

Ethylene Activates the EIN2-EIN3/EIL1 Signaling Pathway in Tapetum and Disturbs Anther Development in Arabidopsis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Growth Conditions and Mutant Identification
2.2. Ethylene Gas Treatment
2.3. Microscopy
2.4. Molecular Cloning
2.5. Real-Time PCR Analyses
3. Results
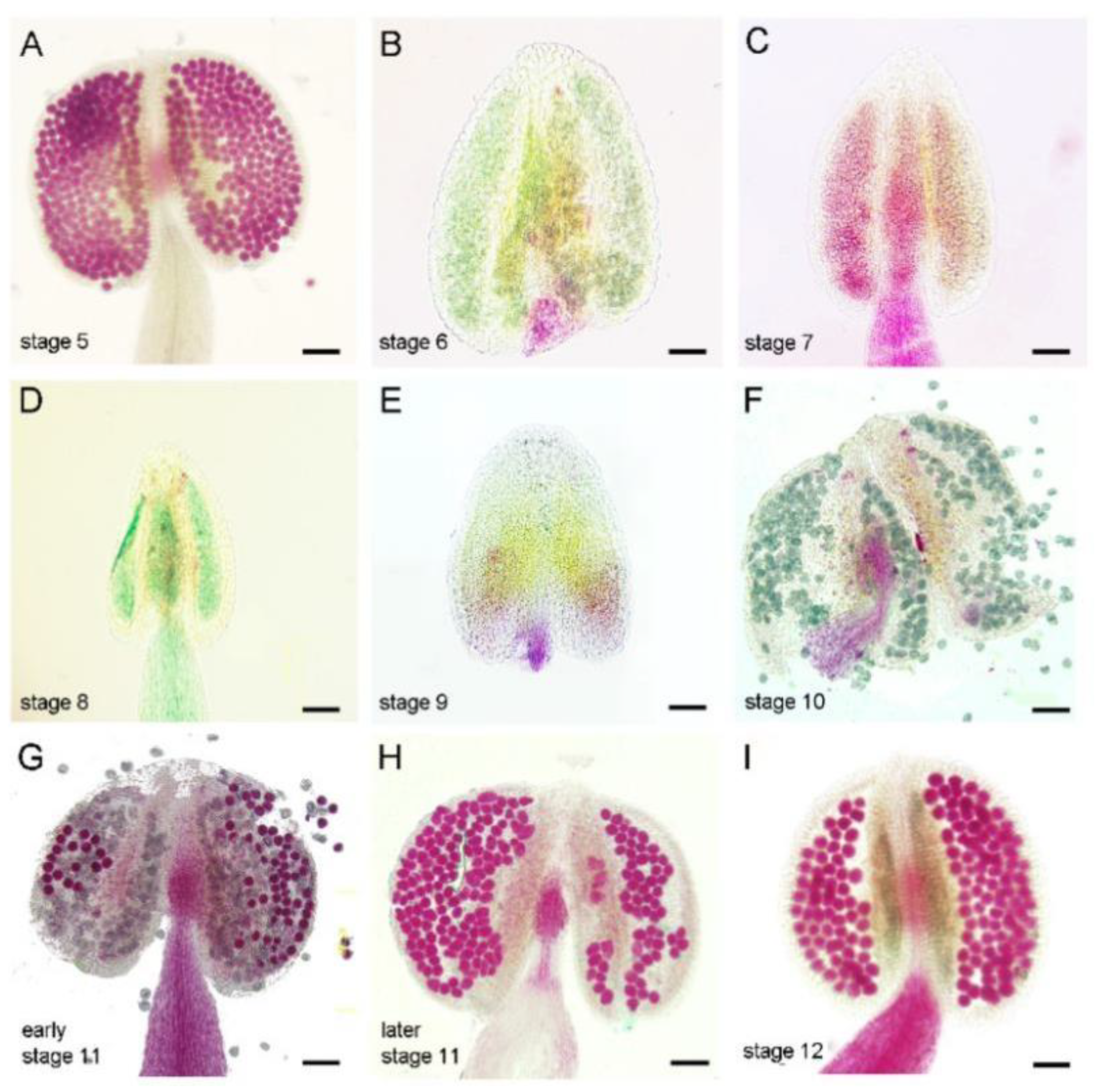
3.1. Exogenous Ethylene Treatment Causes Filament Shortening, Pollen Abortion and Leads to Male Sterility
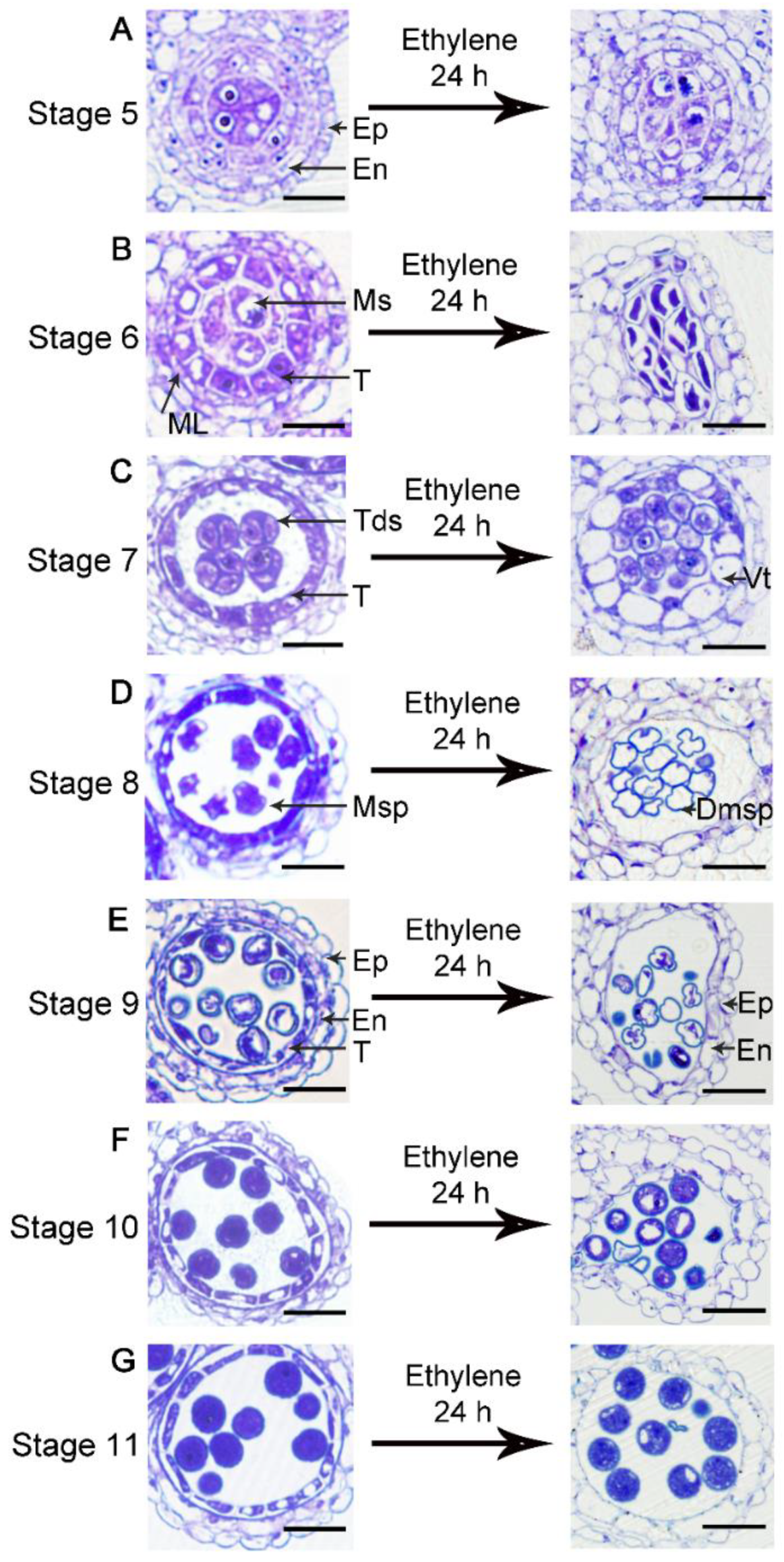
3.2. Ethylene Treatment Leads to Degeneration of the Tapetum and Microspores
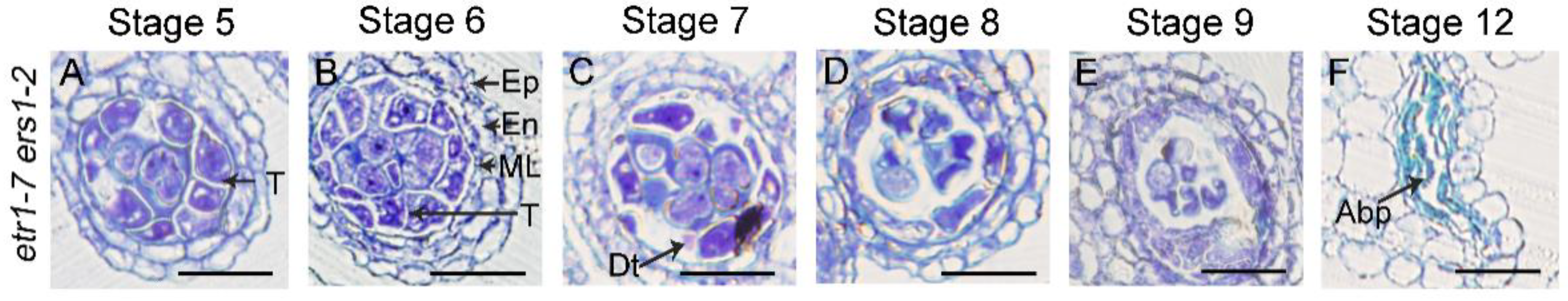
3.3. The Anther Defects Induced by Ethylene Treatment Are EIN2- and EIN3/EIL1-Dependent
3.4. Exogenous Ethylene Treatment Activates EIN2 and EIN3 Transcription in Tapetum
3.5. Exogenous Ethylene Treatment Can Stabilize the Accumulation of EIN3 in Tapetum
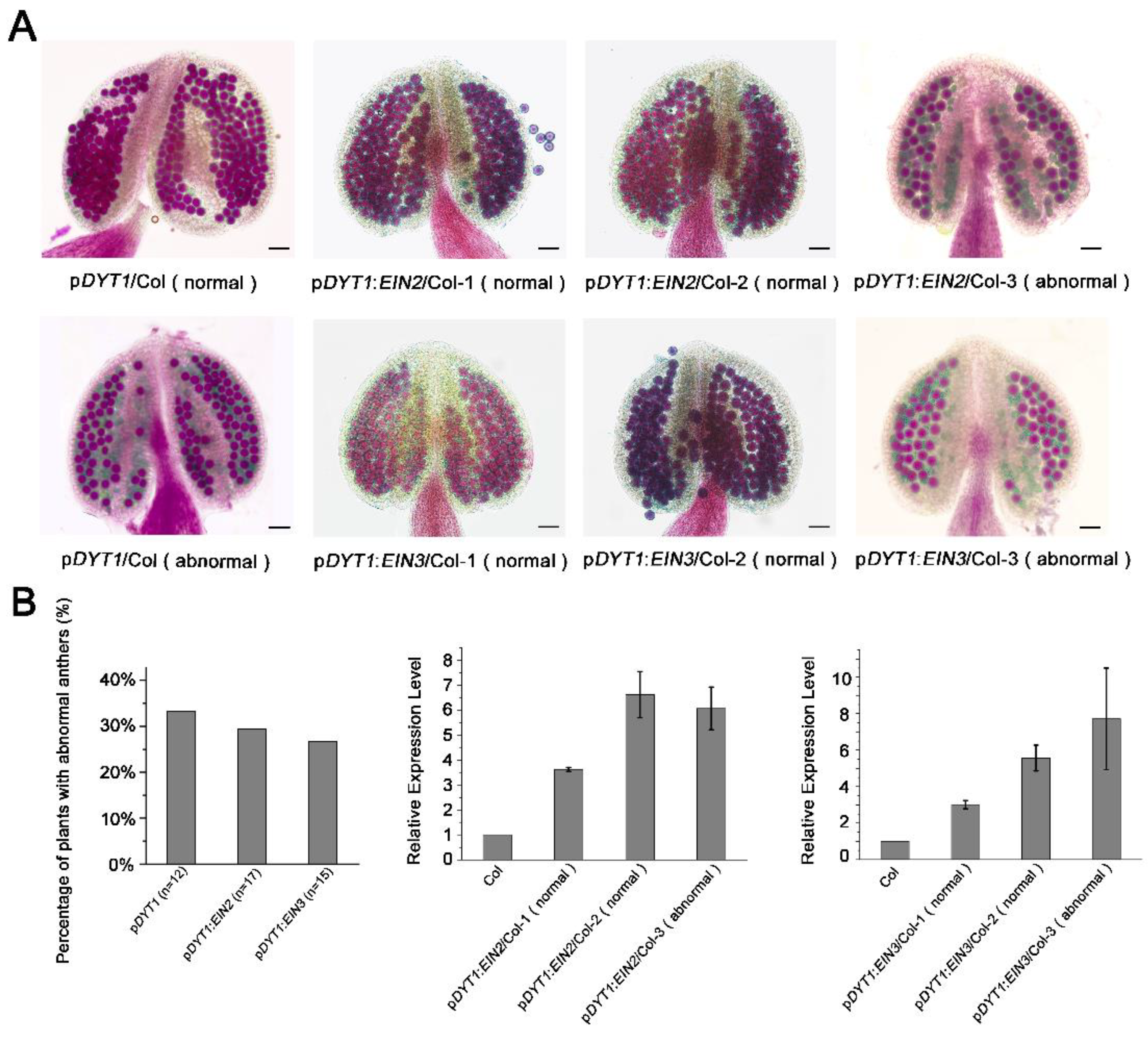
3.6. The Expression of Several ERFs and SAGs Genes Are Induced by Ethylene and the Expression of the Five Key Transcription Factor Genes in Tapetum Development Is Reduced after Ethylene Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubois, M.; Van den Broeck, L.; Inzé, D. The Pivotal Role of Ethylene in Plant Growth. Trends Plant Sci. 2018, 23, 311–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booker, M.A.; DeLong, A. Producing the Ethylene Signal: Regulation and Diversification of Ethylene Biosynthetic Enzymes. Plant Physiol. 2015, 169, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Hua, J.; Meyerowitz, E.M. Ethylene responses are negatively regulated by a receptor gene family in Arabidopsis thaliana. Cell 1998, 94, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Ju, C.; Yoon, G.M.; Shemansky, J.M.; Lin, D.Y.; Ying, Z.I.; Chang, J.; Garrett, W.M.; Kessenbrock, M.; Groth, G.; Tucker, M.L.; et al. CTR1 phosphorylates the central regulator EIN2 to control ethylene hormone signaling from the ER membrane to the nucleus in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 19486–19491. [Google Scholar] [CrossRef] [Green Version]
