1. Introduction
Osteoarthritis (OA) is the most common joint disease and has increasingly affected societies worldwide, especially in middle-aged and elderly populations [
1,
2]. However, current OA treatment strategies, including conservative and surgical treatments, can only serve as the last resort to temporarily relieve the clinical symptoms and joint destruction [
3,
4,
5]. Hence, a new and effective treatment is urgently needed. Furthermore, the onset age of OA has been gradually decreasing, and thus, young people have also increasingly become susceptible to OA [
6,
7]. However, because of poor attention to early symptoms and the lack of reliable biomarkers for early diagnosis before any irreversible damage occurs, many patients in the early stage of OA often miss the optimal treatment opportunity [
8,
9]. In fact, timely attention and effective treatment are both important in OA [
9,
10].
As impaired joint homeostasis is the core of articular cartilage (AC) injury in OA [
11], researchers have employed various strategies to maintain joint homeostasis to protect joint health [
12,
13]. Combined with the definition of “homeostasis”, first proposed by Claude Bernard in 1852 [
14], we believe that joint homeostasis refers to the steady state of all joint structures and can be achieved by maintaining the liquid matrix environment surrounding the joints. As the liquid matrix that surrounds the main structures of the joint, synovial fluid is the primary medium for communication between the tissue cells [
15,
16]. Synoviocytes in the joint can maintain joint homeostasis by releasing bioactive molecules into the synovial fluid, thereby communicating and interacting with the chondrocytes [
17,
18,
19,
20]. During the pathogenesis of OA, this homeostasis is impaired. Moreover, as the disease progresses, inflammatory synoviocytes release bioactive substances to anatomical spaces other than the joint cavity, and thus, these substances can disseminate through blood circulation. Consequently, the homeostasis is impaired not only at the injured joint but also at other locations in the body. Therefore, timely intervention and control of the pathological processes underlying the impaired joint homeostasis induced by synovial initiation is a new therapeutic strategy against OA. However, the mechanisms affecting joint homeostasis are currently unclear. To elucidate the mechanism of bioactive substance delivery in joint homeostasis, we established an in vitro cell model in this study and examined the fibroblast-like synoviocytes (FLSs), the chondrocytes, and their surrounding liquid environment as the major study objects. We hypothesized that a mechanism of bioactive substance delivery from FLSs to chondrocytes could impact joint homeostasis in OA.
MicroRNAs (miRNAs) are endogenous, non-coding, single-stranded small RNAs that degrade or block the translation of their target mRNAs and thereby downregulate corresponding proteins [
21]. From cell-fate decision to signal transduction, miRNAs are widely involved in cellular processes, including the pathogenesis of OA [
22]. In recent years, several miRNAs have been used as biomarkers for the diagnosis or prognosis of several diseases [
23,
24]. The miRNA-mediated novel therapeutic approaches have been applied to various diseases [
21,
25]. In this study, we first identified that miRNA-150-3p (miR-150-3p) is downregulated in circulating extracellular vesicles (EVs) with OA, and we then investigated the intercellular delivery mechanism and the biological functions of this miRNA.
Hence, we urgently needed a miRNA “enricher” to enable us to detect critical miRNAs and a miRNA “transporter” to deliver the miRNAs during the communication among different cells [
26]. EVs, nanomolecular vesicle-like structures made of a double-layer lipid membrane, can be actively released into the extracellular environment and body fluids by almost all types of cells under physiological and pathological conditions. EVs can transport numerous miRNAs and biologically active substances released by secretory cells to nearby or distant recipient cells. These nanostructures act as specific vectors for information transmission between cells and thereby regulate the functions of the recipient cells [
27,
28]. We hypothesized that miRNA-containing EVs secreted by FLSs could serve as extracellular regulators of joint homeostasis and thereby regulate the OA-related pathways by targeting chondrocytes as recipient cells.
The innate immune system is the decisive barrier of the body against infections and is involved in the synovial inflammation and AC catabolic events underlying OA pathogenesis [
29,
30,
31]. The T cell receptor (TCR)-interacting molecule (TRIM) family proteins are closely associated with the innate immune response. We found that, as the target gene of miR-150-3p, Trim14 can regulate the expression of the proteins involved in the nuclear factor kappa B (NF-κB) signal transduction pathway, activate the innate immune response, and modulate joint homeostasis in OA. We also demonstrated the effect of a healthy synovial membrane (SM) on joint homeostasis in OA. We investigated the mechanism of the EV-mediated specific delivery of miR-150-3p from FLSs to chondrocytes, the primary effector cells in the synovium and cartilage that participate in joint homeostasis [
32,
33]. We also investigated the mechanism of joint homeostasis through the involvement of the Trim14/NF-κB/interferon-β (IFN-β) axis in the innate immune response. We preliminarily identified the optimal window for applying miR-150-3p-carrying EVs (EVs-150) as a potential treatment for cartilage protection and OA intervention. These observations provide new insights into whether EV-mediated miRNAs can be used as diagnostic, therapeutic, and prognostic biomarkers in OA, thereby achieving joint protection.
2. Methods
2.1. Construction and Validation of the Animal Model
Specific-pathogen-free female Wistar rats (8 weeks old and 180–220 g) (SLAC, Shanghai, China) were raised for one week under conventional housing conditions to adapt to the environment before the experiments. After being anesthetized via intraperitoneal injection of 2% pentobarbital sodium (Sinopharm, Beijing, China), the rats were fixed in a supine position. Unilateral knee OA was induced within the rats (model group) through anterior cruciate-ligament transection (ALCT) (
Figure 1A). Then, the rats were intramuscularly administered 30,000 U/d penicillin (North China Pharmaceutical Co, Hebei, China) once a day for three consecutive days after the surgery to prevent infection. The mobility of the rats on the surgical side was maintained, and the food and water intake, changes in hair color, incision healing, as well as suture intactness, and incidence of infection were closely monitored. Healthy rats that were not subjected to the surgery were set as the control group and housed under the same conditions. All the animals were provided with water and the conventional pellet food and housed in a quiet and comfortable environment. The study was approved by the Animal Experiment Ethics Committee of China-Japan Friendship Hospital (No. 180117).
Whole blood was collected from the abdominal aorta of the rats in the two groups after 10 weeks and centrifuged at 1900× g for 10 min to separate the serum. These serum samples were then stored at −80 °C until needed. The entire knee joint of the rats in each group was harvested, fixed in 4% paraformaldehyde, and stained with hematoxylin and eosin (HE) to validate the successful establishment of the OA animal model.
