
Mapping the Universe of Eph Receptor and Ephrin Ligand Transcripts in Epithelial and Fiber Cells of the Eye Lens


Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. RNA Isolation from Epithelial Cells
2.3. RNA Isolation from Fiber Cells, Whole Lens Samples, or Positive Control Samples
2.4. Primer Design
2.5. Reverse Transcription and Polymerase Chain Reaction (RT-PCR)
2.6. Gel Electrophoresis and DNA Extraction from Gel Pieces
2.7. Microarray Data Comparison
3. Results
3.1. EphA Transcripts in the Lens
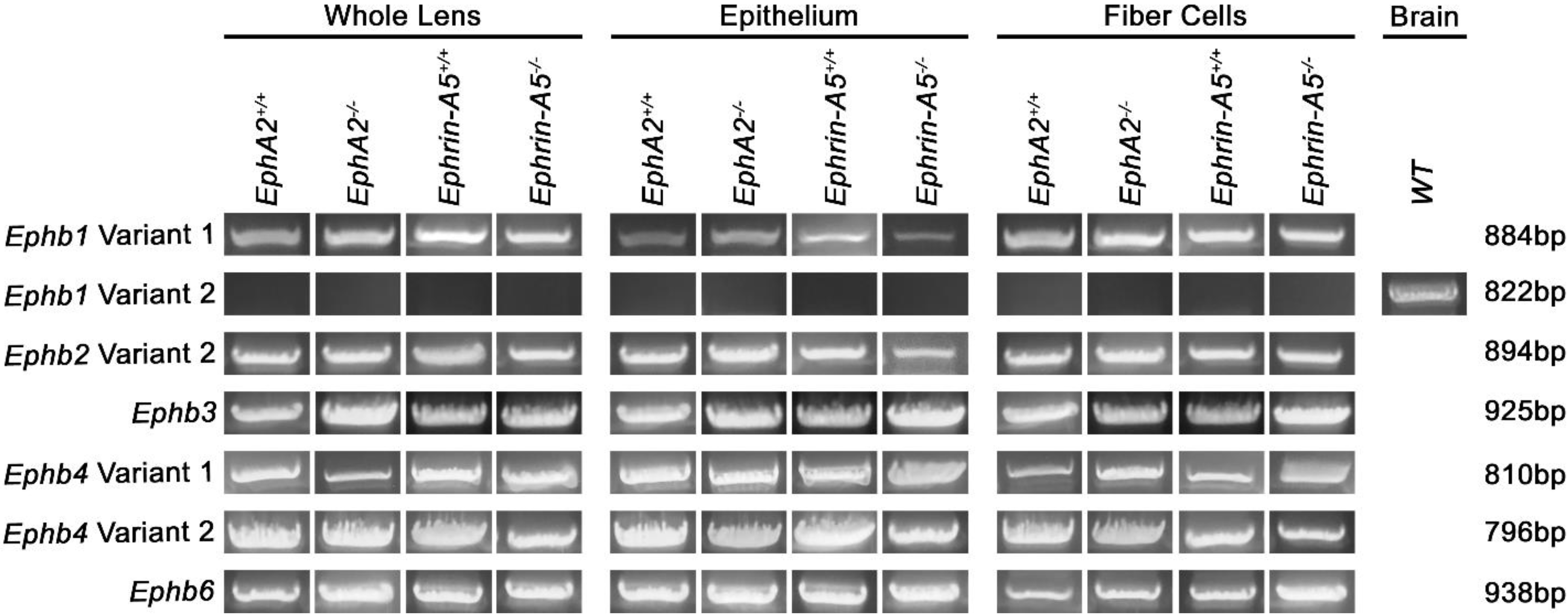
3.2. EphB Transcripts in the Lens
