

Immunomodulatory Effect and Bone Homeostasis Regulation in Osteoblasts Differentiated from hADMSCs via the PD-1/PD-L1 Axis


Abstract
:1. Introduction
2. Materials and Methods
2.1. hADMSCs Culture and Adipogenic or Osteogenic Differentiation
2.2. Oil Red O (ORO) Staining
2.3. Alizarin Red S (ARS) Staining
2.4. RNA Extraction and Real-Time PCR
2.5. Immunofluorescence Staining
2.6. Western Blot Analysis
2.7. Exosome Isolation
2.8. THP-1 Cell Culture and Osteoclasts Differentiation
2.9. Coculture of hADMSCs with Differentiated Cells
2.10. Establishment of In Vitro Bone Matrix System with Osteoblasts and Osteoclasts
2.11. Treatment of PD-L1 and hADMSCs Derived Exosomes
2.12. Inhibition of the PD-1/PD-L1 Pathway
2.13. Enzyme-Linked Immunosorbent Assay (ELISA)
2.14. Nano Particle Tracking Analysis (NTA)
2.15. Statistical Analysis
3. Results
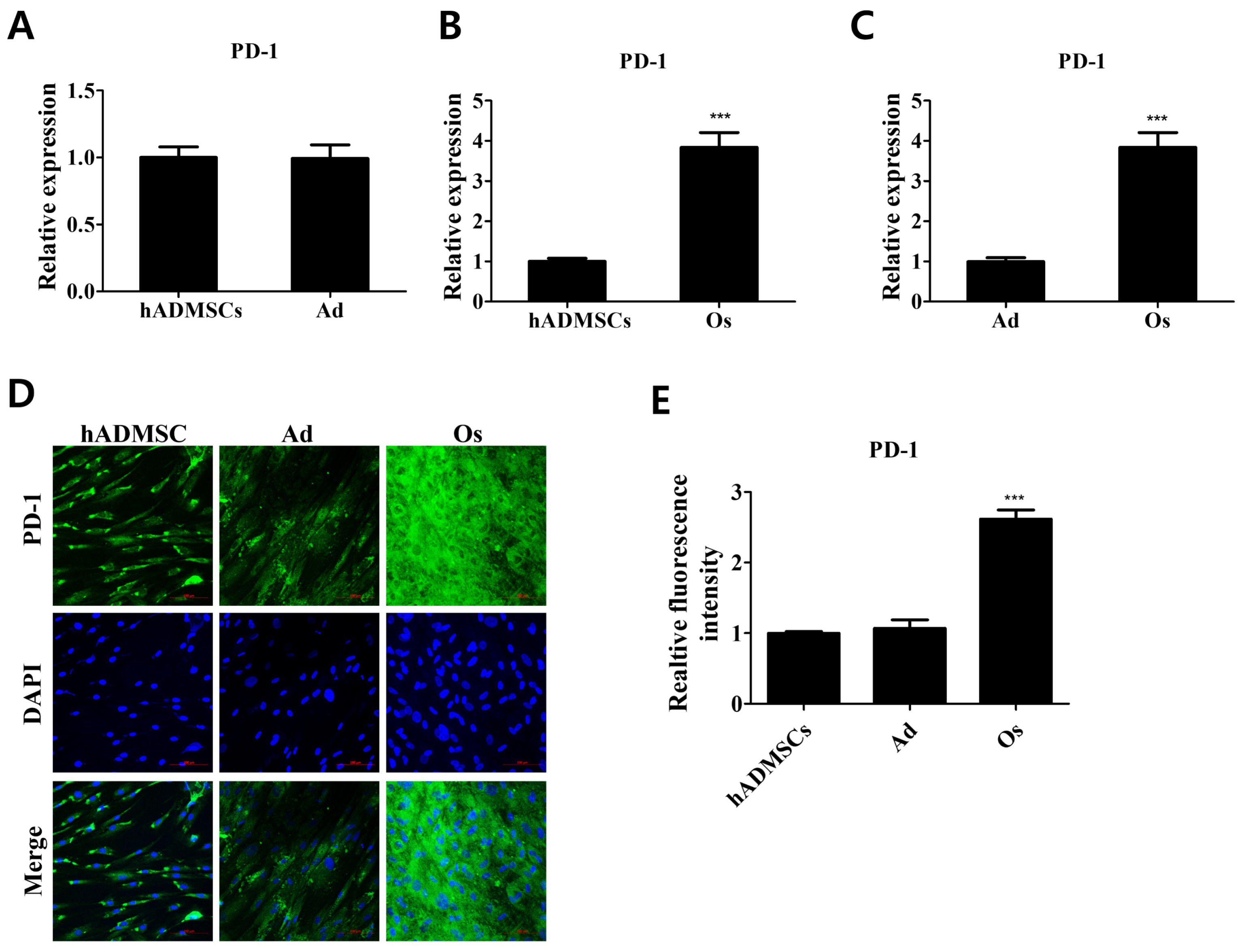
3.1. PD-1 Expression in Differentiated Cells Derived from hADMSCs
3.2. Immunosuppressive Effects Modulating the Expressions of Proinflammatory Cytokines in Differentiated Cells Cocultured with hADMSCs
3.3. Attenuation of the Expression of Proinflammatory Cytokines by Inhibition of the PD-1/PD-L1 Axis in Differentiated Cells Cocultured with hADMSCs
3.4. Regulatory Effects of the PD-1/PD-L1 Axis on Osteogenic Properties of Osteoblasts Cocultured with hADMSCs
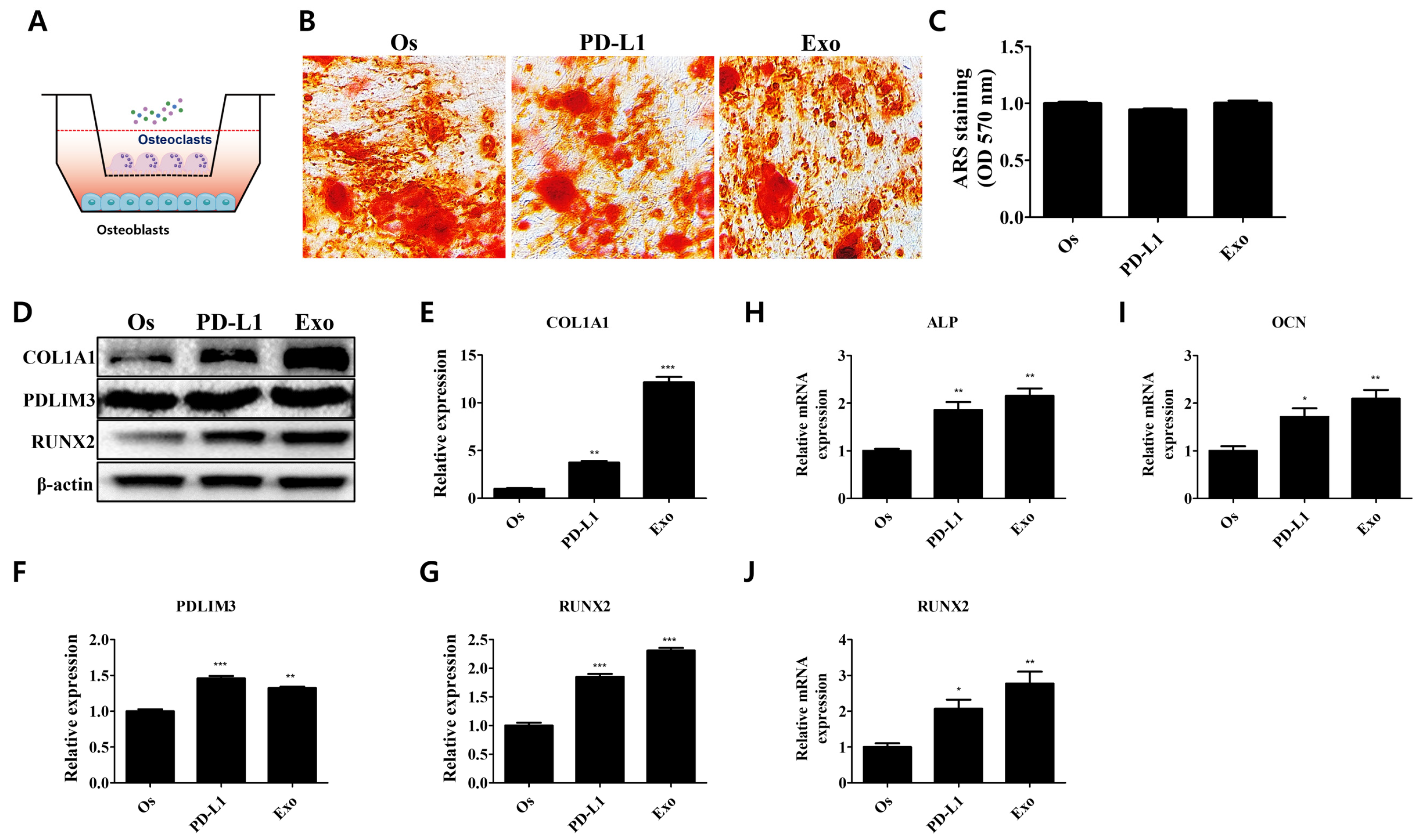
3.5. Effect of Exosomal PD-L1 on Bone Matrix Formation of Osteoblasts
3.6. Regulatory Effect of the PD-1/PD-L1 Axis on Bone Homeostasis in Cocultured Environments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fu, X.; Liu, G.; Halim, A.; Ju, Y.; Luo, Q.; Song, G. Mesenchymal Stem Cell Migration and Tissue Repair. Cells 2019, 8, 784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindroos, B.; Suuronen, R.; Miettinen, S. The Potential of Adipose Stem Cells in Regenerative Medicine. Stem Cell Rev. Rep. 2011, 7, 269–291. [Google Scholar] [CrossRef] [PubMed]
- Sohni, A.; Verfaillie, C.M. Mesenchymal Stem Cells Migration Homing and Tracking. Stem Cells Int. 2013, 2013, 130763. [Google Scholar] [CrossRef]
- Liu, S.; Liu, F.; Zhou, Y.; Jin, B.; Sun, Q.; Guo, S. Immunosuppressive Property of MSCs Mediated by Cell Surface Receptors. Front. Immunol. 2020, 11, 1076. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Lee, Y.J.; Choi, I.; Kim, M.; Sung, J.S. CXCL16/CXCR6 Axis in Adipocytes Differentiated from Human Adipose Derived Mesenchymal Stem Cells Regulates Macrophage Polarization. Cells 2021, 10, 3410. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-C.; Lee, Y.-J.; Shin, M.K.; Sung, J.-S. Regulation of CXCR6 Expression on Adipocytes and Osteoblasts Differentiated from Human Adipose Tissue-Derived Mesenchymal Stem Cells. Stem Cells Int. 2020, 2020, 8870133. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.J.; Kim, H.J.; Won, H.Y.; Min, Y.K.; Hwang, E.S. Acceleration of osteoblast differentiation by a novel osteogenic compound, DMP-PYT, through activation of both the BMP and Wnt pathways. Sci. Rep. 2017, 7, 8455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Yang, J.; Fang, J.; Zhou, Y.; Candi, E.; Wang, J.; Hua, D.; Shao, C.; Shi, Y. The secretion profile of mesenchymal stem cells and potential applications in treating human diseases. Signal Transduct. Target. Ther. 2022, 7, 92. [Google Scholar] [CrossRef]
- Davies, L.C.; Heldring, N.; Kadri, N.; Le Blanc, K. Mesenchymal Stromal Cell Secretion of Programmed Death-1 Ligands Regulates T Cell Mediated Immunosuppression. Stem Cells 2017, 35, 766–776. [Google Scholar] [CrossRef]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am. J. Cancer Res. 2020, 10, 727–742. [Google Scholar]
- Salmaninejad, A.; Khorramshahi, V.; Azani, A.; Soltaninejad, E.; Aslani, S.; Zamani, M.R.; Zal, M.; Nesaei, A.; Hosseini, S.M. PD-1 and cancer: Molecular mechanisms and polymorphisms. Immunogenetics 2018, 70, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Guo, S.; Li, F.; Sun, Q.; Liang, G. Exosomal PD-L1: New Insights Into Tumor Immune Escape Mechanisms and Therapeutic Strategies. Front. Cell Dev. Biol. 2020, 8, 569219. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Huang, A.C.; Zhang, W.; Zhang, G.; Wu, M.; Xu, W.; Yu, Z.; Yang, J.; Wang, B.; Sun, H.; et al. Exosomal PD-L1 contributes to immunosuppression and is associated with anti-PD-1 response. Nature 2018, 560, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Soder, R.; Abhyankar, S.; Abdelhakim, H.; Braun, M.W.; Trinidad, C.V.; Pathak, H.B.; Pessetto, Z.; Deighan, C.; Ganguly, S.; et al. WJMSC-derived small extracellular vesicle enhance T cell suppression through PD-L1. J. Extracell. Vesicles 2021, 10, e12067. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Goulart, V.; Ferreira, L.B.; Duarte, C.A.; de Lima, I.L.; Ferreira, E.R.; de Oliveira, B.C.; Vargas, L.N.; de Moraes, D.D.; Silva, I.B.B.; Faria, R.D.O.; et al. Mesenchymal stem cells from human adipose tissue and bone repair: A literature review. Biotechnol. Res. Innov. 2018, 2, 74–80. [Google Scholar] [CrossRef]
