

Altered Cord Blood Lipid Concentrations Correlate with Birth Weight and Doppler Velocimetry of Fetal Vessels in Human Fetal Growth Restriction Pregnancies

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects and Antenatal Assessments
2.2. Tissue Collection and Processing
2.3. Postnatal Assessments and Case–Control Classifications
2.4. Lipid Extraction of Plasma and Placenta
2.4.1. Triglyceride Analysis
2.4.2. Phospholipid Analysis
2.4.3. Eicosanoid Extraction and Analysis
2.5. Lipid Nomenclature
2.6. Data Presentation and Statistical Analysis
3. Results
3.1. Clinical Characteristics
3.2. Fatty Acid Composition
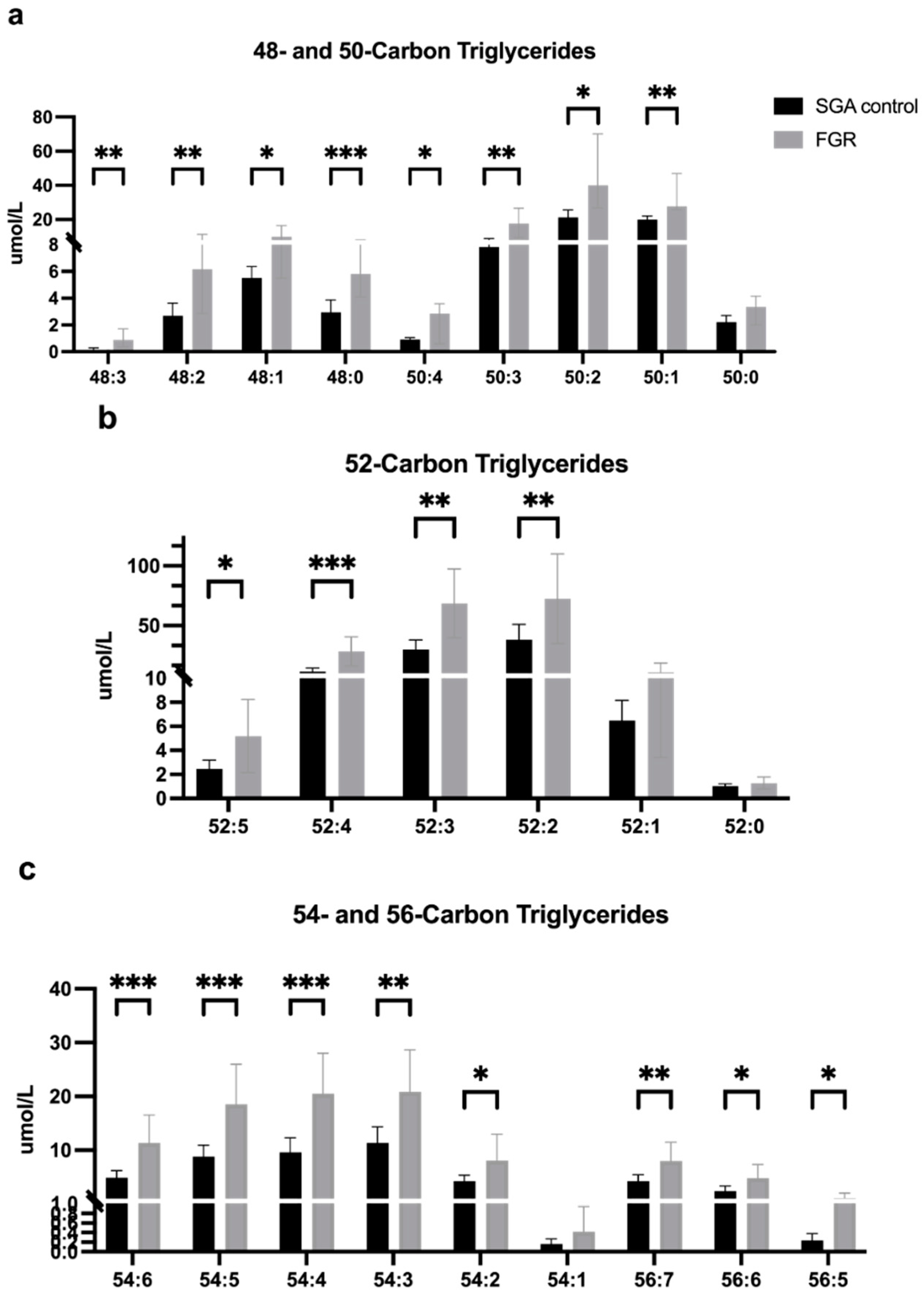
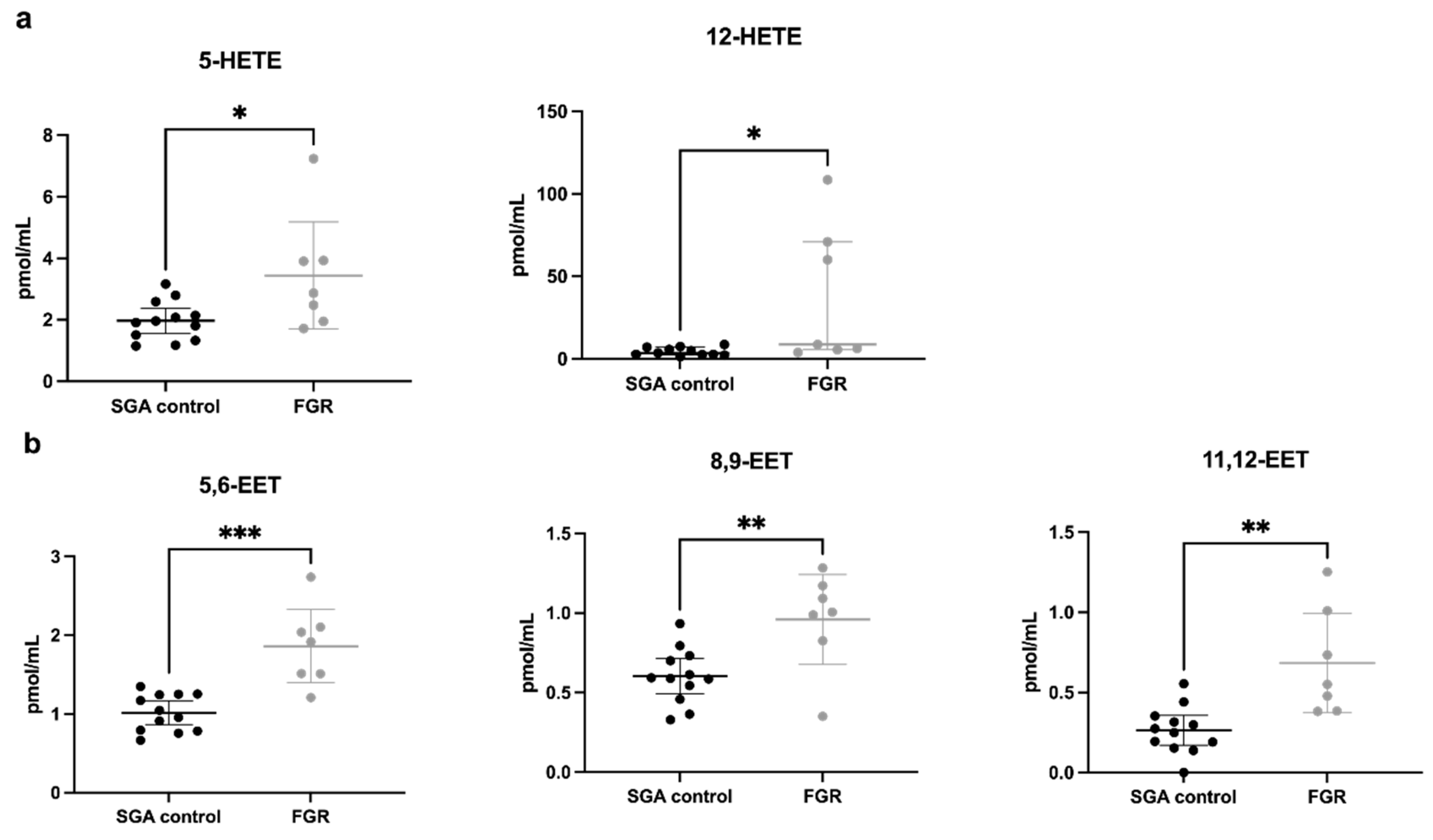
3.2.1. Fatty Acid and Lipid Concentrations in Umbilical Vein Plasma
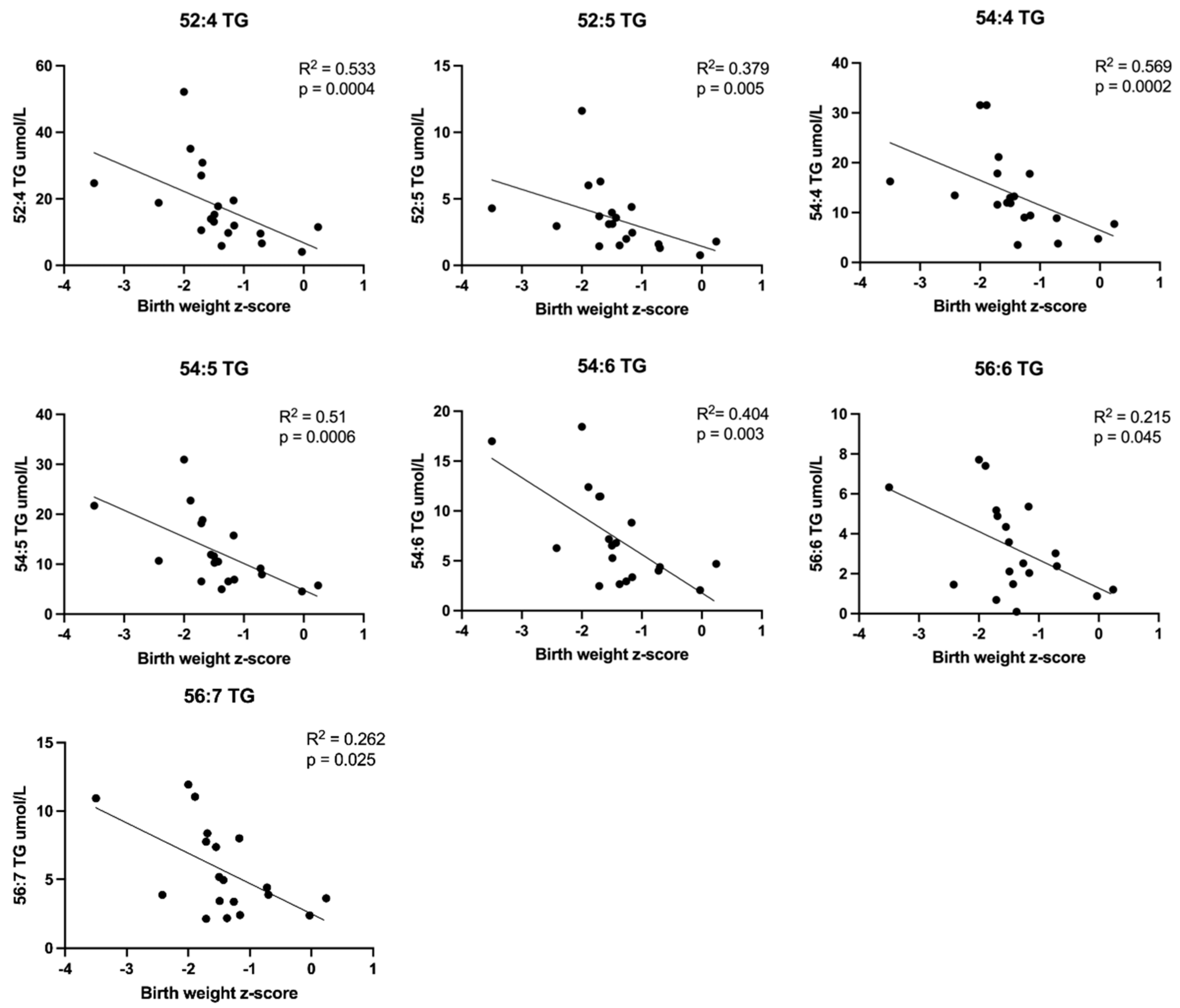
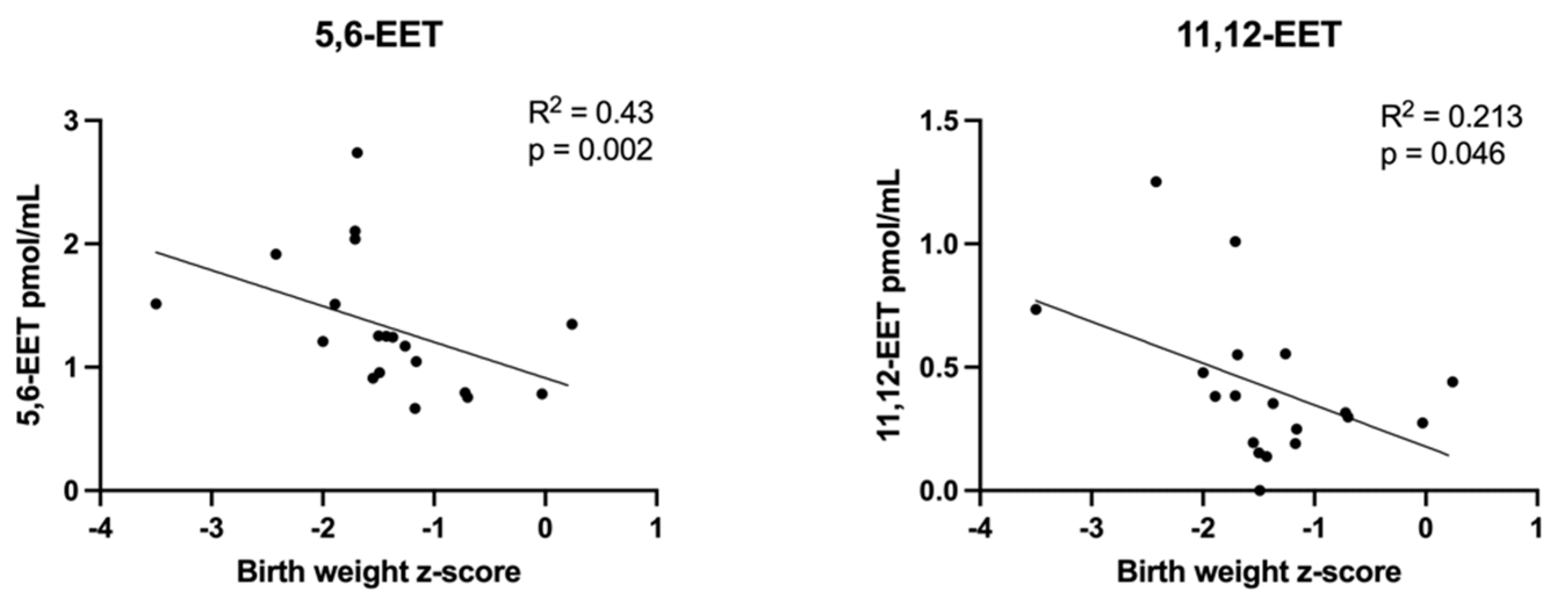
3.2.2. Correlations between Plasma Lipid Concentrations and Birth Weight z-Scores
3.2.3. Correlations between Plasma Lipid Concentrations and Doppler Indices
3.2.4. Fatty Acid and Lipid Concentrations in Placenta Homogenate
3.2.5. Correlations between Placenta Lipid Concentrations and Clinical Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, D.; Shastri, S.; Sharma, P. Intrauterine growth restriction: Antenatal and postnatal aspects. Clin. Med. Insights Pediatr. 2016, 10, 67–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, D.P. In Utero Programming of Chronic Disease. Clin. Sci. 1998, 95, 115–128. [Google Scholar] [CrossRef]
- Hay, W.W., Jr.; Thureen, P.J.; Anderson, M.S. Intrauterine growth restriction. Neoreviews 2001, 2, e129–e138. [Google Scholar] [CrossRef]
- Scifres, C.M.; Nelson, D.M. Intrauterine growth restriction, human placental development and trophoblast cell death. J. Physiol. 2009, 587, 3453–3458. [Google Scholar] [CrossRef] [PubMed]
- Maulik, D.; Frances Evand, J.; Ragolia, L. Fetal growth restriction: Pathogenic mechanisms. Clin. Obstet. Gynecol. 2006, 49, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Giles, W.B.; Trudinger, B.J.; Baird, P.J. Fetal umbilical artery flow velocity waveforms and placental resistance: Pathological correlation. Br. J. Obstet. Gynaecol. 1985, 92, 31–38. [Google Scholar] [CrossRef]
- Trudinger, B.J.; Giles, W.B.; Cook, C.M.; Bombardieri, J.; Collins, L. Fetal umbilical artery flow velocity waveforms and placental resistance: Clinical significance. Br. J. Obstet. Gynaecol. 1985, 92, 23–30. [Google Scholar] [CrossRef]
- Trudinger, B.J.; Cook, C.M.; Giles, W.B.; Ng, S.; Fong, E.; Connelly, A.; Wilcox, W. Fetal umbilical artery velocity waveforms and subsequent neonatal outcome. Br. J. Obstet. Gynaecol. 1991, 98, 378–384. [Google Scholar] [CrossRef]
- Olsen, R.N.; Shepherd, J.; Katheria, A. Postnatal systemic blood flow in neonates with abnormal fetal umbilical artery doppler. ISRN Obstet. Gynecol. 2014, 2014, 957180. [Google Scholar] [CrossRef]
- Kamitomo, M.; Alonso, J.G.; Okai, T.; Longo, L.D.; Gilbert, R.D. Effects of long-term, high-altitude hypoxemia on ovine fetal cardiac output and blood flow distribution. Am. J. Obstet. Gynecol. 1993, 169, 701–707. [Google Scholar] [CrossRef]
- Richardson, B.S.; Bocking, A.D. Metabolic and circulatory adaptations to chronic hypoxia in the fetus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1998, 119, 717–723. [Google Scholar] [CrossRef]
