

Wnt Signaling in the Development of Bone Metastasis

{kind=link}
{kind=link}
Abstract
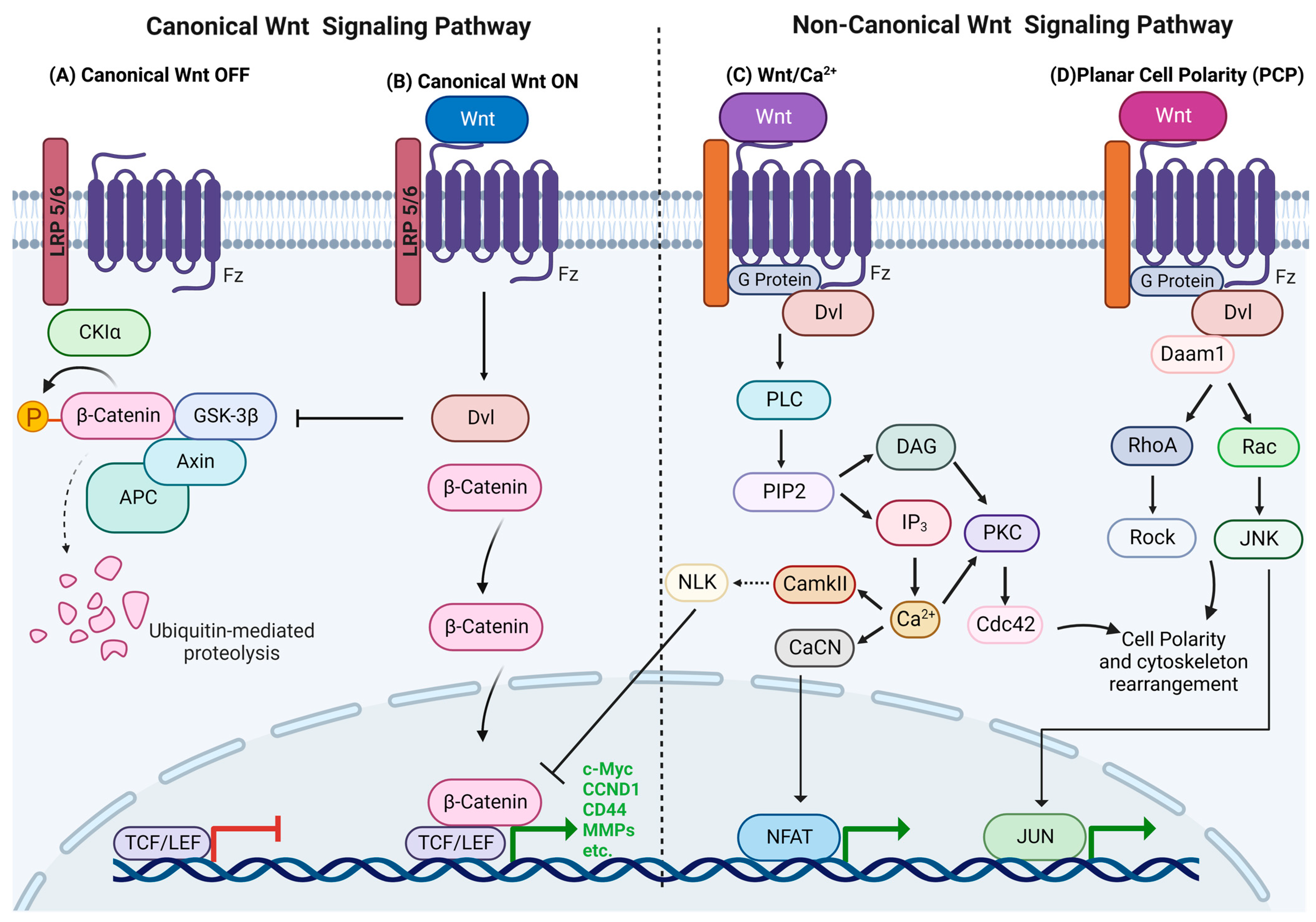
:1. Wnt Signaling
1.1. Canonical Wnt Signaling
1.2. Non-Canonical Wnt Signaling
1.2.1. Wnt/Calcium Pathway
1.2.2. Planar Cell Polarity (PCP)
2. Wnt in Bone Development and Maintenance
3. Bone Metastasis
3.1. Mechanism of Bone Metastasis
3.1.1. Bone Tropism
3.1.2. Process of Bone Metastasis Development
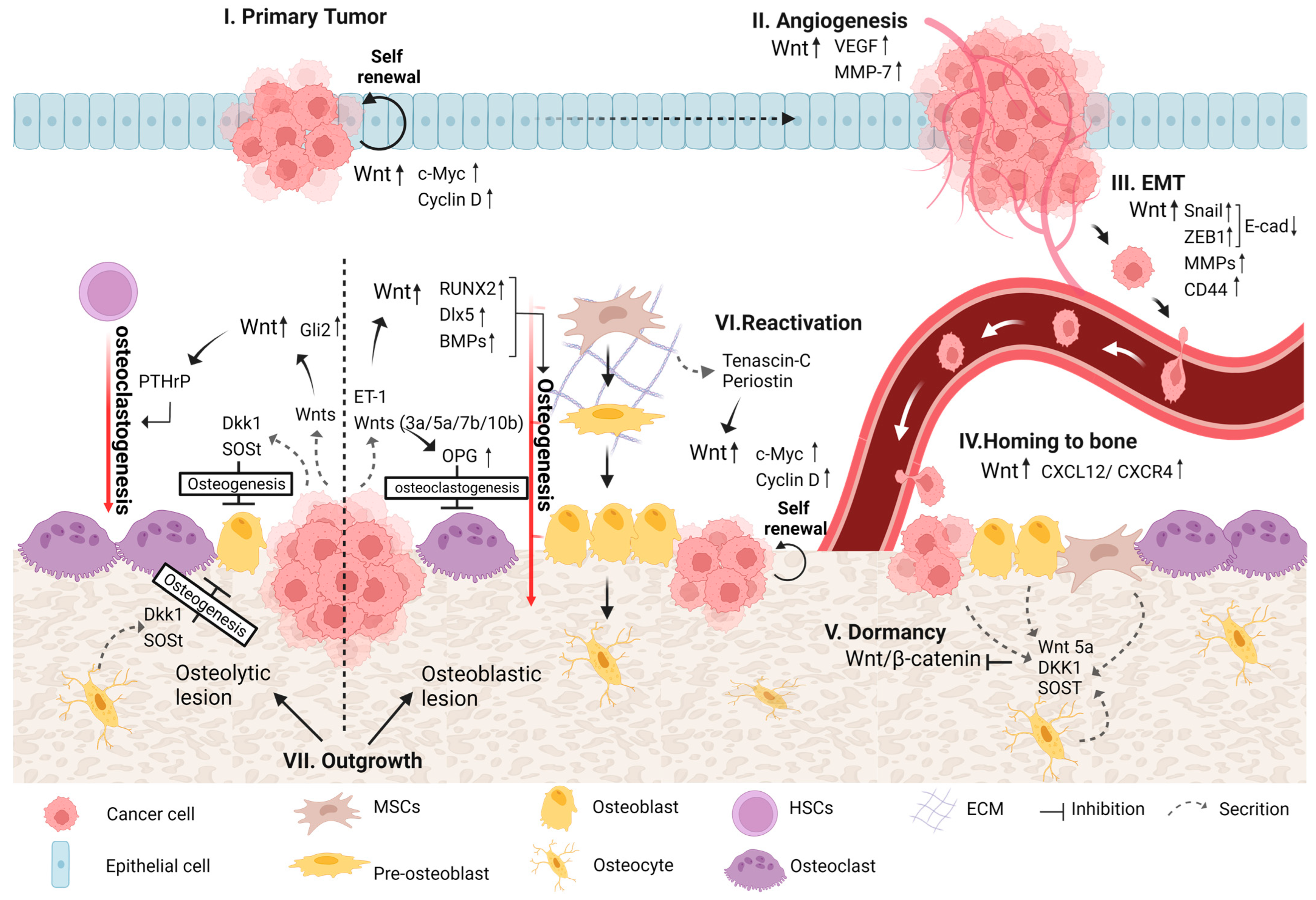
4. Wnt Signaling in Bone Metastasis
4.1. Primary Tumor Development
4.2. Angiogenesis
4.3. EMT
4.4. Homing of DTCs to Bone
4.5. Colonization of DTCs in Bone
4.5.1. Dormancy
4.5.2. Reactivation and Outgrowth
4.5.3. Osteoblastic and Osteolytic Lesions
Osteoblastic Lesion
Osteolytic Lesion
4.5.4. Transmission of Bone Pain
5. Therapeutic Opportunities and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kobayashi, Y.; Uehara, S.; Udagawa, N. Roles of non-canonical Wnt signaling pathways in bone resorption. J. Oral Biosci. 2018, 60, 31–35. [Google Scholar] [CrossRef]
- Qiao, H.; Tang, T. Engineering 3D approaches to model the dynamic microenvironments of cancer bone metastasis. Bone Res. 2018, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Olsen, J.J.; Pohl, S.Ö.-G.; Deshmukh, A.; Visweswaran, M.; Ward, N.C.; Arfuso, F.; Agostino, M.; Dharmarajan, A. The role of Wnt signalling in an-giogenesis. Clin. Biochem. Rev. 2017, 38, 131. [Google Scholar]
- Chen, H.; Liu, H.; Mao, M.; Tan, Y.; Mo, X.; Meng, X.; Cao, M.; Zhong, C.; Liu, Y.; Shan, H.; et al. Crosstalk between autophagy and epithelial-mesenchymal transition and its application in cancer therapy. Mol. Cancer 2019, 18, 101. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, J.; Wald, J.H.; Printsev, I.; Ho, H.-Y.H.; Carraway, K.L. Vangl1 and Vangl2: Planar cell polarity components with a developing role in cancer. Endocrine-Related Cancer 2014, 21, R345–R356. [Google Scholar] [CrossRef] [Green Version]
- Kar, S.; Jasuja, H.; Katti, D.R.; Katti, K.S. Wnt/β-Catenin Signaling Pathway Regulates Osteogenesis for Breast Cancer Bone Metastasis: Experiments in an In Vitro Nanoclay Scaffold Cancer Testbed. ACS Biomater. Sci. Eng. 2019, 6, 2600–2611. [Google Scholar] [CrossRef] [PubMed]
- Zimmerli, D.; Hausmann, G.; Cantù, C.; Basler, K. Pharmacological interventions in the Wnt pathway: Inhibition of Wnt secretion versus disrupting the protein-protein interfaces of nuclear factors. J. Cereb. Blood Flow Metab. 2017, 174, 4600–4610. [Google Scholar] [CrossRef]
- Kim, J.H.; Liu, X.; Wang, J.; Chen, X.; Zhang, H.; Kim, S.H.; Cui, J.; Li, R.; Zhang, W.; Kong, Y.; et al. Wnt signaling in bone formation and its therapeutic potential for bone diseases. Ther. Adv. Musculoskelet. Dis. 2013, 5, 13–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croset, M.; Goehrig, D.; Frackowiak, A.; Bonnelye, E.; Ansieau, S.; Puisieux, A.; Clézardin, P. TWIST1 Expression in Breast Cancer Cells Facilitates Bone Metastasis Formation. J. Bone Miner. Res. 2014, 29, 1886–1899. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K.; Kobayashi, Y.; Koide, M.; Uehara, S.; Okamoto, M.; Ishihara, A.; Kayama, T.; Saito, M.; Marumo, K. The Regulation of Bone Metabolism and Disorders by Wnt Signaling. Int. J. Mol. Sci. 2019, 20, 5525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Wu, D.; Sun, X.; Fan, Y.; Zha, R.; Jalali, A.; Feng, Y.; Li, K.; Sano, T.; Vike, N.; et al. Overexpression of Lrp5 en-hanced the anti-breast cancer effects of osteocytes in bone. Bone Res. 2021, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Cai, T.; Chen, Y. Wnt pathway in osteosarcoma, from oncogenic to therapeutic. J. Cell. Biochem. 2014, 115, 625–631. [Google Scholar] [CrossRef]
- Hoeppner, L.H.; Secreto, F.J.; Westendorf, J.J. Wnt signaling as a therapeutic target for bone diseases. Expert Opin. Ther. Targets 2009, 13, 485–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, K.; Kobayashi, Y.; Udagawa, N.; Uehara, S.; Ishihara, A.; Mizoguchi, T.; Kikuchi, Y.; Takada, I.; Kato, S.; Kani, S.; et al. Wnt5a-Ror2 signaling between osteoblast-lineage cells and osteoclast precursors enhances osteoclastogenesis. Nat. Med. 2012, 18, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Schneider, A.; Jung, Y.; Wang, J.; Dai, J.; Wang, J.; Cook, K.; Osman, N.I.; Koh-Paige, A.J.; Shim, H.; et al. Skeletal localization and neutralization of the SDF-1 (CXCL12)/CXCR4 axis blocks prostate cancer metastasis and growth in osseous sites in vivo. J. Bone Miner. Res. 2005, 20, 318–329. [Google Scholar] [CrossRef]