- Dolgikh, V.A.; Pukhovaya, E.M.; Zemlyanskaya, E.V. Shaping Ethylene Response: The Role of EIN3/EIL1 Transcription Factors. Front. Plant Sci. 2019, 10, 1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Völz, R.; Heydlauff, J.; Ripper, D.; von Lyncker, L.; Groß-Hardt, R. Ethylene signaling is required for synergid degeneration and the establishment of a pollen tube block. Dev. Cell 2013, 25, 310–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Teng, X.D.; Zheng, Q.Q.; Zhao, Y.Y.; Lu, J.Y.; Wang, Y.; Guo, H.; Yang, Z.N. Ethylene signaling is critical for synergid cell functional specification and pollen tube attraction. Plant J. Cell Mol. Biol. 2018, 96, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, Q.; Lyu, M.; Wang, Z.; Song, Z.; Zhong, S.; Gu, H.; Dong, J.; Dresselhaus, T.; Zhong, S.; et al. Lack of ethylene does not affect reproductive success and synergid cell death in Arabidopsis. Mol. Plant 2022, 15, 354–362. [Google Scholar] [CrossRef]
- Hao, Y.J.; Wang, D.H.; Peng, Y.B.; Bai, S.L.; Xu, L.Y.; Li, Y.Q.; Xu, Z.H.; Bai, S.N. DNA damage in the early primordial anther is closely correlated with stamen arrest in the female flower of cucumber (Cucumis sativus L.). Planta 2003, 217, 888–895. [Google Scholar] [CrossRef]
- Bai, S.L.; Peng, Y.B.; Cui, J.X.; Gu, H.T.; Xu, L.Y.; Li, Y.Q.; Xu, Z.H.; Bai, S.N. Developmental analyses reveal early arrests of the spore-bearing parts of reproductive organs in unisexual flowers of cucumber (Cucumis sativus L.). Planta 2004, 220, 230–240. [Google Scholar] [CrossRef]
