
Circulating Microparticles Are Differentially Increased in Lowlanders and Highlanders with High Altitude Induced Pulmonary Hypertension during the Cold Season

 , , , , , , , , ,
, , , , , , , , , 
Abstract
:1. Introduction
2. Materials and Methods
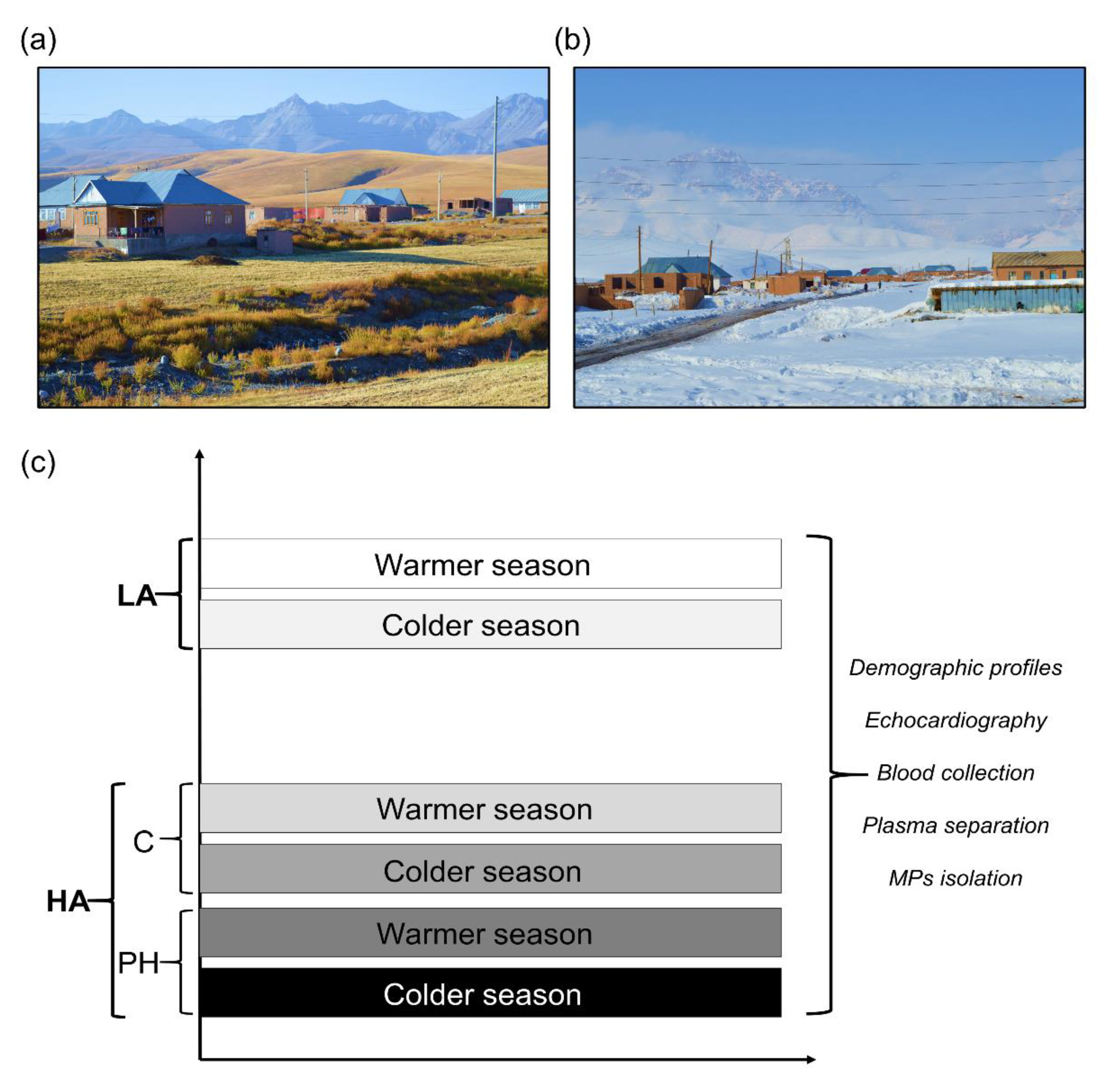
2.1. High Altitude Expedition, Experimental Design, and Human Subjects
2.2. Flow Cytometry Characterization and Quantification of Endothelial and Different Inflammatory Cells-Derived MPs
2.3. Effects of Isolated MPs on Proliferation and Apoptosis of Human Pulmonary Artery Smooth Muscle Cells (PASMCs)
2.4. The Contents of Different MicroRNAs (miRNAs) in Isolated MPs: Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.5. Effects of Isolated MPs on Human Pulmonary Artery Endothelial Cells (hPAECs): Tube Formation Assay
2.6. Effects of Isolated MPs on hPAECs: Western Blot
2.7. Statistical Analysis
3. Results
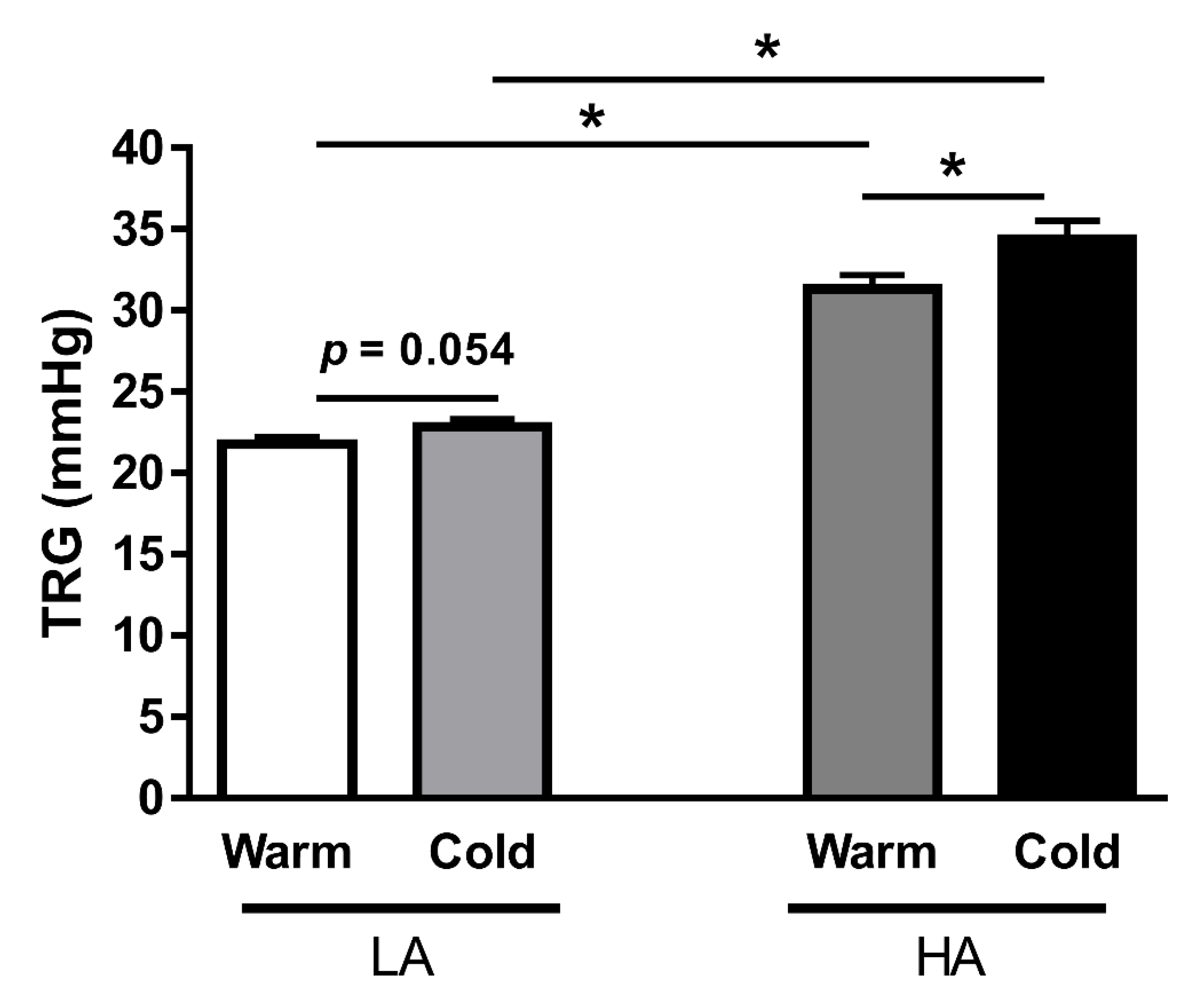
3.1. Long-Term Exposure to Cold Environment Enhances PAP in Kyrgyz Highlanders
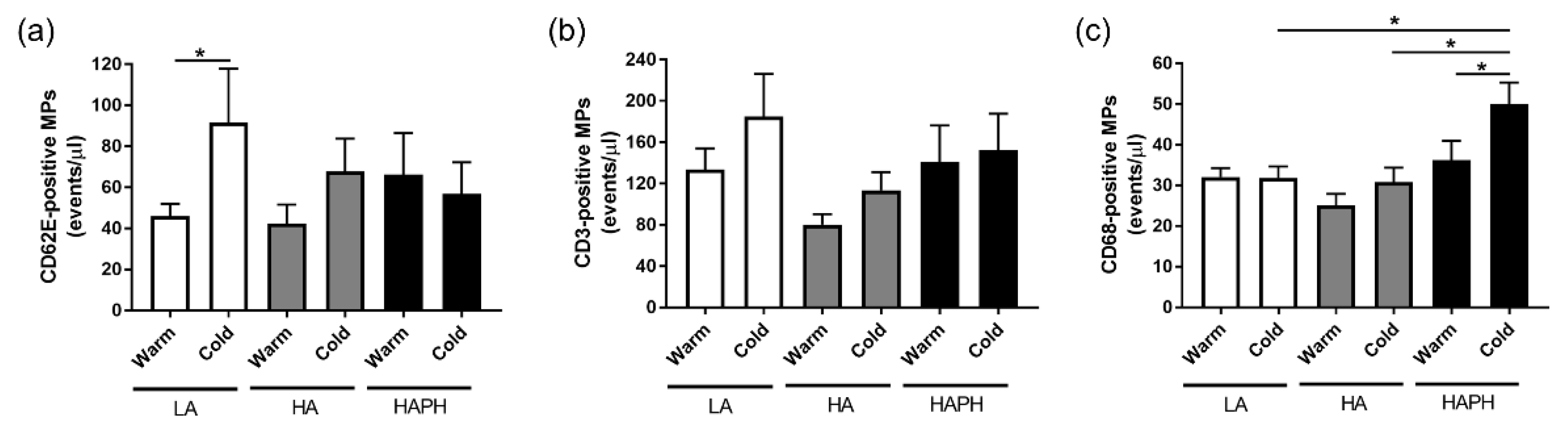
3.2. Effects of High Altitude and Cold on Circulating Levels of Different Endothelial and Inflammatory Cells-Derived MPs
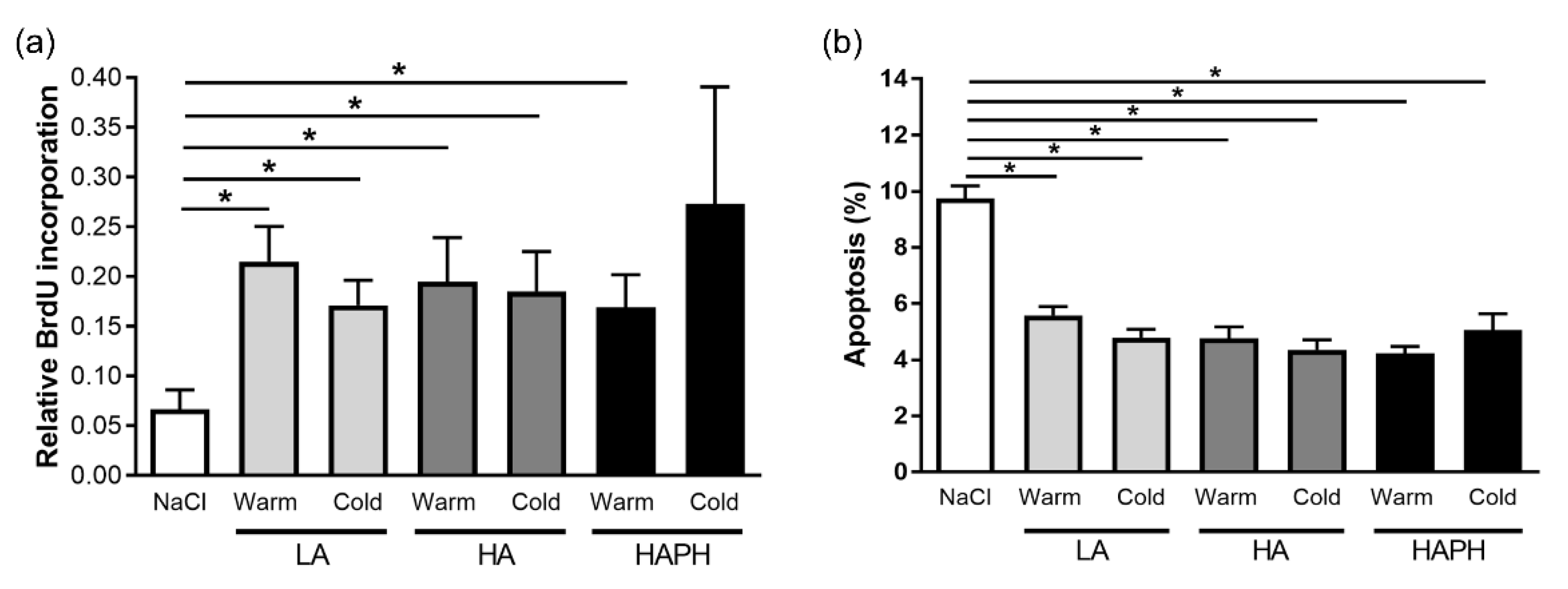
3.3. Effects of Isolated MPs on Human PASMCs
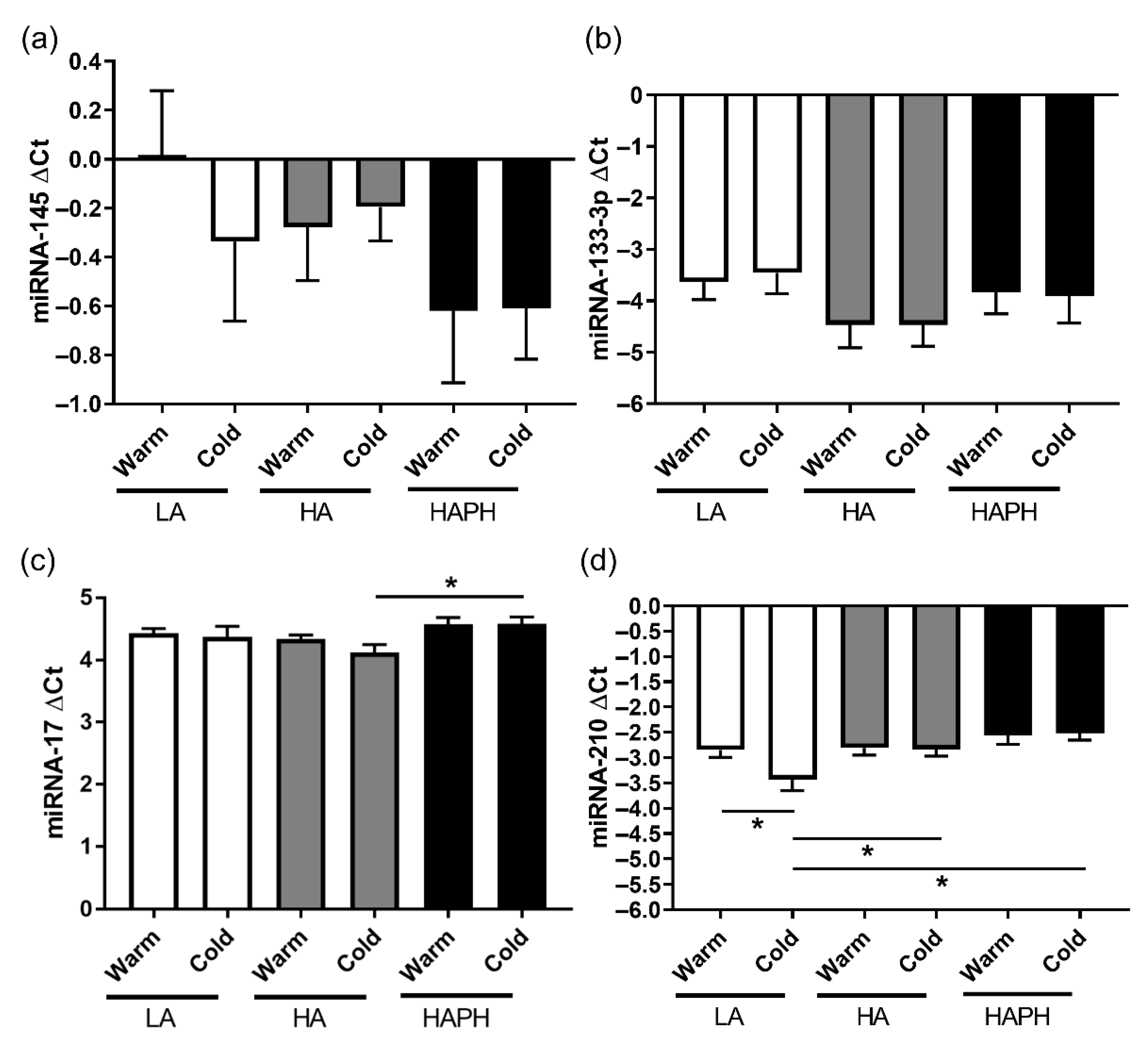
3.4. Effects of High Altitude and Cold on the Content of Different miRNAs in Isolated MPs
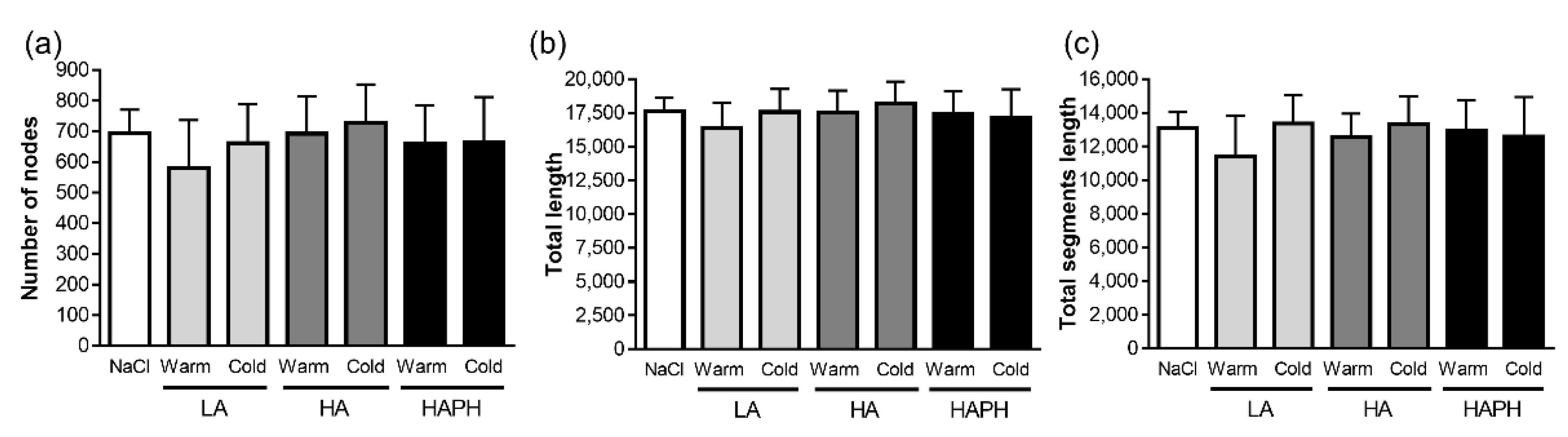
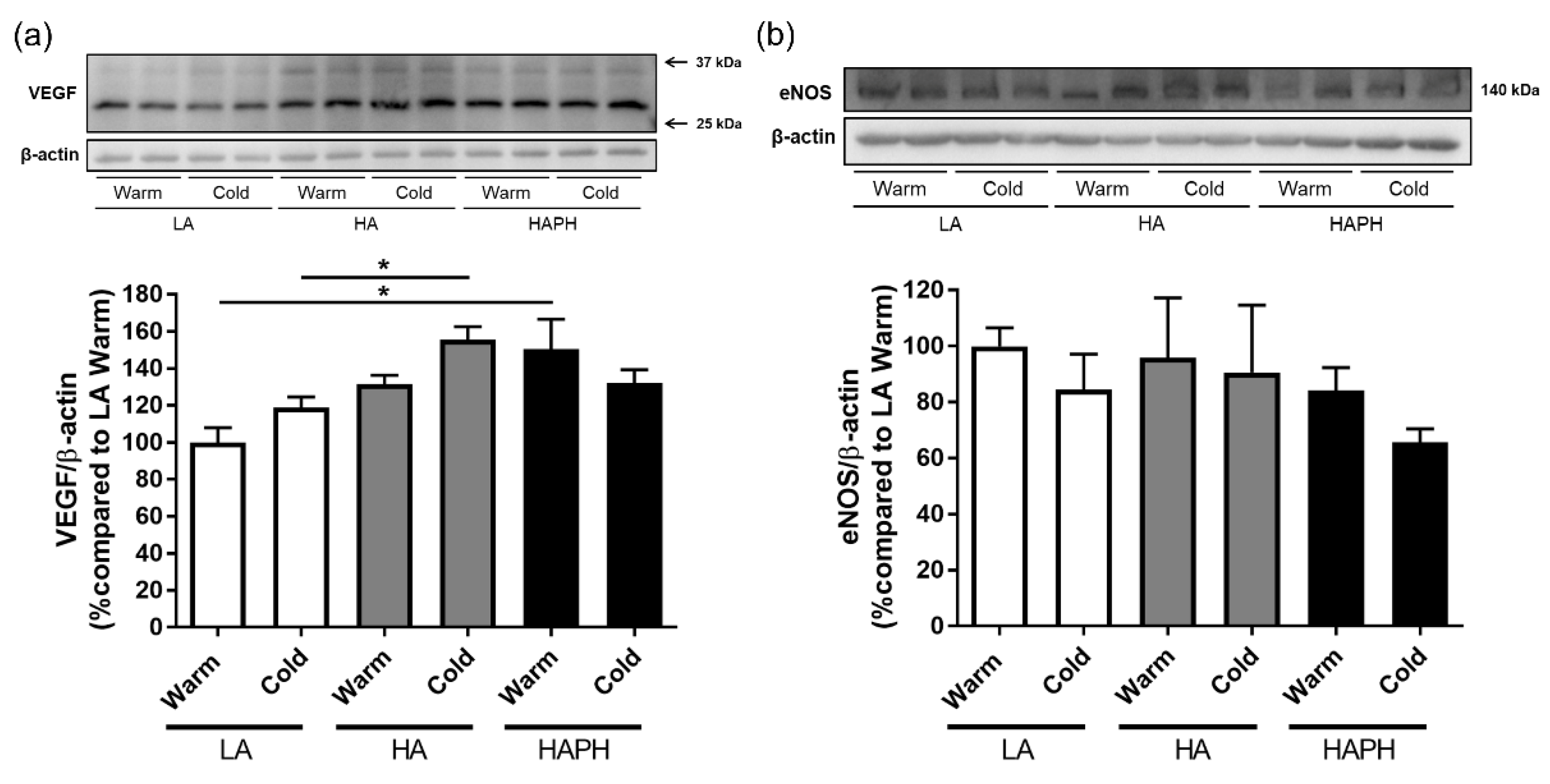
3.5. Effects of Isolated MPs on hPAECs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Penaloza, D.; Arias-Stella, J. The heart and pulmonary circulation at high altitudes: Healthy highlanders and chronic mountain sickness. Circulation 2007, 115, 1132–1146. [Google Scholar] [CrossRef]
- Tipton, M. A case for combined environmental stressor studies. Extrem Physiol. Med. 2012, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.; Mackey, D.R. Brisket disease. In Diseases of Feedlot Cattle, 3rd ed.; Lea and Febiger: Philadelphia, PA, USA, 1979; pp. 257–261. [Google Scholar]
- Will, D.H.; McMurtry, I.F.; Reeves, J.T.; Grover, R.F. Cold-induced pulmonary hypertension in cattle. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1978, 45, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Busch, M.A.; Tucker, A.; Robertshaw, D. Interaction between cold and altitude exposure on pulmonary circulation of cattle. J. Appl. Physiol. 1985, 58, 948–953. [Google Scholar] [CrossRef] [PubMed]
- Chauca, D.; Bligh, J. An additive effect of cold exposure and hypoxia on pulmonary artery pressure in sheep. Res. Vet. Sci. 1976, 21, 123–124. [Google Scholar] [CrossRef]
- Kashimura, O. Effects of acute exposure to cold on pulmonary arterial blood pressure in awake rats. Nihon Eiseigaku Zasshi 1993, 48, 859–863. [Google Scholar] [CrossRef]
