
Recent Developments on the Roles of Calcium Signals and Potential Therapy Targets in Cervical Cancer

Abstract
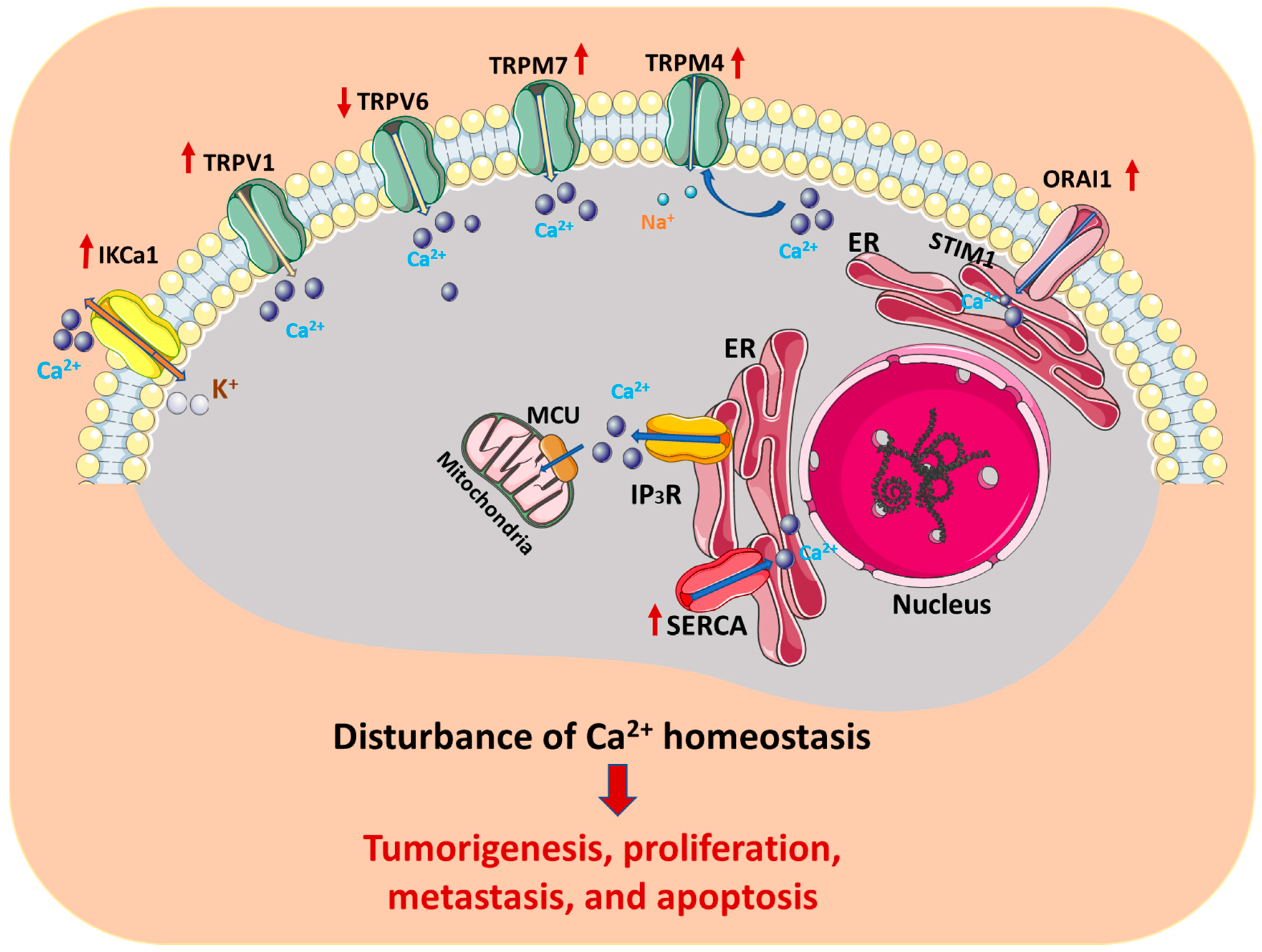
:1. Introduction
2. Ca2+ Channels, Pumps, and Interacting Proteins and Ligands in Cervical Cancer
2.1. Transient Receptor Potential (TRP) Channels
2.2. Ca2+ Release-Activated Ca2+ (CRAC) Channel
2.3. Inositol 1,4,5-Triphosphate Receptor (IP3R) and Ryanodine Receptors (RyR) Channels
2.4. SERCA Pump
2.5. Mitochondrial Calcium Uniporter (MCU) Channel
2.6. Others
3. Summary and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Patergnani, S.; Danese, A.; Bouhamida, E.; Aguiari, G.; Previati, M.; Pinton, P.; Giorgi, C. Various Aspects of Calcium Signaling in the Regulation of Apoptosis, Autophagy, Cell Proliferation, and Cancer. Int. J. Mol. Sci. 2020, 21, 8323. [Google Scholar] [CrossRef] [PubMed]
- Monteith, G.R.; Prevarskaya, N.; Roberts-Thomson, S.J. The calcium-cancer signalling nexus. Nat. Rev. Cancer 2017, 17, 367–380. [Google Scholar] [CrossRef]
- Islam, M.S. Calcium Signaling: From Basic to Bedside. Adv. Exp. Med. Biol. 2020, 1131, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Liu, R.; Yan, X.; Zhang, Q.; Zeng, X.; Yuan, X. Intensive stretch-activated CRT-PMCA1 feedback loop promoted apoptosis of myoblasts through Ca2+ overloading. Apoptosis 2022. [Google Scholar] [CrossRef] [PubMed]
- Fani, G.; La Torre, C.E.; Cascella, R.; Cecchi, C.; Vendruscolo, M.; Chiti, F. Misfolded protein oligomers induce an increase of intracellular Ca2+ causing an escalation of reactive oxidative species. Cell Mol. Life Sci. 2022, 79, 500. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Bouvard, V.; Baan, R.; Straif, K.; Grosse, Y.; Secretan, B.; El Ghissassi, F.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; Galichet, L.; et al. A review of human carcinogens—Part B: Biological agents. Lancet Oncol. 2009, 10, 321–322. [Google Scholar] [CrossRef]
- Marchi, S.; Giorgi, C.; Galluzzi, L.; Pinton, P. Ca2+ Fluxes and Cancer. Mol. Cell 2020, 78, 1055–1069. [Google Scholar] [CrossRef]
- Arbabian, A.; Brouland, J.P.; Gelebart, P.; Kovacs, T.; Bobe, R.; Enouf, J.; Papp, B. Endoplasmic reticulum calcium pumps and cancer. Biofactors 2011, 37, 139–149. [Google Scholar] [CrossRef]
- Han, G.H.; Chay, D.B.; Nam, S.; Cho, H.; Chung, J.Y.; Kim, J.H. The Combination of Transient Receptor Potential Vanilloid Type 1 (TRPV1) and Phosphatase and Tension Homolog (PTEN) is an Effective Prognostic Biomarker in Cervical Cancer. Int. J. Gynecol. Pathol. 2021, 40, 214–223. [Google Scholar] [CrossRef]
- Lucido, C.T.; Wynja, E.; Madeo, M.; Williamson, C.S.; Schwartz, L.E.; Imblum, B.A.; Drapkin, R.; Vermeer, P.D. Innervation of cervical carcinoma is mediated by cancer-derived exosomes. Gynecol. Oncol. 2019, 154, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Xiao, L.; Jang, X.X.; Xiong, Y.; Li, Q.; Yue, X.J.; Wei, Y.J.; Wei, Y.X.; Ma, Y.L.; Yu, Y.H. TRPV6 is a prognostic marker in early-stage cervical squamous cell carcinoma. Tumour Biol. 2016, 37, 15743–15751. [Google Scholar] [CrossRef] [PubMed]
- Narayan, G.; Bourdon, V.; Chaganti, S.; Arias-Pulido, H.; Nandula, S.V.; Rao, P.H.; Gissmann, L.; Durst, M.; Schneider, A.; Pothuri, B.; et al. Gene dosage alterations revealed by cDNA microarray analysis in cervical cancer: Identification of candidate amplified and overexpressed genes. Genes Chromosom. Cancer 2007, 46, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Yu, J.J. MicroRNA-150 suppresses epithelial-mesenchymal transition, invasion, and metastasis in prostate cancer through the TRPM4-mediated beta-catenin signaling pathway. Am. J. Physiol. Cell. Physiol. 2019, 316, C463–C480. [Google Scholar] [CrossRef] [PubMed]