- Iwahori, S.; Lyons, J.M.; Smith, O.E. Sex expression in cucumber plants as affected by 2-chloroethylphosphonic Acid, ethylene, and growth regulators. Plant Physiol. 1970, 46, 412–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, H.; Suge, H. Sex Expression and Ethylene Production in Cucumber Plants as Affected by 1-Aminocyclopropane-1-carboxylic Acid. J. Jpn. Soc. Hortic. Sci. 1982, 51, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Perl-Treves, R. Male to Female Conversion along the Cucumber Shoot: Approaches to Studying Sex Genes and Flora Development in Cucumis sativus; BIOS Scientific: Oxford, UK, 1999. [Google Scholar]
- Mibus, H.; Tatlioglu, T. Molecular characterization and isolation of the F/f gene for femaleness in cucumber (Cucumis sativus L.). TAG Theor. Appl. Genetics. Theor. Und Angew. Genet. 2004, 109, 1669–1676. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Huang, S.; Liu, S.; Pan, J.; Zhang, Z.; Tao, Q.; Shi, Q.; Jia, Z.; Zhang, W.; Chen, H.; et al. Molecular isolation of the M gene suggests that a conserved-residue conversion induces the formation of bisexual flowers in cucumber plants. Genetics 2009, 182, 1381–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boualem, A.; Troadec, C.; Camps, C.; Lemhemdi, A.; Morin, H.; Sari, M.A.; Fraenkel-Zagouri, R.; Kovalski, I.; Dogimont, C.; Perl-Treves, R.; et al. A cucurbit androecy gene reveals how unisexual flowers develop and dioecy emerges. Science 2015, 350, 688–691. [Google Scholar] [CrossRef]
- Duan, Q.H.; Wang, D.H.; Xu, Z.H.; Bai, S.N. Stamen development in Arabidopsis is arrested by organ-specific overexpression of a cucumber ethylene synthesis gene CsACO2. Planta 2008, 228, 537–543. [Google Scholar] [CrossRef]
- Wang, D.H.; Li, F.; Duan, Q.H.; Han, T.; Xu, Z.H.; Bai, S.N. Ethylene perception is involved in female cucumber flower development. Plant J. Cell Mol. Biol. 2010, 61, 862–872. [Google Scholar] [CrossRef]
