
Human Adipose Mesenchymal Stromal/Stem Cells Improve Fat Transplantation Performance

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Expansion of Human AD-MSCs
2.2. Fluorescence-Activated Cell Sorting Analyses (FACS)
2.3. Differentiation Assays
2.4. Prostaglandin E2 (PGE2) Evaluation in AD-MSCs
2.5. Retroviral Transduction of Human AD-MSCs to Express GFP
2.6. Microbiology Test
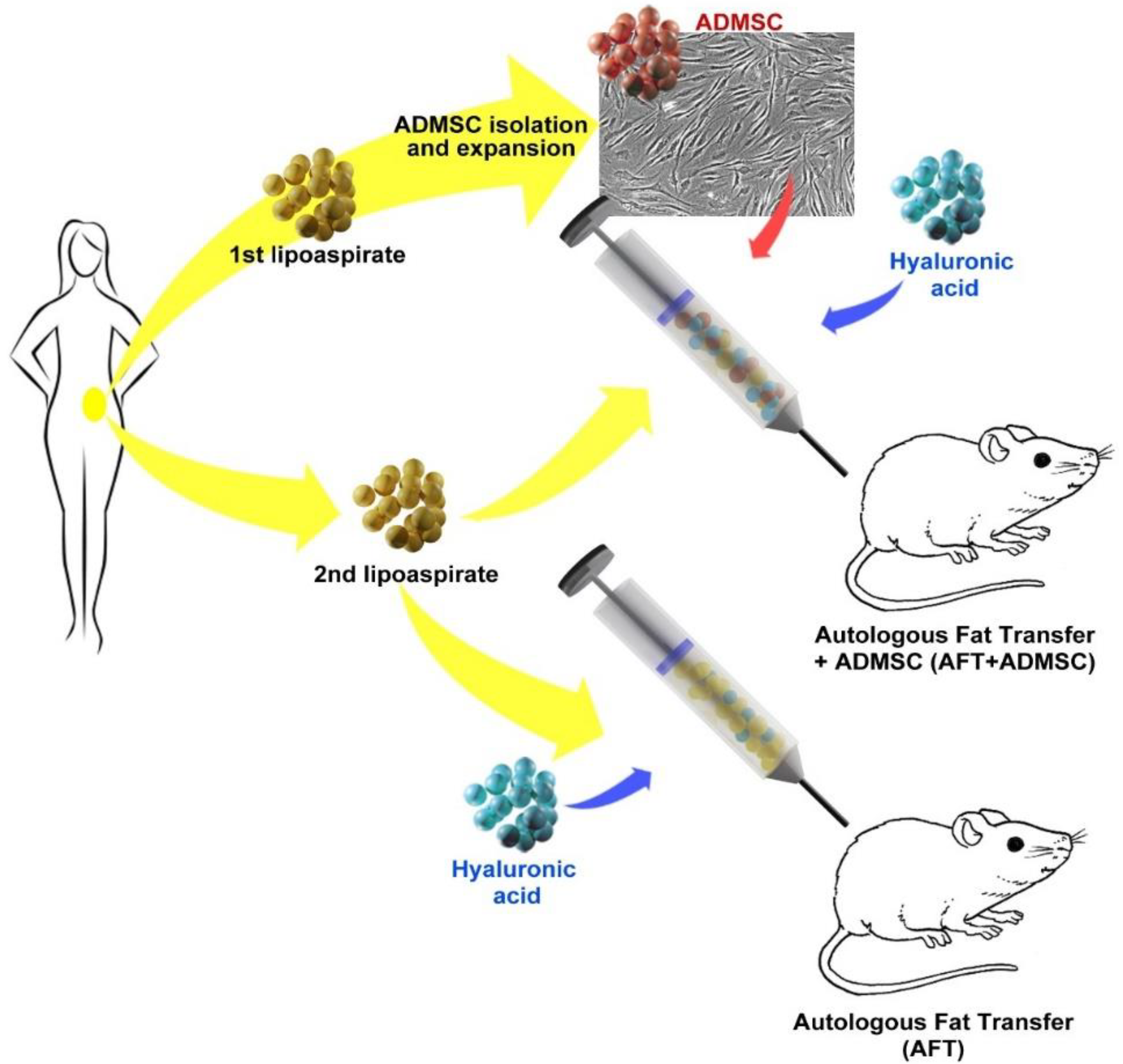
2.7. Human AD-MSC-Assisted Autologous Fat Transplantation in a Xenogeneic Model
2.8. Hematology and Biochemistry
2.9. Tissue Handling and Histology
2.10. Statistical Analysis
3. Results
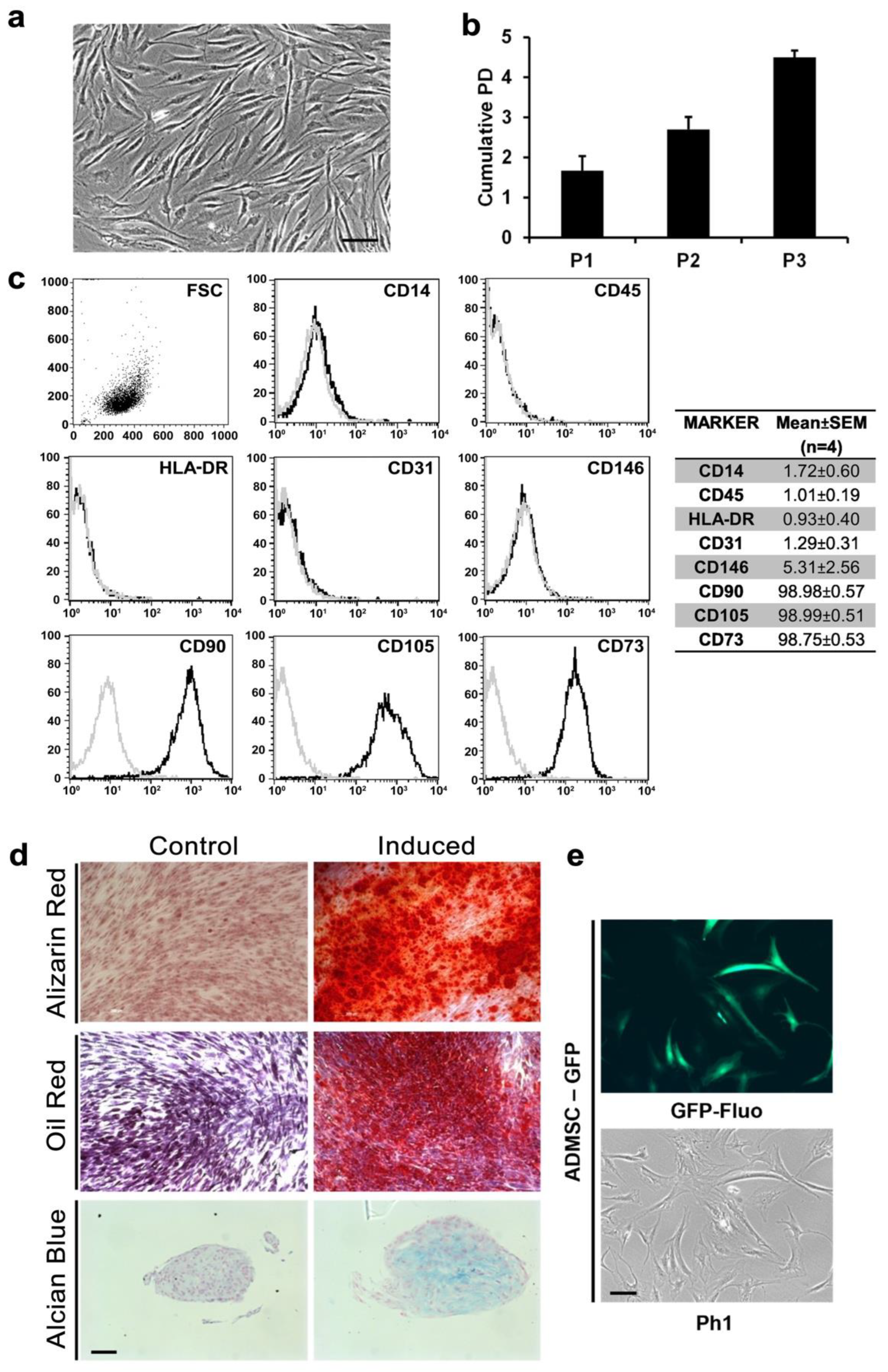
3.1. AD-MSCs Could Be Isolated and Genetically Modified from Small Amounts of Adipose Tissue
3.2. Xenotransplantation Was Well Tolerated by NOD/SCID Mice
3.3. AD-MSC-GFP Differentiated into Adipocytes within 60 Days after Transplantation
3.4. Proangiogenic and Pro-Inflammatory Characterization of AFT-AD-MSC-GFP
3.5. Wild-Type AD-MSCs Preserved Their Proangiogenic and Anti-Inflammatory Properties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krastev, T.K.; Schop, S.J.; Hommes, J.; Piatkowski, A.; van der Hulst, R.R.W.J. Autologous fat transfer to treat fibrosis and scar-related conditions: A systematic review and meta-analysis. J. Plast. Reconstr. Aesthetic Surg. 2020, 73, 2033–2048. [Google Scholar] [CrossRef] [PubMed]
- Raj, S.; Abu-Ghname, A.; Davis, M.J.; Izaddoost, S.A.; Winocour, S.J. Safety and Regulation of Fat Grafting. In Seminars in Plastic Surgery; Thieme Medical Publishers: Leipzig, Germany, 2020; Volume 34, pp. 59–64. [Google Scholar]
- Bauer-Kreisel, P.; Goepferich, A.; Blunk, T. Cell-Delivery Therapeutics for Adipose Tissue Regeneration. Adv. Drug Deliv. Rev. 2010, 62, 798–813. [Google Scholar] [CrossRef]
- Kim, H.Y.; Jung, B.K.; Lew, D.H.; Lee, D.W. Autologous Fat Graft in the Reconstructed Breast: Fat Absorption Rate and Safety based on Sonographic Identification. Arch. Plast. Surg. 2014, 41, 740–747. [Google Scholar] [CrossRef]
- Smith, P.; Adams, W.P.; Lipschitz, A.H.; Chau, B.; Sorokin, E.; Rohrich, R.J.; Brown, S.A. Autologous Human Fat Grafting: Effect of Harvesting and Preparation Techniques on Adipocyte Graft Survival. Plast. Reconstr. Surg. 2006, 117, 1836–1844. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.I.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef]