- Morrison, J.L. Sheep models of intrauterine growth restriction: Fetal adaptations and consequences. Clin. Exp. Pharmacol. Physiol. 2008, 35, 730–743. [Google Scholar] [CrossRef] [PubMed]
- Poudel, R.; McMillen, I.C.; Dunn, S.L.; Zhang, S.; Morrison, J.L. Impact of chronic hypoxemia on blood flow to the brain, heart, and adrenal gland in the late-gestation IUGR sheep fetus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R151–R162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allison, B.J.; Brain, K.L.; Niu, Y.; Kane, A.D.; Herrera, E.A.; Thakor, A.S.; Botting, K.J.; Cross, C.M.; Itani, N.; Skeffington, K.L.; et al. Fetal in vivo continuous cardiovascular function during chronic hypoxia. J. Physiol. 2016, 594, 1247–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gramellini, D.; Folli, M.C.; Raboni, S.; Vadora, E.; Merialdi, A. Cerebral-umbilical Doppler ratio as a predictor of adverse perinatal outcome. Obstet. Gynecol. 1992, 79, 416–420. [Google Scholar] [CrossRef]
- Gordijn, S.J.; Beune, I.M.; Thilaganathan, B.; Papageorghiou, A.; Baschat, A.A.; Baker, P.N.; Silver, F.M.; Wynia, K.; Ganzevoort, W. Consensus definition of fetal growth restriction: A Delphi procedure. Ultrasound. Obstet. Gynecol. 2016, 48, 333–339. [Google Scholar] [CrossRef]
- Savchev, S.; Figueras, F.; Sanz-Cortes, M.; Cruz-Lemini, M.; Triunfo, S.; Botet, F.; Gratacos, E. Evaluation of an optimal gestational age cut-off for the definition of early- and late-onset fetal growth restriction. Fetal. Diagn. Ther. 2014, 36, 99–105. [Google Scholar] [CrossRef]
- Glazier, J.D.; Cetin, I.; Perugino, G.; Ronzoni, S.; Grey, A.M.; Mahendran, D.; Marconi, A.M.; Pardi, G.; Sibley, C.P. Association between the activity of the system A amino acid transport in the microvillous plasma membrane of the human placenta and severity of fetal compromise in intrauterine growth restriction. Pediatr. Res. 1997, 42, 514–519. [Google Scholar] [CrossRef]
- Jansson, T.; Ylven, K.; Wennergren, M.; Powell, T.L. Glucose transport and system A activity in syncytiotrophoblast microvillous and basal plasma membranes in intrauterine growth restriction. Placenta 2002, 23, 392–399. [Google Scholar] [CrossRef]
- Mahendran, D.; Donnai, P.; Glazier, J.D.; D’Souza, S.W.; Boyd, R.D.H.; Sibley, C.P. Amino acid (System A) transporter activity in microvillous membrane vesicles from the placentas of appropriate and small for gestational age babies. Pediatr. Res. 1993, 34, 661–665. [Google Scholar] [CrossRef]
- Johansson, M.; Jansson, T.; Glazier, J.D.; Powell, T.L. Activity and expression of the Na+/H+ exchanger is reduced in syncytiotrophoblast microvillous plasma membranes isolated from preterm intrauterine growth restriction pregnancies. J. Clin. Endocrinol. Metab. 2002, 87, 5686–5694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, M.; Karlsson, L.; Wennergren, M.; Jansson, T.; Powell, T.L. Activity and protein expression of Na+/K+ ATPase are reduced in microvillous syncytiotrophoblast plasma membranes isolated from pregnancies complicated by intrauterine growth restriction. J. Clin. Endocrinol. Metab. 2003, 88, 2831–2837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansson, T.; Wennergren, M.; Illsley, N.P. Glucose transporter protein expression in human placenta throughout gestation and in intrauterine growth restriction. J. Clin. Endocrinol. Metab. 1993, 77, 1554–1562. [Google Scholar] [PubMed]