- Hall, A.E.; Bleecker, A.B. Analysis of combinatorial loss-of-function mutants in the Arabidopsis ethylene receptors reveals that the ers1 etr1 double mutant has severe developmental defects that are EIN2 dependent. Plant Cell 2003, 15, 2032–2041. [Google Scholar] [CrossRef] [Green Version]
- Gu, H.T.; Wang, D.H.; Li, X.; He, C.X.; Xu, Z.H.; Bai, S.N. Characterization of an ethylene-inducible, calcium-dependent nuclease that is differentially expressed in cucumber flower development. New Phytol. 2011, 192, 590–600. [Google Scholar] [CrossRef]
- Sanders, P.M.; Bui, A.Q.; Weterings, K.; McIntire, K.N.; Hsu, Y.-C.; Lee, P.Y.; Truong, M.T.; Beals, T.P.; Goldberg, R.B. Anther developmental defects in Arabidopsis thaliana male-sterile mutants. Sex. Plant Reprod. 1999, 11, 297–322. [Google Scholar] [CrossRef]
- Bouchez, O.; Huard, C.; Lorrain, S.; Roby, D.; Balagué, C. Ethylene is one of the key elements for cell death and defense response control in the Arabidopsis lesion mimic mutant vad1. Plant Physiol. 2007, 145, 465–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Xue, L.; Chintamanani, S.; Germain, H.; Lin, H.; Cui, H.; Cai, R.; Zuo, J.; Tang, X.; Li, X.; et al. Ethylene insensitive3 and ethylene insensitive3-like1 repress salicylic acid induction deficient2 expression to negatively regulate plant innate immunity in Arabidopsis. Plant Cell 2009, 21, 2527–2540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.C.; Qu, X.; Mathews, D.E.; Schaller, G.E. Effect of ethylene pathway mutations upon expression of the ethylene receptor ETR1 from Arabidopsis. Plant Physiol. 2002, 130, 1983–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, M.P. Differential staining of aborted and nonaborted pollen. Stain. Technol. 1969, 44, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Tian, L.; Sun, M.X.; Huang, X.Y.; Zhu, J.; Guan, Y.F.; Jia, Q.S.; Yang, Z.N. AUXIN RESPONSE FACTOR17 is essential for pollen wall pattern formation in Arabidopsis. Plant Physiol. 