- Storti, G.; Scioli, M.G.; Kim, B.-S.; Orlandi, A.; Cervelli, V. Adipose-Derived Stem Cells in Bone Tissue Engineering: Useful Tools with New Applications. Stem Cells Int. 2019, 2019, 3673857. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Calle, J.; Riancho, J.A. The Role of DNA Methylation in Common Skeletal Disorders. Biology 2012, 1, 698–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, R.; Reilly, G.C. In vitro Models of Bone Remodelling and Associated Disorders. Front. Bioeng. Biotechnol. 2018, 6, 134. [Google Scholar] [CrossRef] [Green Version]
- Martin, T.J.; Ng, K.W. Mechanisms by which cells of the osteoblast lineage control osteoclast formation and activity. J. Cell. Biochem. 1994, 56, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Gu, Y.; Liao, Y.; Bang, S.; Donnelly, C.; Chen, O.; Tao, X.; Mirando, A.J.; Hilton, M.J.; Ji, R.-R. PD-1 blockade inhibits osteoclast formation and murine bone cancer pain. J. Clin. Investig. 2020, 130, 3603–3620. [Google Scholar] [CrossRef]
- Zuo, H.; Wan, Y. Inhibition of myeloid PD-L1 suppresses osteoclastogenesis and cancer bone metastasis. Cancer Gene Ther. 2022, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, X.; Zhang, C.; Wang, Y.; Cheng, T.; Duan, L.; Tong, Z.; Tan, S.; Zhang, H.; Saw, P.E.; et al. Tumor cell-intrinsic PD-1 receptor is a tumor suppressor and mediates resistance to PD-1 blockade therapy. Proc. Natl. Acad. Sci. USA 2020, 117, 6640–6650. [Google Scholar] [CrossRef]
- Watkins, L.R.; Maier, S.F.; Goehler, L.E. Immune activation: The role of pro-inflammatory cytokines in inflammation, illness responses and pathological pain states. Pain 1995, 63, 289–302. [Google Scholar] [CrossRef]
- Ayoub, S.; Berbéri, A.; Fayyad-Kazan, M. Cytokines, Masticatory Muscle Inflammation, and Pain: An Update. J. Mol. Neurosci. 2020, 70, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umare, V.; Pradhan, V.; Nadkar, M.; Rajadhyaksha, A.; Patwardhan, M.; Ghosh, K.K.; Nadkarni, A.H. Effect of Proinflammatory Cytokines (IL-6, TNF-α, and IL-1β) on Clinical Manifestations in Indian SLE Patients. Mediat. Inflamm. 2014, 2014, 385297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Mi, J.; Liu, W.; Xiao, S.; Gao, C. Blocking of checkpoint receptor PD-L1 aggravates osteoarthritis in macrophage-dependent manner in the mice model. Int. J. Immunopathol. Pharmacol. 2019, 33, 2058738418820760. [Google Scholar] [CrossRef] [PubMed]