- Kesten, S.; Fraser, J.K. Autologous Adipose Derived Regenerative Cells: A Platform for Therapeutic Applications. Surg. Technol. Int. 2016, 29, 38–44. [Google Scholar]
- Yoshimura, K.; Aoi, N.; Suga, H.; Inoue, K.; Eto, H.; Sato, K.; Kurita, M.; Harii, K.; Hirohi, T. Ectopic Fibrogenesis Induced by Transplantation of Adipose-Derived Progenitor Cell Suspension Immediately after Lipoinjection. Transplantation 2008, 85, 1868–1869. [Google Scholar] [CrossRef]
- Matsumoto, D.; Sato, K.; Gonda, K.; Takaki, Y.; Shigeura, T.; Sato, T.; Aiba-Kojima, E.; Iizuka, F.; Inoue, K.; Suga, H.; et al. Cell-Assisted Lipotransfer: Supportive Use of Human Adipose-Derived Cells for Soft Tissue Augmentation with Lipoinjection. Tissue Eng. 2006, 12, 3375–3382. [Google Scholar] [CrossRef]
- Toyserkani, N.M.; Quaade, M.L.; Sørensen, J.A. Cell-Assisted Lipotransfer: A Systematic Review of Its Efficacy. Aesthetic Plast. Surg. 2016, 40, 309–318. [Google Scholar] [CrossRef]
- Prantl, L.; Brix, E.; Kempa, S.; Felthaus, O.; Eigenberger, A.; Brébant, V.; Anker, A.; Strauss, C. Facial Rejuvenation with Concentrated Lipograft—A 12 Month Follow-Up Study. Cells 2021, 10, 594. [Google Scholar] [CrossRef] [PubMed]
- Piccinno, M.S.; Veronesi, E.; Loschi, P.; Pignatti, M.; Murgia, A.; Grisendi, G.; Castelli, I.; Bernabei, D.; Candini, O.; Conte, P.; et al. Adipose stromal/stem cells assist fat transplantation reducing necrosis and increasing graft performance. Apoptosis 2013, 18, 1274–1289. [Google Scholar] [CrossRef] [Green Version]
- Coleman, S.R. Structural Fat Grafting: More Than a Permanent Filler. Plast. Reconstr. Surg. 2006, 118, 108S–120S. [Google Scholar] [CrossRef]
- Grisendi, G.; Bussolari, R.; Cafarelli, L.; Petak, I.; Rasini, V.; Veronesi, E.; De Santis, G.; Spano, C.; Tagliazzucchi, M.; Barti-Juhasz, H.; et al. Adipose-Derived Mesenchymal Stem Cells as Stable Source of Tumor Necrosis Factor–Related Apoptosis-Inducing Ligand Delivery for Cancer Therapy. Cancer Res. 2010, 70, 3718–3729. [Google Scholar] [CrossRef]