2013, 162, 720–731. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.N.; Zhu, J.; Yu, Y.; Teng, X.D.; Lou, Y.; Xu, X.F.; Liu, J.L.; Yang, Z.N. DYT1 directly regulates the expression of TDF1 for tapetum development and pollen wall formation in Arabidopsis. Plant J. Cell Mol. Biol. 2014, 80, 1005–1013. [Google Scholar] [CrossRef]
- Zhu, J.; Lou, Y.; Shi, Q.S.; Zhang, S.; Zhou, W.T.; Yang, J.; Zhang, C.; Yao, X.Z.; Xu, T.; Liu, J.L.; et al. Slowing development restores the fertility of thermo-sensitive male-sterile plant lines. Nat. Plants 2020, 6, 360–367. [Google Scholar] [CrossRef]
- Xing, S.; Zachgo, S. Pollen lethality: A phenomenon in Arabidopsis RNA interference plants. Plant Physiol. 2007, 145, 330–333. [Google Scholar] [CrossRef] [Green Version]
- Potuschak, T.; Lechner, E.; Parmentier, Y.; Yanagisawa, S.; Grava, S.; Koncz, C.; Genschik, P. EIN3-dependent regulation of plant ethylene hormone signaling by two arabidopsis F box proteins: EBF1 and EBF2. Cell 2003, 115, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Ecker, J.R. Plant responses to ethylene gas are mediated by SCF(EBF1/EBF2)-dependent proteolysis of EIN3 transcription factor. Cell 2003, 115, 667–677. [Google Scholar] [CrossRef]
- An, F.; Zhao, Q.; Ji, Y.; Li, W.; Jiang, Z.; Yu, X.; Zhang, C.; Han, Y.; He, W.; Liu, Y.; et al. Ethylene-induced stabilization of ETHYLENE INSENSITIVE3 and EIN3-LIKE1 is mediated by proteasomal degradation of EIN3 binding F-box 1 and 2 that requires EIN2 in Arabidopsis. Plant Cell 2010, 22, 2384–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masaru, O.-T.; Hideaki, S. Ethylene-inducible DNA binding proteins that interact with an ethylene responsive element. Plant Cell 1995, 7, 173–182. [Google Scholar]
