
Evaluation of the Hematological and Serum Biochemistry Parameters in the Pre-Symptomatic and Symptomatic Stages of ALS Disease to Support Early Diagnosis and Prognosis



Abstract
:1. Introduction
2. Methods
2.1. Ethical Statement
2.2. Animal Studies
2.3. Genotyping of SOD1WT and SOD1G93A Rats
2.4. Evaluation of the Hematological and Serum Biochemistry Parameters
2.5. Magpix Luminex Analysis
2.6. Microwave Digestion of Serum Samples and Inductively Coupled Plasma Mass Spectrometry (ICP-MS)
2.7. Statistical Analysis
3. Results
3.1. Animal Weight, Absolute Organ Weight, and Relative Organ Weights as % Body Weight
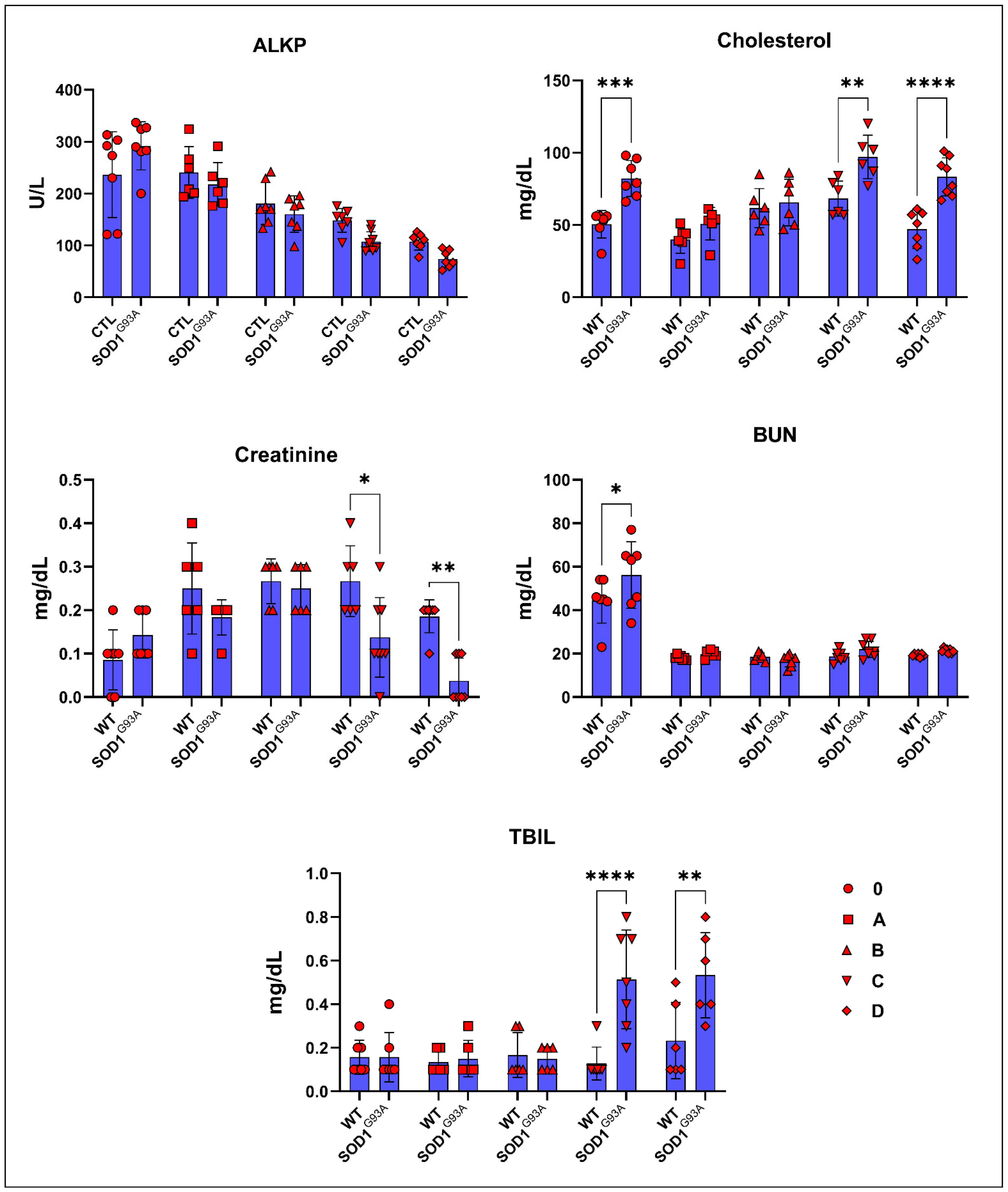
3.2. Serum Biochemistry Parameters of SOD1WT and SOD1G93A Rats
3.3. Serum Hormone and Glucose Levels
3.4. Trace Element and Mineral Levels in the Serum Samples of SOD1WT and SOD1G93A Rats
3.5. Hematological Parameters of SOD1WT and SOD1G93A Rats
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonafede, R.; Mariotti, R. ALS Pathogenesis and Therapeutic Approaches: The Role of Mesenchymal Stem Cells and Extracellular Vesicles. Front. Cell. Neurosci. 2017, 11, 80. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, D.; Ulusu, N.N. Importance of the Serum Biochemical Parameters as Potential Biomarkers for Rapid Diagnosis and Evaluating Preclinical Stage of ALS. Med. Hypotheses 2020, 141, 109736. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.M.; Simmons, Z.; Beard, J.L.; Stephens, H.E.; Connor, J.R. Plasma Biomarkers Associated with ALS and Their Relationship to Iron Homeostasis. Muscle Nerve 2010, 42, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Rothstein, J.D.; Tsai, G.; Kuncl, R.W.; Clawson, L.; Cornblath, D.R.; Drachman, D.B.; Pestronk, A.; Stauch, B.L.; Coyle, J.T. Abnormal Excitatory Amino Acid Metabolism in Amyotrophic Lateral Sclerosis. Ann. Neurol. 1990, 28, 18–25. [Google Scholar] [CrossRef]
- De Vos, K.J.; Grierson, A.J.; Ackerley, S.; Miller, C.C.J. Role of Axonal Transport in Neurodegenerative Diseases. Annu. Rev. Neurosci. 2008, 31, 151–173. [Google Scholar] [CrossRef]
- Rosengren, L.E.; Karlsson, J.-E.; Karlsson, J.-O.; Persson, L.I.; Wikkelsø, C. Patients with Amyotrophic Lateral Sclerosis and Other Neurodegenerative Diseases Have Increased Levels of Neurofilament Protein in CSF. J. Neurochem. 2002, 67, 2013–2018. [Google Scholar] [CrossRef]
- Sun, J.; Carrero, J.J.; Zagai, U.; Evans, M.; Ingre, C.; Pawitan, Y.; Fang, F. Blood Biomarkers and Prognosis of Amyotrophic Lateral Sclerosis. Eur. J. Neurol. 2020, 27, 2125–2133. [Google Scholar] [CrossRef]