- Armisen, R.; Marcelain, K.; Simon, F.; Tapia, J.C.; Toro, J.; Quest, A.F.; Stutzin, A. TRPM4 enhances cell proliferation through up-regulation of the beta-catenin signaling pathway. J. Cell. Physiol. 2011, 226, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gan, L.; Zhang, J. miR-543 inhibites cervical cancer growth and metastasis by targeting TRPM7. Chem. Biol. Interact. 2019, 302, 83–92. [Google Scholar] [CrossRef]
- Dong, R.F.; Zhuang, Y.J.; Wang, Y.; Zhang, Z.Y.; Xu, X.Z.; Mao, Y.R.; Yu, J.J. Tumor suppressor miR-192-5p targets TRPM7 and inhibits proliferation and invasion in cervical cancer. Kaohsiung J. Med. Sci. 2021, 37, 699–708. [Google Scholar] [CrossRef]
- Qi, H.; Lu, L.; Wang, L. Long Noncoding RNA ST7-AS1 Upregulates TRPM7 Expression by Sponging microRNA-543 to Promote Cervical Cancer Progression. Onco Targets Ther. 2020, 13, 7257–7269. [Google Scholar] [CrossRef]
- Chen, Y.T.; Chen, Y.F.; Chiu, W.T.; Liu, K.Y.; Liu, Y.L.; Chang, J.Y.; Chang, H.C.; Shen, M.R. Microtubule-associated histone deacetylase 6 supports the calcium store sensor STIM1 in mediating malignant cell behaviors. Cancer Res. 2013, 73, 4500–4509. [Google Scholar] [CrossRef]
- Chen, Y.F.; Chiu, W.T.; Chen, Y.T.; Lin, P.Y.; Huang, H.J.; Chou, C.Y.; Chang, H.C.; Tang, M.J.; Shen, M.R. Calcium store sensor stromal-interaction molecule 1-dependent signaling plays an important role in cervical cancer growth, migration, and angiogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 15225–15230. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.C.; Chang, T.Y.; Chen, T.C.; Lin, W.S.; Chang, S.C.; Lee, Y.J. ITPR3 gene haplotype is associated with cervical squamous cell carcinoma risk in Taiwanese women. Oncotarget 2017, 8, 10085–10090. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.C.; Chang, T.Y.; Chen, T.C.; Chang, S.C.; Chen, W.F.; Chan, H.W.; Lin, W.S.; Wu, F.T.; Lee, Y.J. Genetic polymorphisms in the ITPKC gene and cervical squamous cell carcinoma risk. Cancer Immunol. Immunother. 2012, 61, 2153–2159. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ouyang, Z.; Zhang, Q.; Wang, L.; Shen, Y.; Wu, X.; Gu, Y.; Shu, Y.; Yu, B.; Wu, X.; et al. SBF-1 exerts strong anticervical cancer effect through inducing endoplasmic reticulum stress-associated cell death via targeting sarco/endoplasmic reticulum Ca(2+)-ATPase 2. Cell Death Dis. 2014, 5, e1581. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Sang, L.; Wang, X. S100 Calcium Binding Protein A11 (S100A11) Promotes The Proliferation, Migration And Invasion Of Cervical Cancer Cells, And Activates Wnt/beta-Catenin Signaling. Onco Targets Ther. 2019, 12, 8675–8685. [Google Scholar] [CrossRef]
- Wang, X.; Yang, J.; Qian, J.; Liu, Z.; Chen, H.; Cui, Z. S100A14, a mediator of epithelial-mesenchymal transition, regulates proliferation, migration and invasion of human cervical cancer cells. Am. J. Cancer Res. 2015, 5, 1484–1495. [Google Scholar]
- Liu, L.; Zhan, P.; Nie, D.; Fan, L.; Lin, H.; Gao, L.; Mao, X. Intermediate-Conductance-Ca2-Activated K Channel IKCa1 Is Upregulated and Promotes Cell Proliferation in Cervical Cancer. Med. Sci. Monit. Basic Res. 2017, 23, 45–57. [Google Scholar] [CrossRef]
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome Biol. 2011, 12, 218. [Google Scholar] [CrossRef]