- Lorenzo, O.; Piqueras, R.; Sánchez-Serrano, J.J.; Solano, R. ETHYLENE RESPONSE FACTOR1 Integrates Signals from Ethylene and Jasmonate Pathways in Plant Defense. Plant Cell 2003, 15, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, S.Y.; Ohta, M.; Usui, A.; Shinshi, H.; Takagi, M.O. Arabidopsis Ethylene-Responsive Element Binding Factors Act as Transcriptional Activators or Respressors of GCC Box-Mediated Gene Expression. Plant Cell 2000, 12, 393–404. [Google Scholar]
- Jing, H.C.; Schippers, J.H.; Hille, J.; Dijkwel, P.P. Ethylene-induced leaf senescence depends on age-related changes and OLD genes in Arabidopsis. J. Exp. Bot. 2005, 56, 2915–2923. [Google Scholar] [CrossRef] [PubMed]
- Sharabi-Schwager, M.; Samach, A.; Porat, R. Overexpression of the CBF2 transcriptional activator in Arabidopsis suppresses the responsiveness of leaf tissue to the stress hormone ethylene. Plant Biol. 2010, 12, 630–638. [Google Scholar] [CrossRef]
- Liu, W.; Yin, H.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J. Comparative Transcriptome Analysis of Flower Senescence of Camellia lutchuensis. Curr. Genom. 2022, 23, 66–76. [Google Scholar] [CrossRef]
- Li, D.-D.; Xue, J.-S.; Zhu, J.; Yang, Z.-N. Gene Regulatory Network for Tapetum Development in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 1559. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Lou, Y.; Xu, X.; Yang, Z.N. A genetic pathway for tapetum development and function in Arabidopsis. J. Integr. Plant Biol. 2011, 53, 892–900. [Google Scholar] [CrossRef]
- Bleecker, A.B.; Estelle, M.A.; Somerville, C.; Kende, H. Insensitivity to Ethylene Conferred by a Dominant Mutation in Arabidopsis thaliana. Science 1988, 241, 1086–1089. [Google Scholar] [CrossRef] [PubMed]
- Ariizumi, T.; Toriyama, K. Genetic Regulation of Sporopollenin Synthesis and Pollen Exine Development. Annu. Rev. Plant Biol. 2011, 62, 437–460. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Guo, X.; Zhang, J.; Liu, Y.; Wang, B.; Li, H.; Lu, H. betaVPE is involved in tapetal degradation and pollen development by activating proprotease maturation in Arabidopsis thaliana. J. Exp. Bot. 2020, 71, 1943–1955. [Google Scholar] [CrossRef] [PubMed]