- Nguyen, D.X.; Chiang, A.C.; Zhang, X.H.-F.; Kim, J.Y.; Kris, M.G.; Ladanyi, M.; Gerald, W.L.; Massagué, J. WNT/TCF signaling through LEF1 and HOXB9 mediates lung adenocarcinoma metastasis. Cell 2009, 138, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Eyre, R.; Alférez, D.G.; Santiago-Gómez, A.; Spence, K.; McConnell, J.C.; Hart, C.; Simões, B.M.; Lefley, D.; Tulotta, C.; Storer, J.; et al. Microenvironmental IL1β promotes breast cancer metastatic colonisation in the bone via activation of Wnt signalling. Nat. Commun. 2019, 10, 5016. [Google Scholar] [CrossRef] [Green Version]
- Gokturk, F.; Erkoc-Kaya, D.; Arikoglu, H. Juglone can inhibit angiogenesis and metastasis in pancreatic cancer cells by tar-geting Wnt/beta-catenin signaling. Bratisl. Med. J. 2021, 122, 132–137. [Google Scholar] [CrossRef]
- Sterling, J.A.; Edwards, J.R.; Martin, T.J.; Mundy, G.R. Advances in the biology of bone metastasis: How the skeleton affects tumor behavior. Bone 2011, 48, 6–15. [Google Scholar] [CrossRef]
- Khoon, M.C.S. Experimental models of bone metastasis: Opportunities for the study of cancer dormancy. Adv. Drug Deliv. Rev. 2015, 94, 141–150. [Google Scholar] [CrossRef]
- Micalizzi, D.S.; Farabaugh, S.M.; Ford, H.L. Epithelial-mesenchymal transition in cancer: Parallels between normal devel-opment and tumor progression. J. Mammary Gland Biol. Neoplasia 2010, 15, 117–134. [Google Scholar] [CrossRef]
- Ke, H.Z.; Richards, W.G.; Li, X.; Ominsky, M.S. Sclerostin and Dickkopf-1 as Therapeutic Targets in Bone Diseases. Endocr. Rev. 2012, 33, 747–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Y.; Chippada-Venkata, U.D.; Oh, W.K. Roles of Matrix Metalloproteinases and Their Natural Inhibitors in Prostate Cancer Progression. Cancers 2014, 6, 1298–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramovitch-Gottlib, L.; Gross, T.; Naveh, D.; Geresh, S.; Rosenwaks, S.; Bar, I.; Vago, R. Low level laser irradiation stimulates osteogenic phenotype of mesenchymal stem cells seeded on a three-dimensional biomatrix. Lasers Med. Sci. 2005, 20, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Bonewald, L.F.; Johnson, M.L. Osteocytes, mechanosensing and Wnt signaling. Bone 2008, 42, 606–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayakumar, S.; Liu, G.; Rus, I.A.; Yao, S.; Chen, Y.; Akiri, G.; Grumolato, L.; Aaronson, S.A. High-Frequency Canonical Wnt Activation in Multiple Sarcoma Subtypes Drives Proliferation through a TCF/β-Catenin Target Gene, CDC25A. Cancer Cell 2011, 19, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, N.; Young, B.; Abramovitz, M.; Bouzyk, M.; Barwick, B.; De, P.; Leyland-Jones, B. Differential Activation of Wnt-β-Catenin Pathway in Triple Negative Breast Cancer Increases MMP7 in a PTEN Dependent Manner. PLoS ONE 2013, 8, e77425. [Google Scholar] [CrossRef] [Green Version]
- Bennett, C.N.; Longo, K.A.; Wright, W.S.; Suva, L.J.; Lane, T.F.; Hankenson, K.D.; MacDougald, O.A. Regulation of osteoblastogenesis and bone mass by Wnt10b. Proc. Natl. Acad. Sci. USA 2005, 102, 3324–3329. [Google Scholar] [CrossRef] [Green Version]
- Mu, J.; Zhu, D.; Shen, Z.; Ning, S.; Liu, Y.; Chen, J.; Li, Y.; Li, Z. The repressive effect of miR-148a on Wnt/β-catenin signaling in-volved in Glabridin-induced anti-angiogenesis in human breast cancer cells. BMC Cancer 2017, 17, 307. [Google Scholar] [CrossRef] [Green Version]
- Port, F.; Basler, K. Wnt Trafficking: New Insights into Wnt Maturation, Secretion and Spreading. Traffic 2010, 11, 1265–1271. [Google Scholar] [CrossRef]
- Shah, K.; Panchal, S.; Patel, B. Porcupine inhibitors: Novel and emerging anti-cancer therapeutics targeting the Wnt signaling pathway. Pharmacol. Res. 2021, 167, 105532. [Google Scholar] [CrossRef] [PubMed]
- Sakunrangsit, N.; Ketchart, W. Plumbagin inhibits cancer stem-like cells, angiogenesis and suppresses cell proliferation and invasion by targeting Wnt/β-catenin pathway in endocrine resistant breast cancer. Pharmacological 2019, 150, 104517. [Google Scholar] [CrossRef] [PubMed]