- Vangeel, L.; Voets, T. Transient Receptor Potential Channels and Calcium Signaling. Cold Spring Harb. Perspect Biol. 2019, 11, a035048. [Google Scholar] [CrossRef]
- Li, H. TRP Channel Classification. Adv. Exp. Med. Biol. 2017, 976, 1–8. [Google Scholar] [CrossRef]
- Karki, T.; Tojkander, S. TRPV Protein Family-From Mechanosensing to Cancer Invasion. Biomolecules 2021, 11, 1019. [Google Scholar] [CrossRef]
- Borgstrom, A.; Peinelt, C.; Stoklosa, P. TRPM4 in Cancer—A New Potential Drug Target. Biomolecules 2021, 11, 229. [Google Scholar] [CrossRef] [PubMed]
- Aghazadeh Tabrizi, M.; Baraldi, P.G.; Baraldi, S.; Gessi, S.; Merighi, S.; Borea, P.A. Medicinal Chemistry, Pharmacology, and Clinical Implications of TRPV1 Receptor Antagonists. Med. Res. Rev. 2017, 37, 936–983. [Google Scholar] [CrossRef] [PubMed]
- Zhai, K.; Liskova, A.; Kubatka, P.; Busselberg, D. Calcium Entry through TRPV1: A Potential Target for the Regulation of Proliferation and Apoptosis in Cancerous and Healthy Cells. Int. J. Mol. Sci. 2020, 21, 4177. [Google Scholar] [CrossRef] [PubMed]
- De La Chapa, J.; Valdez, M.; Ruiz, F., 3rd; Gonzales, K.; Mitchell, W.; McHardy, S.F.; Hart, M.; Polusani, S.R.; Gonzales, C.B. Synthesis and SAR of novel capsazepine analogs with significant anti-cancer effects in multiple cancer types. Bioorg. Med. Chem. 2019, 27, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Ramer, R.; Merkord, J.; Rohde, H.; Hinz, B. Cannabidiol inhibits cancer cell invasion via upregulation of tissue inhibitor of matrix metalloproteinases-1. Biochem. Pharmacol. 2010, 79, 955–966. [Google Scholar] [CrossRef] [PubMed]
- Yelshanskaya, M.V.; Nadezhdin, K.D.; Kurnikova, M.G.; Sobolevsky, A.I. Structure and function of the calcium-selective TRP channel TRPV6. J. Physiol. 2021, 599, 2673–2697. [Google Scholar] [CrossRef]
- Bolanz, K.A.; Hediger, M.A.; Landowski, C.P. The role of TRPV6 in breast carcinogenesis. Mol. Cancer Ther. 2008, 7, 271–279. [Google Scholar] [CrossRef]
- Xu, X.; Li, N.; Wang, Y.; Yu, J.; Mi, J. Calcium channel TRPV6 promotes breast cancer metastasis by NFATC2IP. Cancer Lett. 2021, 519, 150–160. [Google Scholar] [CrossRef]
- Song, H.; Dong, M.; Zhou, J.; Sheng, W.; Li, X.; Gao, W. Expression and prognostic significance of TRPV6 in the development and progression of pancreatic cancer. Oncol. Rep. 2018, 39, 1432–1440. [Google Scholar] [CrossRef]
- Fan, H.; Shen, Y.X.; Yuan, Y.F. Expression and prognostic roles of TRPV5 and TRPV6 in non-small cell lung cancer after curative resection. Asian Pac. J. Cancer Prev. 2014, 15, 2559–2563. [Google Scholar] [CrossRef]
- So, C.L.; Milevskiy, M.J.G.; Monteith, G.R. Transient receptor potential cation channel subfamily V and breast cancer. Lab. Investig. 2020, 100, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Fixemer, T.; Wissenbach, U.; Flockerzi, V.; Bonkhoff, H. Expression of the Ca2+-selective cation channel TRPV6 in human prostate cancer: A novel prognostic marker for tumor progression. Oncogene 2003, 22, 7858–7861. [Google Scholar] [CrossRef] [PubMed]
- Mathar, I.; Jacobs, G.; Kecskes, M.; Menigoz, A.; Philippaert, K.; Vennekens, R. Trpm4. Handb. Exp. Pharmacol. 2014, 222, 461–487. [Google Scholar] [CrossRef] [PubMed]
- Fleig, A.; Penner, R. The TRPM ion channel subfamily: Molecular, biophysical and functional features. Trends Pharmacol. Sci. 2004, 25, 633–639. [Google Scholar] [CrossRef]