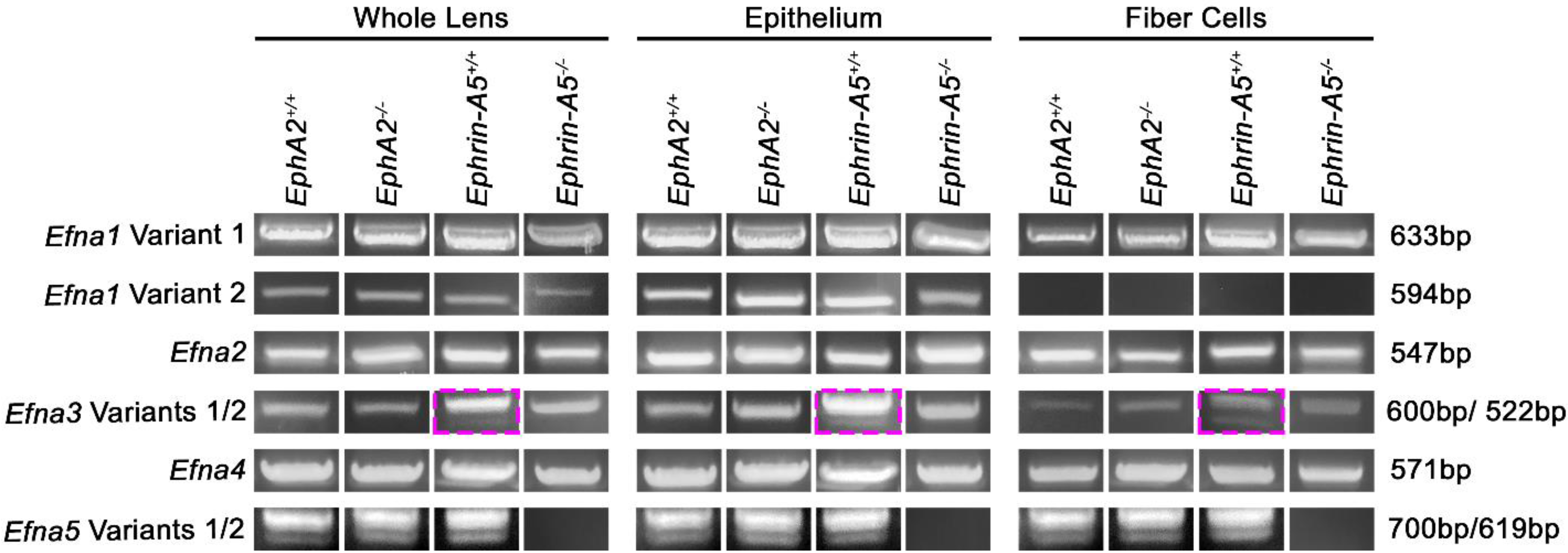
3.3. Ephrin-A Transcripts in the Lens
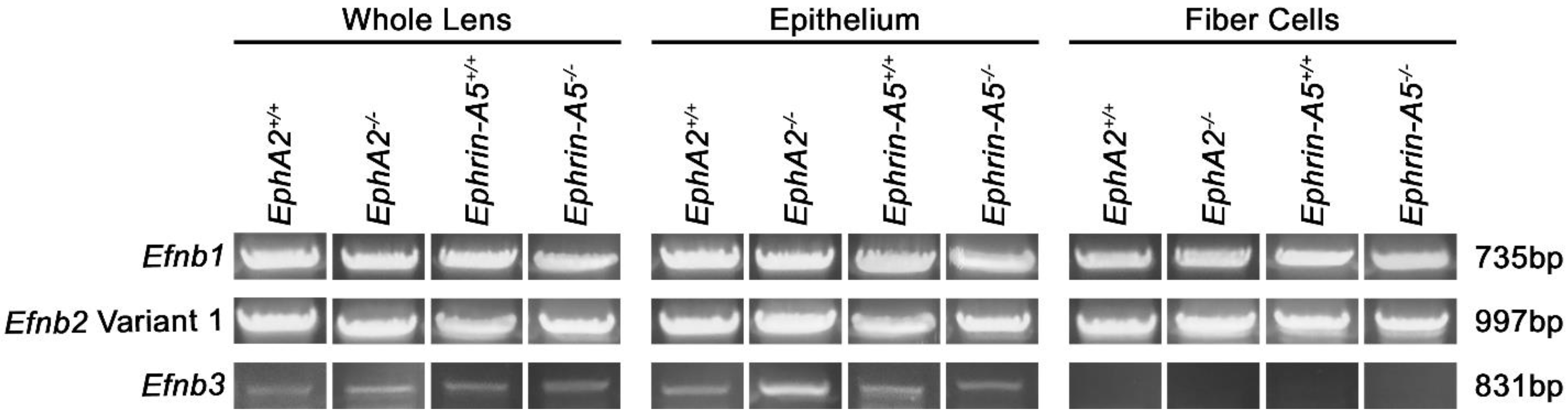
3.4. Ephrin-Bs Transcripts in the Lens
3.5. Data Comparison with Adult Lens Microarray Data
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lovicu, F.J.; Robinson, M.L. Development of the Ocular Lens; Cambridge University Press: Cambridge, UK, 2004; 398 p. [Google Scholar]
- WHO, World Health Organization: World Report on Vision. 2019. Available online: https://www.who.int/publications/i/item/9789241516570 (accessed on 8 October 2019).