2.2. EV Extraction and Identification
The EVs were isolated from the serum samples from each group by using differential centrifugation as follows: 500× g for 10 min, 2000× g for 30 min, and 10,000× g for 30 min. Afterwards, the supernatants containing the EVs were collected, filtered through 0.22 µm sterile filters into centrifuge tubes, topped up to 20 mL with sterile PBS, and then centrifuged at 120,000× g for 1 h (Optima XE, Beckman, Fort Collins, CO, USA). All of the above procedures were conducted at 4 °C in sterile centrifuge tubes. Finally, after carefully removing the supernatant, the pellets were resuspended with sterile PBS and stored at −80 °C for subsequent experiments.
The extracted EVs were identified using transmission electron microscopy (TEM, Tecnai f20, Philips, Amsterdam, Netherlands), nanoparticle tracking analysis (NTA, NS300, NanoSight, Malvern, UK), and Western blotting (WB). After the extracted particles were stained with phosphotungstic-acid solution, TEM was used to observe and photograph the extracted particles and determine whether their morphology was consistent with the general characteristics of EVs. NTA was used to measure the diameter and concentration of the extracted particles (diluted for 30 folds in sterile phosphate-buffered saline [PBS, Corning Incorporated, Corning, NY, USA]). WB was used to detect EV marker proteins, including CD9, CD63, and HSP70 (Abcam, Cambridge, MA, USA), and the detailed specific procedures are illustrated in the “WB” section.
2.3. miRNA Sequencing and Validation
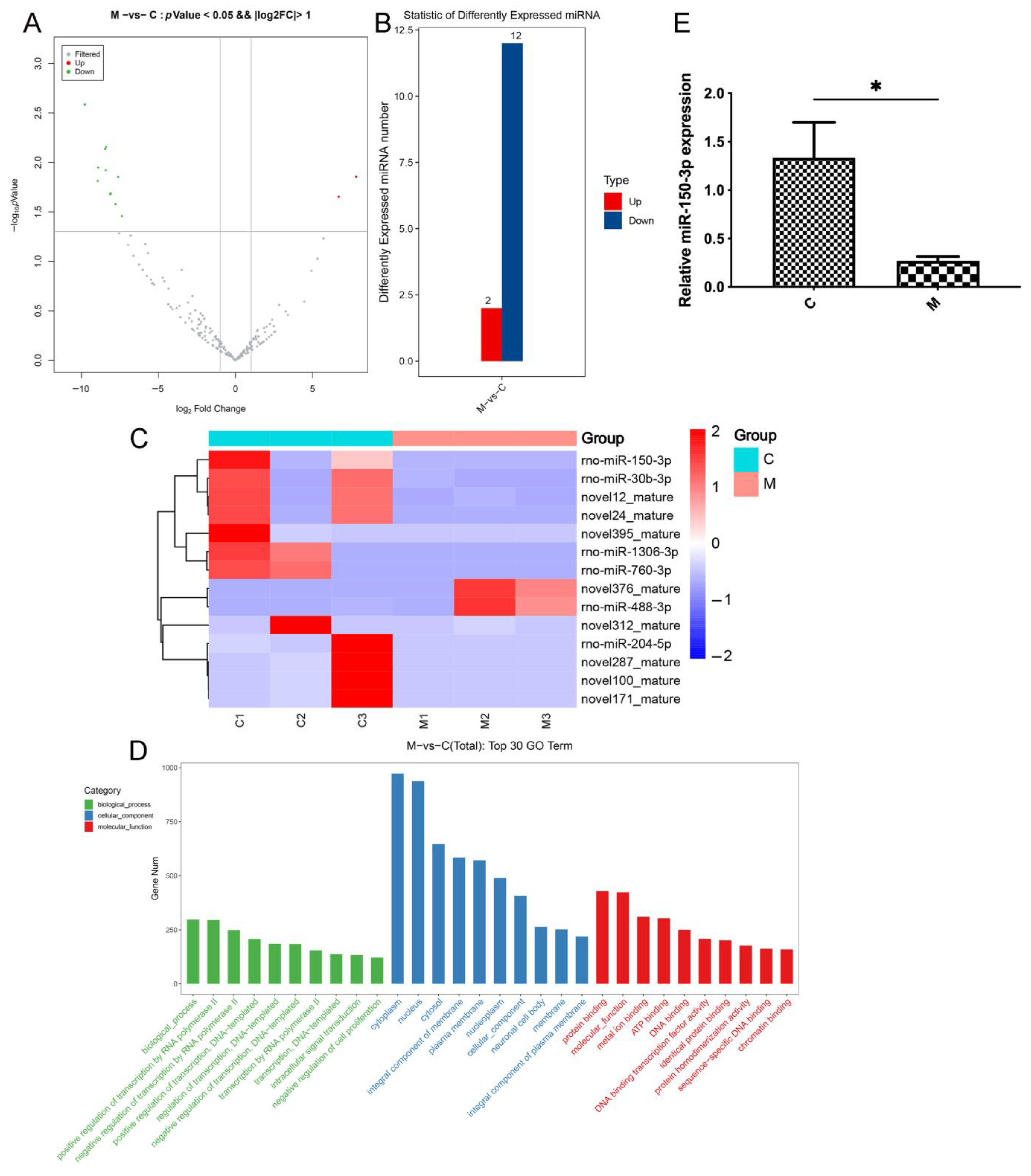
The EV miRNAs were analyzed using high-throughput sequencing. The procedures of the bioinformatic analysis were as follows: The raw data were processed and statistically analyzed; then, quality control, length-distribution analysis, database comparison and filtration, gene comparison, and analysis of the gene-expression level were conducted on known and predicted miRNAs to identify the differentially expressed (DE) miRNAs (YanzaiBio Co, Shanghai, China). The DE miRNAs were then distributed on a volcano plot, and a heatmap was employed to cluster these miRNAs according to their expression levels. The DE miRNAs were validated using reverse transcription-quantitative polymerase chain reaction (qPCR) and subjected to gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses to predict their target genes, associated pathways, and putative functional roles.
2.4. Construction of the In Vitro Cell Model
2.4.1. Cell Isolation and Culture
The OA rat model was constructed using the methods described in the sub-section “Construction and validation of the animal model”, which should be performed under sterile conditions for the subsequent cell culture. The SM was harvested from the knee joint of the surgical side and rinsed several times with Dulbecco’s PBS (DPBS, Corning Incorporated, Corning, NY, USA). After the removal of the attached surrounding tissues (fat, ligaments, fascia, etc.), the synovium was minced and digested with 5 mL type II collagenase (Sigma-Aldrich, St. Louis, MO, USA) for 4 h at 37 °C. The solution was gently shaken several times during the digestion. The collected cell suspension was filtered through a 100-mesh filter and centrifuged at 300× g for 5 min. Then, the cells were seeded into culture flasks and passaged when the cell density reached 70–80%. The cells passaged 3–5 times were used in the experiments. The isolation and culture of the healthy synoviocytes and chondrocytes were performed using the same methods as were used with the OA synoviocytes, except that the chondrocytes were harvested from the AC and then digested for ≥6 h. The morphology of the isolated and cultured synoviocytes and chondrocytes was examined under a light microscope (IX-70, Olympus, Tokyo, Japan).