- Grisendi, G.; Annerén, C.; Cafarelli, L.; Sternieri, R.; Veronesi, E.; Cervo, G.L.; Luminari, S.; Maur, M.; Frassoldati, A.; Palazzi, G.; et al. GMP-manufactured density gradient media for optimized mesenchymal stromal/stem cell isolation and expansion. Cytotherapy 2010, 12, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Pajarinen, J.; Nabeshima, A.; Lu, L.; Nathan, K.; Jämsen, E.; Yao, Z.; Goodman, S.B. Preconditioning of murine mesenchymal stem cells synergistically enhanced immunomodulation and osteogenesis. Stem Cell Res. Ther. 2017, 8, 277. [Google Scholar] [CrossRef]
- Zhu, M.; Zhou, Z.; Chen, Y.; Schreiber, R.; Ransom, J.T.; Fraser, J.K.; Hedrick, M.H.; Pinkernell, K.; Kuo, H.-C. Supplementation of Fat Grafts with Adipose-Derived Regenerative Cells Improves Long-Term Graft Retention. Ann. Plast. Surg. 2010, 64, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Rasini, V.; Bussolari, R.; Chen, X.; Hofmann, T.J.; Spano, C.; Bernabei, D.; Veronesi, E.; Bertoni, F.; Paolucci, P.; et al. Restoration and reversible expansion of the osteoblastic hematopoietic stem cell niche after marrow radioablation. Blood 2009, 114, 2333–2343. [Google Scholar] [CrossRef]
- Debels, H.; Galea, L.; Han, X.-L.; Palmer, J.; Van Rooijen, N.; Morrison, W.; Abberton, K. Macrophages Play a Key Role in Angiogenesis and Adipogenesis in a Mouse Tissue Engineering Model. Tissue Eng. Part A 2013, 19, 2615–2625. [Google Scholar] [CrossRef]
- Charles River Research Model, Technical Shit about NOD SCID Mouse Hematology. Available online: https://larc.ucsf.edu/sites/larc.ucsf.edu/files/wysiwyg/RM-catalog-2019-US-full-version.pdf (accessed on 19 June 2022).
- Ansari, A.M.; Ahmed, A.K.; Matsangos, A.E.; Lay, F.; Born, L.J.; Marti, G.; Harmon, J.W.; Sun, Z. Cellular GFP Toxicity and Immunogenicity: Potential Confounders in in Vivo Cell Tracking Experiments. Stem Cell Rev. Rep. 2016, 12, 553–559. [Google Scholar] [CrossRef]
- Moustaki, M.; Papadopoulos, O.; Verikokos, C.; Karypidis, D.; Masud, D.; Kostakis, A.; Papastefanaki, F.; Roubelakis, M.G.; Perrea, D. Application of adipose-derived stromal cells in fat grafting: Basic science and literature review. Exp. Ther. Med. 2017, 14, 2415–2423. [Google Scholar] [CrossRef]
- Fisher, C.; Grahovac, T.L.; Schafer, M.E.; Shippert, R.D.; Marra, K.; Rubin, J.P. Comparison of Harvest and Processing Techniques for Fat Grafting and Adipose Stem Cell Isolation. Plast. Reconstr. Surg. 2013, 132, 351–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielins, E.R.; Brett, E.A.; Longaker, M.T.; Wan, D.C. Autologous Fat Grafting: The Science Behind the Surgery. Aesthetic Surg. J. 2016, 36, 488–496. [Google Scholar] [CrossRef]