- Strid, H.; Bucht, E.; Jansson, T.; Wennergren, M.; Powell, T.L. ATP dependent Ca2+ transport across basal membrane of human syncytiotrophoblast in pregnancies complicated by intrauterine growth restriction or diabetes. Placenta 2003, 24, 445–452. [Google Scholar] [CrossRef]
- Chassen, S.S.; Ferchaud-Roucher, V.; Gupta, M.B.; Jansson, T.; Powell, T.L. Alterations in placental long chain polyunsaturated fatty acid metabolism in human intrauterine growth restriction. Clin. Sci. 2018, 132, 595–607. [Google Scholar] [CrossRef]
- Haggarty, P. Fatty acid supply to the human fetus. Annu. Rev. Nutr. 2010, 30, 237–255. [Google Scholar] [CrossRef]
- Larciprete, G.; Valensise, H.; Di Pierro, G.; Vasapollo, B.; Casalino, B.; Arduini, D.; Jarvis, S.; Cirese, E. Intrauterine growth restriction and fetal body composition. Ultrasound Obstet. Gynecol. 2005, 26, 258–262. [Google Scholar] [CrossRef]
- Steller, J.G.; Gumina, D.; Driver, C.; Palmer, C.; Brown, L.D.; Reeves, S.; Hobbins, J.C.; Galan, H.L. 3D fractional limb volume identifies reduced subcutaneous and lean mass in fetal growth restriction. J. Ultrasound Med. 2022, 41, 1623–1632. [Google Scholar] [CrossRef]
- Haggarty, P.; Page, K.; Abramovich, D.R.; Ashton, J.; Brown, D. Long-chain polyunsaturated fatty acid transport across the perfused human placenta. Placenta 1997, 18, 635–642. [Google Scholar] [CrossRef]
- Hadlock, F.P.; Harrist, R.B.; Martinez-Poyer, J. In utero analysis of fetal growth: A sonographic weight standard. Radiology 1991, 181, 129–133. [Google Scholar] [CrossRef]
- Arbeille, P.H.; Roncin, A.; Berson, M.; Patat, F.; Pourcelot, L. Exploration of the fetal cerebral blood flow by duplex doppler- linear array system in normal and pathological pregnancies. Ultrasound Med. Biol. 1987, 13, 329–337. [Google Scholar] [CrossRef]
- Fenton, T.R.; Kim, J.H. A systematic review and meta-analysis to revise the Fenton growth chart for preterm infants. BMC Pediatrics 2013, 13, 59–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid. Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krank, J.; Murphy, R.C.; Barkley, R.M.; Duchoslav, E.; McAnoy, A. Qualitative analysis and quantitative assessment of changes in neutral glycerol lipid molecular species within cells. Methods. Enzymol. 2007, 432, 1–20. [Google Scholar]
- Zarini, S.; Gijon, M.A.; Ransome, A.E.; Murphy, R.C.; Sala, A. Transcellular biosynthesis of cysteinyl leukotrienes in vivo during mouse peritoneal inflammation. Proc. Natl. Acad. Sci. USA 2009, 106, 8296–8301. [Google Scholar] [CrossRef] [Green Version]
- Liebisch, G.; Fahy, E.; Aoki, J.; Dennis, E.A.; Durand, T.; Eising, C.S.; Fedorova, M.; Feussner, I.; Griffiths, W.J.; Kofeler, H.; et al. Update on LIPID MAPS classification, nomenclature, and shorthand notation for MS-derived lipid structures. J. Lipid. Res. 2020, 61, 1539–1555. [Google Scholar] [CrossRef]
- Madden, J.V.; Flatley, C.J.; Kumar, S. Term small-for-gestational age infants from low-risk women are at significantly greater risk of adverse neonatal outcomes. Am. J. Obstet. Gynecol. 2018, 196, 499–507. [Google Scholar] [CrossRef] [Green Version]
