
Natural Cross-Kingdom Spread of Apple Scar Skin Viroid from Apple Trees to Fungi


 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
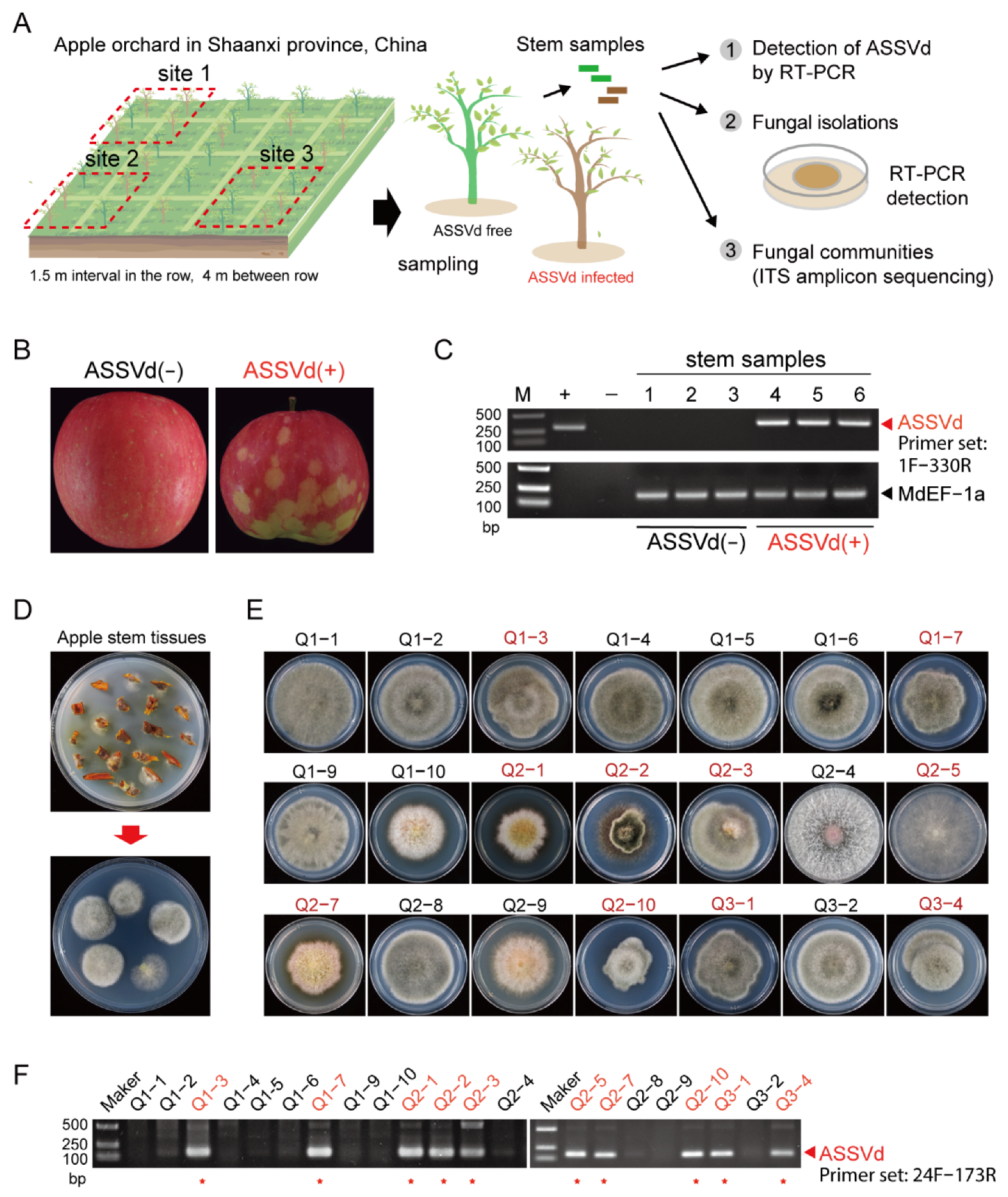
2.1. Collection of Plant Samples
2.2. RNA Extraction, RT-PCR and Sequencing
2.3. DNA Extraction
2.4. Fungal Isolation and Single Spore Preparation
2.5. Fungal Virulence Assay
2.6. Fungal Species Identification
2.7. Fungal Amplicon Sequencing
2.8. Bioinformatics and Statistical Analyses
3. Results
3.1. The Presence of ASSVd in Fungal Isolates
3.2. Phenotypic Effects of ASSVd on Fungal Isolates
3.3. Effect of ASSVd on the Composition of Plant-Associated Fungi
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diener, T. Circular RNAs: Relics of precellular evolution? Proc. Natl. Acad. Sci. USA 1989, 86, 9370–9374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Serio, F.; Flores, R.; Verhoeven, J.T.J.; Li, S.F.; Pallás, V.; Randles, J.W.; Sano, T.; Vidalakis, G.; Owens, R.A. Current status of viroid taxonomy. Arch. Virol. 2014, 159, 3467–3478. [Google Scholar] [CrossRef] [PubMed]
- De la Peña, M.; Cervera, A. Circular RNAs with hammerhead ribozymes encoded in eukaryotic genomes: The enemy at home. Proc. Natl. Acad. Sci. USA 2017, 14, 985–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsagris, E.M.; Martínez de Alba, Á.E.; Gozmanova, M.; Kalantidis, K. Viroids. Cell. Microbiol. 2008, 10, 2168–2179. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, A.; Flores, R.; Randles, J.W.; Palukaitis, P. Viroids and Satellites; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Moelling, K.; Broecker, F.J.I.J.o.M.S. Viroids and the Origin of Life. Int. J. Mol. Sci. 2021, 22, 3476. [Google Scholar] [CrossRef]
- Di Serio, F.; Li, S.F.; Matoušek, J.; Owens, R.A.; Pallás, V.; Randles, J.W.; Sano, T.; Verhoeven, J.T.J.; Vidalakis, G.; Flores, R. ICTV virus taxonomy profile: Avsunviroidae. J. Gen. Virol. 2018, 99, 611–612. [Google Scholar] [CrossRef]
- Di Serio, F.; Owens, R.A.; Li, S.-F.; Matoušek, J.; Pallás, V.; Randles, J.W.; Sano, T.; Verhoeven, J.T.J.; Vidalakis, G.; Flores, R. ICTV virus taxonomy profile: Pospiviroidae. J. Gen. Virol. 2021, 102, 001543. [Google Scholar] [CrossRef]
- Flores, R.; Gas, M.E.; Molina-Serrano, D.; Nohales, M.À.; Carbonell, A.; Gago, S.; De la Peña, M.; Daròs, J.A. Viroid Replication: Rolling-Circles, Enzymes and Ribozymes. Viruses. 2009, 1(2), 317–334. [Google Scholar] [CrossRef] [Green Version]
- Daròs, J.A.; Flores, R. A chloroplast protein binds a viroid RNA in vivo and facilitates its hammerhead mediated self-cleavage. EMBO J. 2002, 21(4), 749–759. [Google Scholar] [CrossRef] [Green Version]
- Flores, R.; Hernández, C.; Alba, A.E.M.D.; Daròs, J.A.; Serio, F.D. Viroids and viroid-host interactions. Annu. Rev. Phytopathol. 2005, 43, 117–139. [Google Scholar] [CrossRef]