- Amarasekara, D.; Kim, S.; Rho, J. Regulation of Osteoblast Differentiation by Cytokine Networks. Int. J. Mol. Sci. 2021, 22, 2851. [Google Scholar] [CrossRef] [PubMed]
- Metzger, C.E.; Narayanan, S.A. The Role of Osteocytes in Inflammatory Bone Loss. Front. Endocrinol. 2019, 10, 285. [Google Scholar] [CrossRef]
- Lacey, D.; Simmons, P.; Graves, S.; Hamilton, J. Proinflammatory cytokines inhibit osteogenic differentiation from stem cells: Implications for bone repair during inflammation. Osteoarthr. Cartil. 2009, 17, 735–742. [Google Scholar] [CrossRef] [Green Version]
- Kaneshiro, S.; Ebina, K.; Shi, K.; Higuchi, C.; Hirao, M.; Okamoto, M.; Hashimoto, J. IL-6 negatively regulates osteoblast differentiation through the SHP2/MEK2 and SHP2/Akt2 pathways in vitro. J. Bone Miner. Metab. 2014, 32, 378–392. [Google Scholar] [CrossRef] [PubMed]
- Lehti, M.S.; Henriksson, H.; Rummukainen, P.; Wang, F.; Uusitalo-Kylmälä, L.; Kiviranta, R.; Heino, T.; Kotaja, N.; Sironen, A. Cilia-related protein SPEF2 regulates osteoblast differentiation. Sci. Rep. 2018, 8, 859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.-H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef] [PubMed]
- Moll, G.; Hoogduijn, M.J.; Ankrum, J.A. Editorial: Safety, Efficacy and Mechanisms of Action of Mesenchymal Stem Cell Therapies. Front. Immunol. 2020, 11, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Jiang, J.; Gu, Z.; Zhang, J.; Chen, Y.; Liu, X. Mesenchymal stromal cell therapies: Immunomodulatory properties and clinical progress. Stem Cell Res. Ther. 2020, 11, 345. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.-C.; Kang, K.-S. Functional enhancement strategies for immunomodulation of mesenchymal stem cells and their therapeutic application. Stem Cell Res. Ther. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wen, M.; Cao, Y.; Wu, B.; Xiao, T.; Cao, R.; Wang, Q.; Liu, X.; Xue, H.; Yu, Y.; Lin, J.; et al. PD-L1 degradation is regulated by electrostatic membrane association of its cytoplasmic domain. Nat. Commun. 2021, 12, 5106. [Google Scholar] [CrossRef]
- Akinleye, A.; Rasool, Z. Immune checkpoint inhibitors of PD-L1 as cancer therapeutics. J. Hematol. Oncol. 2019, 12, 92. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Zhu, Z.; Chen, X.; Zhang, H.; Huang, J.; Gu, S.; Zhao, X. The Importance of Exosomal PD-L1 in Cancer Progression and Its Potential as a Therapeutic Target. Cells 2021, 10, 3247. [Google Scholar] [CrossRef]