- McCowan, L.M.; Figueras, F.; Anderson, N.H. Evidence-based national guidelines for the management of suspected fetal growth restriction: Comparison, consensus, and controversy. Am. J. Obstet. Gyencol. 2018, 281, 855–868. [Google Scholar] [CrossRef] [Green Version]
- Ego, A.; Subtil, D.; Grange, G.; Thiebaugeorges, O.; Senat, M.V.; Vayssiere, C.; Zeitlin, J. Customized versus population-based birth weight standards for identifying growth restricted infants: A French multicenter study. Am. J. Obstet. Gynecol. 2006, 194, 1042–1049. [Google Scholar] [CrossRef]
- Getahun, D.; Ananth, C.V.; Kinzler, W.L. Risk factors for antepartum and intrapartum stillbirth: A population-based study. Am. J. Obstet. Gynecol. 2007, 196, 499–507. [Google Scholar] [CrossRef]
- Chassen, S.S.; Ferchaud-Roucher, V.; Palmer, C.; Li, C.; Jansson, T.; Nathanielsz, P.W.; Powell, T.L. Placental fatty acid transport across late gestation in a baboon model of intrauterine growth restriction. J. Physiol. 2020, 598, 2469–2489. [Google Scholar] [CrossRef] [PubMed]
- Mylonis, I.; Sembongi, H.; Befani, C.; Liakos, P.; Siniossoglou, S.; Gimos, G. Hypoxia causes triglyceride accumulation by HIF-1-mediated stimulation of lipin1 expression. J. Cell. Sci. 2012, 125, 3485–3493. [Google Scholar] [PubMed] [Green Version]
- Kim, W.Y.; Safran, M.; Buckley, M.R.M.; Ebert, B.L.; Glickman, J.; Bosenberg, M.; Regan, M.; Kaelin, W.G., Jr. Failure to prolyl hydroxylate hypoxia-inducible factor alpha phenocopies VHL inactivation in vivo. EMBO J. 2006, 25, 4650–4662. [Google Scholar] [CrossRef] [PubMed]
- Nath, B.; Levin, I.; Csak, T.; Petrasek, J.; Mueller, C.; Kodys, K.; Catalano, D.; Mandrekar, P.; Szabo, G. Hepatocyte-specific hypoxia-inducible factor-1a is a determinant of lipid accumulation and liver injury in alcohol-induced steatosis in mice. Hepatology 2011, 53, 1526–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bostrom, P.; Magnusson, B.; Svensson, P.-A.; Wiklund, O.; Boren, J.; Carlsson, L.M.S.; Stahlman, M.; Olofsson, S.-O.; Hulten, L.M. Hypoxia converts human macrophages into triglyceride-loaded foam cells. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1871–1876. [Google Scholar] [CrossRef] [Green Version]
- Patterson, T.M.; Parker, C.R., Jr. Intrapartum stress and lipid status of term infants: Relation to fetal adrenal production of dehydroepiandrosterone sulphate. J. Matern. Fetal Neonatal. Med. 2008, 21, 783–787. [Google Scholar] [CrossRef]
- Tsang, R.; Glueck, C.J.; Evans, G.; Steiner, P.M. Cord blood hypertriglyceridemia. Am. J. Dis. Child. 1974, 127, 78–82. [Google Scholar] [CrossRef]
- Anderson, G.E.; Friis-Hansen, B. Neonatal hypertriglyceridemia: A new index of antepartum-intrapartum fetal stress. Acta. Paediatr. Scand. 1976, 65, 369–374. [Google Scholar] [CrossRef]
- Gil-Sanchez, A.; Larque, E.; Demmelmair, H.; Acien, M.I.; Faber, F.L.; Parrilla, J.J.; Koletzko, B. Maternal-fetal in vivo transfer of [13C] docosahexaenoic and other fatty acids across the human placenta 12 h after maternal oral intake. Am. J. Clin. Nutr. 2010, 92, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Lagarde, M.; Bernoud, N.; Brossard, N.; Lemaitre-Delaunay, D.; Thies, F.; Croset, M.; Lecerf, J. Lysophosphatidylcholine as a preferred carrier form of docosahexaenoic acid to the brain. J. Mol. Neurosci. 2001, 16, 201–204. [Google Scholar] [CrossRef]
