev, G., & Georgieva, M. (2022). Actin-Related Protein 4 and Linker Histone Sustain Yeast Replicative Ageing. Cells, 11(17), 2754. https://doi.org/10.3390/cells11172754