Flow cytometry (FCM) was utilized to detect the viability of the synoviocytes and chondrocytes, along with the synoviocyte surface markers CD90 (FLSs), CD68 (macrophages), and CD3 (T cells), and the mesenchymal stem cell (MSC) surface markers CD29 and CD105 were also labeled. Vimentin, the characteristic labeled protein of FLSs, was tagged with the SM and FLSs through immunohistochemical (IHC) and immunofluorescence (IF) analysis to further identify FLSs. Toluidine-blue staining was used to examine the developing glycosaminoglycans in the extracellular matrix of the isolated chondrocytes, and their expression of type II collagen (COLII) was assessed via IF analysis. The production of glycosaminoglycans and COLII is characteristic of chondrocytes.
2.4.2. Model Construction
To study the mechanisms of EV-mediated miRNA delivery and its role in OA, we constructed an in vitro OA model. Two different cell types were co-cultured without contacting each other using a transwell system (8 µm, Corning Incorporated, Corning, NY, USA). In this cell model, FLSs and chondrocytes were cultured in the upper and lower chambers as secretory and recipient cells, respectively, with Dulbecco’s modified Eagle’s medium/F12 (DMEM/F12, Gibco, Carlsbad, CA, USA), containing 10% fetal bovine serum (FBS, Gibco, Carlsbad, CA, USA). Bioactive factors can realize the communication between the two cell types through this liquid environment and across the permeable polycarbonate membrane of the transwell system and exert their biological functions.
2.5. miRNA Transportation in the In Vitro OA Model
miR-150-3p mimics were labeled with Cy3 to observe their location. FLSs were transfected with the Cy3-labeled mimics (GenerayBiotech, Shanghai, China) according to the manufacturer’s instructions. The sequences were as follows: miR-150-3p mimic, forward 5′-CUGGUACAGGCCUGGGGGA-3′, reverse 5′-CCCCAGGCCUGUACCAGUU-3′.
The transfected FLSs were seeded in the upper chamber and co-cultured with chondrocytes pre-seeded in the lower chamber. To assess whether the miR-150-3p mimics transfected into the FLSs in the upper chamber had been released and then absorbed by the chondrocytes in the lower chamber, the localization of the red Cy3 fluorescence was characterized. As a control, another group of FLSs was treated with Cy3 that was not conjugated to miR-150-3p and then co-cultured with the chondrocytes. Under a fluorescence microscope, the chondrocytes were examined for the presence of the red Cy3 fluorescence signal to address the effect of Cy3 on them alone.
In addition, to assess whether EVs are involved in the delivery of miR-150-3p from FLSs to chondrocytes, we treated another set of the in vitro OA model transfected with the Cy3-labeled miR-150-3p mimics with the EV inhibitor GW4869 (Sigma-Aldrich, St. Louis, MO, USA) (10 µM).
2.6. FLS–EV Extraction and Identification
Given the high levels of miR-150-3p in the circulating EVs of healthy rats, we considered that miR-150-3p might positively impact the injured joints in OA. However, to determine whether EVs-150 in the circulation are associated with FLSs, which participate in OA progression, and whether FLSs can secrete EVs, we isolated FLSs from healthy rats and cultured them as described above. The FLSs were passaged until they reached the required cell quantity. Then, they were collected, rinsed three times with PBS, and transferred to a fresh serum-free EV medium (Gibco, Carlsbad, CA, USA). After 48 h, the culture supernatant was collected and its EV content was extracted via differential centrifugation as described above. TEM, NTA, and WB (CD9, TSG101, and HSP70 [Abcam, Cambridge, MA, USA]) were used to characterize the EVs.
2.7. EV-Uptake Assay
To detect the uptake of FLS–EVs by chondrocytes in vitro, we labeled the FLS–EVs using the PKH67 green-fluorescent kit (Sigma-Aldrich, St. Louis, MO, USA). For subsequent experiments, the EVs were re-extracted via differential centrifugation and quantitated using a bicinchoninic acid (BCA, Sigma-Aldrich, St. Louis, MO, USA) assay. To observe the uptake of EVs by chondrocytes, the chondrocyte nuclei were first labeled with the Hoechst stain (Sigma-Aldrich, St. Louis, MO, USA) for 15 min to aid the localization of the cells, and then, the cells were rinsed three times with PBS. With the addition of the EVs (50 mg/L final concentration), the EV uptake of the chondrocytes was observed and recorded after 1, 6, 12, and 24 h. The other group was pre-treated with GW4869 for 24 h before the addition of the EVs, and the change in fluorescence intensity after the EV blockage was observed.
2.8. In Vitro EV-150 Functional Assays
To further study the functions of EVs, we treated the chondrocytes with GW4869 or FLS–EVs isolated from healthy rats and compared these cells with untreated chondrocytes. qPCR was used to evaluate the effect of the EV treatment on the expression of miR-150-3p in the chondrocytes, to confirm that this miRNA was enriched in EVs and transported by them and to determine whether GW4869 terminated this process.