- Kashimura, O. Mechanisms of cold-induced pulmonary hypertension in rats. Nihon Eiseigaku Zasshi 1995, 50, 616–621. [Google Scholar] [CrossRef]
- Watanabe, K.; Koizumi, T.; Ruan, Z.; Kubo, K.; Sakai, A.; Shibamoto, T. Reduced pulmonary vascular reactivity after cold exposure to acute hypoxia: A role of nitric oxide (NO). High Alt. Med. Biol. 2007, 8, 43–49. [Google Scholar] [CrossRef]
- Crosswhite, P.; Chen, K.; Sun, Z. AAV delivery of tumor necrosis factor-alpha short hairpin RNA attenuates cold-induced pulmonary hypertension and pulmonary arterial remodeling. Hypertension 2014, 64, 1141–1150. [Google Scholar] [CrossRef]
- Crosswhite, P.; Sun, Z. Inhibition of phosphodiesterase-1 attenuates cold-induced pulmonary hypertension. Hypertension 2013, 61, 585–592. [Google Scholar] [CrossRef] [Green Version]
- Li, J.C.; Pan, J.Q.; Huang, G.Q.; Tan, X.; Sun, W.D.; Liu, Y.J.; Wang, X.L. Expression of PDGF-beta receptor in broilers with pulmonary hypertension induced by cold temperature and its association with pulmonary vascular remodeling. Res. Vet. Sci. 2010, 88, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, A.; Seimetz, M.; Sydykov, A.; Pak, O.; Veit, F.; Luitel, H.; Weissmann, N.; Schermuly, R.T.; Kosanovic, D. Cold-induced pulmonary hypertension: A distinctive pathological entity of the pulmonary circulation? PVRI Chron. 2015, 2, 14–17. [Google Scholar]
- Sydykov, A.; Maripov, A.; Muratali Uulu, K.; Kushubakova, N.; Petrovic, A.; Vroom, C.; Cholponbaeva, M.; Duishobaev, M.; Satybaldyev, S.; Satieva, N.; et al. Pulmonary Vascular Pressure Response to Acute Cold Exposure in Kyrgyz Highlanders. High Alt. Med. Biol. 2019, 20, 375–382. [Google Scholar] [CrossRef]
- Mause, S.F.; Weber, C. Microparticles: Protagonists of a novel communication network for intercellular information exchange. Circ. Res. 2010, 107, 1047–1057. [Google Scholar] [CrossRef]
- Vince, R.V.; Chrismas, B.; Midgley, A.W.; McNaughton, L.R.; Madden, L.A. Hypoxia mediated release of endothelial microparticles and increased association of S100A12 with circulating neutrophils. Oxid. Med. Cell Longev. 2009, 2, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Lichtenauer, M.; Goebel, B.; Fritzenwanger, M.; Förster, M.; Betge, S.; Lauten, A.; Figulla, H.R.; Jung, C. Simulated temporary hypoxia triggers the release of CD31+/Annexin+ endothelial microparticles: A prospective pilot study in humans. Clin. Hemorheol. Microcirc. 2015, 61, 83–90. [Google Scholar] [CrossRef]
- Ayers, L.; Stoewhas, A.C.; Ferry, B.; Latshang, T.D.; Lo Cascio, C.M.; Sadler, R.; Stadelmann, K.; Tesler, N.; Huber, R.; Achermann, P.; et al. Circulating levels of cell-derived microparticles are reduced by mild hypobaric hypoxia: Data from a randomised controlled trial. Eur. J. Appl. Physiol. 2014, 114, 1067–1073. [Google Scholar] [CrossRef]