- Komiya, Y.; Habas, R. Wnt signal transduction pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.-Y.; Li, C.-J.; Yiang, G.-T.; Cheng, Y.-L.; Tsai, A.P.; Hou, Y.-T.; Ho, Y.-C.; Hou, M.-F.; Chu, P.-Y. Molecular Regulation of Bone Metastasis Pathogenesis. Cell. Physiol. Biochem. 2018, 46, 1423–1438. [Google Scholar] [CrossRef]
- Giancotti, F.G. Mechanisms Governing Metastatic Dormancy and Reactivation. Cell 2013, 155, 750–764. [Google Scholar] [CrossRef] [Green Version]
- Baksh, D.; Tuan, R.S. Canonical and non-canonical wnts differentially affect the development potential of primary isolate of human bone marrow mesenchymal stem cells. J. Cell. Physiol. 2007, 212, 817–826. [Google Scholar] [CrossRef]
- Schlange, T.; Matsuda, Y.; Lienhard, S.; Huber, A.; E Hynes, N. Autocrine WNT signaling contributes to breast cancer cell proliferation via the canonical WNT pathway and EGFR transactivation. Breast Cancer Res. 2007, 9, R63. [Google Scholar] [CrossRef] [Green Version]
- Gupta, G.P.; Massagué, J. Cancer Metastasis: Building a Framework. Cell 2006, 127, 679–695. Available online: http://www.sciencedirect.com/science/article/pii/S0092867406014140 (accessed on 1 November 2022). [CrossRef] [Green Version]
- Dey, N.; Barwick, B.G.; Moreno, C.S.; Ordanic-Kodani, M.; Chen, Z.; Oprea-Ilies, G.; Tang, W.; Catzavelos, C.; Kerstann, K.F.; Sledge, G.W.; et al. Wnt signaling in triple negative breast cancer is associated with metastasis. BMC Cancer 2013, 13, 537. [Google Scholar] [CrossRef] [Green Version]
- He, J.-J.; Wang, X.; Liang, C.; Yao, X.; Zhang, Z.-S.; Yang, R.-H.; Fang, D. Wnt5b/Ryk-mediated membrane trafficking of P2X3 receptors contributes to bone cancer pain. Exp. Neurol. 2020, 334, 113482. [Google Scholar] [CrossRef]
- Alliegro, M. Angiogenesis. In xPharm: The Comprehensive Pharmacology Reference; Enna, S.J., Bylund, D.B., Eds.; Elsevier: New York, NY, USA, 2007; pp. 1–6. Available online: http://www.sciencedirect.com/science/article/pii/B9780080552323630193 (accessed on 1 November 2022).
- Chung, R.; Wong, D.; Macsai, C.; Piergentili, A.; Del Bello, F.; Quaglia, W.; Xian, C.J. Roles of Wnt/β-catenin signalling pathway in the bony repair of injured growth plate cartilage in young rats. Bone 2013, 52, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.O.; Johnson, M.L. Wnt Signaling and Bone Cell Activity. In Principles of Bone Biology; Elsevier: Amsterdam, The Netherlands, 2020; 28p. [Google Scholar]
- Nishita, M.; Enomoto, M.; Yamagata, K.; Minami, Y. Cell/tissue-tropic functions of Wnt5a signaling in normal and cancer cells. Trends Cell Biol. 2010, 20, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Simmons, J.K.; Hildreth, B.; Supsavhad, W.; Elshafae, S.M.; Hassan, B.B.; Dirksen, W.P.; Toribio, R.E.; Rosol, T.J. Animal Models of Bone Metastasis. Vet. Pathol. 2015, 52, 827–841. [Google Scholar] [CrossRef] [PubMed]
- Lowy, A.M.; Clements, W.M.; Bishop, J.; Kong, L.; Bonney, T.; Sisco, K.; Aronow, B.; Fenoglio-Preiser, C.; Groden, J. β-Catenin/Wnt Signaling Regulates Expression of the Membrane Type 3 Matrix Metalloproteinase in Gastric Cancer. Cancer Res. 2006, 66, 4734–4741. [Google Scholar] [CrossRef] [Green Version]
- Yuzugullu, H.; Benhaj, K.; Ozturk, N.; Senturk, S.; Celik, E.; Toylu, A.; Tasdemir, N.; Yilmaz, M.; Erdal, E.; Akcali, K.C.; et al. Canonical Wnt signaling is antagonized by noncanonical Wnt5a in hepatocellular carcinoma cells. Mol. Cancer 2009, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, E.G.; Delgado-Calle, J. The Emerging Role of Osteocytes in Cancer in Bone. JBMR Plus 2019, 3, e10186. [Google Scholar] [CrossRef] [Green Version]
- Cong, N.; Du, P.; Zhang, A.; Shen, F.; Su, J.; Pu, P.; Wang, T.; Zjang, J.; Kang, C.; Zhang, Q. Downregulated microRNA-200a promotes EMT and tumor growth through the wnt/β-catenin pathway by targeting the E-cadherin repressors ZEB1/ZEB2 in gastric adenocarcinoma. Oncol. Rep. 2013, 29, 1579–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, M.Q.; Maeda, Y.; Taipaleenmaki, H.; Zhang, W.; Jafferji, M.; Gordon, J.A.R.; Li, Z.; Croce, C.M.; van Wijnen, A.J.; Stein, J.L.; et al. miR-218 Directs a Wnt Signaling Circuit to Promote Differentiation of Osteoblasts and Osteomimicry of Metastatic Cancer Cells. J. Biol. Chem. 2012, 287, 42084–42092. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Ethen, N.J.; Williams, B.O. WNT signaling in bone development and homeostasis. Wiley Interdiscip. Rev. Dev. Biol. 2014, 3, 489–500. [Google Scholar] [CrossRef] [Green Version]