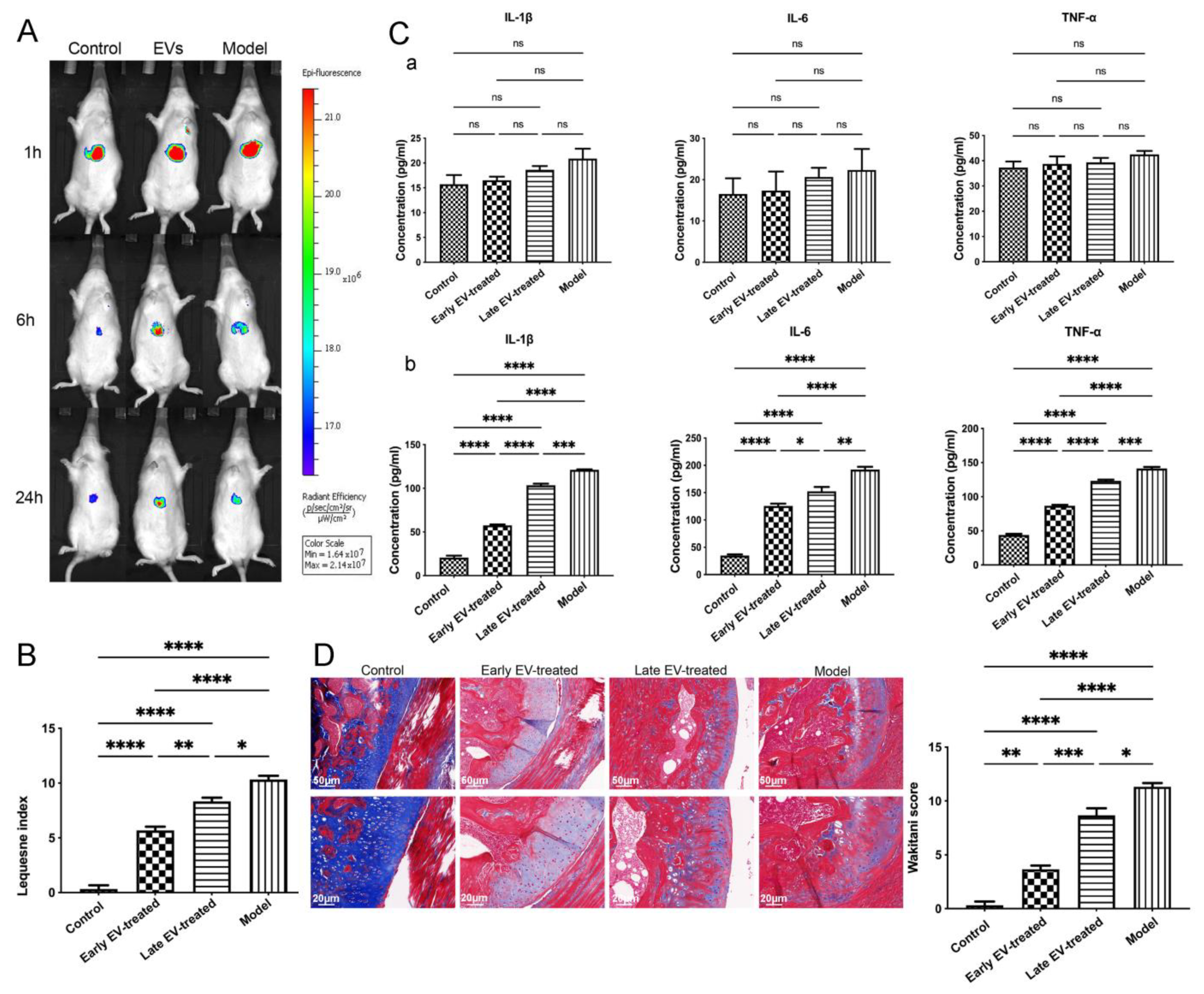
To assess the regulatory effect of EVs-150 on the chondrocytes of OA, the expression levels of COLII and aggrecan (ACAN) were measured using qPCR and WB. The concentrations of the pro-inflammatory cytokines interleukin-1β (IL-1β), IL-6, and tumor necrosis factor-α (TNF-α) in the chondrocyte culture medium were measured using an enzyme-linked immunosorbent assay (ELISA).
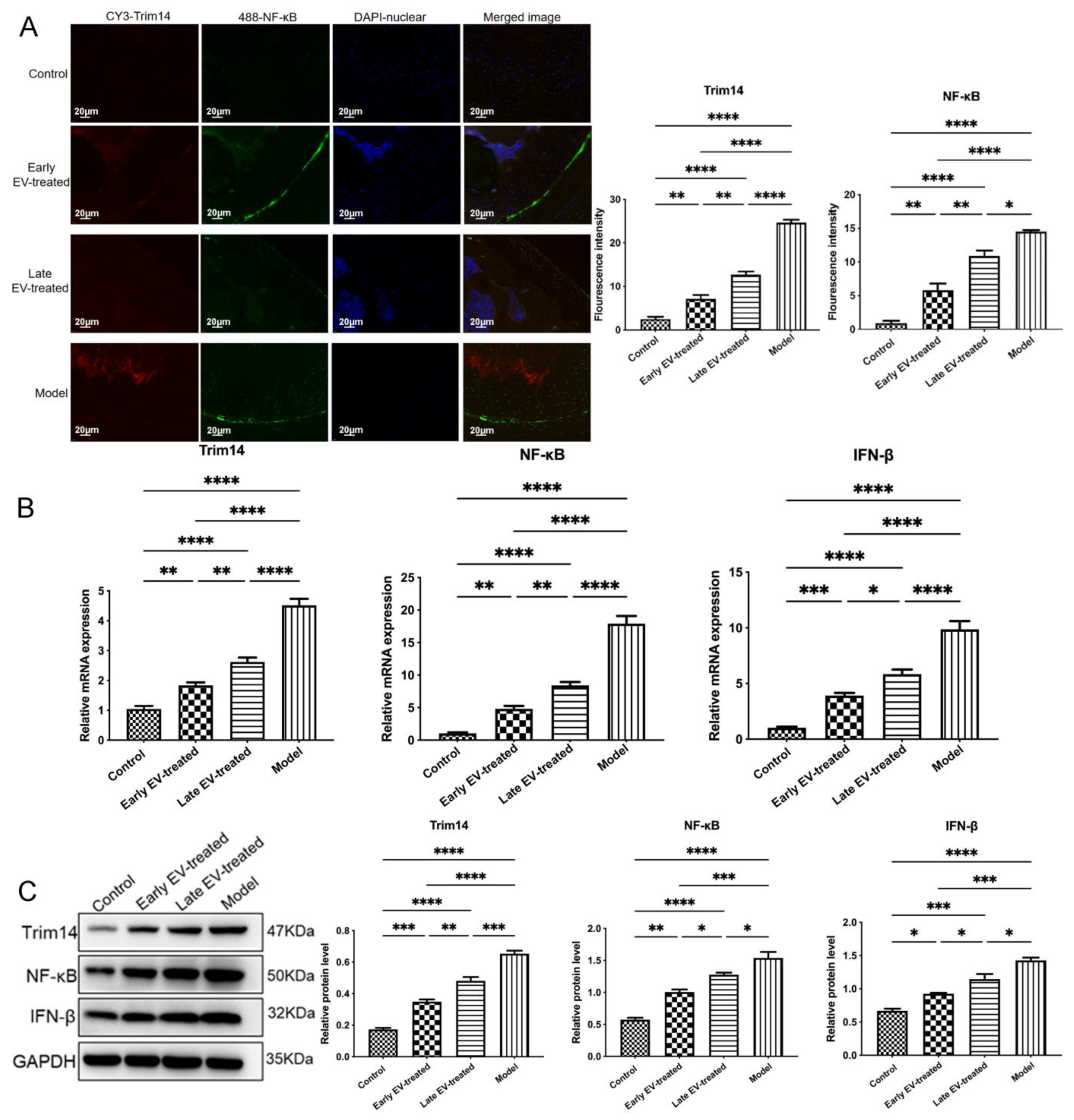
Furthermore, to investigate the mechanism whereby FLS–EVs from healthy rats regulate OA, we predicted the target genes of miR-150-3p and associated pathways via GO enrichment analysis. Given the role of the innate immune response in OA that we showed in our previous study [
34], the effects of EVs-150 on the expression of the target gene Trim14 and the downstream genes NF-κB and IFN-β, three genes involved in the innate immune response, were analyzed using qPCR and WB.