- Thies, F.; Pillon, C.; Moliere, P.; Lagarde, M.; Lecerf, J. Preferential incorporation of sn-2 lysoPC DHA over unesterified DHA in the young rat brain. Am. J. Physiol 1994, 267, R1273–R1279. [Google Scholar] [CrossRef] [PubMed]
- Sugasini, D.; Thomas, R.; Yalagala, P.C.R.; Tai, L.M.; Subbaiah, P.V. Dietary docosahexaenoic acid (DHA) as lysophosphatidylcholinne, but not as free acid, enriches brain DHA and improves memory in adult mice. Sci. Rep. 2017, 7, 11263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferchaud-Roucher, V.; Kramer, A.; Silva, E.; Pantham, P.; Weintraub, S.T.; Jansson, T.; Powell, T.L. A potential role for lysophosphatidylcholine in the delivery of long chain polyunsaturated fatty acids to the fetal circulation. Biochim. Biophys. Acta. Mol. Cell. Biol. Lipids 2019, 1864, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.E.; Bazan, N.G. Changing fatty acid content of growth cone lipids prior to synaptogenesis. J. Neurochem. 1992, 59, 318–325. [Google Scholar] [CrossRef]
- Kim, H.Y.; Edsall, L.; Ma, Y.C. Specificity of polyunsaturated fatty acid release from rat brain synaptosomes. Lipids 1996, 31, S229–S233. [Google Scholar] [CrossRef]
- Barcelo-Coblijn, G.; Kitajka, K.; Puskas, L.G.; Hogyes, E.; Zvara, A.; Hackler, L., Jr.; Farkas, T. Gene expression and molecular composition of phospholipids in rat brain in relation to dietary n-6 to n-3 fatty acid ratio. BBA 2003, 1632, 72–79. [Google Scholar] [CrossRef]
- Kitajka, K.; Sinclair, A.J.; Weisinger, R.S.; Weisinger, H.S.; Mathai, M.; Jayasooriya, A.P.; Halver, J.E.; Puskas, L.G. Effects of dietary omega-3 polyunsaturated fatty acids on brain gene expression. Proc. Natl. Acad. Sci. USA 2004, 101, 10931–10936. [Google Scholar] [CrossRef] [Green Version]
- Crawford, M.A.; Costeloe, K.; Ghebremeskel, K.; Phylactos, A.; Skirvin, L.; Stacey, F. Are deficits of arachidonic and docosahexaenoic acids responsible for the neural and vascular complications of preterm babies? Am. J. Clin. Nutr. 1997, 66, 1032S–1041S. [Google Scholar] [CrossRef] [Green Version]
- Janssen, C.I.F.; Kiliaan, A.J. Long-chain polyunsaturated fatty acids (LCPUFA) from genesis to senescence: The influence of LCPUFA on neural development, aging, and neurodegeneration. Prog. Lipid. Res. 2014, 53, 1–17. [Google Scholar] [CrossRef]
- Dobbing, J.; Sands, J. Quantitative growth and development of human brain. Arch. Dis. Child. 1973, 48, 757–767. [Google Scholar] [CrossRef] [Green Version]
- Martinez, M. Tissue levels of polyunsaturated fatty acids during early human development. J. Pediatr. 1992, 120, S129–S138. [Google Scholar] [CrossRef]
- Esteban, F.J.; Padilla, N.; Sanz-Cortes, M.; de Miras, J.R.; Bargallo, N.; Villoslada, P.; Gratacos, E. Fractal-dimension analysis detects cerebral changes in preterm infants with and without intrauterine growth restriction. Neuroimage 2010, 53, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Padilla, N.; Falcon, C.; Sanz-Cortes, M.; Figueras, F.; Bargallo, N.; Crispi, F.; Eixarch, E.; Arranz, A.; Botet, F.; Gratacos, E. Differential effects of intrauterine growth restriction on brain structure and development in preterm infants: A magnetic resonance imaging study. Brain Res. 2011, 1382, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Padilla, N.; Junque, C.; Figueras, F.; Sanz-Cortes, M.; Bargallo, N.; Arranz, A.; Donaire, A.; Figueras, J.; Gratacos, E. Differential vulnerability of gray matter and white matter to intrauterine growth restriction in preterm infants at 12 months corrected age. Brain Res. 2014, 1545, 1–22. [Google Scholar] [CrossRef]