- Berry, V.; Pontikos, N.; Albarca-Aguilera, M.; Plagnol, V.; Massouras, A.; Prescott, D.; Moore, A.T.; Arno, G.; Cheetham, M.E.; Michaelides, M. A recurrent splice-site mutation in EPHA2 causing congenital posterior nuclear cataract. Ophthalmic Genet. 2017, 39, 236–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dave, A.; Laurie, K.; Staffieri, S.E.; Taranath, D.; Mackey, D.A.; Mitchell, P.; Wang, J.J.; Craig, J.; Burdon, K.P.; Sharma, S. Mutations in the EPHA2 Gene Are a Major Contributor to Inherited Cataracts in South-Eastern Australia. PLoS ONE 2013, 8, e72518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaul, H.; Riazuddin, S.A.; Shahid, M.; Kousar, S.; Butt, N.H.; Zafar, A.U.; Khan, S.N.; Husnain, T.; Akram, J.; Hejtmancik, J.F.; et al. Autosomal recessive congenital cataract linked to EPHA2 in a consanguineous Pakistani family. Mol. Vis. 2010, 16, 511–517. [Google Scholar] [PubMed]
- Li, D.; Wang, S.; Ye, H.; Tang, Y.; Qiu, X.; Fan, Q.; Rong, X.; Liu, X.; Chen, Y.; Yang, J.; et al. Distribution of gene mutations in sporadic congenital cataract in a Han Chinese population. Mol. Vis. 2016, 22, 589–598. [Google Scholar]
- Park, J.E.; Son, A.I.; Hua, R.; Wang, L.; Zhang, X.; Zhou, R. Human Cataract Mutations in EPHA2 SAM Domain Alter Receptor Stability and Function. PLoS ONE 2012, 7, e36564. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Y.; Zhu, S.; Li, J.; Yao, K. A Novel Human Congenital Cataract Mutation in EPHA2 Kinase Domain (p.G668D) Alters Receptor Stability and Function. Investig. Opthalmol. Vis. Sci. 2019, 60, 4717–4726. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Hua, R.; Xiao, W.; Burdon, K.P.; Bhattacharya, S.S.; Craig, J.E.; Shang, D.; Zhao, X.; Mackey, D.A.; Moore, A.T.; et al. Mutations of the EPHA2 receptor tyrosine kinase gene cause autosomal dominant congenital cataract. Hum. Mutat. 2009, 30, E603–E611. [Google Scholar] [CrossRef]
- Jun, G.; Guo, H.; Klein, B.E.K.; Klein, R.; Wang, J.J.; Mitchell, P.; Miao, H.; Lee, K.E.; Joshi, T.; Buck, M.; et al. EPHA2 Is Associated with Age-Related Cortical Cataract in Mice and Humans. PLoS Genet. 2009, 5, e1000584. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; Zhou, N.; Zhang, N.; Qi, Y. Mutational Screening of EFNA5 in Chinese Age-Related Cataract Patients. Ophthalmic Res. 2014, 52, 124–129. [Google Scholar] [CrossRef]
- Sundaresan, P.; Ravindran, R.D.; Vashist, P.; Shanker, A.; Nitsch, D.; Talwar, B.; Maraini, G.; Camparini, M.; Nonyane, B.A.S.; Smeeth, L.; et al. EPHA2 Polymorphisms and Age-Related Cataract in India. PLoS ONE 2012, 7, e33001. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.; Hou, S.; Jiang, Z.; Hu, Z.; Yang, P.; Ye, J. Association of EPHA2 polymorphisms and age-related cortical cataract in a Han Chinese population. Mol. Vis. 2011, 17, 1553–1558. [Google Scholar]