- Vu, L.T.; Bowser, R. Fluid-Based Biomarkers for Amyotrophic Lateral Sclerosis. Neurotherapeutics 2017, 14, 119–134. [Google Scholar] [CrossRef] [Green Version]
- Aydemir, D.; Sarayloo, E.; Ulusu, N.N. Rosiglitazone-Induced Changes in the Oxidative Stress Metabolism and Fatty Acid Composition in Relation with Trace Element Status in the Primary Adipocytes. J. Med. Biochem. 2019, 39, 267. [Google Scholar] [CrossRef] [Green Version]
- Aydemir, D.; Simsek, G.; Ulusu, N.N. Dataset of the Analyzing Trace Elements and Minerals via ICP-MS: Method Validation for the Mammalian Tissue and Serum Samples. Data Brief 2020, 29, 105218. [Google Scholar] [CrossRef]
- Fang, F.; Ingre, C.; Roos, P.; Kamel, F.; Piehl, F. Risk Factors for Amyotrophic Lateral Sclerosis. Clin. Epidemiol. 2015, 7, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longinetti, E.; Fang, F. Epidemiology of Amyotrophic Lateral Sclerosis: An Update of Recent Literature. Curr. Opin. Neurol. 2019, 32, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Janse van Mantgem, M.R.; van Eijk, R.P.A.; van der Burgh, H.K.; Tan, H.H.G.; Westeneng, H.-J.; van Es, M.A.; Veldink, J.H.; van den Berg, L.H. Prognostic Value of Weight Loss in Patients with Amyotrophic Lateral Sclerosis: A Population-Based Study. J. Neurol. Neurosurg. Psychiatry 2020, 91, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Moglia, C.; Calvo, A.; Grassano, M.; Canosa, A.; Manera, U.; D’Ovidio, F.; Bombaci, A.; Bersano, E.; Mazzini, L.; Mora, G.; et al. Early Weight Loss in Amyotrophic Lateral Sclerosis: Outcome Relevance and Clinical Correlates in a Population-Based Cohort. J. Neurol. Neurosurg. Psychiatry 2019, 90, 666–673. [Google Scholar] [CrossRef]
- Ngo, S.T.; Steyn, F.J.; Huang, L.; Mantovani, S.; Pfluger, C.M.M.; Woodruff, T.M.; O’Sullivan, J.D.; Henderson, R.D.; McCombe, P.A. Altered Expression of Metabolic Proteins and Adipokines in Patients with Amyotrophic Lateral Sclerosis. J. Neurol. Sci. 2015, 357, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Lazic, S.E.; Semenova, E.; Williams, D.P. Determining Organ Weight Toxicity with Bayesian Causal Models: Improving on the Analysis of Relative Organ Weights. Sci. Rep. 2020, 10, 6625. [Google Scholar] [CrossRef] [Green Version]
- Bharadwaj, S.; Ginoya, S.; Tandon, P.; Gohel, T.D.; Guirguis, J.; Vallabh, H.; Jevenn, A.; Hanouneh, I. Malnutrition: Laboratory Markers vs Nutritional Assessment. Gastroenterol. Rep. 2016, 4, gow013. [Google Scholar] [CrossRef] [Green Version]