- Pichler Hefti, J.; Leichtle, A.; Stutz, M.; Hefti, U.; Geiser, T.; Huber, A.R.; Merz, T.M. Increased endothelial microparticles and oxidative stress at extreme altitude. Eur. J. Appl. Physiol. 2016, 116, 739–748. [Google Scholar] [CrossRef]
- Tremblay, J.C.; Thom, S.R.; Yang, M.; Ainslie, P.N. Oscillatory shear stress, flow-mediated dilatation, and circulating microparticles at sea level and high altitude. Atherosclerosis 2017, 256, 115–122. [Google Scholar] [CrossRef]
- Tual-Chalot, S.; Guibert, C.; Muller, B.; Savineau, J.P.; Andriantsitohaina, R.; Martinez, M.C. Circulating microparticles from pulmonary hypertensive rats induce endothelial dysfunction. Am. J. Respir. Crit. Care Med. 2010, 182, 261–268. [Google Scholar] [CrossRef]
- Kosanovic, D.; Deo, U.; Gall, H.; Selvakumar, B.; Herold, S.; Weiss, A.; Petrovic, A.; Sydykov, A.; Ghofrani, H.A.; Schermuly, R.T. Enhanced circulating levels of CD3 cells-derived extracellular vesicles in different forms of pulmonary hypertension. Pulm. Circ. 2019, 9, 2045894019864357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosanovic, D.; Platzek, S.M.; Petrovic, A.; Sydykov, A.; Maripov, A.; Mamazhakypov, A.; Sartmyrzaeva, M.; Muratali Uulu, K.; Cholponbaeva, M.; Toktosunova, A.; et al. Circulating Apoptotic Signals During Acute and Chronic Exposure to High Altitude in Kyrgyz Population. Front. Physiol. 2019, 10, 54. [Google Scholar] [CrossRef] [PubMed]
- Sommer, N.; Hüttemann, M.; Pak, O.; Scheibe, S.; Knoepp, F.; Sinkler, C.; Malczyk, M.; Gierhardt, M.; Esfandiary, A.; Kraut, S.; et al. Mitochondrial Complex IV Subunit 4 Isoform 2 Is Essential for Acute Pulmonary Oxygen Sensing. Circ. Res. 2017, 121, 424–438. [Google Scholar] [CrossRef]
- Rozenbaum, Z.; Topilsky, Y.; Khoury, S.; Assi, M.; Balchyunayte, A.; Laufer-Perl, M.; Berliner, S.; Pereg, D.; Entin-Meer, M.; Havakuk, O. Relationship between climate and hemodynamics according to echocardiography. J. Appl. Physiol. 2019, 126, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Miehe, G.; Winiger, M.; Böhner, J.; Yili, Z. The Climatic Diagram Map of High Asia: Purpose and Concepts. Erdkunde 2001, 55, 94–97. [Google Scholar] [CrossRef]
- Boulanger, C.M.; Amabile, N.; Tedgui, A. Circulating microparticles: A potential prognostic marker for atherosclerotic vascular disease. Hypertension 2006, 48, 180–186. [Google Scholar] [CrossRef]
- Amabile, N.; Heiss, C.; Real, W.M.; Minasi, P.; McGlothlin, D.; Rame, E.J.; Grossman, W.; De Marco, T.; Yeghiazarians, Y. Circulating endothelial microparticle levels predict hemodynamic severity of pulmonary hypertension. Am. J. Respir. Crit. Care Med. 2008, 177, 1268–1275. [Google Scholar] [CrossRef]