- Murugan, S.; Cheng, C. Roles of Eph-Ephrin Signaling in the Eye Lens Cataractogenesis, Biomechanics, and Homeostasis. Front. Cell Dev. Biol. 2022, 10. [Google Scholar] [CrossRef]
- Pitulescu, M.E.; Adams, R.H. Eph/ephrin molecules—a hub for signaling and endocytosis. Genes Dev. 2010, 24, 2480–2492. [Google Scholar] [CrossRef] [Green Version]
- Pasquale, E.B. Eph receptors and ephrins in cancer: Bidirectional signalling and beyond. Nat. Cancer 2010, 10, 165–180. [Google Scholar] [CrossRef] [Green Version]
- Darling, T.K.; Lamb, T.J. Emerging Roles for Eph Receptors and Ephrin Ligands in Immunity. Front. Immunol. 2019, 10, 1473. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-Q.; Takakura, N.; Oike, Y.; Inada, T.; Gale, N.W.; Yancopoulos, G.D.; Suda, T. Stromal cells expressing ephrin-B2 promote the growth and sprouting of ephrin-B2+ endothelial cells. Blood 2001, 98, 1028–1037. [Google Scholar] [CrossRef] [Green Version]
- Davy, A.; Robbins, S.M. Ephrin-A5 modulates cell adhesion and morphology in an integrin-dependent manner. EMBO J. 2000, 19, 5396–5405. [Google Scholar] [CrossRef] [Green Version]
- Davy, A.; Gale, N.W.; Murray, E.W.; Klinghoffer, R.A.; Soriano, P.; Feuerstein, C.; Robbins, S.M. Compartmentalized signaling by GPI-anchored ephrin-A5 requires the Fyn tyrosine kinase to regulate cellular adhesion. Genes Dev. 1999, 13, 3125–3135. [Google Scholar] [CrossRef] [Green Version]
- Holmberg, J.; Clarke, D.L.; Frisén, J. Regulation of repulsion versus adhesion by different splice forms of an Eph receptor. Nat. 2000, 408, 203–206. [Google Scholar] [CrossRef]
- Lisabeth, E.M.; Falivelli, G.; Pasquale, E.B. Eph Receptor Signaling and Ephrins. Cold Spring Harb. Perspect. Biol. 2013, 5, a009159. [Google Scholar] [CrossRef] [Green Version]
- Noberini, R.; de la Torre, E.R.; Pasquale, E.B. Profiling Eph receptor expression in cells and tissues: A targeted mass spec-trometry approach. Cell Adh. Migr. 2012, 6, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Pasquale, E.B. Eph–ephrin promiscuity is now crystal clear. Nat. Neurosci. 2004, 7, 417–418. [Google Scholar] [CrossRef]
- Pasquale, E.B. Eph–receptor signalling casts a wide net on cell behaviour. Nat. Rev. Mol. Cell Biol. 2005, 6, 462–475. [Google Scholar] [CrossRef]
- Himanen, J.-P.; Saha, N.; Nikolov, D.B. Cell–cell signaling via Eph receptors and ephrins. Curr. Opin. Cell Biol. 2007, 19, 534–542. [Google Scholar] [CrossRef] [Green Version]
- Kullander, K.; Klein, R. Mechanisms and functions of eph and ephrin signalling. Nat. Rev. Mol. Cell Biol. 2002, 3, 475–486. [Google Scholar] [CrossRef]
- Piatigorsky, J. Lens differentiation in vertebrates. A review of cellular and molecular features. Differentiation 1981, 19, 134–153. [Google Scholar] [CrossRef]