- Chiò, A.; Calvo, A.; Bovio, G.; Canosa, A.; Bertuzzo, D.; Galmozzi, F.; Cugnasco, P.; Clerico, M.; De Mercanti, S.; Bersano, E.; et al. Amyotrophic Lateral Sclerosis Outcome Measures and the Role of Albumin and Creatinine. JAMA Neurol. 2014, 71, 1134. [Google Scholar] [CrossRef]
- Ishida, S.; Hashimoto, I.; Seike, T.; Abe, Y.; Nakaya, Y.; Nakanishi, H. Serum Albumin Levels Correlate with Inflammation Rather than Nutrition Supply in Burns Patients: A Retrospective Study. J. Med. Investig. 2014, 61, 361–368. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.F.; Shaw, P.J.; De Vos, K.J. The Role of Mitochondria in Amyotrophic Lateral Sclerosis. Neurosci. Lett. 2019, 710, 132933. [Google Scholar] [CrossRef]
- Lương, K.; Nguyễn, L.T.H. Roles of Vitamin D in Amyotrophic Lateral Sclerosis: Possible Genetic and Cellular Signaling Mechanisms. Mol. Brain 2013, 6, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.; Wu, P.; Wen, T.; Jia, R.; Zhang, R.; Jin, J.; Hu, F.; Chen, Q.Y.; Dang, J. Comparative Assessment of Blood Metal/Metalloid Levels, Clinical Heterogeneity, and Disease Severity in Amyotrophic Lateral Sclerosis Patients. Neurotoxicology 2022, 89, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Hu, W.; Xu, L.; Luo, H.; Wang, N.; Zhang, Q. Decreased Serum Creatinine Levels Predict Short Survival in Amyotrophic Lateral Sclerosis. Ann. Clin. Transl. Neurol. 2021, 8, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Lanznaster, D.; Bruno, C.; Bourgeais, J.; Emond, P.; Zemmoura, I.; Lefèvre, A.; Reynier, P.; Eymieux, S.; Blanchard, E.; Vourc’h, P.; et al. Metabolic Profile and Pathological Alterations in the Muscle of Patients with Early-Stage Amyotrophic Lateral Sclerosis. Biomedicines 2022, 10, 1307. [Google Scholar] [CrossRef]
- Abdel-Khalik, J.; Yutuc, E.; Crick, P.J.; Gustafsson, J.-Å.; Warner, M.; Roman, G.; Talbot, K.; Gray, E.; Griffiths, W.J.; Turner, M.R.; et al. Defective Cholesterol Metabolism in Amyotrophic Lateral Sclerosis. J. Lipid Res. 2017, 58, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Dodge, J.C.; Yu, J.; Sardi, S.P.; Shihabuddin, L.S. Sterol Auto-Oxidation Adversely Affects Human Motor Neuron Viability and Is a Neuropathological Feature of Amyotrophic Lateral Sclerosis. Sci. Rep. 2021, 11, 803. [Google Scholar] [CrossRef]
- González De Aguilar, J.-L. Lipid Biomarkers for Amyotrophic Lateral Sclerosis. Front. Neurol. 2019, 10, 284. [Google Scholar] [CrossRef] [Green Version]