- Kovalskaya, N.; Hammond, R.W. Molecular biology of viroid–host interactions and disease control strategies. Plant Sci. 2014, 228, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delan-Forino, C.; Maurel, M.C.; Torchet, C. Replication of avocado sunblotch viroid in the yeast Saccharomyces cerevisiae. J. Virol. 2011, 85, 3229–3238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latifi, A.; Bernard, C. Replication of Avocado Sunblotch Viroid in the Cyanobacterium Nostoc Sp. PCC 7120. J. Plant Pathol. Microbiol. 2016, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Bian, R.; Andika, I.B.; Niu, E.; Liu, Q.; Kondo, H.; Yang, L.; Zhou, H.; Pang, T.; Lian, Z.; et al. Symptomatic plant viroid infections in phytopathogenic fungi. Proc. Natl. Acad. Sci. USA 2019, 116, 13042–13050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afanasenko, O.; Khiutti, A.; Mironenko, N.; Lashina, N. Transmission of potato spindle tuber viroid between Phytophthora infestans and host plants. Vavilov J. Gen. Breed. 2022, 26, 272. [Google Scholar] [CrossRef]
- Müller, D.B.; Vogel, C.; Bai, Y.; Vorholt, J.A. The plant microbiota: Systems-level insights and perspectives. Annu. Rev. Genet. 2016, 50, 211–234. [Google Scholar] [CrossRef] [Green Version]
- Petre, B.; Lorrain, C.; Stukenbrock, E.H.; Duplessis, S. Host-specialized transcriptome of plant-associated organisms. Curr. Opin. Plant Biol. 2020, 56, 81–88. [Google Scholar] [CrossRef]
- Wang, B.; Qiu, Y.L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 2006, 16, 299–363. [Google Scholar] [CrossRef]
- Manoharachary, C.; Mukerji, K.G. Rhizosphere biology—An overview. In Microbial Activity in the Rhizoshere; Springer Science: Berlin/Heidelberg, Germany, 2006; pp. 1–15. [Google Scholar]
- Aziz, U.; Rehmani, M.S.; Wang, L.; Luo, X.; Xian, B.; Wei, S.; Wang, G.; Shu, K. Toward a Molecular Understanding of Rhizosphere, Phyllosphere, and Spermosphere Interactions in Plant Growth and Stress Response. Crit. Rev. Plant Sci. 2021, 40, 479–500. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Xiong, C.; Wei, Z.; Chen, Q.L.; Ma, B.; Zhou, S.Y.D.; Tan, J.; Zhang, L.M.; Cui, H.L.; Duan, G.L. Impacts of global change on the phyllosphere microbiome. New Phytol. 2022, 234, 1977–1986. [Google Scholar] [CrossRef]
- Von Arx, J.A. Plant Pathogenic Fungi; Schweizerbart Science Publications: Stuttgart, Germany, 1987. [Google Scholar]
- Gong, T.; Xin, X.F. Phyllosphere microbiota: Community dynamics and its interaction with plant hosts. J. Integr. Plant Biol. 2021, 63, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Gomes, T.; Pereira, J.A.; Benhadi, J.; Lino-Neto, T.; Baptista, P. Endophytic and epiphytic phyllosphere fungal communities are shaped by different environmental factors in a Mediterranean ecosystem. Microb. Ecol. 2018, 76, 668–679. [Google Scholar] [CrossRef] [Green Version]
- Ohtsuka, Y. A new diseased of apple, on the abnormality of fruit. J. Jpn. Soc. Hortic. Sci. 1935, 6, 44–53. [Google Scholar]