- Rubin, E.M.; Guo, Y.; Tu, K.; Xie, J.; Zi, X.; Hoang, B.H. Wnt Inhibitory Factor 1 Decreases Tumorigenesis and Metastasis in Osteosarcoma. Mol. Cancer Ther. 2010, 9, 731–741. [Google Scholar] [CrossRef] [Green Version]
- Coleman, R. Metastatic bone disease: Clinical features, pathophysiology and treatment strategies. Cancer Treat. Rev. 2001, 27, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, K.; Hase, M.; Zha, R.; Feng, Y.; Li, B.-Y.; Yokota, H. Suppression of breast cancer-associated bone loss with osteoblast proteomes via Hsp90ab1/moesin-mediated inhibition of TGFβ/FN1/CD44 signaling. Theranostics 2022, 12, 929. [Google Scholar] [CrossRef] [PubMed]
- Kerbel, R.S. Tumor angiogenesis. N. Engl. J. Med. 2008, 358, 2039–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, D.; Jones, C.; Rzadzinska, A.; Mark, S.; Zhang, X.; Steel, K.P.; Dai, X.; Chen, P. Wnt5a functions in planar cell polarity regulation in mice. Dev. Biol. 2007, 306, 121–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gay, L.J.; Malanchi, I. The sleeping ugly: Tumour microenvironment’s act to make or break the spell of dormancy. Biochim. Biophys. Acta 2017, 1868, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Decker, K.F.; Zhou, T.; Chen, J.; Qi, Z.; Jacobs, K.; Weilbaecher, K.N.; Corey, E.; Long, F.; Jia, L. Role of WNT7B-induced Noncanonical Pathway in Ad-vanced Prostate CancerWNT7B in Advanced Prostate Cancer. Mol. Cancer Res. 2013, 11, 482–493. [Google Scholar] [CrossRef] [Green Version]
- Nelson, W.J.; Nusse, R. Convergence of Wnt, ß-Catenin, and Cadherin Pathways. Science 2004, 303, 1483–1487. [Google Scholar] [CrossRef] [Green Version]
- Van Camp, J.K.; Beckers, S.; Zegers, D.; Van Hul, W. Wnt Signaling and the Control of Human Stem Cell Fate. Stem Cell Rev. Rep. 2013, 10, 207–229. [Google Scholar] [CrossRef]
- Zhu, M.; Liu, C.; Li, S.; Zhang, S.; Yao, Q.; Song, Q. Sclerostin induced tumor growth, bone metastasis and osteolysis in breast cancer. Sci. Rep. 2017, 7, 11399. [Google Scholar] [CrossRef]
- Chang, S.-C.; Magee, A.I. Acyltransferases for secreted signalling proteins (Review). Mol. Membr. Biol. 2009, 26, 104–113. [Google Scholar] [CrossRef]
- Wang, Y.; Nathans, J. Tissue/planar cell polarity in vertebrates: New insights and new questions. Development 2007, 134, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Subik, K.; Shu, L.; Wu, C.; Liang, Q.; Hicks, D.; Boyce, B.; Schiffhauer, L.; Chen, D.; Chen, C.; Tang, P.; et al. The ubiquitin E3 ligase WWP1 decreases CXCL12-mediated MDA231 breast cancer cell migration and bone metastasis. Bone 2012, 50, 813–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, S.; Sng, N.; Carretero, J.; Welner, R.; Hayashi, Y.; Yamamoto, M.; Tan, A.J.; Yamaguchi, N.; Yasuda, H.; Li, D.; et al. β-catenin contributes to lung tumor de-velopment induced by EGFR mutations. Cancer Res. 2014, 74, 5891–5902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Bhat, R.A.; Seestaller-Wehr, L.M.; Fukayama, S.; Mangine, A.; Moran, R.A.; Komm, B.S.; Bodine, P.V.N.; Billiard, J. The Orphan Receptor Tyrosine Kinase Ror2 Promotes Osteoblast Differentiation and Enhances ex Vivo Bone Formation. Mol. Endocrinol. 2007, 21, 376–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, G.; Nie, J.-J.; Liu, X.; Ding, Z.; Luo, P.; Liu, Y.; Zhang, B.; Wang, R.; Liu, X.; Hai, Y.; et al. Zinc oxide nanoparticles inhibit osteosarcoma metastasis by downregulating β-catenin via HIF-1α/BNIP3/LC3B-mediated mitophagy pathway. Bioact. Mater. 2023, 19, 690–702. Available online: https://www.sciencedirect.com/science/article/pii/S2452199X22002171 (accessed on 1 November 2022). [CrossRef]
- Levinger, I.; Zagouri, R.; Ventura, Y.; Vago, R. Effects of three dimensional microenvironment on tumorigenicity of fibro-sarcoma in vitro. Cancer Stud. Mol. Med.—Open J. 2014, 1, 15–25. [Google Scholar] [CrossRef]