- Bakouboula, B.; Morel, O.; Faure, A.; Zobairi, F.; Jesel, L.; Trinh, A.; Zupan, M.; Canuet, M.; Grunebaum, L.; Brunette, A.; et al. Procoagulant membrane microparticles correlate with the severity of pulmonary arterial hypertension. Am. J. Respir. Crit. Care Med. 2008, 177, 536–543. [Google Scholar] [CrossRef]
- Curtis, A.M.; Edelberg, J.; Jonas, R.; Rogers, W.T.; Moore, J.S.; Syed, W.; Mohler, E.R.3rd. Endothelial microparticles: Sophisticated vesicles modulating vascular function. Vasc. Med. 2013, 18, 204–214. [Google Scholar] [CrossRef]
- Yong, P.J.; Koh, C.H.; Shim, W.S. Endothelial microparticles: Missing link in endothelial dysfunction? Eur. J. Prev. Cardiol. 2013, 20, 496–512. [Google Scholar] [CrossRef]
- Michiels, C.; Arnould, T.; Remacle, J. Endothelial cell responses to hypoxia: Initiation of a cascade of cellular interactions. Biochim. Biophys. Acta 2000, 1497, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wiley, K.E.; Davenport, A.P. Nitric oxide-mediated modulation of the endothelin-1 signalling pathway in the human cardiovascular system. Br. J. Pharmacol. 2001, 132, 213–220. [Google Scholar] [CrossRef]
- Morganti, A.; Giussani, M.; Sala, C.; Gazzano, G.; Marana, I.; Pierini, A.; Savoia, M.T.; Ghio, F.; Cogo, A.; Zanchetti, A. Effects of exposure to high altitude on plasma endothelin-1 levels in normal subjects. J. Hypertens 1995, 13, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Goerre, S.; Wenk, M.; Bartsch, P.; Luscher, T.F.; Niroomand, F.; Hohenhaus, E.; Oelz, O.; Reinhart, W.H. Endothelin-1 in pulmonary hypertension associated with high-altitude exposure. Circulation 1995, 91, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Modesti, P.A.; Vanni, S.; Morabito, M.; Modesti, A.; Marchetta, M.; Gamberi, T.; Sofi, F.; Savia, G.; Mancia, G.; Gensini, G.F.; et al. Role of endothelin-1 in exposure to high altitude: Acute Mountain Sickness and Endothelin-1 (ACME-1) study. Circulation 2006, 114, 1410–1416. [Google Scholar] [CrossRef] [PubMed]
- Droma, Y.; Hayano, T.; Takabayashi, Y.; Koizumi, T.; Kubo, K.; Kobayashi, T.; Sekiguchi, M. Endothelin-1 and interleukin-8 in high altitude pulmonary oedema. Eur. Respir. J. 1996, 9, 1947–1949. [Google Scholar] [CrossRef]
- Sartori, C.; Vollenweider, L.; Loffler, B.M.; Delabays, A.; Nicod, P.; Bartsch, P.; Scherrer, U. Exaggerated endothelin release in high-altitude pulmonary edema. Circulation 1999, 99, 2665–2668. [Google Scholar] [CrossRef]
- Brewster, L.M.; Garcia, V.P.; Levy, M.V.; Stockelman, K.A.; Goulding, A.; DeSouza, N.M.; Greiner, J.J.; Hijmans, J.G.; DeSouza, C.A. Endothelin-1-induced endothelial microvesicles impair endothelial cell function. J. Appl. Physiol. 2020, 128, 1497–1505. [Google Scholar] [CrossRef]