To further study the functions of EVs-150, we used TargetScan to predict the targets of miR-150-3p. Consequently, we predicted Trim14 as a potential miR-150-3p target and identified the putative binding site of miR-150-3p on Trim14. These results were validated using the dual luciferase assay. To this end, we constructed a set of pGL3 luciferase reporter vectors with the promoter region composed of the identified miR-150-3p binding sites on the Trim14 promoter. Next, 293T cells (Cell Bank of Chinese Academy of Sciences, Shanghai, China) were transfected with a vector expressing miR-150-3p or the negative control (NC) vector. The sequences (Generay Biotech, Shanghai, China) used for the dual-luciferase assay were: pGL3-Trim14-3′ UTR-WT, TTCTGAGCTGGGGTTTGTGCTGGC; pGL3-Trim14-3′ UTR-MUT, TAGTGAGGTGCCGTTACAGGTGCC; miR-150-3p mimic NC, forward 5′-UUCUCCGAACGUGUCACGUTT-3′, reverse 5′-ACGUGACACGUUCGGAGAATT-3′.
2.9. Regulation of the Innate Immune Signaling by miR-150-3p
To elucidate the molecular mechanism underlying the effect of miR-150-3p on joint homeostasis during OA, we transfected FLSs with a miR-150-3p mimic, miR-150-3p inhibitor, or NC by using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Then, the cells were transferred to the upper chamber of the transwell system in the cell model and evaluated for their ability to influence the chondrocytes in the lower chamber. The pro-inflammatory cytokines IL-1β, IL-6, and TNF-α in the culture medium were quantitated using ELISA. The expression levels of COLII, ACAN, Trim14, NF-κB, and IFN-β in the chondrocytes of each group were determined via qPCR and WB. The sequences of the oligonucleotides (Generay Biotech, Shanghai, China) used were as follows: miR-150-3p inhibitor, 5′-UCCCCCAGGCCUGUACCAG-3′; miR-150-3p inhibitor NC, 5′-GGCCUCACCGGGUGUAAAUCAG-3′.
2.10. In Vivo EV-150 Functional Assays
Healthy FLS-derived EVs (H-FLS–EVs) were labeled with the DiR fluorescent dye (KeyGEN, Nanjing, China). The labeled EVs were re-extracted to remove impurities and then injected into rats with OA via the caudal vein (the construction of the OA rat model is described above). The fluorescence distribution in the animals was observed after 1, 6, and 24 h via live imaging (IVIS Lumina XRMS Series III, PerkinElmer, Waltham, MA, USA). Likewise, the DiR fluorescent dye without EVs was injected into another set of healthy or OA rats, as a control.
The H-FLS–EVs were injected inside the rats through the tail vein, and four-week treatment in the early stages (early EV-treated group) of OA was compared with the same treatment period in the late stages (late EV-treated group). The rats were grouped and processed as follows: In the early EV-treated group, EV injection (500 µg/kg) was conducted on days 1 and 4, from the fourth to the seventh week after the surgery; in the late EV-treated group, EV injection was conducted on days 1 and 4, from the seventh to the tenth week after the surgery; in the model group, the animals were not subjected to any intervention after the surgery; and in the control group, healthy rats from the same batch of animals that had been used to construct the OA model were used. The behavior of the rats within each group was assessed based on the modified Lequesne MG index [
35]. It had the stimulus–response of local pain, altered gait, joint motion range, and joint swelling degree. Blood and joint fluid were collected from each group on the last day of week 10, and the concentrations of the pro-inflammatory cytokines IL-1β, IL-6, and TNF-α were measured using ELISA. The cartilage tissues were harvested and stained with Masson to perform histological grading through the Wakitani scoring system. qPCR was used to measure the miR-150-3p level in the serum EVs of each group. The effects of EVs-150 on the expression levels of COLII, ACAN, Trim14, NF-κB, and IFN-β in the AC of each group were determined via qPCR, WB, and IF to investigate the regulatory functions of EVs-150 in OA.
2.11. Cell Viability and FLS Characterization of FLSs through FCM
The FLSs or chondrocytes were digested with 0.25% trypsin containing 0.05% EDTA (Gibco), centrifuged at 300× g for 5 min, collected, and then washed twice in the stain buffer (BD Pharmingen, San Diego, CA, USA). After the cell pellets were resuspended in the stain buffer, the cell concentration in each sample was adjusted to 1 × 107 cells/mL. Then, 100 μL of each cell suspension (approximately 1 × 106 cells) was transferred into an FCM tube. Subsequently, Annexin V-FITC/PI, FITC-labeled anti-CD3 antibody, PE-labeled anti-CD29 antibody (eBioscience, San Diego, CA, USA), PE-labeled anti-CD90 antibody, PE-labeled anti-CD68 antibody, and FITC-labeled anti-CD105 antibody (Abcam, Cambridge, MA, USA) were added into each sample. The samples were then incubated for 20 min at room temperature (20 ± 5 °C) in the dark. The isotype control was likewise prepared using the isotype antibody (BD Pharmingen) at a volume equal to the total volume of the anti-CD antibodies. After the incubation, the samples were mixed with 2 mL stain buffer and centrifuged at 300× g for 5 min. The supernatant was discarded, and each cell pellet was resuspended in 0.5 mL stain buffer. FCM analysis (LSRII, BD Biosciences, San Jose, CA, USA) was promptly performed, and approximately 10,000 cells per sample were analyzed. The collected data were exported in the FCS3.0 format and analyzed using the Flowjo software (Tree Star Inc., Ashland, OR, USA). The proportion and intensity of the antigen expression were determined based on the isotype controls.
2.12. Histological Analysis
The entire knee joint or femoral condyle with complete AC was harvested, fixed in 4% paraformaldehyde (Sigma-Aldrich, St. Louis, MO, USA) for 24 h, and then decalcified using the EDTA (Servicebio Tech., Wuhan, China) method for 28 d with daily replacement of the decalcifying solution. After dehydration by an ethanol gradient, vitrification by xylene, and paraffin embedding, the trimmed wax block was cut into 4 μm-thick sections using a microtome (RM2016, Leica, Wetzlar, Germany). The tissue sections were then dewaxed and rehydrated using conventional procedures for subsequent use.
The entire joints from the control and model groups were stained with HE (Servicebio Tech., Wuhan, China). The morphological characteristics of the tissues were observed under a microscope to validate the successful construction of the OA model. AC morphology and collagen maturity were assessed via Masson’s staining (Sigma-Aldrich, St. Louis, MO, USA) to evaluate the OA progression after the EV-150 treatment.