- Kuszak, J.R.; Mazurkiewicz, M.; Jison, L.; Madurski, A.; Ngando, A.; Zoltoski, R.K. Quantitative analysis of animal model lens anatomy: Accommodative range is related to fiber structure and organiza-tion. Vet Ophthalmol 2006, 9, 266–280. [Google Scholar] [CrossRef]
- Kuszak, J.; Zoltoski, R.; Sivertson, C. Fibre cell organization in crystalline lenses. Exp. Eye Res. 2003, 78, 673–687. [Google Scholar] [CrossRef]
- Cheng, C.; Ansari, M.M.; Cooper, J.A.; Gong, X. EphA2 and Src regulate equatorial cell morphogenesis during lens development. Development 2013, 140, 4237–4245. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Fowler, V.M.; Gong, X. EphA2 and ephrin-A5 are not a receptor-ligand pair in the ocular lens. Exp. Eye Res. 2017, 162, 9–17. [Google Scholar] [CrossRef]
- Zhou, Y.; Shiels, A. Epha2 and Efna5 participate in lens cell pattern-formation. Differentiation 2018, 102, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C. EphA2 and Ephrin-A5 Guide Eye Lens Suture Alignment and Influence Whole Lens Resilience. Investig. Opthalmol. Vis. Sci. 2021, 62, 3. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Wang, K.; Hoshino, M.; Uesugi, K.; Yagi, N.; Pierscionek, B. EphA2 Affects Development of the Eye Lens Nucleus and the Gradient of Refractive Index. Investig. Opthalmol. Vis. Sci. 2022, 63, 2. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Gong, X. Diverse Roles of Eph/ephrin Signaling in the Mouse Lens. PLoS ONE 2011, 6, e28147. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; De Maria, A.; Bennett, T.; Shiels, A.; Bassnett, S. A Role for Epha2 in Cell Migration and Refractive Organization of the Ocular Lens. Investig. Opthalmol. Vis. Sci. 2012, 53, 551–559. [Google Scholar] [CrossRef] [Green Version]
- Parreno, J.; Emin, G.; Vu, M.P.; Clark, J.T.; Aryal, S.; Patel, S.D.; Cheng, C. Methodologies to unlock the molecular expression and cellular structure of ocular lens epithelial cells. Front. Cell Dev. Biol. 2022, 10, 1827. [Google Scholar] [CrossRef]
- Frisén, J.; Yates, P.A.; McLaughlin, T.; Friedman, G.C.; O’Leary, D.D.; Barbacid, M. Ephrin-A5 (AL-1/RAGS) is essential for proper retinal axon guidance and topographic mapping in the mammalian vis-ual system. Neuron 1998, 20, 235–243. [Google Scholar] [CrossRef] [Green Version]
- Brantley-Sieders, D.M.; Caughron, J.; Hicks, D.; Pozzi, A.; Ruiz, J.C.; Chen, J. EphA2 receptor tyrosine kinase regulates endothelial cell migration and vascular assembly through phosphoinositide 3-kinase-mediated Rac1 GTPase activation. J. Cell Sci. 2004, 117, 2037–2049. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Nowak, R.B.; Amadeo, M.B.; Biswas, S.K.; Lo, W.K.; Fowler, V.M. Tropomyosin 3.5 protects the F-actin networks required for tissue biomechanical properties. J. Cell Sci. 2018, 131, jcs222042. [Google Scholar] [CrossRef] [Green Version]