- Diehl, P.; Aleker, M.; Helbing, T.; Sossong, V.; Germann, M.; Sorichter, S.; Bode, C.; Moser, M. Increased platelet, leukocyte and endothelial microparticles predict enhanced coagulation and vascular inflammation in pulmonary hypertension. J. Thromb. Thrombolysis 2011, 31, 173–179. [Google Scholar] [CrossRef]
- Mohan, A.; Agarwal, S.; Clauss, M.; Britt, N.S.; Dhillon, N.K. Extracellular vesicles: Novel communicators in lung diseases. Respir. Res. 2020, 21, 175. [Google Scholar] [CrossRef]
- Diehl, P.; Fricke, A.; Sander, L.; Stamm, J.; Bassler, N.; Htun, N.; Ziemann, M.; Helbing, T.; El-Osta, A.; Jowett, J.B.; et al. Microparticles: Major transport vehicles for distinct microRNAs in circulation. Cardiovasc. Res. 2012, 93, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Boucherat, O.; Potus, F.; Bonnet, S. microRNA and Pulmonary Hypertension. Adv. Exp. Med. Biol. 2015, 888, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Pullamsetti, S.S.; Doebele, C.; Fischer, A.; Savai, R.; Kojonazarov, B.; Dahal, B.K.; Ghofrani, H.A.; Weissmann, N.; Grimminger, F.; Bonauer, A.; et al. Inhibition of microRNA-17 improves lung and heart function in experimental pulmonary hypertension. Am. J. Respir. Crit. Care Med. 2012, 185, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, C.J.; Wharton, J.; Boon, R.A.; Roexe, T.; Tsang, H.; Wojciak-Stothard, B.; Chakrabarti, A.; Howard, L.S.; Gibbs, J.S.; Lawrie, A.; et al. Reduced microRNA-150 is associated with poor survival in pulmonary arterial hypertension. Am. J. Respir. Crit. Care Med. 2013, 187, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Chun, H.J.; Bonnet, S.; Chan, S.Y. Translational Advances in the Field of Pulmonary Hypertension. Translating MicroRNA Biology in Pulmonary Hypertension. It Will Take More Than “miR” Words. Am. J. Respir. Crit. Care Med. 2017, 195, 167–178. [Google Scholar] [CrossRef]
- Miao, C.; Chang, J.; Zhang, G. Recent research progress of microRNAs in hypertension pathogenesis, with a focus on the roles of miRNAs in pulmonary arterial hypertension. Mol. Biol. Rep. 2018, 45, 2883–2896. [Google Scholar] [CrossRef]
- Kulshreshtha, R.; Ferracin, M.; Wojcik, S.E.; Garzon, R.; Alder, H.; Agosto-Perez, F.J.; Davuluri, R.; Liu, C.G.; Croce, C.M.; Negrini, M.; et al. A microRNA signature of hypoxia. Mol. Cell Biol. 2007, 27, 1859–1867. [Google Scholar] [CrossRef]
- Bavelloni, A.; Ramazzotti, G.; Poli, A.; Piazzi, M.; Focaccia, E.; Blalock, W.; Faenza, I. MiRNA-210: A Current Overview. Anticancer Res. 2017, 37, 6511–6521. [Google Scholar] [CrossRef]
- Hui, X.; Al-Ward, H.; Shaher, F.; Liu, C.Y.; Liu, N. The Role of miR-210 in the Biological System: A Current Overview. Hum. Hered 2020, 84, 233–239. [Google Scholar] [CrossRef]