- Kappel, S.; Stoklosa, P.; Hauert, B.; Ross-Kaschitza, D.; Borgstrom, A.; Baur, R.; Galvan, J.A.; Zlobec, I.; Peinelt, C. TRPM4 is highly expressed in human colorectal tumor buds and contributes to proliferation, cell cycle, and invasion of colorectal cancer cells. Mol. Oncol. 2019, 13, 2393–2405. [Google Scholar] [CrossRef]
- Sagredo, A.I.; Sagredo, E.A.; Pola, V.; Echeverria, C.; Andaur, R.; Michea, L.; Stutzin, A.; Simon, F.; Marcelain, K.; Armisen, R. TRPM4 channel is involved in regulating epithelial to mesenchymal transition, migration, and invasion of prostate cancer cell lines. J. Cell. Physiol. 2019, 234, 2037–2050. [Google Scholar] [CrossRef]
- Rivas, J.; Diaz, N.; Silva, I.; Morales, D.; Lavanderos, B.; Alvarez, A.; Saldias, M.P.; Pulgar, E.; Cruz, P.; Maureira, D.; et al. KCTD5, a novel TRPM4-regulatory protein required for cell migration as a new predictor for breast cancer prognosis. FASEB J. 2020, 34, 7847–7865. [Google Scholar] [CrossRef]
- Guinamard, R.; Hof, T.; Del Negro, C.A. The TRPM4 channel inhibitor 9-phenanthrol. Br. J. Pharmacol. 2014, 171, 1600–1613. [Google Scholar] [CrossRef]
- Zou, Z.G.; Rios, F.J.; Montezano, A.C.; Touyz, R.M. TRPM7, Magnesium, and Signaling. Int. J. Mol. Sci. 2019, 20, 1877. [Google Scholar] [CrossRef]
- Chubanov, V.; Gudermann, T. Mapping TRPM7 Function by NS8593. Int. J. Mol. Sci. 2020, 21, 7017. [Google Scholar] [CrossRef]
- Pugliese, D.; Armuzzi, A.; Castri, F.; Benvenuto, R.; Mangoni, A.; Guidi, L.; Gasbarrini, A.; Rapaccini, G.L.; Wolf, F.I.; Trapani, V. TRPM7 is overexpressed in human IBD-related and sporadic colorectal cancer and correlates with tumor grade. Dig. Liver Dis. 2020, 52, 1188–1194. [Google Scholar] [CrossRef] [PubMed]
- Yee, N.S. Role of TRPM7 in Cancer: Potential as Molecular Biomarker and Therapeutic Target. Pharmaceuticals 2017, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhao, X.; Duan, S. The role of miR-543 in human cancerous and noncancerous diseases. J. Cell. Physiol. 2021, 236, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Numata, T.; Sato-Numata, K.; Okada, Y. TRPM7 is involved in acid-induced necrotic cell death in a manner sensitive to progesterone in human cervical cancer cells. Physiol. Rep. 2019, 7, e14157. [Google Scholar] [CrossRef] [PubMed]
- Gaunt, H.J.; Vasudev, N.S.; Beech, D.J. Transient receptor potential canonical 4 and 5 proteins as targets in cancer therapeutics. Eur. Biophys. J. 2016, 45, 611–620. [Google Scholar] [CrossRef]
- Sun, Y.; Ye, C.; Tian, W.; Ye, W.; Gao, Y.Y.; Feng, Y.D.; Zhang, H.N.; Ma, G.Y.; Wang, S.J.; Cao, W.; et al. TRPC1 promotes the genesis and progression of colorectal cancer via activating CaM-mediated PI3K/AKT signaling axis. Oncogenesis 2021, 10, 67. [Google Scholar] [CrossRef]
- Dhennin-Duthille, I.; Gautier, M.; Faouzi, M.; Guilbert, A.; Brevet, M.; Vaudry, D.; Ahidouch, A.; Sevestre, H.; Ouadid-Ahidouch, H. High expression of transient receptor potential channels in human breast cancer epithelial cells and tissues: Correlation with pathological parameters. Cell Physiol. Biochem. 2011, 28, 813–822. [Google Scholar] [CrossRef]
- Asghar, M.Y.; Tornquist, K. Transient Receptor Potential Canonical (TRPC) Channels as Modulators of Migration and Invasion. Int. J. Mol. Sci. 2020, 21, 1739. [Google Scholar] [CrossRef]
- Zomot, E.; Achildiev Cohen, H.; Dagan, I.; Militsin, R.; Palty, R. Bidirectional regulation of calcium release-activated calcium (CRAC) channel by SARAF. J. Cell. Biol. 2021, 220, e202104007. [Google Scholar] [CrossRef]