- Kakrana, A.; Yang, A.; Anand, D.; Djordjevic, D.; Ramachandruni, D.; Singh, A.; Huang, H.; Ho, J.W.K.; A Lachke, S. iSyTE 2.0: A database for expression-based gene discovery in the eye. Nucleic Acids Res. 2017, 46, D875–D885. [Google Scholar] [CrossRef]
- Cheng, C.; Gao, J.; Sun, X.; Mathias, R.T. Eph-ephrin Signaling Affects Eye Lens Fiber Cell Intracellular Voltage and Membrane Conductance. Front. Physiol. 2021, 12, 2072. [Google Scholar] [CrossRef]
- Patel, S.D.; Anand, D.; Motohashi, H.; Katsuoka, F.; Yamamoto, M.; Lachke, S.A. Deficiency of the bZIP transcription factors Mafg and Mafk causes misexpression of genes in distinct pathways and results in lens embryonic developmental defects. Front. Cell Dev. Biol. 2022, 10, 981893. [Google Scholar] [CrossRef]
- Agrawal, S.A.; Anand, D.; Siddam, A.D.; Kakrana, A.; Dash, S.; Scheiblin, D.A.; Dang, C.A.; Terrell, A.M.; Waters, S.M.; Singh, A.; et al. Compound mouse mutants of bZIP transcription factors Mafg and Mafk reveal a regulatory network of non-crystallin genes associated with cataract. Qual. Life Res. 2015, 134, 717–735. [Google Scholar] [CrossRef] [Green Version]
- Anand, D.; Agrawal, S.A.; Slavotinek, A.; Lachke, S.A. Mutation update of transcription factor genes FOXE3, HSF4, MAF, and PITX3 causing cataracts and other develop-mental ocular defects. Hum. Mutat. 2018, 39, 471–494. [Google Scholar]
- Jamieson, R.V.; Perveen, R.; Kerr, B.; Carette, M.; Yardley, J.; Heon, E.; Wirth, M.G.; Van Heyningen, V.; Donnai, D.; Munier, F.; et al. Domain disruption and mutation of the bZIP transcription factor, MAF, associated with cataract, ocular anterior segment dysgenesis and coloboma. Hum. Mol. Genet. 2002, 11, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Kawauchi, S.; Takahashi, S.; Nakajima, O.; Ogino, H.; Morita, M.; Nishizawa, M.; Yasuda, K.; Yamamoto, M. Regulation of Lens Fiber Cell Differentiation by Transcription Factor c-Maf. J. Biol. Chem. 1999, 274, 19254–19260. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.I.; Li, T.; Ho, I.-C.; Grusby, M.J.; Glimcher, L.H. Requirement for the c-Maf transcription factor in crystallin gene regulation and lens development. Proc. Natl. Acad. Sci. 1999, 96, 3781–3785. [Google Scholar] [CrossRef] [Green Version]
- Ring, B.Z.; Cordes, S.P.; A Overbeek, P.; Barsh, G.S. Regulation of mouse lens fiber cell development and differentiation by the Maf gene. Development 2000, 127. [Google Scholar] [CrossRef]
- Parreno, J.; Amadeo, M.B.; Kwon, E.H.; Fowler, V.M. Tropomyosin 3.1 Association With Actin Stress Fibers is Required for Lens Epithelial to Mesenchymal Transition. Investig. Opthalmol. Vis. Sci. 2020, 61, 2. [Google Scholar] [CrossRef]
- Himanen, J.-P.; Chumley, M.J.; Lackmann, M.; Li, C.; A Barton, W.; Jeffrey, P.D.; Vearing, C.; Geleick, D.; A Feldheim, D.; Boyd, A.W.; et al. Repelling class discrimination: Ephrin-A5 binds to and activates EphB2 receptor signaling. Nat. Neurosci. 2004, 7, 501–509. [Google Scholar] [CrossRef]