- Baron, R.; Kneissel, M. WNT signaling in bone homeostasis and disease: From human mutations to treatments. Nat. Med. 2013, 19, 179–192. [Google Scholar] [CrossRef]
- Chang, A.-C.; Chen, P.-C.; Lin, Y.-F.; Su, C.-M.; Liu, J.-F.; Lin, T.-H.; Chuang, S.-M.; Tang, C.-H. Osteoblast-secreted WISP-1 promotes adherence of prostate cancer cells to bone via the VCAM-1/integrin α4β1 system. Cancer Lett. 2018, 426, 47–56. [Google Scholar] [CrossRef]
- Basu, S.; Cheriyamundath, S.; Ben-Ze’ev, A. Cell–cell adhesion: Linking Wnt/β-catenin signaling with partial EMT and stemness traits in tumorigenesis. F1000Research 2018, 7, F1000. [Google Scholar] [CrossRef] [Green Version]
- Sottnik, J.L.; Hall, C.L.; Zhang, J.; Keller, E.T. Wnt and Wnt inhibitors in bone metastasis. BoneKEy Rep. 2012, 1, 101. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-K.; Huang, Z.-J.; Liu, S.; Liu, Y.-P.; Song, A.A.; Song, X.-J. WNT signaling underlies the pathogenesis of neuro-pathic pain in rodents. J. Clin. Investig. 2013, 123, 2268–2286. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.G.; Yang, J.; Vazquez, E.S.; Rose, D.; Vakar-Lopez, F.; Mathew, P.; Lopez, A.; Logothetis, C.J.; Lin, S.-H.; Navone, N.M. Low-density lipoprotein receptor-related protein 5 (LRP5) mediates the prostate cancer-induced formation of new bone. Oncogene 2007, 27, 596–603. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Virshup, D.M. Wnt Signaling and Drug Resistance in Cancer. Mol. Pharmacol. 2020, 97, 72–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Kang, Y. Organotropism of Breast Cancer Metastasis. J. Mammary Gland. Biol. Neoplasia 2007, 12, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.R.; Edwards, C.M.; Edwards, J.R. Modeling the Human Bone–Tumor Niche: Reducing and Replacing the Need for Animal Data. JBMR Plus 2020, 4, e10356. [Google Scholar] [CrossRef] [Green Version]
- Halbleib, J.M.; Nelson, W.J. Cadherins in development: Cell adhesion, sorting, and tissue morphogenesis. Genes Dev. 2006, 20, 3199–3214. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, H.; Takahashi, R.-U.; Prieto-Vila, M.; Kawamura, Y.; Kondo, S.; Shirota, T.; Ochiya, T. CD44 exerts a functional role during EMT induction in cisplatin-resistant head and neck cancer cells. Oncotarget 2018, 9, 10029–10041. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Wang, M.; Fu, Y.; Castro, N.J.; Fu, S.W.; Zhang, L.G. Engineering a biomimetic three-dimensional nanostructured bone model for breast cancer bone metastasis study. Acta Biomater. 2014, 14, 164–174. [Google Scholar] [CrossRef]
- Kuhl, M. The WNT/calcium pathway: Biochemical mediators, tools and future requirements. Front. Biosci.-Landmark 2004, 9, 967–974. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, W.; Bado, I.; Zhang, X.H.-F. Bone Tropism in Cancer Metastases. Cold Spring Harb. Perspect. Med. 2019, 10, a036848. [Google Scholar] [CrossRef]
- Hirata, T.; Park, S.C.; Muldong, M.T.; Wu, C.N.; Yamaguchi, T.; Strasner, A.; Raheem, O.; Kumon, H.; Sah, R.L.; Cacalano, N.A.; et al. Specific bone region localization of oste-olytic versus osteoblastic lesions in a patient-derived xenograft model of bone metastatic prostate cancer. Asian J. Urol. 2016, 3, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Saneyoshi, T.; Kume, S.; Amasaki, Y.; Mikoshiba, K. The Wnt/calcium pathway activates NF-AT and promotes ventral cell fate in Xenopus embryos. Nature 2002, 417, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Qi, L.; Liang, Z.; Song, W.; Liu, Y.; Wang, Y.; Sun, B.; Zhang, B.; Cao, W. Transforming growth factor-β1 induces EMT by the transactivation of epidermal growth factor signaling through HA/CD44 in lung and breast cancer cells. Int. J. Mol. Med. 2015, 36, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadanandam, A.; Futakuchi, M.; A Lyssiotis, C.; Gibb, W.J.; Singh, R.K. A Cross-Species Analysis of a Mouse Model of Breast Cancer-Specific Osteolysis and Human Bone Metastases Using Gene Expression Profiling. BMC Cancer 2011, 11, 304. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yang, J.; Bao, M.; Zeng, K.; Fu, S.; Wang, C.; Ye, L. Wnt signaling in bone metastasis: Mechanisms and therapeutic opportunities. Life Sci. 2018, 208, 33–45. [Google Scholar] [CrossRef]