- Karajibani, M.; Montazerifar, F.; Hosseini, R.; Suni, F.; Dashipour, A.R.; Fadaaeimokhtarkanlo, M. The Relationship Between Malnutrition and Liver Enzymes in Hospitalized Children in Zahedan: A Case-Control Study. Zahedan J. Res. Med. Sci. 2021, 23. [Google Scholar] [CrossRef]
- Jayanti, S.; Moretti, R.; Tiribelli, C.; Gazzin, S. Bilirubin and Inflammation in Neurodegenerative and Other Neurological Diseases. Neuroimmunol. Neuroinflamm. 2020, 7, 92–108. [Google Scholar] [CrossRef]
- Tefera, T.W.; Steyn, F.J.; Ngo, S.T.; Borges, K. CNS Glucose Metabolism in Amyotrophic Lateral Sclerosis: A Therapeutic Target? Cell Biosci. 2021, 11, 14. [Google Scholar] [CrossRef]
- Jones, A.G.; Hattersley, A.T. The Clinical Utility of C-peptide Measurement in the Care of Patients with Diabetes. Diabet. Med. 2013, 30, 803–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karra, E.; Chandarana, K.; Batterham, R.L. The Role of Peptide YY in Appetite Regulation and Obesity. J. Physiol. 2009, 587, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Sartipy, P.; Loskutoff, D.J. Monocyte Chemoattractant Protein 1 in Obesity and Insulin Resistance. Proc. Natl. Acad. Sci. USA 2003, 100, 7265–7270. [Google Scholar] [CrossRef] [Green Version]
- Greco, A.; Chiesa, M.R.; Da Prato, I.; Romanelli, A.M.; Dolciotti, C.; Cavallini, G.; Masciandaro, S.M.; Scilingo, E.P.; Del Carratore, R.; Bongioanni, P. Using Blood Data for the Differential Diagnosis and Prognosis of Motor Neuron Diseases: A New Dataset for Machine Learning Applications. Sci. Rep. 2021, 11, 3371. [Google Scholar] [CrossRef] [PubMed]
- Åberg, M.; Nyberg, J.; Robertson, J.; Kuhn, G.; Schiöler, L.; Nissbrandt, H.; Waern, M.; Torén, K. Risk Factors in Swedish Young Men for Amyotrophic Lateral Sclerosis in Adulthood. J. Neurol. 2018, 265, 460–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Q.-Q.; Chen, Y.; Cao, B.; Ou, R.W.; Zhang, L.; Hou, Y.; Gao, X.; Shang, H. Blood Hemoglobin A1c Levels and Amyotrophic Lateral Sclerosis Survival. Mol. Neurodegener. 2017, 12, 69. [Google Scholar] [CrossRef] [Green Version]
- McCombe, P.A.; Lee, J.D.; Woodruff, T.M.; Henderson, R.D. The Peripheral Immune System and Amyotrophic Lateral Sclerosis. Front. Neurol. 2020, 11, 279. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Gascon, R.; Miller, R.G.; Gelinas, D.F.; Mass, J.; Hadlock, K.; Jin, X.; Reis, J.; Narvaez, A.; McGrath, M.S. Evidence for Systemic Immune System Alterations in Sporadic Amyotrophic Lateral Sclerosis (SALS). J. Neuroimmunol. 2005, 159, 215–224. [Google Scholar] [CrossRef]