- Mantha, S.; Pillai, S.; Khayambashi, P.; Upadhyay, A.; Zhang, Y.; Tao, O.; Pham, H.M.; Tran, S.D. Smart Hydrogels in Tissue Engineering and Regenerative Medicine. Materials 2019, 12, 3323. [Google Scholar] [CrossRef]
- Skelton, D.; Satake, N.; Kohn, D. The enhanced green fluorescent protein (eGFP) is minimally immunogenic in C57BL/6 mice. Gene Ther. 2001, 8, 1813–1814. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, H.J.; Song, Y.S. Tracking Transplanted Stem Cells Using Magnetic Resonance Imaging and the Nanoparticle Labeling Method in Urology. BioMed Res. Int. 2015, 2015, 231805. [Google Scholar] [CrossRef]
- Guo, Y.; Su, L.; Wu, J.; Zhang, N.; Zhang, X.; Zhang, G.; Li, T.; Wang, J.; Liu, C. Assessment of the green florescence protein labeling method for tracking implanted mesenchymal stem cells. Cytotechnology 2012, 64, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Wei, N.; Sun, Z.; Yu, J.; Jia, Y.; Zheng, P.; Tang, H.; Chen, J. Immunological Responses to Transgene-Modified Neural Stem Cells After Transplantation. Front. Immunol. 2021, 12, 697203. [Google Scholar] [CrossRef] [PubMed]
- Stillaert, F.; Findlay, M.; Palmer, J.; Idrizi, R.; Cheang, S.; Messina, A.; Abberton, K.; Morrison, W.; Thompson, E.W. Host Rather than Graft Origin of Matrigel-Induced Adipose Tissue in the Murine Tissue-Engineering Chamber. Tissue Eng. 2007, 13, 2291–2300. [Google Scholar] [CrossRef]
- Bowles, A.C.; Wise, R.M.; Gerstein, B.Y.; Thomas, R.C.; Ogelman, R.; Febbo, I.; Bunnell, B.A. Immunomodulatory Effects of Adipose Stromal Vascular Fraction Cells Promote Alternative Activation Macrophages to Repair Tissue Damage. Stem Cells 2017, 35, 2198–2207. [Google Scholar] [CrossRef]
- Lu, X.; Han, J.; Xu, X.-P.; Xu, J.; Liu, L.; Huang, Y.; Yang, Y.; Qiu, H. PGE2 Promotes the Migration of Mesenchymal Stem Cells through the Activation of FAK and ERK1/2 Pathway. Stem Cells Int. 2017, 2017, 8178643. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Daaka, Y. PGE2 promotes angiogenesis through EP4 and PKA Cγ pathway. Blood 2011, 118, 5355–5364. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.-C.; Kim, H.-S.; Shin, T.-H.; Kang, I.; Lee, J.Y.; Kim, J.-J.; Kang, H.K.; Seo, Y.; Lee, S.; Yu, K.-R.; et al. PGE2 maintains self-renewal of human adult stem cells via EP2-mediated autocrine signaling and its production is regulated by cell-to-cell contact. Sci. Rep. 2016, 6, 26298. [Google Scholar] [CrossRef]