- Takemoto, M.; Fukuda, T.; Sonoda, R.; Murakami, F.; Tanaka, H.; Yamamoto, N. Ephrin-B3-EphA4 interactions regulate the growth of specific thalamocortical axon populations in vitro. Eur. J. Neurosci. 2002, 16, 1168–1172. [Google Scholar] [CrossRef] [PubMed]
- Gale, N.W.; Holland, S.J.; Valenzuela, D.M.; Flenniken, A.; Pan, L.; Ryan, T.E.; Henkemeyer, M.; Strebhardt, K.; Hirai, H.; Wilkinson, D.; et al. Eph Receptors and Ligands Comprise Two Major Specificity Subclasses and Are Reciprocally Compartmentalized during Embryogenesis. Neuron 1996, 17, 9–19. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Whole Lens | Epithelium | Fiber Cells | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EphA2 | Ephrin-A5 | EphA2 | Ephrin-A5 | EphA2 | Ephrin-A5 | |||||||
| +/+ | -/- | +/+ | -/- | +/+ | -/- | +/+ | -/- | +/+ | -/- | +/+ | -/- | |
| Epha1 | ✓ | ✓ | ✓ | |||||||||
| Epha2 | ✓ | KO | ✓ | ✓ | ✓ | KO | ✓ | ✓ | ✓ | KO | ✓ | ✓ |
| Epha3Variant 1 | ✓ | ✓ | ✓ | |||||||||
| Epha3 Variant 2 | n.d. | n.d. | n.d. | |||||||||
| Epha4 | ✓ | ✓ | ✓ | |||||||||
| Epha5 Variant 1 | n.d. | n.d. | n.d. | |||||||||
| Epha5 Variant 2 | n.d. | n.d. | n.d. | |||||||||
| Epha5 Variant 3 | ✓ | ✓ | ✓ | |||||||||
| Epha5 Variant 4 | n.d. | n.d. | n.d. | |||||||||
| Epha5 Variant 5 | n.d. | n.d. | n.d. | |||||||||
| Epha5 Variant 6 | n.d. | n.d. | n.d. | |||||||||
| Epha5 Variant 7 | n.d. | n.d. | n.d. | |||||||||
| Epha5 Variant 8 | n.d. | n.d. | n.d. | |||||||||
| Epha5 Variant 9 | ✓ | ✓ | ✓ | |||||||||
| Epha5 Variant 10 | n.d. | n.d. | n.d. | |||||||||
| Epha5 Variant 11 | n.d. | n.d. | n.d. | |||||||||
| Epha5Variant 12 | ✓ | ✓ | ✓ | |||||||||
| Epha5 Variant 13 | n.d. | n.d. | n.d. | |||||||||
| Epha5 Variant 14 | ✓ | ✓ | ✓ | |||||||||
| Epha6 | n.d. | n.d. | n.d. | |||||||||
| Epha7 Variant 1 | ✓ | ✓ | n.d. | ✓ | n.d. | ✓ | ||||||
| Epha7 Variant 2 | ✓ | ✓ | ✓ | |||||||||
| Epha7 Variant 3 | n.d. | n.d. | n.d. | |||||||||
| Epha8 | ✓ | ✓ | ✓ | |||||||||
| Epha10 Variant 1 | n.d. | n.d. | n.d. | |||||||||
| Epha10 Variant 2 | n.d. | n.d. | n.d. | |||||||||
| Ephb1 Variant 1 | ✓ | ✓ | ✓ | |||||||||
| Ephb1 Variant 2 | n.d. | n.d. | n.d. | |||||||||
| Ephb2 Variant 1 | n.d. | n.d. | n.d. | |||||||||
| Ephb2 Variant 2 | ✓ | ✓ | ✓ | |||||||||
| Ephb3 | ✓ | ✓ | ✓ | |||||||||
| Ephb4 Variant 1 | ✓ | ✓ | ✓ | |||||||||
| Ephb4 Variant 2 | ✓ | ✓ | ✓ | |||||||||
| Ephb6 Variant 1 | ✓ | ✓ | ✓ | |||||||||
| Ephb6 Variant 2 | ✓ | ✓ | ✓ | |||||||||
| Efna1 Variant 1 | ✓ | ✓ | ✓ | |||||||||
| Efna1 Variant 2 | ✓ | ✓ | n.d. | |||||||||
| Efna2 | ✓ | ✓ | ✓ | |||||||||
| Efna3 Variant 1 * | ✓ | ✓ | ✓ | |||||||||
| Efna3 Variant 2 * | n.d. | n.d. | ✓ | n.d. | n.d. | n.d. | ✓ | n.d. | n.d. | n.d. | ✓ | n.d. |
| Efna4 | ✓ | ✓ | ✓ | |||||||||