- Chiu, I.M.; Chen, A.; Zheng, Y.; Kosaras, B.; Tsiftsoglou, S.A.; Vartanian, T.K.; Brown, R.H.; Carroll, M.C. T Lymphocytes Potentiate Endogenous Neuroprotective Inflammation in a Mouse Model of ALS. Proc. Natl. Acad. Sci. USA 2008, 105, 17913–17918. [Google Scholar] [CrossRef] [Green Version]
- Pierce, C.N.; Larson, D.F. Inflammatory Cytokine Inhibition of Erythropoiesis in Patients Implanted with a Mechanical Circulatory Assist Device. Perfusion 2005, 20, 83–90. [Google Scholar] [CrossRef]
- Aydemir, D.; Öztaşcı, B.; Barlas, N.; Ulusu, N.N. Effects of Butylparaben on Antioxidant Enzyme Activities and Histopathological Changes in Rat Tissues. Arch. Ind. Hyg. Toxicol. 2019, 70, 315–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydemir, D.; Hashemkhani, M.; Acar, H.Y.; Ulusu, N.N. Evaluation of the Biocompatibility of the GSH-Coated Ag2S Quantum Dots in Vitro: A Perfect Example for the Non-Toxic Optical Probes. Mol. Biol. Rep. 2020, 47, 4117–4129. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, M.; Aydemir, D.; Bacanlı, M.; Anlar, H.G.; Ulusu, N.N.; Aksoy, Y. Protective Effects of Antioxidant Chlorophyllin in Chemically Induced Breast Cancer Model In Vivo. Biol. Trace Elem. Res. 2021, 199, 4475–4488. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, D.; Karabulut, G.; Şimşek, G.; Gok, M.; Barlas, N.; Ulusu, N.N. Impact of the Di(2-Ethylhexyl) Phthalate Administration on Trace Element and Mineral Levels in Relation of Kidney and Liver Damage in Rats. Biol. Trace Elem. Res. 2018, 186, 474–488. [Google Scholar] [CrossRef]
- Aydemir, D.; Ulusu, N.N. Comment on the: Molecular mechanism of CAT and SOD activity change under MPA-CdTe quantum dots induced oxidative stress in the mouse primary hepatocytes (Spectrochim Acta A Mol Biomol Spectrosc. 2019 Sep 5; 220:117104). Spectrochim. Acta-A Mol. Biomol. Spectrosc. 2020, 229, 117792. [Google Scholar] [CrossRef]
- Fracchiolla, G.; Laghezza, A.; Piemontese, L.; Tortorella, P.; Mazza, F.; Montanari, R.; Pochetti, G.; Lavecchia, A.; Novellino, E.; Pierno, S.; et al. New 2-Aryloxy-3-Phenyl-Propanoic Acids as Peroxisome Proliferator-Activated Receptors α/γ Dual Agonists with Improved Potency and Reduced Adverse Effects on Skeletal Muscle Function. J. Med. Chem. 2009, 52, 6382–6393. [Google Scholar] [CrossRef]
- Camerino, G.M.; Fonzino, A.; Conte, E.; de Bellis, M.; Mele, A.; Liantonio, A.; Tricarico, D.; Tarantino, N.; Dobrowolny, G.; Musarò, A.; et al. Elucidating the Contribution of Skeletal Muscle Ion Channels to Amyotrophic Lateral Sclerosis in Search of New Therapeutic Options. Sci. Rep. 2019, 9, 3185. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serum Biochemistry Parameters | ||||||||