- Kohn, A.D.; Moon, R.T. Wnt and calcium signaling: β-Catenin-independent pathways. Cell Calcium 2005, 38, 439–446. [Google Scholar] [CrossRef]
- Mundy, G. Metastasis to bone: Causes, consequences and therapeutic opportunities. Nat. Rev. Cancer 2002, 2, 584–593. [Google Scholar] [CrossRef]
- Brown, R.L.; Reinke, L.M.; Damerow, M.S.; Perez, D.; Chodosh, L.A.; Yang, J.; Cheng, C. CD44 splice isoform switching in human and mouse epithelium is essential for epithelial-mesenchymal transition and breast cancer progression. J. Clin. Investig. 2011, 121, 1064–1074. [Google Scholar] [CrossRef] [Green Version]
- Chirgwin, J.M.; Guise, T.A. Does prostate-specific antigen contribute to bone metastases? Clin. Cancer Res. 2006, 12, 1395–1397. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Su, Y.; Zhu, H.; Wang, X.; Li, X.; Dai, C.; Xu, C.; Zheng, T.; Mao, C.; Chen, D. Interleukin-23 receptor signaling mediates cancer dormancy and radioresistance in human esophageal squamous carcinoma cells via the Wnt/Notch pathway. J. Mol. Med. 2018, 97, 177–188. [Google Scholar] [CrossRef] [Green Version]
- Easwaran, V.; Lee, S.H.; Inge, L.; Guo, L.; Goldbeck, C.; Garrett, E.; Wiesmann, M.; Garcia, P.D.; Fuller, J.H.; Chan, V.; et al. β-Catenin regulates vascular endothelial growth factor expression in colon cancer. Cancer Res. 2003, 63, 3145–3153. [Google Scholar] [PubMed]
- Taipaleenmäki, H.; Farina, N.H.; van Wijnen, A.J.; Stein, J.L.; Hesse, E.; Stein, G.S.; Lian, J.B. Antagonizing miR-218-5p attenuates Wnt signaling and reduces metastatic bone disease of triple negative breast cancer cells. Oncotarget 2016, 7, 79032–79046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, N.M.; Summers, M.A.; McDonald, M.M. Tumor cell dormancy and reactivation in bone: Skeletal biology and thera-peutic opportunities. JBMR Plus 2019, 3, e10125. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzotta, S.; Neves, C.; Bonner, R.J.; Bernardo, A.S.; Docherty, K.; Hoppler, S. Distinctive Roles of Canonical and Non-canonical Wnt Signaling in Human Embryonic Cardiomyocyte Development. Stem Cell Rep. 2016, 7, 764–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willert, K.; Nusse, R. Wnt proteins. Cold Spring Harbor Perspect. Biol. 2012, 4, a007864. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Park, S.-Y.; Jun, Y.; Kim, J.-Y.; Nam, J.-S. Roles of Wnt Target Genes in the Journey of Cancer Stem Cells. Int. J. Mol. Sci. 2017, 18, 1604. [Google Scholar] [CrossRef] [Green Version]
- Gray, R.S.; Roszko, I.; Solnica-Krezel, L. Planar Cell Polarity: Coordinating Morphogenetic Cell Behaviors with Embryonic Polarity. Dev. Cell 2011, 21, 120–133. [Google Scholar] [CrossRef] [Green Version]
- Ban, J.; Fock, V.; Aryee, D.N.T.; Kovar, H. Mechanisms, Diagnosis and Treatment of Bone Metastases. Cells 2021, 10, 2944. [Google Scholar] [CrossRef]
- Tian, E.; Zhan, F.; Walker, R.; Rasmussen, E.; Ma, Y.; Barlogie, B.; Shaughnessy, J.D., Jr. The Role of the Wnt-Signaling Antagonist DKK1 in the Development of Osteolytic Lesions in Multiple Myeloma. N. Engl. J. Med. 2003, 349, 2483–2494. [Google Scholar] [CrossRef]
- Gross-Aviv, T.; DiCarlo, B.B.; French, M.M.; Athanasiou, K.A.; Vago, R. A study of crystalline biomaterials for articular car-tilage bioengineering. Mater. Sci. Eng. C 2008, 28, 1388–1400. [Google Scholar] [CrossRef]
- Hou, L.; Wang, X.; Zhou, Y.; Ma, H.; Wang, Z.; He, J.; Hu, H.; Guan, W.; Ma, Y. Inhibitory effect and mechanism of mesenchymal stem cells on liver cancer cells. Tumor Biol. 2013, 35, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [Green Version]
- Peinado, H.; Olmeda, D.; Cano, A. Snail, Zeb and bHLH factors in tumour progression: An alliance against the epithelial phenotype? Nat. Rev. Cancer 2007, 7, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Dodge, M.E.; Tang, W.-J.; Lu, J.; Ma, Z.; Fan, C.-W.; Wei, S.; Hao, W.; Kilgore, J.; Williams, N.S.; et al. Small molecule–mediated disruption of Wnt-dependent signaling in tissue regeneration and cancer. Nat. Chem. Biol. 2009, 5, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, A.; Lin, S.-H. Osteolytic and Osteoblastic Bone Metastases: Two Extremes of the Same Spectrum? In Prevention of Bone Metastases; Springer: Berlin/Heidelberg, Germany, 2012; 9p. [Google Scholar]