- Parekh, A.B. Store-operated CRAC channels: Function in health and disease. Nat. Rev. Drug Discov. 2010, 9, 399–410. [Google Scholar] [CrossRef]
- Khan, H.Y.; Mazahir, I.; Reddy, S.; Fazili, F.; Azmi, A. Roles of CRAC channel in cancer: Implications for therapeutic development. Expert Rev. Precis. Med. Drug Dev. 2020, 5, 371–382. [Google Scholar] [CrossRef]
- Chalmers, S.B.; Monteith, G.R. ORAI channels and cancer. Cell Calcium. 2018, 74, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Hammad, A.S.; Machaca, K. Store Operated Calcium Entry in Cell Migration and Cancer Metastasis. Cells 2021, 10, 1246. [Google Scholar] [CrossRef]
- Jardin, I.; Rosado, J.A. STIM and calcium channel complexes in cancer. Biochim. Biophys. Acta 2016, 1863, 1418–1426. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Jan, L.Y. Targeting potassium channels in cancer. J. Cell. Biol. 2014, 206, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zhang, J.J.; Huang, X.Y. Orai1 and STIM1 are critical for breast tumor cell migration and metastasis. Cancer Cell 2009, 15, 124–134. [Google Scholar] [CrossRef]
- Santulli, G.; Lewis, D.; des Georges, A.; Marks, A.R.; Frank, J. Ryanodine Receptor Structure and Function in Health and Disease. Subcell Biochem. 2018, 87, 329–352. [Google Scholar] [CrossRef]
- Seo, M.D.; Velamakanni, S.; Ishiyama, N.; Stathopulos, P.B.; Rossi, A.M.; Khan, S.A.; Dale, P.; Li, C.; Ames, J.B.; Ikura, M.; et al. Structural and functional conservation of key domains in InsP3 and ryanodine receptors. Nature 2012, 483, 108–112. [Google Scholar] [CrossRef]
- Jia, X.; Yang, Q.; Gao, C.; Chen, X.; Li, Y.; Su, H.; Zheng, Y.; Zhang, S.; Wang, Z.; Wang, H.; et al. Stimulation of vascular smooth muscle cell proliferation by stiff matrix via the IKCa channel-dependent Ca2+ signaling. J. Cell. Physiol. 2021, 236, 6897–6906. [Google Scholar] [CrossRef]
- Berridge, M.J. Inositol trisphosphate and calcium signalling mechanisms. Biochim. Biophys. Acta 2009, 1793, 933–940. [Google Scholar] [CrossRef]
- Berridge, M.J. The Inositol Trisphosphate/Calcium Signaling Pathway in Health and Disease. Physiol. Rev. 2016, 96, 1261–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroder, D.; Todter, K.; Gonzalez, B.; Franco-Echevarria, E.; Rohaly, G.; Blecher, C.; Lin, H.Y.; Mayr, G.W.; Windhorst, S. The new InsP3Kinase inhibitor BIP-4 is competitive to InsP3 and blocks proliferation and adhesion of lung cancer cells. Biochem. Pharmacol. 2015, 96, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Ueasilamongkol, P.; Khamphaya, T.; Guerra, M.T.; Rodrigues, M.A.; Gomes, D.A.; Kong, Y.; Wei, W.; Jain, D.; Trampert, D.C.; Ananthanarayanan, M.; et al. Type 3 Inositol 1,4,5-Trisphosphate Receptor Is Increased and Enhances Malignant Properties in Cholangiocarcinoma. Hepatology 2020, 71, 583–599. [Google Scholar] [CrossRef] [PubMed]
- Rezuchova, I.; Hudecova, S.; Soltysova, A.; Matuskova, M.; Durinikova, E.; Chovancova, B.; Zuzcak, M.; Cihova, M.; Burikova, M.; Penesova, A.; et al. Type 3 inositol 1,4,5-trisphosphate receptor has antiapoptotic and proliferative role in cancer cells. Cell Death Dis. 2019, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Imboden, J.B.; Pattison, G. Regulation of inositol 1,4,5-trisphosphate kinase activity after stimulation of human T cell antigen receptor. J. Clin. Investig. 1987, 79, 1538–1541. [Google Scholar] [CrossRef]
- Zalk, R.; Marks, A.R. Ca2+ Release Channels Join the ‘Resolution Revolution’. Trends Biochem. Sci. 2017, 42, 543–555. [Google Scholar] [CrossRef]