- Blissenbach, B.; Nakas, C.T.; Krönke, M.; Geiser, T.; Merz, T.M.; Pichler Hefti, J. Hypoxia-induced changes in plasma micro-RNAs correlate with pulmonary artery pressure at high altitude. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 314, L157–L164. [Google Scholar] [CrossRef]
- Lacedonia, D.; Scioscia, G.; Pia Palladino, G.; Gallo, C.; Carpagnano, G.E.; Sabato, R.; Foschino Barbaro, M.P. MicroRNA expression profile during different conditions of hypoxia. Oncotarget 2018, 9, 35114–35122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Megha, S.; Basu, U.; Kav, N.N.V. Regulation of low temperature stress in plants by microRNAs. Plant Cell Environ. 2018, 41, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Frigault, J.J.; Morin, M.D.; Morin, P.J. Differential expression and emerging functions of non-coding RNAs in cold adaptation. J. Comp. Physiol. B 2017, 187, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Zhen, L.; Guo, W.; Peng, M.; Liu, Y.; Zang, S.; Ji, H.; Li, S.; Yang, H. Identification of cold-responsive miRNAs in rats by deep sequencing. J. Therm Biol. 2017, 66, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, K.; Qi, G.; Yan, R.; Yang, Y.; Li, Y.; Wang, S.; Bai, Z.; Ge, R.L. Adipose-derived exosomal miR-210/92a cluster inhibits adipose browning via the FGFR-1 signaling pathway in high-altitude hypoxia. Sci. Rep. 2020, 10, 14390. [Google Scholar] [CrossRef]
- Leung, D.W.; Cachianes, G.; Kuang, W.J.; Goeddel, D.V.; Ferrara, N. Vascular endothelial growth factor is a secreted angiogenic mitogen. Science 1989, 246, 1306–1309. [Google Scholar] [CrossRef]
- Liu, J.; Wang, W.; Wang, L.; Chen, S.; Tian, B.; Huang, K.; Corrigan, C.J.; Ying, S.; Wang, W.; Wang, C. IL-33 Initiates Vascular Remodelling in Hypoxic Pulmonary Hypertension by up-Regulating HIF-1α and VEGF Expression in Vascular Endothelial Cells. EBioMedicine 2018, 33, 196–210. [Google Scholar] [CrossRef]
- Borgia, F.; Custurone, P.; Li Pomi, F.; Cordiano, R.; Alessandrello, C.; Gangemi, S. IL-31: State of the Art for an Inflammation-Oriented Interleukin. Int. J. Mol. Sci. 2022, 23, 6507. [Google Scholar] [CrossRef]
- Melincovici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Fan, N.; Liu, C.; Ren, M. Effect of different high altitudes on vascular endothelial function in healthy people. Medicine 2020, 99, e19292. [Google Scholar] [CrossRef]
- Walter, R.; Maggiorini, M.; Scherrer, U.; Contesse, J.; Reinhart, W.H. Effects of high-altitude exposure on vascular endothelial growth factor levels in man. Eur. J. Appl. Physiol. 2001, 85, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Droma, Y.; Kinjo, T.; Nozawa, S.; Kobayashi, N.; Yasuo, M.; Kitaguchi, Y.; Ota, M.; Basnyat, B.; Hanaoka, M. Hypoxia-tolerant vascular endothelial growth factor-A (VEGF-A) and the genetic variations of VEGFA in Sherpa highlanders. bioRxiv 2020. [Google Scholar] [CrossRef]