- Pang, H.; Ma, N.; Shen, W.; Zhao, Q.; Wang, J.; Duan, L.; Chen, W.; Zhang, N.; Zhao, Z.; Liu, L.; et al. Effects of DKK1 overexpression on bone metastasis of SBC-3 cells. Oncol. Lett. 2018, 15, 6739–6744. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, X.; Zhang, H.; Li, X.; Li, X.; Cong, M.; Peng, F.; Yu, J.; Zhang, X.; Yang, Q.; Hu, G. Differential effects on lung and bone metastasis of breast cancer by Wnt signalling inhibitor DKK1. Nature 2017, 19, 1274–1285. [Google Scholar] [CrossRef]
- Nandana, S.; Tripathi, M.; Duan, P.; Chu, C.-Y.; Mishra, R.; Liu, C.; Jin, R.; Yamashita, H.; Zayzafoon, M.; Bhowmick, N.A.; et al. Bone Metastasis of Prostate Cancer Can Be Ther-apeutically Targeted at the TBX2–WNT Signaling AxisTBX2–WNT Signaling Axis in Prostate Cancer Bone Metastasis. Cancer Res. 2017, 77, 1331–1344. [Google Scholar] [CrossRef] [Green Version]
- Ling, L.; Nurcombe, V.; Cool, S.M. Wnt signaling controls the fate of mesenchymal stem cells. Gene 2009, 433, 1–7. [Google Scholar] [CrossRef]
- Hall, C.L.; Keller, E.T. The role of Wnts in bone metastases. Cancer Metastasis Rev. 2006, 25, 551–558. [Google Scholar] [CrossRef]
- Clarke, B.L. Anti-sclerostin antibodies: Utility in treatment of osteoporosis. Maturitas 2014, 78, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Huo, C.; Kao, Y.-H.; Chuu, C.-P. Androgen receptor inhibits epithelial–mesenchymal transition, migration, and invasion of PC-3 prostate cancer cells. Cancer Lett. 2015, 369, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Buczacki, S.J.; Popova, S.; Biggs, E.; Koukorava, C.; Buzzelli, J.; Vermeulen, L.; Hazelwood, L.; Francies, H.; Garnett, M.J.; Winton, D.J. Itraconazole targets cell cycle hetero-geneity in colorectal cancer. J. Exp. Med. 2018, 215, 1891–1912. [Google Scholar] [CrossRef] [PubMed]
- Rosanò, L.; Spinella, F.; Bagnato, A. Endothelin 1 in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer 2013, 13, 637–651. [Google Scholar] [CrossRef] [PubMed]
- Clézardin, P.; Coleman, R.; Puppo, M.; Ottewell, P.; Bonnelye, E.; Paycha, F.; Confavreux, C.B.; Holen, I. Bone metastasis: Mechanisms, therapies, and biomarkers. Physiol. Rev. 2021, 101, 797–855. [Google Scholar] [CrossRef]
- Hu, T.; Li, C. Convergence between Wnt-β-catenin and EGFR signaling in cancer. Mol. Cancer 2010, 9, 236. [Google Scholar] [CrossRef] [Green Version]
- Ren, D.; Dai, Y.; Yang, Q.; Zhang, X.; Guo, W.; Ye, L.; Huang, S.; Chen, X.; Lai, Y.; Du, H.; et al. Wnt5a induces and maintains prostate cancer cells dormancy in bone. J. Exp. Med. 2019, 216, 428–449. [Google Scholar] [CrossRef]
- Peretz, H.; Talpalar, A.E.; Vago, R.; Baranes, D. Superior Survival and Durability of Neurons and Astrocytes on 3-Dimensional Aragonite Biomatrices. Tissue Eng. 2007, 13, 461–472. [Google Scholar] [CrossRef]
- Ben Ghedalia-Peled, N.; Cohen-Erez, I.; Rapaport, H.; Vago, R. Aggressiveness of 4T1 breast cancer cells hampered by Wnt production-2 inhibitor nanoparticles: An in vitro study. Int. J. Pharm. 2021, 596, 120208. [Google Scholar] [CrossRef]
- Herr, P.; Hausmann, G.; Basler, K. WNT secretion and signalling in human disease. Trends Mol. Med. 2012, 18, 483–493. [Google Scholar] [CrossRef]
- Zhou, Y.-Q.; Tian, X.-B.; Tian, Y.-K.; Mei, W.; Liu, D.-Q.; Ye, D.-W. Wnt signaling: A prospective therapeutic target for chronic pain. Pharmacol. Ther. 2022, 231, 107984. Available online: https://www.sciencedirect.com/science/article/pii/S0163725821001868 (accessed on 1 November 2022). [CrossRef] [PubMed]
- Clines, G.A.; Mohammad, K.S.; Bao, Y.; Stephens, O.W.; Suva, L.J.; Shaughnessy, J.D., Jr.; Fox, J.W.; Chirgwin, J.M.; Guise, T.A. Dickkopf Homolog 1 Mediates Endothelin-1-Stimulated New Bone Formation. Mol. Endocrinol. 2007, 21, 486–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, Y.; Chen, Y. Wnt signaling pathway: Implications for therapy in lung cancer and bone metastasis. Cancer Lett. 2014, 353, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.K. A Study of Osteocyte Apoptosis by Region and Quadrant in Murine Cortical Bone. Master’s Thesis, California Polytechnic State University, San Luis Obispo, CA, USA, 2011. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, N.; Kurzrock, R. Targeting the Wnt/beta-catenin pathway in cancer: Update on effectors and inhibitors. Cancer Treat. Rev. 2017, 62, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Polakis, P. Wnt signaling in cancer. Cold Spring Harbor Perspect. Biol. 2012, 4, a008052. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.W.; Merkel, A.R.; Page, J.M.; Ruppender, N.S.; Guelcher, S.A.; Sterling, J.A. Wnt signaling induces gene ex-pression of factors associated with bone destruction in lung and breast cancer. Clin. Exp. Metastasis 2014, 31, 945–959. [Google Scholar] [CrossRef]