- Lanner, J.T.; Georgiou, D.K.; Joshi, A.D.; Hamilton, S.L. Ryanodine receptors: Structure, expression, molecular details, and function in calcium release. Cold Spring Harb. Perspect Biol. 2010, 2, a003996. [Google Scholar] [CrossRef]
- Giannini, G.; Clementi, E.; Ceci, R.; Marziali, G.; Sorrentino, V. Expression of a ryanodine receptor-Ca2+ channel that is regulated by TGF-beta. Science 1992, 257, 91–94. [Google Scholar] [CrossRef]
- Schmitt, K.; Molfenter, B.; Laureano, N.K.; Tawk, B.; Bieg, M.; Hostench, X.P.; Weichenhan, D.; Ullrich, N.D.; Shang, V.; Richter, D.; et al. Somatic mutations and promotor methylation of the ryanodine receptor 2 is a common event in the pathogenesis of head and neck cancer. Int. J. Cancer 2019, 145, 3299–3310. [Google Scholar] [CrossRef]
- Mariot, P.; Prevarskaya, N.; Roudbaraki, M.M.; Le Bourhis, X.; Van Coppenolle, F.; Vanoverberghe, K.; Skryma, R. Evidence of functional ryanodine receptor involved in apoptosis of prostate cancer (LNCaP) cells. Prostate 2000, 43, 205–214. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, Y.; Song, F.; Zheng, H.; Hu, L.; Lu, H.; Liu, P.; Hao, X.; Zhang, W.; Chen, K. Functional SNP in the microRNA-367 binding site in the 3’UTR of the calcium channel ryanodine receptor gene 3 (RYR3) affects breast cancer risk and calcification. Proc. Natl. Acad. Sci. USA 2011, 108, 13653–13658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul, M.; Ramlal, S.; Hoosein, N. Ryanodine receptor expression correlates with tumor grade in breast cancer. Pathol. Oncol. Res. 2008, 14, 157–160. [Google Scholar] [CrossRef]
- Law, B.Y.K.; Michelangeli, F.; Qu, Y.Q.; Xu, S.W.; Han, Y.; Mok, S.W.F.; Dias, I.; Javed, M.U.; Chan, W.K.; Xue, W.W.; et al. Neferine induces autophagy-dependent cell death in apoptosis-resistant cancers via ryanodine receptor and Ca2+-dependent mechanism. Sci. Rep. 2019, 9, 20034. [Google Scholar] [CrossRef] [PubMed]
- Christodoulou, P.; Yiallouris, A.; Michail, A.; Christodoulou, M.I.; Politis, P.K.; Patrikios, I. Altered SERCA Expression in Breast Cancer. Medicina 2021, 57, 1074. [Google Scholar] [CrossRef]
- Li, W.; Song, R.; Fang, X.; Wang, L.; Chen, W.; Tang, P.; Yu, B.; Sun, Y.; Xu, Q. SBF-1, a synthetic steroidal glycoside, inhibits melanoma growth and metastasis through blocking interaction between PDK1 and AKT3. Biochem. Pharmacol. 2012, 84, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Fang, X.; Gao, Y.; Shi, K.; Sun, L.; Yu, B.; Luo, Q.; Xu, Q. SBF-1 inhibits contact hypersensitivity in mice through down-regulation of T-cell-mediated responses. BMC Pharmacol. Toxicol. 2019, 20, 86. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Y.; Li, Z.; Lin, S.; Wang, H.; Sun, J.; Lan, C.; Wu, L.; Sun, D.; Huang, C.; et al. Mitochondrial Calcium Uniporter Drives Metastasis and Confers a Targetable Cystine Dependency in Pancreatic Cancer. Cancer Res. 2022, 82, 2254–2268. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Wang, X.; Lai, Y.; Lin, W.; Huang, Y.; Yin, H.; Hou, R.; Zhang, F. Mitochondrial calcium uniporter promotes cell proliferation and migration in esophageal cancer. Oncol. Lett. 2021, 22, 686. [Google Scholar] [CrossRef] [PubMed]
- Vultur, A.; Gibhardt, C.S.; Stanisz, H.; Bogeski, I. The role of the mitochondrial calcium uniporter (MCU) complex in cancer. Pflugers Arch. 2018, 470, 1149–1163. [Google Scholar] [CrossRef]
- De Stefani, D.; Raffaello, A.; Teardo, E.; Szabo, I.; Rizzuto, R. A forty-kilodalton protein of the inner membrane is the mitochondrial calcium uniporter. Nature 2011, 476, 336–340. [Google Scholar] [CrossRef]