- Hashimoto, J.; Koganezawa, H. Nucleotide sequence and secondary structure of apple scar skin viroid. Nucleic Acids Res. 1987, 15, 7045–7052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadidi, A.; Hansen, A.; Parish, C.; Yang, X. Scar skin and dapple apple viroids are seed-borne and persistent in infected apple trees. Res. Virol. 1991, 142, 289–296. [Google Scholar] [CrossRef]
- Walia, Y.; Dhir, S.; Zaidi, A.A.; Hallan, V. Apple scar skin viroid naked RNA is actively transmitted by the whitefly Trialeurodes vaporariorum. J. RNA Biol. 2015, 12, 1131–1138. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.R.; Lee, S.H.; Lee, D.H.; Kim, J.S.; Park, J.W. Transmission of Apple scar skin viroid by grafting, using contaminated pruning equipment, and planting infected seeds. J. Plant Pathol. J. 2006, 22, 63–67. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.Y.; Lee, H.J.; Kim, N.K.; Oh, J.; Lee, S.H.; Kim, H.; Moon, J.S.; Jeong, R.D. Occurrence pattern of viral infection on pear in korea and genetic characterization of apple scar skin viroid isolates. Hortic. Sci. Technol. 2019, 767–778. [Google Scholar]
- Sun, L.; Suzuki, N. Intragenic rearrangements of a mycoreovirus induced by the multifunctional protein p29 encoded by the prototypic hypovirus CHV1-EP713. RNA 2008, 14, 2557–2571. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Bian, R.; Liu, Q.; Yang, L.; Pang, T.; Salaipeth, L.; Andika, I.B.; Kondo, H.; Sun, L. Identification of a Novel Hypovirulence-Inducing Hypovirus From Alternaria alternata. Front. Microbiol. 2019, 10, 1076. [Google Scholar] [CrossRef] [Green Version]
- Davis, R.H.; De Serres, F.J. Genetic and microbiological research techniques for Neurospora crassa. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1970; Volume 17, pp. 79–143. [Google Scholar]
- Kumar, V.; Soni, R.; Jain, L.; Dash, B.; Goel, R. Endophytic fungi: Recent advances in identification and explorations. In Advances in Endophytic Fungal Research; Springer: Berlin/Heidelberg, Germany, 2019; pp. 267–281. [Google Scholar]
- Hadidi, A.; Barba, M.J.V. Economic impact of pome and stone fruit viruses and viroids. In Virus and Virus-Like Diseases of Pome and Stone Fruits; APS Publications (American Phytopathological Society): Saint Paul, MN, USA, 2011; Volume 1, pp. 1–7. [Google Scholar]
- Op De Beeck, M.; Lievens, B.; Busschaert, P.; Declerck, S.; Vangronsveld, J.; Colpaert, J.V. Comparison and validation of some ITS primer pairs useful for fungal metabarcoding studies. PLoS ONE 2014, 9, e97629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Bian, R.; Andika, I.B.; Niu, E.; Liu, Q.; Kondo, H.; Yang, L.; Zhou, H.; Pang, T.; Lian, Z.; et al. Reply to Serra et al.: Nucleotide substitutions in plant viroid genomes that multiply in phytopathogenic fungi. Proc. Natl. Acad. Sci. USA 2020, 117, 10129–10130. [Google Scholar] [CrossRef]