- Grienenberger, E.; Quilichini, T.D. The Toughest Material in the Plant Kingdom: An Update on Sporopollenin. Front. Plant Sci. 2021, 12, 703864. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Hu, W.; Yang, Z.-N. The contributions of sporophytic tapetum to pollen formation. Seed Biol. 2022, 1, 1–13. [Google Scholar] [CrossRef]
- Kawanabe, T.; Ariizumi, T.; Kawai-Yamada, M.; Uchimiya, H.; Toriyama, K. Abolition of the tapetum suicide program ruins microsporogenesis. Plant Cell Physiol. 2006, 47, 784–787. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Liu, D.; Lv, X.; Wang, Y.; Xun, Z.; Liu, Z.; Li, F.; Lu, H. The cysteine protease CEP1, a key executor involved in tapetal programmed cell death, regulates pollen development in Arabidopsis. Plant Cell 2014, 26, 2939–2961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, Y.; Zhu, J.; Yang, Z. Molecular Cell Biology of Pollen Walls. In Applied Plant Cell Biology: Cellular Tools and Approaches for Plant Biotechnology; Nick, P., Opatrny, Z., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 179–205. [Google Scholar] [CrossRef]
- Skirycz, A.; Claeys, H.; De Bodt, S.; Oikawa, A.; Shinoda, S.; Andriankaja, M.; Maleux, K.; Eloy, N.B.; Coppens, F.; Yoo, S.D.; et al. Pause-and-stop: The effects of osmotic stress on cell proliferation during early leaf development in Arabidopsis and a role for ethylene signaling in cell cycle arrest. Plant Cell 2011, 23, 1876–1888. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Claeys, H.; Van den Broeck, L.; Inzé, D. Time of day determines Arabidopsis transcriptome and growth dynamics under mild drought. Plant Cell Environ. 2017, 40, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Savada, R.P.; Ozga, J.A.; Jayasinghege, C.P.A.; Waduthanthri, K.D.; Reinecke, D.M. Heat stress differentially modifies ethylene biosynthesis and signaling in pea floral and fruit tissues. Plant Mol. Biol. 2017, 95, 313–331. [Google Scholar] [CrossRef]