- Bresnick, A.R.; Weber, D.J.; Zimmer, D.B. S100 proteins in cancer. Nat. Rev. Cancer 2015, 15, 96–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, D.B.; Weber, D.J. The Calcium-Dependent Interaction of S100B with Its Protein Targets. Cardiovasc. Psychiatry Neurol. 2010, 2010, 728052. [Google Scholar] [CrossRef]
- Tian, T.; Li, X.; Hua, Z.; Ma, J.; Wu, X.; Liu, Z.; Chen, H.; Cui, Z. S100A7 promotes the migration, invasion and metastasis of human cervical cancer cells through epithelial-mesenchymal transition. Oncotarget 2017, 8, 24964–24977. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; He, Y.; Wang, X.L.; Zhang, Y.X.; Wu, Y.M. Differentially expressed proteins among normal cervix, cervical intraepithelial neoplasia and cervical squamous cell carcinoma. Clin. Transl. Oncol. 2015, 17, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.M.; Fakler, B.; Rivard, A.; Wayman, G.; Johnson-Pais, T.; Keen, J.E.; Ishii, T.; Hirschberg, B.; Bond, C.T.; Lutsenko, S.; et al. Mechanism of calcium gating in small-conductance calcium-activated potassium channels. Nature 1998, 395, 503–507. [Google Scholar] [CrossRef]
- Toyama, K.; Wulff, H.; Chandy, K.G.; Azam, P.; Raman, G.; Saito, T.; Fujiwara, Y.; Mattson, D.L.; Das, S.; Melvin, J.E.; et al. The intermediate-conductance calcium-activated potassium channel KCa3.1 contributes to atherogenesis in mice and humans. J. Clin. Investig. 2008, 118, 3025–3037. [Google Scholar] [CrossRef]
- Pedarzani, P.; Stocker, M. Molecular and cellular basis of small--and intermediate-conductance, calcium-activated potassium channel function in the brain. Cell Mol. Life Sci. 2008, 65, 3196–3217. [Google Scholar] [CrossRef]
- Hua, X.; Deuse, T.; Chen, Y.J.; Wulff, H.; Stubbendorff, M.; Kohler, R.; Miura, H.; Langer, F.; Reichenspurner, H.; Robbins, R.C.; et al. The potassium channel KCa3.1 as new therapeutic target for the prevention of obliterative airway disease. Transplantation 2013, 95, 285–292. [Google Scholar] [CrossRef]
- Kiefer, H.; Blume, A.J.; Kaback, H.R. Membrane potential changes during mitogenic stimulation of mouse spleen lymphocytes. Proc. Natl. Acad. Sci. USA 1980, 77, 2200–2204. [Google Scholar] [CrossRef]
- Strobl, J.S.; Wonderlin, W.F.; Flynn, D.C. Mitogenic signal transduction in human breast cancer cells. Gen. Pharmacol. 1995, 26, 1643–1649. [Google Scholar] [CrossRef]
- Jager, H.; Dreker, T.; Buck, A.; Giehl, K.; Gress, T.; Grissmer, S. Blockage of intermediate-conductance Ca2+-activated K+ channels inhibit human pancreatic cancer cell growth in vitro. Mol. Pharmacol. 2004, 65, 630–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.W.; Liu, J.W.; Zhang, R.C.; Yin, Q.; Shen, W.Z.; Yi, J.L. Inhibitory effects of blockage of intermediate conductance Ca2+-activated K+ channels on proliferation of hepatocellular carcinoma cells. J. Huazhong Univ. Sci. Technol. Med. Sci. 2013, 33, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.H.; Shen, B.; Yao, H.L.; Jia, Y.C.; Ren, J.; Feng, Y.J.; Wang, Y.Z. Blockage of intermediate-conductance-Ca2+-activated K+ channels inhibits progression of human endometrial cancer. Oncogene 2007, 26, 5107–5114. [Google Scholar] [CrossRef] [PubMed]
- Chamlali, M.; Kouba, S.; Rodat-Despoix, L.; Todesca, L.M.; Petho, Z.; Schwab, A.; Ouadid-Ahidouch, H. Orai3 Calcium Channel Regulates Breast Cancer Cell Migration through Calcium-Dependent and -Independent Mechanisms. Cells 2021, 10, 3487. [Google Scholar] [CrossRef]