- Batalle, D.; Eixarch, E.; Figueras, F.; Munoz-Moreno, E.; Bargallo, N.; Illa, M.; Acosta-Rojas, R.; Amat-Roldan, I.; Gratacos, E. Altered small-world topology of structural brain networks in infants with intrauterine growth restriction and its association with later neurodevelopmental outcome. Neuroimage 2012, 60, 1352–1366. [Google Scholar] [CrossRef]
- Munoz-Moreno, E.; Fischi-Gomez, E.; Batalle, D.; Borradori-Tolsa, C.; Eixarch, E.; Thiran, J.P.; Gratacos, E.; Huppi, P.S. Structural brain network reorganization and social cognition related to adverse perinatal condition from infancy to early adolescence. Front. Neurosci. 2016, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.I.; Huppi, P.S.; Mallard, C. The consequences of fetal growth restriction on brain structure and neurodevelopmental outcome. J. Physiol. 2016, 594, 807–823. [Google Scholar] [CrossRef] [Green Version]
- Welin, A.K.; Svedin, P.; Lapatto, R.; Sultan, B.; Hagberg, H.; Gressens, P.; Kjellmer, I.; Mallard, C. Melatonin reduces inflammation and cell death in white matter in the mid-gestation fetal sheep following umbilical cord occlusion. Pediatr. Res. 2007, 61, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Burke, C.; Gobe, G. Pontosubicular apoptosis (“necrosis”) in human neonates with intrauterine growth retardation and placental infarction. Virchows. Arch. 2005, 446, 640–645. [Google Scholar] [CrossRef]
- Ramenghi, L.A.; De Carli, A.; Fumagalli, M.; Mosca, F.; Martinelli, A.; Mandia, L.; Fumagalli, M.; Triulzi, F.; Mosca, F.; Cetin, I. Cerebral maturation in IUGR and appropriate for gestational age preterm babies. Reprod. Sci. 2011, 18, 469–475. [Google Scholar] [CrossRef]
- Businelli, C.; De Wit, C.; Visser, G.H.A.; Pistorius, I.R. Ultrasound evaluation of cortical brain development in fetuses with intrauterine growth restriction. J. Matern. Neonatal. Med. 2015, 28, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Dudink, I.; Huppi, P.S.; Sizonenko, S.V.; Castillo-Melendez, M.; Sutherland, A.E.; Allison, B.J.; Miller, S.L. Altered trajectory of neurodevelopment associated with fetal growth restriction. Exp. Neurol. 2022, 347, 113885. [Google Scholar] [CrossRef] [PubMed]
- Morsing, E.; Malova, M.; Kahn, A.; Latt, J.; Bjorkman-Burtscher, I.M.; Marsal, K.; Ley, D. Brain volumes and developmental outcome in childhood following fetal growth restriction leading to very preterm birth. Front. Physiol. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes. Nutrients 2010, 2, 355–374. [Google Scholar] [CrossRef] [Green Version]
- Biri, A.; Bozkurt, N.; Turp, A.; Kavutcu, M.; Himmetoglu, O.; Durak, I. Role of oxidative stress in intrauterine growth restriction. Gynecol. Obstet. Invest. 2007, 64, 187–192. [Google Scholar] [CrossRef]
- Schoots, M.H.; Gordijn, S.J.; Scherjon, S.A.; van Goor, H.; Hillebrands, J.L. Oxidative stress in placental pathology. Placenta 2018, 69, 153–161. [Google Scholar] [CrossRef]