|---|---|---|---|---|---|---|---|---|
| Brain | Lung | Liver | Spleen | Heart | Kidney | Testis | ||
| Group 0 | WT | 0.828 ± 0.119 | 0.609 ± 0.047 | 4.752 ± 0.875 | 0.236 ± 0.021 | 0.356 ± 0.035 | 0.832 ± 0.071 | 1.291± 0.082 |
| G93A | 0.923 ± 0.164 | 0.660 ± 0.040 | 4.057 ± 0.595 | 0.246 ± 0.018 | 0.367 ± 0.033 | 0.856 ± 0.034 | 1.226 ± 0.029 | |
| Group A | WT | 0.607 ± 0.045 | 0.505 ± 0.037 | 4.196 ± 0.443 | 0.203 ± 0.030 | 0.338 ± 0.016 | 0.803 ± 0.093 | 1.028 ± 0.063 |
| G93A | 0.578 ± 0.084 | 0.476 ± 0.072 | 4.292 ± 0.595 | 0.198 ± 0.020 | 0.329 ± 0.013 | 0.761 ± 0.078 | 1.073 ± 0.100 | |
| Group B | WT | 0.502 ± 0.041 | 0.449 ± 0.078 | 3.903 ± 0.194 | 0.171 ± 0.022 | 0.317 ± 0.026 | 0.724 ± 0.057 | 0.854 ± 0.069 |
| G93A | 0.473 ± 0.053 | 0.486 ± 0.056 | 3.671 ± 0.427 | 0.189 ± 0.059 | 0.284 ± 0.031 | 0.688 ± 0.225 | 0.824 ± 0.054 | |
| Group C | WT | 0.432 ± 0.061 | 0.526 ± 0.103 | 3.528 ± 0.195 | 0.156 ± 0.010 | 0.278 ± 0.014 | 0.657 ± 0.049 | 0.793 ± 0.111 |
| G93A | 0.561 ± 0.075 b | 0.601 ± 0.088 | 3.977 ± 0.316 b | 0.164 ± 0.015 | 0.333 ± 0.021 c | 0.755 ± 0.049 c | 0.876 ± 0.128 | |
| Group D | WT | 0.429 ± 0.042 | 0.518 ± 0.096 | 3.355 ± 0.273 | 0.163 ± 0.030 | 0.297 ± 0.021 | 0.632 ± 0.038 | 0.795 ± 0.101 b |
| G93A | 0.657 ± 0.119 c | 0.660 ± 0.125 a | 3.886 ± 0.445 a | 0.164 ± 0.022 | 0.341 ± 0.026 c | 0.872 ± 0.084 c | 1.06 ± 0.217 | |
| Serum Biochemistry Parameters | |||||||
|---|---|---|---|---|---|---|---|
| Albumin (g/dL) | ALT (U/L) | Amylase (U/L) | Calcium (mg/dL) | Globulin (g/dL) | TP (g/dL) | ||
| Group 0 | WT | 3.11 ± 0.76 | 53.50 ± 15.33 | 1327 ± 256.8 | 10.39 ± 0.42 | 2.657 ± 0.67 | 5.743 ± 1.43 |
| G93A | 2.80 ± 0.08 | 57.00 ± 6.30 | 1249 ± 298.9 | 9.957 ± 0.37 | 2.443 ± 0.20 | 5.214 ± 0.25 | |
| Group A | WT | 2.80 ± 0.35 | 76.20 ± 24.16 | 1653 ± 390.4 | 10.72 ± 0.94 | 2.90 ± 0.45 | 5.700 ± 0.71 |
| G93A | 2.61 ± 0.02 | 69.83 ± 10.03 | 1624 ± 284.7 | 10.06 ± 0.11 | 2.483 ± 0.18 | 5.100 ± 0.43 | |
| Group B | WT | 2.71 ± 0.24 | 77.33 ± 12.33 | 1556 ± 129.4 | 10.08 ± 0.23 | 2.90 ± 0.12 | 5.617 ± 0.34 |
| G93A | 3.17 ± 1.30 | 74.17 ± 19.76 | 1516 ± 302.1 | 10.23 ± 0.47 | 2.933 ± 0.47 | 5.667± 0.25 | |
| Group C | WT | 2.80 ± 0.23 | 65.60 ± 13.83 | 1555 ± 136.6 | 9.980 ± 0.32 | 2.880 ± 0.30 | 5.680 ± 0.47 |