- Appenzeller, O.; Minko, T.; Qualls, C.; Pozharov, V.; Gamboa, J.; Gamboa, A.; Wang, Y. Gene expression, autonomic function and chronic hypoxia:lessons from the Andes. Clin. Auton Res. 2006, 16, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Luks, A.M.; Swenson, E.R.; Bartsch, P. Acute high-altitude sickness. Eur. Respir. Rev 2017, 26, 160096. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, J.; Jiang, D.; Wuren, T.; Ma, S.; Du, Y.; Yi, X.; Wu, S. The plasma level changes of VEGF and soluble VEGF receptor-1 are associated with high-altitude pulmonary edema. J. Med. Investig. 2018, 65, 64–68. [Google Scholar] [CrossRef]
- Ge, R.L.; Mo, V.Y.; Januzzi, J.L.; Jin, G.; Yang, Y.; Han, S.; Wood, M.J.; Levine, B.D. B-type natriuretic peptide, vascular endothelial growth factor, endothelin-1, and nitric oxide synthase in chronic mountain sickness. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H1427–H1433. [Google Scholar] [CrossRef]
- Ma, L.; Chen, Y.; Jin, G.; Yang, Y.; Ga, Q.; Ge, R.L. Vascular Endothelial Growth Factor as a Prognostic Parameter in Subjects with “Plateau Red Face”. High Alt. Med. Biol. 2015, 16, 147–153. [Google Scholar] [CrossRef]
- Joo, C.H.; Allan, R.; Drust, B.; Close, G.L.; Jeong, T.S.; Bartlett, J.D.; Mawhinney, C.; Louhelainen, J.; Morton, J.P.; Gregson, W. Passive and post-exercise cold-water immersion augments PGC-1α and VEGF expression in human skeletal muscle. Eur. J. Appl. Physiol. 2016, 116, 2315–2326. [Google Scholar] [CrossRef]
- Kim, J.C.; Yi, H.K.; Hwang, P.H.; Yoon, J.S.; Kim, H.J.; Kawano, F.; Ohira, Y.; Kim, C.K. Effects of cold-water immersion on VEGF mRNA and protein expression in heart and skeletal muscles of rats. Acta Physiol. Scand 2005, 183, 389–397. [Google Scholar] [CrossRef]
- Banchero, N.; Kayar, S.R.; Lechner, A.J. Increased capillarity in skeletal muscle of growing guinea pigs acclimated to cold and hypoxia. Respir. Physiol. 1985, 62, 245–255. [Google Scholar] [CrossRef]
- Hudlická, O. Growth of capillaries in skeletal and cardiac muscle. Circ. Res. 1982, 50, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Sillau, A.H.; Aquin, L.; Lechner, A.J.; Bui, M.V.; Banchero, N. Increased capillary supply in skeletal muscle of guinea pigs acclimated to cold. Respir. Physiol. 1980, 42, 233–245. [Google Scholar] [CrossRef]
- Aliotta, J.M.; Pereira, M.; Amaral, A.; Sorokina, A.; Igbinoba, Z.; Hasslinger, A.; El-Bizri, R.; Rounds, S.I.; Quesenberry, P.J.; Klinger, J.R. Induction of pulmonary hypertensive changes by extracellular vesicles from monocrotaline-treated mice. Cardiovasc. Res. 2013, 100, 354–362. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Experimental Group | TRG (mmHg) Warm | TRG (mmHg) Cold | Age (Years) | f/m Ratio (%) | Associated Figure(s) |
|---|---|---|---|---|---|---|