- Peyambari, M.; Roossinck, M.J. Characterizing mycoviruses. In Plant Pathogenic Fungi and Oomycetes; Springer: Berlin/Heidelberg, Germany, 2018; pp. 13–24. [Google Scholar]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal–plant interface. Nat. Rev. Genet. 2005, 3, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, S.; Badar, U.; Shoeb, E.; Hashim, G.; AbouHaidar, M.; Hefferon, K. An inside look into biological miniatures: Molecular mechanisms of viroids. Int. J. Mol. Sci. 2021, 22, 2795. [Google Scholar] [CrossRef]
- Navarro, B.; Flores, R.; Di Serio, F. Advances in Viroid-Host Interactions. Annu. Rev. Virol. 2021, 8, 305–325. [Google Scholar] [CrossRef]
- Hadidi, A.; Sun, L.; Randles, J.W. Modes of Viroid Transmission. Cells 2022, 11, 719. [Google Scholar] [CrossRef]
- Roossinck, M.J. Viruses in the phytobiome. Opin. Virol. 2019, 37, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Roossinck, M.J. Multiplexed interactions: Viruses of endophytic fungi. Adv. Virus Res. 2013, 86, 37–58. [Google Scholar] [PubMed]
- Andika, I.B.; Wei, S.; Cao, C.; Salaipeth, L.; Kondo, H.; Sun, L.J.P.o.t.N.A.o.S. Phytopathogenic fungus hosts a plant virus: A naturally occurring cross-kingdom viral infection. Proc. Natl. Acad. Sci. USA 2017, 114, 12267–12272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, R.; Andika, I.B.; Pang, T.; Lian, Z.; Wei, S.; Niu, E.; Wu, Y.; Kondo, H.; Liu, X.; Sun, L. Facilitative and synergistic interactions between fungal and plant viruses. Proc. Natl. Acad. Sci. USA 2020, 117, 3779–3788. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Liu, J.; Pang, J.; Kondo, H.; Chi, S.; Zhang, J.; Sun, L.; Andika, I.B. Common but nonpersistent acquisitions of plant viruses by plant-associated fungi. Viruses 2022, 14, 2279. [Google Scholar] [CrossRef]
- Zand Karimi, H.; Baldrich, P.; Rutter, B.D.; Borniego, L.; Zajt, K.K.; Meyers, B.C.; Innes, R.W. Arabidopsis apoplastic fluid contains sRNA-and circular RNA–protein complexes that are located outside extracellular vesicles. The Plant Cell 2022, 34, 1863–1881. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Botella, L.; Suzuki, N. Mycovirus Diversity and Evolution Revealed/Inferred from Recent Studies. Annu. Rev. Phytopathol. 2022, 60, 307–336. [Google Scholar] [CrossRef]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef]
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [Green Version]
- Thomma, B.P. Alternaria spp.: From general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef]
- Logrieco, A.; Moretti, A.; Solfrizzo, M. Alternaria toxins and plant diseases: An overview of origin, occurrence and risks. World Mycotoxin J. 2009, 2, 129–140. [Google Scholar] [CrossRef]