- Miller, J.D.; Arteca, R.N.; Pell, E.J. Senescence-associated gene expression during ozone-induced leaf senescence in Arabidopsis. Plant Physiol. 1999, 120, 1015–1024. [Google Scholar] [CrossRef] [Green Version]
- Keates, S.E.; Kostman, T.A.; Anderson, J.D.; Bailey, B.A. Altered gene expression in three plant species in response to treatment with Nep1, a fungal protein that causes necrosis. Plant Physiol. 2003, 132, 1610–1622. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, B.-S.; Zhu, Y.-X.; Zhang, Y.-F.; Zhong, X.; Pan, K.-Y.; Jiang, Y.; Wen, C.-K.; Yang, Z.-N.; Yao, X. Ethylene Activates the EIN2-EIN3/EIL1 Signaling Pathway in Tapetum and Disturbs Anther Development in Arabidopsis. Cells 2022, 11, 3177. https://doi.org/10.3390/cells11193177
Zhu B-S, Zhu Y-X, Zhang Y-F, Zhong X, Pan K-Y, Jiang Y, Wen C-K, Yang Z-N, Yao X. Ethylene Activates the EIN2-EIN3/EIL1 Signaling Pathway in Tapetum and Disturbs Anther Development in Arabidopsis. Cells. 2022; 11(19):3177. https://doi.org/10.3390/cells11193177
Chicago/Turabian StyleZhu, Ben-Shun, Ying-Xiu Zhu, Yan-Fei Zhang, Xiang Zhong, Keng-Yu Pan, Yu Jiang, Chi-Kuang Wen, Zhong-Nan Yang, and Xiaozhen Yao. 2022. "Ethylene Activates the EIN2-EIN3/EIL1 Signaling Pathway in Tapetum and Disturbs Anther Development in Arabidopsis" Cells 11, no. 19: 3177. https://doi.org/10.3390/cells11193177
APA StyleZhu, B. -S., Zhu, Y. -X., Zhang, Y. -F., Zhong, X., Pan, K. -Y., Jiang, Y., Wen, C. -K., Yang, Z. -N., & Yao, X. (2022). Ethylene Activates the EIN2-EIN3/EIL1 Signaling Pathway in Tapetum and Disturbs Anther Development in Arabidopsis. Cells, 11(19), 3177. https://doi.org/10.3390/cells11193177