| MPs numbers and miRNA content | LA (n = 12) | 20.0 ± 0.5 | 23.2 ± 0.6 * | 52.3 ± 2.1 | 42/58 | 3 and 5 |
| HA (n = 12) | 21.1 ± 0.6 | 24.0 ± 0.9 * | 37.6 ± 2.8 | 50/50 | 3 and 5 | |
| HAPH (n = 12) | 44.3 ± 1.0 §,$ | 49.0 ± 1.1 *,§,$ | 50.8 ± 3.6 | 75/25 | 3 and 5 | |
| Effects of MPs on PASMCs | LA (n = 7) | 19.3 ± 0.7 | 21.8 ± 0.4 * | 51.9 ± 3.0 | 43/57 | 4 |
| HA (n = 7) | 20.3 ± 1.0 | 22.2 ± 0.8 * | 38.1 ± 4.5 | 43/57 | 4 | |
| HAPH (n = 7) | 45.2 ± 1.4 §,$ | 50.1 ± 1.4 *,§,$ | 55.6 ± 5.2 | 71/29 | 4 | |
| Effects of MPs on PAECs: angiogenesis | LA (n = 3) | 18.1 ± 0.6 | 21.0 ± 0.6 | 58.3 ± 2.7 | 67/33 | 6 |
| HA (n = 3) | 20.4 ± 0.8 | 22.1 ± 0.9 | 44.0 ± 2.6 | 67/33 | 6 | |
| HAPH (n = 3) | 46.0 ± 3.3 §,$ | 52.8 ± 1.8 *,§,$ | 52.7 ± 11.3 | 67/33 | 6 | |
| Effects of MPs on PAECs: Western Blot | LA (n = 4) | 17.8 ± 0.5 | 21.1 ± 0.4 * | 56.7 ± 2.5 | 50/50 | 7 |
| HA (n = 4) | 19.3 ± 1.2 | 21.3 ± 1.0 | 41.0 ± 3.5 | 50/50 | 7 | |
| HAPH (n = 4) | 44.6 ± 2.7 §,$ | 52.6 ± 1.3 *,§,$ | 53.7 ± 8.1 | 75/25 | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sydykov, A.; Petrovic, A.; Maripov, A.M.; Gredic, M.; Bermes, D.G.; Kushubakova, N.; Muratali Uulu, K.; Pilz, C.; Cholponbaeva, M.; Duishobaev, M.; et al. Circulating Microparticles Are Differentially Increased in Lowlanders and Highlanders with High Altitude Induced Pulmonary Hypertension during the Cold Season. Cells 2022, 11, 2932. https://doi.org/10.3390/cells11192932
Sydykov A, Petrovic A, Maripov AM, Gredic M, Bermes DG, Kushubakova N, Muratali Uulu K, Pilz C, Cholponbaeva M, Duishobaev M, et al. Circulating Microparticles Are Differentially Increased in Lowlanders and Highlanders with High Altitude Induced Pulmonary Hypertension during the Cold Season. Cells. 2022; 11(19):2932. https://doi.org/10.3390/cells11192932
Chicago/Turabian StyleSydykov, Akylbek, Aleksandar Petrovic, Abdirashit M. Maripov, Marija Gredic, Daniel Gerd Bermes, Nadira Kushubakova, Kubatbek Muratali Uulu, Christina Pilz, Meerim Cholponbaeva, Melis Duishobaev, and et al. 2022. "Circulating Microparticles Are Differentially Increased in Lowlanders and Highlanders with High Altitude Induced Pulmonary Hypertension during the Cold Season" Cells 11, no. 19: 2932. https://doi.org/10.3390/cells11192932
APA StyleSydykov, A., Petrovic, A., Maripov, A. M., Gredic, M., Bermes, D. G., Kushubakova, N., Muratali Uulu, K., Pilz, C., Cholponbaeva, M., Duishobaev, M., Satybaldyev, S., Satieva, N., Mamazhakypov, A., Sartmyrzaeva, M., Omurzakova, N., Kerimbekova, Z., Baktybek, N., Kulchoroeva, C., Pak, O., ... Kosanovic, D. (2022). Circulating Microparticles Are Differentially Increased in Lowlanders and Highlanders with High Altitude Induced Pulmonary Hypertension during the Cold Season. Cells, 11(19), 2932. https://doi.org/10.3390/cells11192932