2.13. Toluidine Blue Staining of Chondrocytes
Chondrocyte suspension was collected, and 3.5 × 104 cells were transferred into each well of a 24-well plate containing sterile coverslips. After overnight standing, the culture supernatant was removed, and the cells were incubated in 4% paraformaldehyde for 20 min at room temperature. Afterwards, the cells were washed with PBS and stained with toluidine blue (Solarbio, Beijing, China) for 30 min. Finally, they were rinsed with anhydrous ethanol and observed and photographed under a microscope.
2.14. IHC/IF Analysis
2.14.1. IHC Analysis of SM
The SM tissue required for separated FLSs was labeled Vimentin to determine the purity of FLSs in SM. After the SM sections, antigen repair was undergone and then incubation with the anti-Vimentin primary antibody (Abcam, Cambridge, MA, USA), and goat anti-rabbit HRP secondary antibody (Abcam, Cambridge, MA, USA) was introduced for the final detection. The nuclei were counterstained using hematoxylin (Servicebio Tech., Wuhan, China) and sealed. The positive FLS expression was observed using an inverted fluorescence microscope.
2.14.2. IF Analysis of FLSs and Chondrocytes
FLSs or chondrocytes grown on coverslips were fixed in 4% paraformaldehyde, permeabilized and blocked using conventional procedures, and then incubated with the anti-Vimentin or anti-COLII primary antibody (Abcam, Cambridge, MA, USA). Goat anti-rabbit IgG-555 fluorescent secondary antibody (Invitrogen, Carlsbad, CA, USA) was employed for the detection, and the samples were mounted with a mounting agent containing DAPI (Burlingame, CA, USA) to stain the nuclei.
2.14.3. IF Analysis of AC
To evaluate the therapeutic potential of EVs-150 in OA, IF colocalization analysis was performed to observe the extent of the AC injury and repair and the changes in the related signaling pathways. After antigen retrieval, tissue sections were incubated with the anti-COLII, anti-ACAN, anti–NF-κB (all from Abcam, Cambridge, MA, USA), and anti-Trim14 (Novus Biologicals, Littleton, CO, USA) antibodies as the primary antibodies and then with the Cy3- or Alexa fluor-488-labeled secondary antibody (Abcam, Cambridge, MA, USA) for fluorescent detection. The cell nucleus was stained with DAPI. The tissue sections were immediately examined and photographed under an inverted fluorescent microscope (IX-73, Olympus, Tokyo, Japan) to determine the fluorescence intensity and localization.
2.15. WB Analysis
The cells or tissues were lysed, and the total protein concentration was measured using the BCA assay. Equal quantities of proteins were resolved using SDS-PAGE and then transferred onto a PVDF membrane (Merck Millipore, Billerica, MA, USA). Next, the membrane was blocked at 37 °C for 2 h and then incubated with the primary antibody (anti-COLII, anti-ACAN, anti-NF-κB, anti-IFN-β, anti-GAPDH [all from Abcam, Cambridge, MA, USA], or anti-Trim14 [Novus Biologicals] antibody) at 4 °C overnight. The membrane was washed three times with PBST (PBS with Tween 20, Solarbio) and then incubated with the horseradish-peroxidase-labeled anti-mouse or anti-rabbit IgG secondary antibody (Jackson Immunoresearch, West Grove, PA, USA) (diluted 1: 10,000 with PBST) at 37 °C for 2 h. Finally, the membrane was washed five times with PBST and developed using chemiluminescence (BeyoECL Star [Beyotime, Shanghai, China]). The results were analyzed using the ImageJ software (NIH, Bethesda, MA, USA).
2.16. qPCR
Total RNA was extracted, and cDNA was synthesized using the RNeasy Mini Kit (QIAGEN, Duesseldorf, Germany), PrimeScript RT Master Mix, PrimeScript II 1st Strand cDNA Synthesis Kit (TaKaRa, Tokyo, Japan), and Power SYBR Green PCR Master Mix (Thermo Fisher Scientific, Waltham, MA, USA) according to the instructions of the manufacturers. The relative expression levels of miR-150-3p, COLII, ACAN, Trim14, NF-κB, and IFN-β were measured via qPCR using the ΔΔCt method. For normalization, U6 and cel-miR39 were used as the internal and external small RNA controls (for miR-150-3p), and GADPH was used as the internal control for the mRNAs. The results were normalized, and the specificity of the amplification was determined based on the presence of a single peak in the melting curve. The sequences of the primers (Sangon Biotech, Shanghai, China) used were as follows: miR-150-3p, stem-loop 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCCCCC-3′, forward 5′-GCGCCTGGTACAGGCCT-3′; universal downstream primer for stem-loop method, 5′-GTGCAGGGTCCGAGGT-3′; U6, reverse transcription 5′-CGCTTCACGAATTTGCGTGTCAT-3′, forward 5′-GCTTCGGCAGCACATATACTAAAAT-3′, reverse 5′-CGCTTCACGAATTTGCGTGTCAT-3′; cel-miR-39, stem-loop 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCAAGCT-3′, forward 5′-GGCCTCACCGGGTGTAAATCAG-3′; COLII, forward 5′-ACTTAACATCCAAGGCCGCT-3′, reverse 5′-ACAATATTTGCCTCAGTTTGTGC-3′; ACAN, forward 5′-AGAGGCAGAGGGACTTTCGGT-3′, reverse 5′-AGAGGCAGAGGGACTTTCGGT-3′; Trim14, forward 5′-GTGAATACTTACAGTGCCTTGC-3′, reverse 5′-GACCCAGACCAGAACCCT-3′; NF-κB, forward 5′-GCACGGATGACAGAGGCGTGTATAAGG-3′, reverse 5′-GGCGGATGATCTCCTTCTCTCTGTCTG-3′; IFN-β, forward 5′-ATTGCGTTCCTGCTGTGC-3′, reverse 5′-GTCCGAATGCTAGTGCTTTGTC-3′; GAPDH, forward 5′-AGACAGCCGCATCTTCTTGT-3′, reverse 5′-CTTGCCGTGGGTAGAGTCAT-3′.
2.17. Enzyme-Linked Immunosorbent Assay
The IL-1β, IL-6, and TNF-α levels in cell-culture media, sera, and synovial fluid were measured using ELISA according to the instructions of the manufacturer of the ELISA kit (Abcam, Cambridge, MA, USA).
2.18. Statistical Analysis
All the data are expressed as mean ± SEM and statistically processed using the SPSS 22.0 software (SPSS, Inc., Chicago, IL, USA) and GraphPad Prism 9.0 software (GraphPad Software, Inc., La Jolla, CA, USA). Comparison between the two groups and among more groups was performed using the Student’s t-test and one-way ANOVA, respectively. p < 0.05 was considered to indicate statistical significance.