| Efna5 Variant 1 | ✓ | ✓ | ✓ | KO | ✓ | ✓ | ✓ | KO | ✓ | ✓ | ✓ | KO |
| Efna5 Variant 2 | ✓ | ✓ | ✓ | KO | ✓ | ✓ | ✓ | KO | ✓ | ✓ | ✓ | KO |
| Efnb1 | ✓ | ✓ | ✓ | |||||||||
| Efnb2 Variant 1 | ✓ | ✓ | ✓ | |||||||||
| Efnb2 Variant 2 | n.d. | n.d. | n.d. | |||||||||
| Efnb3 | ✓ | ✓ | n.d. | |||||||||
| Gene | iSyte 2.0 | NCBI | ||||
|---|---|---|---|---|---|---|
| Affymetrix 430 2.0 | Illumina WG-6 v2.0 | |||||
| P28 Epi | P56 | P30 | P42 | P52 | Other tissues with high expression | |
| Epha1 | ✓ | ✓ | ✓ | ✓ | ✓ | Duodenum, intestines, lung |
| Epha2 | ✓ | ✓ | ✓ | ✓ | ✓ | Duodenum, intestines, lung |
| Epha3 | ✓ | ✓ | ✓ | ✓ | ✓ | Embryonic/adult brain, embryonic limb |
| Epha4 | ✓ | ✓ | ✓ | ✓ | ✓ | Embryonic/adult brain, embryonic limb, heart |
| Epha5 | ✓ | ✓ | ✓ | ✓ | ✓ | Embryonic/adult brain |
| Epha6 | ✓ | ✓ | n.d. | n.d. | n.d. | Adult brain |
| Epha7 | ✓ | ✓ | ✓ | ✓ | ✓ | Embryonic/adult brain, embryonic limb |
| Epha8 1 | n.d | n.d | n.d | n.d | n.d | Embryonic brain, adult cerebellum |
| Epha10 2 | N/A | N/A | N/A | N/A | N/A | Embryonic/adult brain, testis |
| Ephb1 | ✓ | ✓ | ✓ | ✓ | ✓ | Embryonic/adult brain |
| Ephb2 | ✓ | ✓ | ✓ | ✓ | ✓ | Embryonic brain, adrenal, colon, intestines |
| Ephb3 | ✓ | ✓ | n.d. | n.d. | n.d. | Embryonic limb, colon, lung, stomach |
| Ephb4 | ✓ | ✓ | ✓ | ✓ | ✓ | Embryonic limb, colon, lung, ovary |
| Ephb6 | ✓ | ✓ | ✓ | ✓ | ✓ | Thymus, adult cortex |
| Efna1 | ✓ | ✓ | ✓ | ✓ | ✓ | Duodenum, lung, intestines |
| Efna2 | ✓ | ✓ | n.d. | n.d. | n.d. | Embryonic brain, embryonic liver, ovary |
| Efna3 | ✓ | ✓ | n.d. | n.d. | n.d. | Embryonic brain, embryonic limb, stomach |
| Efna4 | ✓ | ✓ | ✓ | ✓ | ✓ | Embryonic limb, duodenum, ovary |
| Efna5 | ✓ | ✓ | ✓ | ✓ | ✓ | Embryonic brain, embryonic limb, bladder |
| Efnb1 | ✓ | ✓ | ✓ | ✓ | ✓ | Colon, duodenum, lung, ovary |
| Efnb2 | ✓ | ✓ | ✓ | ✓ | ✓ | Colon, lung |
| Efnb3 3 | ✓ | ✓ | ✓ | ✓ | ✓ | Embryonic brain, embryonic limb, heart |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vu, M.P.; Cheng, C. Mapping the Universe of Eph Receptor and Ephrin Ligand Transcripts in Epithelial and Fiber Cells of the Eye Lens. Cells 2022, 11, 3291. https://doi.org/10.3390/cells11203291
Vu MP, Cheng C. Mapping the Universe of Eph Receptor and Ephrin Ligand Transcripts in Epithelial and Fiber Cells of the Eye Lens. Cells. 2022; 11(20):3291. https://doi.org/10.3390/cells11203291
Chicago/Turabian StyleVu, Michael P., and Catherine Cheng. 2022. "Mapping the Universe of Eph Receptor and Ephrin Ligand Transcripts in Epithelial and Fiber Cells of the Eye Lens" Cells 11, no. 20: 3291. https://doi.org/10.3390/cells11203291
APA StyleVu, M. P., & Cheng, C. (2022). Mapping the Universe of Eph Receptor and Ephrin Ligand Transcripts in Epithelial and Fiber Cells of the Eye Lens. Cells, 11(20), 3291. https://doi.org/10.3390/cells11203291