- Akamatsu, H.; Taga, M.; Kodama, M.; Johnson, R.; Otani, H.; Kohmoto, K. Molecular karyotypes for Alternaria plant pathogens known to produce host-specific toxins. Curr. Genet. 1999, 35, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Xiong, C.; Gao, C.; Tsui, C.K.M.; Wang, M.M.; Zhou, X.; Zhang, A.M.; Cai, L. Disease-induced changes in plant microbiome assembly and functional adaptation. Microbiome 2021, 9, 187. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wei, Z.; Friman, V.P.; Gu, S.H.; Wang, X.-F.; Eisenhauer, N.; Yang, T.J.; Ma, J.; Shen, Q.-R.; Xu, Y.-C.J.M. Probiotic diversity enhances rhizosphere microbiome function and plant disease suppression. mBio 2016, 7, e01790. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, P.J.P.; Colaianni, N.R.; Fitzpatrick, C.R.; Dangl, J.L. Beyond pathogens: Microbiota interactions with the plant immune system. Curr. Opin. Microbiol. 2019, 49, 7–17. [Google Scholar] [CrossRef]
- Pereira, A.P.; de Souza, A.J.; de Chaves, M.G.; Fracetto, G.G.; Garcia, K.G.; Paulo Filho, F.; Cardoso, E.J. Mechanisms of the phytomicrobiome for enhancing soil fertility and health. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2021; pp. 1–14. [Google Scholar]
- Fitzpatrick, C.R.; Salas-González, I.; Conway, J.M.; Finkel, O.M.; Gilbert, S.; Russ, D.; Teixeira, P.J.P.L.; Dangl, J.L. The plant microbiome: From ecology to reductionism and beyond. Annu. Rev. Microbiol. 2020, 74, 81–100. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fungal Genus | Fungi Isolated from ASSVd-Infected Plant Tissues | Fungi Isolated from Non-Infected Plant Tissues |
|---|---|---|
| No. Infected/No. Tested Strains | No. Strains | |
| Alternaria tenuissima | 0 | 7 |
| Alternaria compacta | 0 | 42 |
| Alternaria alternata | 77/100 | 192 |
| Alternaria brassicicola | 0 | 10 |
| Epicoccum nigrum | 2/4 | 15 |
| Curvularia spicifera | 0 | 1 |
| Talaromyces verruculosus | 0 | 5 |
| Botryosphaeria dothidea | 1/1 | 2 |
| Diaporthe phaseolorum | 1/10 | 13 |
| Unclassified | 0/2 | 14 |
| Total | 81/117 | 301 |
| Sample | Sobs | Shannon | Simpson | Ace |
|---|---|---|---|---|
| ASSVd (−) | 112.67 ± 9.07 a | 2.16 ± 0.45 a | 0.23 ± 0.10 a | 139.65 ± 13.94 a |
| ASSVd (+) | 122.67 ± 1.53 a | 2.17 ± 0.26 a | 0.21 ± 0.10 a | 151.52 ± 5.78 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, M.; Wei, S.; Bian, R.; Luo, J.; Khan, H.A.; Tai, H.; Kondo, H.; Hadidi, A.; Andika, I.B.; Sun, L. Natural Cross-Kingdom Spread of Apple Scar Skin Viroid from Apple Trees to Fungi. Cells 2022, 11, 3686. https://doi.org/10.3390/cells11223686
Tian M, Wei S, Bian R, Luo J, Khan HA, Tai H, Kondo H, Hadidi A, Andika IB, Sun L. Natural Cross-Kingdom Spread of Apple Scar Skin Viroid from Apple Trees to Fungi. Cells. 2022; 11(22):3686. https://doi.org/10.3390/cells11223686
Chicago/Turabian StyleTian, Mengyuan, Shuang Wei, Ruiling Bian, Jingxian Luo, Haris Ahmed Khan, Huanhuan Tai, Hideki Kondo, Ahmed Hadidi, Ida Bagus Andika, and Liying Sun. 2022. "Natural Cross-Kingdom Spread of Apple Scar Skin Viroid from Apple Trees to Fungi" Cells 11, no. 22: 3686. https://doi.org/10.3390/cells11223686
APA StyleTian, M., Wei, S., Bian, R., Luo, J., Khan, H. A., Tai, H., Kondo, H., Hadidi, A., Andika, I. B., & Sun, L. (2022). Natural Cross-Kingdom Spread of Apple Scar Skin Viroid from Apple Trees to Fungi. Cells, 11(22), 3686. https://doi.org/10.3390/cells11223686