- Fraser, J.K.; Wulur, I.; Alfonso, Z.; Hedrick, M.H. Fat Tissue: An Underappreciated Source of Stem Cells for Biotechnology. Trends Biotechnol. 2006, 24, 150–154. [Google Scholar] [CrossRef]
- Dong, Z.; Peng, Z.; Chang, Q.; Zhan, W.; Zeng, Z.; Zhang, S.; Lu, F. The Angiogenic and Adipogenic Modes of Adipose Tissue after Free Fat Grafting. Plast. Reconstr. Surg. 2015, 135, 556e–567e. [Google Scholar] [CrossRef] [PubMed]
- Kølle, S.-F.T.; Fischer-Nielsen, A.; Mathiasen, A.B.; Elberg, J.J.; Oliveri, R.S.; Glovinski, P.V.; Kastrup, J.; Kirchhoff, M.; Rasmussen, B.S.; Talman, M.-L.M.; et al. Enrichment of autologous fat grafts with ex-vivo expanded adipose tissue-derived stem cells for graft survival: A randomised placebo-controlled trial. Lancet 2013, 382, 1113–1120. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piccinno, M.S.; Petrachi, T.; Pignatti, M.; Murgia, A.; Grisendi, G.; Candini, O.; Resca, E.; Bergamini, V.; Ganzerli, F.; Portone, A.; et al. Human Adipose Mesenchymal Stromal/Stem Cells Improve Fat Transplantation Performance. Cells 2022, 11, 2799. https://doi.org/10.3390/cells11182799
Piccinno MS, Petrachi T, Pignatti M, Murgia A, Grisendi G, Candini O, Resca E, Bergamini V, Ganzerli F, Portone A, et al. Human Adipose Mesenchymal Stromal/Stem Cells Improve Fat Transplantation Performance. Cells. 2022; 11(18):2799. https://doi.org/10.3390/cells11182799
Chicago/Turabian StylePiccinno, Maria Serena, Tiziana Petrachi, Marco Pignatti, Alba Murgia, Giulia Grisendi, Olivia Candini, Elisa Resca, Valentina Bergamini, Francesco Ganzerli, Alberto Portone, and et al. 2022. "Human Adipose Mesenchymal Stromal/Stem Cells Improve Fat Transplantation Performance" Cells 11, no. 18: 2799. https://doi.org/10.3390/cells11182799
APA StylePiccinno, M. S., Petrachi, T., Pignatti, M., Murgia, A., Grisendi, G., Candini, O., Resca, E., Bergamini, V., Ganzerli, F., Portone, A., Mastrolia, I., Chiavelli, C., Castelli, I., Bernabei, D., Tagliazucchi, M., Bonetti, E., Lolli, F., De Santis, G., Dominici, M., & Veronesi, E. (2022). Human Adipose Mesenchymal Stromal/Stem Cells Improve Fat Transplantation Performance. Cells, 11(18), 2799. https://doi.org/10.3390/cells11182799