| G93A | 2.98 ± 0.22 | 65.11 ± 8.22 | 1467 ± 213.1 | 10.40 ± 0.56 | 3,170 ± 0.23 | 6.060 ± 0.27 | |
| Group D | WT | 2.86 ± 0.05 | 58.67 ± 19.35 | 1559 ± 227.2 | 10.00 ± 0.43 | 2.800 ± 0.43 | 5.583 ± 0.31 |
| G93A | 3.02 ± 0.25 | 59.20 ± 13.75 | 1366 ± 387.4 | 10.35 ± 0.57 | 2.810 ± 0.35 | 5.820 ± 0.46 | |
| Hematological Parameters | ||||||
|---|---|---|---|---|---|---|
| RBC (10−6/µL) | HGB (g/dL) | MCH (pg) | MPV (fL) | PDW (%) | ||
| Group 0 | WT | 7.024 ± 1.26 | 13.58 ± 3.03 | 19.10 ± 1.21 | 7.280 ± 0.28 | 8.320 ± 0.81 |
| G93A | 6.528 ± 1.15 | 13.28 ± 2.73 | 16.77 ± 5.02 | 7.967 ± 0.70 | 9.083 ± 1.38 | |
| Group A | WT | 7.640 ± 1.62 | 14.66 ± 3.77 | 18.99 ± 1.07 | 8.286 ± 1.48 | 8.743 ± 1.01 |
| G93A | 7.475 ± 0.80 | 14.18 ± 2.03 | 17.64 ± 2.90 | 8.960 ± 1.89 | 8.620 ± 0.94 | |
| Group B | WT | 9.156 ± 1.86 | 17.63 ± 3.51 | 19.11 ± 0.94 | 8.600 ± 1.42 | 8.829 ± 0.87 |
| G93A | 9.109 ± 1.24 | 16.77 ± 2.96 | 18.61 ± 1.06 | 8.500 ± 1.62 | 8.914 ± 1.69 | |
| Group C | WT | 8.926 ± 0.35 | 16.14 ± 0.83 | 18.02 ± 0.57 | 10.32 ± 2.89 | 10.42 ± 2.59 |
| G93A | 8.416 ± 0.78 | 15.22 ± 2.21 | 17.96 ± 1.09 | 9.120 ± 1.11 | 8.720 ± 1.04 | |
| Group D | WT | 7.803 ± 1.38 | 14.48 ± 3.03 | 29.53 ± 1.99 | 9.700 ± 2.67 | 9.967 ± 2.62 |
| G93A | 8.151 ± 1.25 | 15.15 ± 3.05 | 31.69 ± 2.07 | 9.225 ± 1.58 | 8.788 ± 0.90 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aydemir, D.; Surucu, S.; Basak, A.N.; Ulusu, N.N. Evaluation of the Hematological and Serum Biochemistry Parameters in the Pre-Symptomatic and Symptomatic Stages of ALS Disease to Support Early Diagnosis and Prognosis. Cells 2022, 11, 3569. https://doi.org/10.3390/cells11223569
Aydemir D, Surucu S, Basak AN, Ulusu NN. Evaluation of the Hematological and Serum Biochemistry Parameters in the Pre-Symptomatic and Symptomatic Stages of ALS Disease to Support Early Diagnosis and Prognosis. Cells. 2022; 11(22):3569. https://doi.org/10.3390/cells11223569
Chicago/Turabian StyleAydemir, Duygu, Selcuk Surucu, Ayse Nazli Basak, and Nuriye Nuray Ulusu. 2022. "Evaluation of the Hematological and Serum Biochemistry Parameters in the Pre-Symptomatic and Symptomatic Stages of ALS Disease to Support Early Diagnosis and Prognosis" Cells 11, no. 22: 3569. https://doi.org/10.3390/cells11223569
APA StyleAydemir, D., Surucu, S., Basak, A. N., & Ulusu, N. N. (2022). Evaluation of the Hematological and Serum Biochemistry Parameters in the Pre-Symptomatic and Symptomatic Stages of ALS Disease to Support Early Diagnosis and Prognosis. Cells, 11(22), 3569. https://doi.org/10.3390/cells11223569