- Di Tinco, R.; Bertani, G.; Pisciotta, A.; Bertoni, L.; Pignatti, E.; Maccaferri, M.; Bertacchini, J.; Sena, P.; Vallarola, A.; Tupler, R.; et al. Role of PD-L1 in licensing immunoregulatory function of dental pulp mesenchymal stem cells. Stem Cell Res. Ther. 2021, 12, 598. [Google Scholar] [CrossRef]
- Blair, H.C.; Larrouture, Q.C.; Li, Y.; Lin, H.; Beer-Stoltz, D.; Liu, L.; Nelson, D.J. Osteoblast Differentiation and Bone Matrix Formation In Vivo and In Vitro. Tissue Eng. B Rev. 2017, 23, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Bourne, L.E.; Wheeler-Jones, C.P.; Orriss, I.R. Regulation of mineralisation in bone and vascular tissue: A comparative review. J. Endocrinol. 2021, 248, R51–R65. [Google Scholar] [CrossRef] [PubMed]
- Lodder, E.M.; Scicluna, B.P.; Beekman, L.; Arends, D.; Moerland, P.D.; Tanck, M.W.; Adriaens, M.E.; Bezzina, C.R. Integrative Genomic Approach Identifies Multiple Genes Involved in Cardiac Collagen Deposition. Circ. Cardiovasc. Genet. 2014, 7, 790–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Fang, J.; Lv, J.; Pan, Z.; Yin, X.; Cheng, H.; Guo, X. Novel polymorphisms in PDLIM3 and PDLIM5 gene encoding Z-line proteins increase risk of idiopathic dilated cardiomyopathy. J. Cell Mol. Med. 2019, 23, 7054–7062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salhotra, A.; Shah, H.N.; Levi, B.; Longaker, M.T. Mechanisms of bone development and repair. Nat. Rev. Mol. Cell Biol. 2020, 21, 696–711. [Google Scholar] [CrossRef] [PubMed]
- Borciani, G.; Montalbano, G.; Baldini, N.; Cerqueni, G.; Vitale-Brovarone, C.; Ciapetti, G. Co–culture systems of osteoblasts and osteoclasts: Simulating in vitro bone remodeling in regenerative approaches. Acta Biomater. 2020, 108, 22–45. [Google Scholar] [CrossRef]
- Jimi, E.; Hirata, S.; Osawa, K.; Terashita, M.; Kitamura, C.; Fukushima, H. The Current and Future Therapies of Bone Regeneration to Repair Bone Defects. Int. J. Dent. 2012, 2012, 148261. [Google Scholar] [CrossRef] [Green Version]
- Schröder, H.; Wang, X.; Wiens, M.; Diehl-Seifert, B.; Kropf, K.; Schloßmacher, U.; Müller, W. Silicate modulates the cross-talk between osteoblasts (SaOS-2) and osteoclasts (RAW 264.7 cells): Inhibition of osteoclast growth and differentiation. J. Cell. Biochem. 2012, 113, 3197–3206. [Google Scholar] [CrossRef]
- Remmers, S.J.A.; de Wildt, B.W.M.; Vis, M.A.M.; Spaander, E.S.R.; de Vries, R.B.M.; Ito, K.; Hofmann, S. Osteoblast-osteoclast co-cultures: A systematic review and map of available literature. PLoS ONE 2021, 16, e0257724. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequences | |
|---|---|---|
| PD-1 | Forward | CCCTGGTGGTTGGTGTCGT |
| Reverse | GCCTGGCTCCTATTGTCCCTC | |
| TNF-α | Forward | AGAGAAGCCAACTACAGACC |
| Reverse | CAGTATGTGAGAGGAAGAGAA | |
| IL-6 | Forward | CAGAACAGATTTGAGAGTAGTGA |
| Reverse | CGCAGAATGAGATGAGTTGT | |
| IL-1β | Forward | GGCTTATTACAGTGGCAATG |
| Reverse | TAGTGGTGGTCGGAGATT | |
| ALP | Forward | TCATCCGTGGTTGTATCA |
| Reverse | GTGGTCTCAGTAGCATCT | |
| RUNX2 | Forward | AAGGCAGTTACATATCAATACAG |
| Reverse | GAGGCAGAGGCTTCAATA | |
| OCN | Forward | CGGAGTAGTCATCATTGTG |
| Reverse | CGAGTGTTCATTCTGTTCA | |
| FABP4 | Forward | TCAAGAGCACCATAACCTT |
| Reverse | TTCCACCACCAGTTTATCA | |
| Adiponectin | Forward | ACCACTATGATGGCTCCACT |
| Reverse | GGTGAAGAGCATAGCCTTGT | |
| PPARγ | Forward | CGAAGACATTCCATTCACAA |
| Reverse | CACAGACACGACATTCAAT | |
| GAPDH | Forward | TATGACAACAGCCTCAAGAT |
| Reverse | GAGTCCTTCCACGATACC |
| Target | Catalogue Number | Dilution |
|---|---|---|
| ALP | SC-271431 | 1/500 |
| COL1A1 | Ab34710 | 1/500 |
| OCN | SC-7449 | 1/500 |
| RUNX2 | SC-10758 | 1/500 |
| PDLIM3 | SC-98652 | 1/500 |
| β-actin | SC-47778 | 1/500 |
| PD-1 | AF1086 | 1/200 |
| Anti-rabbit IgG HRP-linked antibody | 7074S | 1/1500 |
| m-IgGκ BP-HRP | SC-516102 | 1/1500 |
| Donkey anti-goat IgG NorthernLights | NL003 | 1/1000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-C.; Shin, M.K.; Jang, B.-Y.; Lee, S.-H.; Kim, M.; Sung, J.-S. Immunomodulatory Effect and Bone Homeostasis Regulation in Osteoblasts Differentiated from hADMSCs via the PD-1/PD-L1 Axis. Cells 2022, 11, 3152. https://doi.org/10.3390/cells11193152
Lee S-C, Shin MK, Jang B-Y, Lee S-H, Kim M, Sung J-S. Immunomodulatory Effect and Bone Homeostasis Regulation in Osteoblasts Differentiated from hADMSCs via the PD-1/PD-L1 Axis. Cells. 2022; 11(19):3152. https://doi.org/10.3390/cells11193152
Chicago/Turabian StyleLee, Seung-Cheol, Min Kyoung Shin, Bo-Young Jang, Seung-Ho Lee, Min Kim, and Jung-Suk Sung. 2022. "Immunomodulatory Effect and Bone Homeostasis Regulation in Osteoblasts Differentiated from hADMSCs via the PD-1/PD-L1 Axis" Cells 11, no. 19: 3152. https://doi.org/10.3390/cells11193152
APA StyleLee, S. -C., Shin, M. K., Jang, B. -Y., Lee, S. -H., Kim, M., & Sung, J. -S. (2022). Immunomodulatory Effect and Bone Homeostasis Regulation in Osteoblasts Differentiated from hADMSCs via the PD-1/PD-L1 Axis. Cells, 11(19), 3152. https://doi.org/10.3390/cells11193152