4. Discussions
A healthy SM is critical for maintaining joint homeostasis and cartilage integrity [
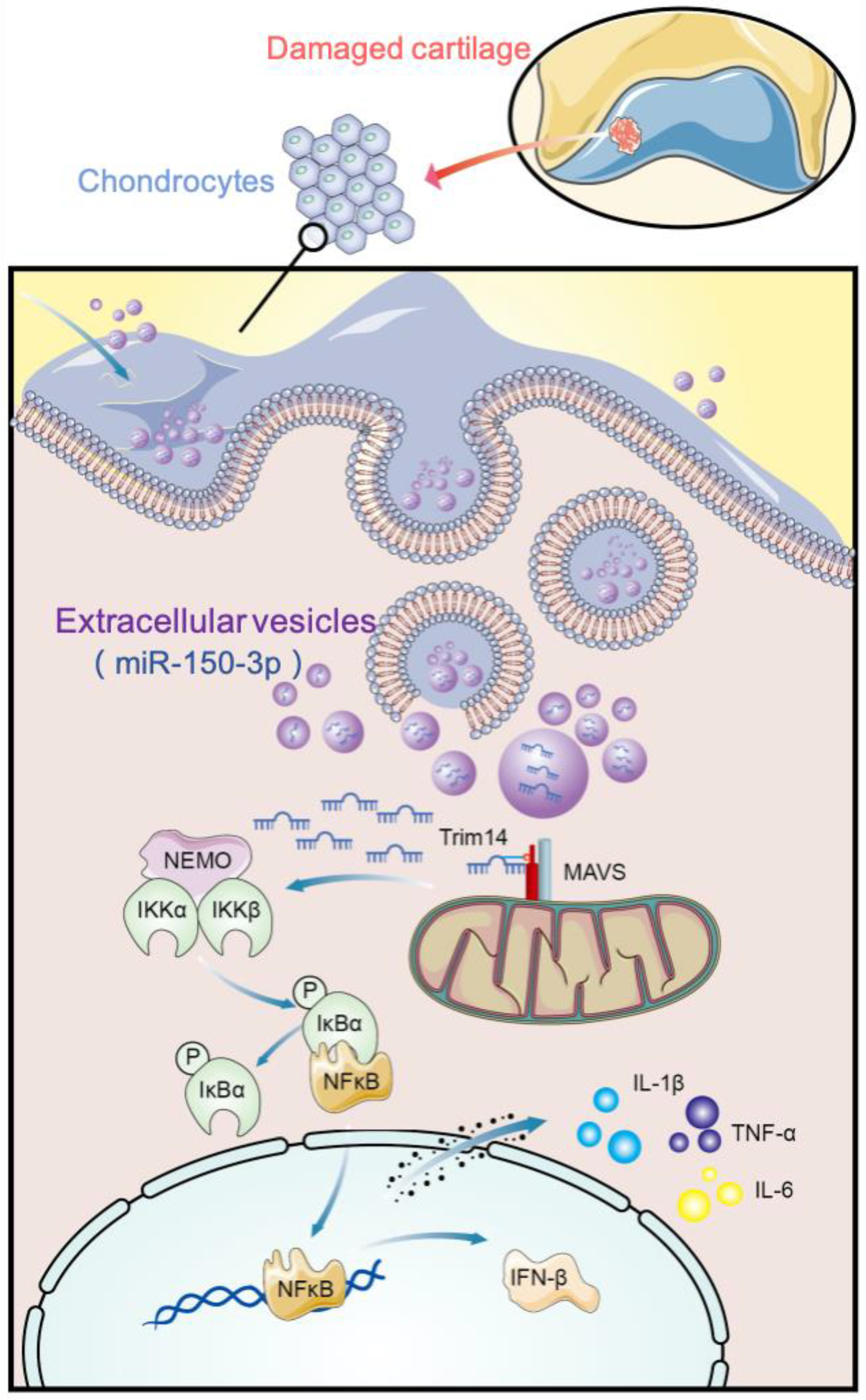
37]. However, the details of the interactions among the tissues in the joints and their specific contribution to the maintenance of joint homeostasis are still unclear. In this study, we demonstrated that H-FLS–EVs could act as vectors for cell-to-cell communication and participate in the maintenance of joint homeostasis through the innate immune response by delivering miR-150-3p to the chondrocytes. Thus, we preliminarily proved the importance of “early intervention”. These results show the therapeutic potentials of H-FLS–EVs in protecting joints and delaying the pathogenesis in OA through miRNA-mediated gene regulation.
As most previous OA studies have focused only on the impact of aberrant joint biomechanics on OA pathology [
38], other mechanisms promoting cartilage injury and progressive OA have been neglected. The joint should be viewed as a complex structure to be comprehensively analyzed, a structure wherein the tissue components all take concerted actions to maintain the homeostasis of the internal environment [
39]. Hence, in this study, we proposed the concept of “joint homeostasis”. The inflammatory state of the culture medium in direct contact with the two joint-derived cell types in the in vitro model and the synovial fluid in the in vivo model was used as the reference evidence to assess joint homeostasis. Considering the anatomical adjacency and functional interdependence of the AC and SM, alongside the importance of synovial pathology in AC degradation and OA progression, as was demonstrated in our previous study [
34], the SM, AC, and the liquid environment between the two were selected as the main subjects to study the mechanism and effect of FLS–EVs on chondrocytes.
miRNAs are regulatory molecules packaged into EVs and delivered from secretory cells to recipient cells to exert functions [
23,
40,
41]. Through bioinformatic analysis, miR-150-3p was depicted at a high level in EVs in the serum of healthy rats. However, it was significantly lower in OA rats, indicating that miR-150-3p could be an indispensable miRNA in joint homeostasis and cartilage protection.
We hypothesized that FLS–EVs could influence OA progression by acting as extracellular factors that regulate joint homeostasis and AC health. Hence, to further and better simulate the interaction system between the SM and AC and to determine the substance delivery and regulatory relationship between the two, we constructed an in vitro cell model, which mimics the joint cavity and thereby simulates the paracrine mechanism of substances released by FLSs and transported to chondrocytes via the microenvironment. This model also acts as a microscopic circulatory system that simulates the long-distance paracrine mechanism involving chondrocytes as the signaling cells secreting substances that are circulated through the blood to the joints [
42].
The past decade has witnessed a rapid increase in the understanding of the sources, characteristics, functions, and pathways of EVs. So far, the studies believe that almost all mammalian cell types, including fibroblasts and dendritic cell lines, MSCs, and dendritic cells, are EV secretory parental cells. MSCs from bone marrow, umbilical cord, and other tissues are the most commonly used EV secretory cells due to their differentiation potential, secretion, immune regulation, and therapeutic functions. They have been widely reported in treating a variety of diseases [
43,
44]. In recent years, the therapeutic potential of EVs obtained from MSCs in OA has also received increasing attention. Researchers have observed that it can protect OA joints from injury by promoting cartilage repair, suppressing synovial inflammation, and mediating subchondral bone remodeling [
28].