- Reyes-Hernandez, C.G.; Ramiro-Cortijo, D.; Rodriguez-Rodrigeuz, P.; Giambelluca, S.; Simonato, M.; Gonzalez, M.C.; Lopez de Pablo, A.L.; Lopez-Gimenez, M.R.; Cogo, P.; Saenz de Pipaon, M.; et al. Effects of arachidonic and docosahexaenoic acid supplementation during gestation in rats. Implication of placental oxidative stress. Int. J. Mol. Sci. 2018, 19, 3863. [Google Scholar] [CrossRef] [Green Version]
- Campbell, W.B.; Harder, D.R. Endothelium-derived hyperpolarizing factors and vascular cytochrome P450 metabolites of arachidonic acid in the regulation of tone. Circ. Res. 1999, 84, 484–488. [Google Scholar] [CrossRef] [Green Version]
- Roman, R.J. P-450 metabolites of arachidonic acid in the control of cardiovascular function. Physiol. Rev. 2002, 82, 131–185. [Google Scholar] [CrossRef] [Green Version]
- Larsen, B.T.; Gutterman, D.D.; Hatoum, O.A. Emerging role of epoxyeicosatrienoic acids in coronary vascular function. Eur. J. Clin. Invest. 2006, 36, 293–300. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y.; Schmelzer, K.; Lee, T.S.; Fang, X.; Zhu, Y.; Spector, A.A.; Gill, S.; Morisseau, C.; Hammock, B.D.; et al. The anti-inflammatory effect of laminar flow: The role of PPARγ, epoxyeicosatrienoic acids, and soluble epoxide hydrolase. Proc. Natl. Acad. Sci. USA 2005, 102, 16747–16752. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.M.; Lodhi, I.J. Structural and functional roles of ether lipids. Protein Cell. 2018, 9, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Clifton, V.L. Review: Sex and the human placenta: Mediating differential strategies of fetal growth and survival. Placenta 2010, 31, S33–S39. [Google Scholar] [CrossRef]
- Grigore, D.; Ojeda, N.B.; Alexander, B.T. Sex differences in the fetal programming of hypertension. Gend. Med. 2008, 5, S121–S132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, J.C. Sexual dimorphism of body composition. Best Pract. Res. Clin. Endocrinol. 2007, 21, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Childs, C.E.; Romeu-Nadal, M.; Burdge, G.C.; Calder, P.C. Gender differences in the n-3 fatty acid content of tissues. Proc. Nutr. Soc. 2008, 67, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Sedlmeier, E.M.; Brunner, S.; Much, D.; Pagel, P.; Ulbrich, S.E.; Meyer, H.H.D.; Amann-Gassner, U.; Hauner, H.; Bader, B.L. Human placental transcriptome shows sexually dimorphic gene expression and responsiveness to maternal dietary n-3 long-chain polyunsaturated fatty acid intervention during pregnancy. BMC Genomics 2014, 15, 941. [Google Scholar] [CrossRef] [Green Version]
- Morales-Rosello, J.; Khalil, A.; Morlando, M.; Papageorghiou, A.; Bhide, A.; Thilaganathan, B. Changes in fetal doppler indices as a marker of failure to reach growth potential at term. Ultrasound Obstet. Gynecol. 2014, 43, 303–310. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | SGA Controls (n = 12) | FGR Cases (n = 8) | P Value | ||
|---|---|---|---|---|---|
| Mean/Median | 95% CI/IQR | Mean/Median | 95% CI/IQR | ||
| Maternal, n (%) | |||||
| Age (years) | 29.25 | [25.3,33.2] | 27.13 | [23.1,31.1] | 0.426 |
| Race | 0.495 | ||||
| White | 10 (83%) | 8 (100%) | |||
| Not reported | 2 (17%) | ||||
| Chronic hypertension | >0.999 | ||||
| Yes | 1 (8%) | 1 (12%) | |||
| No | 11 (92%) | 7 (88%) | |||
| PIH | |||||
| No | 12 (100%) | 8 (100%) | |||
| Preeclampsia | >0.999 | ||||
| Yes | 1 (8%) | 0 | |||