- Dang, D.; Prasad, H.; Rao, R. Secretory pathway Ca2+-ATPases promote in vitro microcalcifications in breast cancer cells. Mol. Carcinog. 2017, 56, 2474–2485. [Google Scholar] [CrossRef]
- Curry, M.C.; Roberts-Thomson, S.J.; Monteith, G.R. Plasma membrane calcium ATPases and cancer. Biofactors 2011, 37, 132–138. [Google Scholar] [CrossRef]
- Rodrigues, T.; Estevez, G.N.N.; Tersariol, I. Na+/Ca2+ exchangers: Unexploited opportunities for cancer therapy? Biochem. Pharmacol. 2019, 163, 357–361. [Google Scholar] [CrossRef]
- Sun, Y.; Li, M.; Liu, G.; Zhang, X.; Zhi, L.; Zhao, J.; Wang, G. The function of Piezo1 in colon cancer metastasis and its potential regulatory mechanism. J. Cancer Res. Clin. Oncol. 2020, 146, 1139–1152. [Google Scholar] [CrossRef]
- Han, Y.; Liu, C.; Zhang, D.; Men, H.; Huo, L.; Geng, Q.; Wang, S.; Gao, Y.; Zhang, W.; Zhang, Y.; et al. Mechanosensitive ion channel Piezo1 promotes prostate cancer development through the activation of the Akt/mTOR pathway and acceleration of cell cycle. Int. J. Oncol. 2019, 55, 629–644. [Google Scholar] [CrossRef]
- Zhao, Z.; Ji, M.; Wang, Q.; He, N.; Li, Y. Ca2+ signaling modulation using cancer cell membrane coated chitosan nanoparticles to combat multidrug resistance of cancer. Carbohydr. Polym. 2020, 238, 116073. [Google Scholar] [CrossRef]


{kind=link}
| Ca2+ Channels/Pumps | Expression | Effects on Cervical Cancer | References |
|---|---|---|---|
| TRPV1 | Increased | Increased hazard ratio for overall survival, cell viability, and colony formation | [10,11] |
| TRPV6 | Decreased | Lead to poor prognosis | [12] |
| TRPM4 | Increased | Promote cancer cell proliferation | [13,14,15] |
| TRPM7 | Increased | Promote cancer cell proliferation and invasion | [16,17,18] |
| Orai1 and STIM1 | Increased | Cancer cell proliferation, migration, and angiogenesis increase; correlates with prognosis | [19,20] |
| IP3R3/ITPKC | / | Genetic polymorphism is associated with an increased risk of cervical squamous cell carcinoma | [21,22] |
| SERCA2 | Increased | Positive correlation with clinical stage | [23] |
| S100A7, S100A9, S100A11, S100A14 | Increased | Correlation with tumor grade and lymph node metastasis; promotes cancer cell proliferation, migration, and invasion | [24,25] |
| IKCa1 | Increased | Positively correlated with malignancy, promoting dedifferentiation and cancer cell proliferation | [26] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, J.; Deng, F.; Ding, H.; Fu, M.; Xu, T.; Ji, B.; Feng, L.; Li, M.; Qiu, J.; Gao, Q. Recent Developments on the Roles of Calcium Signals and Potential Therapy Targets in Cervical Cancer. Cells 2022, 11, 3003. https://doi.org/10.3390/cells11193003
Lei J, Deng F, Ding H, Fu M, Xu T, Ji B, Feng L, Li M, Qiu J, Gao Q. Recent Developments on the Roles of Calcium Signals and Potential Therapy Targets in Cervical Cancer. Cells. 2022; 11(19):3003. https://doi.org/10.3390/cells11193003
Chicago/Turabian StyleLei, Jiahui, Fengying Deng, Hongmei Ding, Mengyu Fu, Ting Xu, Bingyu Ji, Lin Feng, Min Li, Junlan Qiu, and Qinqin Gao. 2022. "Recent Developments on the Roles of Calcium Signals and Potential Therapy Targets in Cervical Cancer" Cells 11, no. 19: 3003. https://doi.org/10.3390/cells11193003
APA StyleLei, J., Deng, F., Ding, H., Fu, M., Xu, T., Ji, B., Feng, L., Li, M., Qiu, J., & Gao, Q. (2022). Recent Developments on the Roles of Calcium Signals and Potential Therapy Targets in Cervical Cancer. Cells, 11(19), 3003. https://doi.org/10.3390/cells11193003