Unlike previous studies utilizing mesenchymal stem cells as the primary secretory cells [
45,
46,
47], our study showed that FLSs, the most common cell type in the SM with OA, could also secrete EVs. It is primarily based on the importance of FLSs in maintaining internal joint homeostasis in OA. In the closed joint cavity, chondrocytes depend on various bioactive substances secreted into the synovial fluid using synoviocytes to maintain joint health or alleviate, induce, or exacerbate joint pathology [
48,
49,
50]. FLSs, the primary cell type that constitutes the inner membrane structure of the SM, no longer act as innocent bystanders in the progression of joint disease but actively influence joint inflammation and destruction [
51]. FLSs, as an initiation factor, and the synovial fluid, as the mediator generated by them, regulate the transportation of substances among cells and facilitate the primary conditions for the interactions among the various tissues in the environment [
50,
52]. FLSs and FLS–EVs are closely associated with joint homeostasis and cartilage health, but they have rarely been reported. FLS–EVs are a new attempt to treat OA based on joint anatomy and OA pathogenesis, which is different from previous studies [
45,
46,
47]. In the field of OA, the analysis of miRNAs in FLS–EVs can better understand the pathogenesis of OA and develop new treatment strategies suitable for patients [
53]. This study could provide some new ideas and evidence for the further application of FLS–EVs in OA research. However, their transformation and production will be long-term, and in-depth studies are needed to validate the efficacy and optimize the process [
54].
These outstanding issues and challenges in developing EV-related technologies have been juxtaposed with the promising development of the material delivery functions of EVs in treating and diagnosing diseases. Based on some studies, not all EVs can carry miRNAs to participate in cell-to-cell communication and thus play a part in biological processes. Moreover, the stoichiometry of miRNAs in EVs and whether EVs are more likely to function through protein or RNA are inconsistent across different studies [
55,
56,
57]. These studies reflect the main reasons for limiting the application of EVs, due to the high heterogeneity of cell types, experimental methods, dose regimens, and even experimental operations used in many studies, leading to the inconsistency of the source, quality, and related technologies of EVs. The latest guidance requirements for EVs (MISEV2018) attempt to solve and unify these problems [
58]. However, due to the reasons mentioned above, the recognition of EV function and the effect of EVs have not reached a consensus until now. In addition, it is difficult to monitor and maintain cell viability, potency, and EV transformation during the manufacturing process of EV preparations. Achieving a standardized method for producing and treating a wide variety of EV preparations is impossible.
In the future, newer technologies and scientific methods may apply EVs in OA research. Several studies have confirmed the biological function of EV-mediated miRNAs, the communication effect, and the advantage between cells [
19,
23]. For instance, Zhou et al. [
53] believed that although synovial miRNAs are rapidly degraded, EVs can stabilize these molecules. Thus, EVs enhance their ability to alter the proliferation, differentiation, survival, and inflammatory activity of other cells.
However, previous studies have focused on investigating various pathological states [
59,
60], and a few have examined the possible roles of bioactive substances obtained from healthy synoviocytes. We proposed the idea of using H-FLS–EVs as a therapeutic measure in OA. However, the study on EV-mediated miRNAs is currently in its initial stage, and whether they can be used for joint protection in OA remains unclear and needs further investigation.
Our findings have clarified the intercellular transport mechanism of miRNAs and demonstrated that EVs can carry miR-150-3p from FLSs to chondrocytes. Healthy FLSs, like mesenchymal stem cells [
61], can also play a beneficial role in OA by releasing EVs, and this effect is sufficient to repair the joint and AC significantly. Our study repeatedly emphasized the importance of a good SM condition in maintaining joint health. On this basis, we determined that the EVs-150-mediated intervention of OA at an early stage is more effective than later. Thus, an early diagnosis is crucial to apply this intervention as soon as possible.
EV-mediated miRNAs have sufficient sensitivity and specificity and the potential to serve as simple and accurate biomarkers of OA [
62,
63]. The changes in the level of EVs-150 are critical for the expression levels of COLII and ACAN and the activities of related pathways, as well as the levels of downstream inflammatory mediators, indicating that the OA treatment strategy is based on EVs-150.
There are few reports on the biological functions of the miR-150-3p-mediated regulation of Trim14 expression. Our study showed that Trim14 is a direct target gene of miR-150-3p, and EVs-150 can influence OA by modulating the innate immune response by regulating Trim14 expression and the NF-κB signal transduction pathway. The NF-κB signaling plays a critical role in the activating of the innate immune response in OA. Many key effectors during OA development and progression are under the control of NF-κB. Many pathways involved in NF-κB activation have been well studied [
64,
65], but research on the regulation mechanism of Trim14–NF-κB in OA has rarely been reported. Previous studies have reported that Trim14 is a mitochondrial adaptor located on the outer mitochondrial membrane and is involved in the innate immune response to infections and tumors [
66,
67]. Our study found that EVs-150 can target Trim14 and regulate its expression to affect sterile inflammation, such as OA. We observed that miR-150-3p downregulates Trim14 and thereby reduces the activities of the related NF-κB pathways and that H-FLS–EVs can be used to inhibit the Trim14/NF-κB/IFNβ axis for the treatment of OA.
The rats used in our experiments are highly anatomically suitable for studying various human diseases, including OA and other diseases involving joint injury [
68,
69]. Rat genes are highly homologous to human genes and are commonly used for OA research [
70]. We selected the rat model of OA based on ALCT, a modeling method in common use for OA, which can change the stability of joints and simulate joint wear and degeneration in OA [
71].
The crosstalk among cells is essential during OA pathogenesis [
72,
73,
74], and the crosstalk of the Trim14/NF-κB/IFN-β axis and other pathways may also occur in OA. We did not exclude the possibility of substance transportation among other structures in joints. Whether EV-mediated miR-150-3p in the circulation can stimulate the pathogenesis of OA at distal sites, and how it can be used as a diagnostic marker and therapeutic agent in OA, needs in-depth analyses. FLS–EVs from healthy SM can regulate the Trim14/NF-κB/IFN-β axis of the innate immune response by delivering miR-150-3p to chondrocytes to maintain joint homeostasis, repair AC injury, and delay OA progression. The earlier this intervention is provided, the more effective it is in preventing joint degeneration. These important findings may serve as a basis for further studies on the mechanisms of OA and provide an attractive source of biomarker candidates for monitoring joint health during OA. They may also lead to the development of novel approaches for the treatment of OA.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}