| No | 11 (92%) | 8 (100%) | |||
| Mode of delivery | 0.255 | ||||
| Vaginal | 11 (92%) | 5 (62% | |||
| C-section | 1 (8%) | 3 (38%) | |||
| Prenatal, n (%) | |||||
| GA at recruitment | >0.999 | ||||
| <32 weeks | 5 (42%) | 4 (50%) | |||
| 7 (58%) | 4 (50%) | ||||
| Ultrasound measurements * | |||||
| GA at ultrasound (weeks) | 36.57 | [36.1,37.1] | 34.11 | [31.1,37.1] | 0.032 |
| EFW (grams) | 2403 | [2263,2543] | 1713 | [1121,2305] | 0.005 |
| EFW percentile (%) | 10.5 | [16.5] | 1.5 | [3.5] | 0.008 |
| AC (cm) | 31.18 | [2.03] | 26.84 | [7.2] | 0.069 |
| AC percentile (%) | 24.83 | [10.2,39.5] | 2.5 | [0.7,4.3] | 0.015 |
| UA PI | 0.87 | [0.8,0.9] | 1.16 | [0.8,1.5] | 0.019 |
| UA PI percentile (%) | 59.5 | [47.5,71.5] | 78.67 | [60.8,96.6] | 0.049 |
| MCA PI | 1.53 | [1.3,1.8] | 1.45 | [1.2,1.7] | 0.641 |
| MCA PI percentile (%) | 16.15 | [34.1] | 11.23 | [25.6] | 0.384 |
| CPR | 1.76 | [1.5,2] | 1.35 | [0.9,1.8] | 0.060 |
| CPR percentile (%) | 34.46 | [42.6] | 4.61 | [37.1] | 0.083 |
| Birth | |||||
| Infant sex, n (%) | >0.999 | ||||
| Male | 4 (33%) | 2 (25%) | |||
| Female | 8 (67%) | 6 (75%) | |||
| GA at delivery (weeks) | 38 | [37.4,38.7] | 36.52 | [34.8,38.2] | 0.038 |
| Placenta weight (grams) | 535.30 | [472.9,597.7] | 374.40 | [242.4,506.4] | 0.01 |
| Birth weight (grams) | 2590 | [467] | 2120 | [600] | 0.0002 |
| Birth weight percentile (%) | 11 | [16.8] | 3.5 | [2.8] | <0.0001 |
| Birth length (cm) | 46.97 | [46.4,47.6] | 42.54 | [38,47.1] | 0.012 |
| Birth length percentile (%) | 21 | [20] | 7.5 | [12.3] | 0.02 |
| Birth HC (cm) | 32.82 | [31.9,33.8] | 31.64 | [29.9,33.4] | 0.143 |
| Birth HC percentile (%) | 15.50 | [50.3] | 12 | [40] | 0.521 |
| HC:BW ratio | 12.73 | [2.5] | 14.88 | [1.5] | 0.0001 |
| Z–scores | |||||
| Birth weight | −1.215 | [0.8] | −1.80 | [0.6] | <0.0001 |
| Birth length | −0.79 | [−1.2,−0.4] | −1.80 | [−2.9,−0.8] | 0.027 |
| Birth HC | −0.55 | [−1.1,−0.02] | −0.97 | [−1.7,−0.2] | 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chassen, S.S.; Zemski-Berry, K.; Raymond-Whish, S.; Driver, C.; Hobbins, J.C.; Powell, T.L. Altered Cord Blood Lipid Concentrations Correlate with Birth Weight and Doppler Velocimetry of Fetal Vessels in Human Fetal Growth Restriction Pregnancies. Cells 2022, 11, 3110. https://doi.org/10.3390/cells11193110
Chassen SS, Zemski-Berry K, Raymond-Whish S, Driver C, Hobbins JC, Powell TL. Altered Cord Blood Lipid Concentrations Correlate with Birth Weight and Doppler Velocimetry of Fetal Vessels in Human Fetal Growth Restriction Pregnancies. Cells. 2022; 11(19):3110. https://doi.org/10.3390/cells11193110
Chicago/Turabian StyleChassen, Stephanie S., Karin Zemski-Berry, Stefanie Raymond-Whish, Camille Driver, John C. Hobbins, and Theresa L. Powell. 2022. "Altered Cord Blood Lipid Concentrations Correlate with Birth Weight and Doppler Velocimetry of Fetal Vessels in Human Fetal Growth Restriction Pregnancies" Cells 11, no. 19: 3110. https://doi.org/10.3390/cells11193110
APA StyleChassen, S. S., Zemski-Berry, K., Raymond-Whish, S., Driver, C., Hobbins, J. C., & Powell, T. L. (2022). Altered Cord Blood Lipid Concentrations Correlate with Birth Weight and Doppler Velocimetry of Fetal Vessels in Human Fetal Growth Restriction Pregnancies. Cells, 11(19), 3110. https://doi.org/10.3390/cells11193110