- Schlessinger, K.; Hall, A.; Tolwinski, N. Wnt signaling pathways meet Rho GTPases. Genes Dev. 2009, 23, 265–277. [Google Scholar] [CrossRef] [Green Version]
- Sowder, M.E.; Johnson, R.W. Bone as a Preferential Site for Metastasis. JBMR Plus 2018, 3, e10126. [Google Scholar] [CrossRef]
- Simons, M.; Mlodzik, M. Planar Cell Polarity Signaling: From Fly Development to Human Disease. Annu. Rev. Genet. 2008, 42, 517–540. [Google Scholar] [CrossRef]
- Ford, C.; Punnia-Moorthy, G.; Henry, C.; Llamosas, E.; Nixdorf, S.; Olivier, J.; Caduff, R.; Ward, R.; Heinzelmann-Schwarz, V. The non-canonical Wnt ligand, Wnt5a, is upregulated and associated with epithelial to mesenchymal transition in epithelial ovarian cancer. Gynecol. Oncol. 2014, 134, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Azad, B.B.; Nimmagadda, S. The Intricate Role of CXCR4 in Cancer. In Advances in Cancer Research; Elsevier: Amsterdam, The Netherlands, 2014; Volume 124, 52p. [Google Scholar]
- Davis, K.W.; Blankenbaker, D.G. (Eds.) Target Lesion of Bone. In Expertddx: Musculoskeletal, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2018; 4p, Available online: https://www.sciencedirect.com/science/article/pii/B9780323524834501277 (accessed on 1 November 2022).
- Huang, E.H.; Singh, B.; Cristofanilli, M.; Gelovani, J.; Wei, C.; Vincent, L.; Cook, K.R.; Lucci, A. A CXCR4 Antagonist CTCE-9908 Inhibits Primary Tumor Growth and Metastasis of Breast Cancer. J. Surg. Res. 2009, 155, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Takada, I.; Mihara, M.; Suzawa, M.; Ohtake, F.; Kobayashi, S.; Igarashi, M.; Youn, M.-Y.; Takeyama, K.-I.; Nakamura, T.; Mezaki, Y.; et al. A histone lysine methyltransferase activated by non-canonical Wnt signalling suppresses PPAR-γ transactivation. Nature 2007, 9, 1273–1285. [Google Scholar] [CrossRef]
- Yu, B.; Chang, J.; Liu, Y.; Li, J.; Kevork, K.; Al-Hezaimi, K.; Graves, D.T.; Park, N.-H.; Wang, C.-Y. Wnt4 signaling prevents skeletal aging and inflammation by inhibiting nuclear factor-κB. Nat. Med. 2014, 20, 1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappariello, A.; Capulli, M. The Vicious Cycle of Breast Cancer-Induced Bone Metastases, a Complex Biological and Therapeutic Target. Curr. Mol. Biol. Rep. 2018, 4, 123–131. [Google Scholar] [CrossRef]
- Macedo, F.; Ladeira, K.; Pinho, F.; Saraiva, N.; Bonito, N.; Pinto, L.; Gonçalves, F. Bone metastases: An overview. Oncol. Rev. 2017, 11, 321. [Google Scholar] [CrossRef] [Green Version]
- Kolb, A.D.; Shupp, A.B.; Mukhopadhyay, D.; Marini, F.C.; Bussard, K.M. Osteoblasts are “educated” by crosstalk with metastatic breast cancer cells in the bone tumor microenvironment. Breast Cancer Res. 2019, 21, 31. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Zylstra-Diegel, C.R.; Schumacher, C.A.; Baker, J.J.; Carpenter, A.C.; Rao, S.; Yao, W.; Guan, M.; Helms, J.A.; Lane, N.E.; et al. Wntless functions in mature osteoblasts to regulate bone mass. Proc. Natl. Acad. Sci. USA 2012, 109, E2197–E2204. [Google Scholar] [CrossRef] [Green Version]
- Bordonaro, M.; Tewari, S.; Cicco, C.E.; Atamna, W.; Lazarova, D.L. A Switch from Canonical to Noncanonical Wnt Signaling Mediates Drug Resistance in Colon Cancer Cells. PLoS ONE 2011, 6, e27308. [Google Scholar] [CrossRef]
- Huang, Y.; Zhao, K.; Hu, Y.; Zhou, Y.; Luo, X.; Li, X.; Wei, L.; Li, Z.; You, Q.; Guo, Q.; et al. Wogonoside inhibits angiogenesis in breast cancer via sup-pressing Wnt/β-catenin pathway. Mol. Carcinog. 2016, 55, 1598–1612. [Google Scholar] [CrossRef]
- Kaplan, Z.; Zielske, S.P.; Ibrahim, K.G.; Cackowski, F.C. Wnt and β-Catenin Signaling in the Bone Metastasis of Prostate Cancer. Life 2021, 11, 1099. [Google Scholar] [CrossRef] [PubMed]
- Aktary, Z.; Bertrand, J.U.; Larue, L. The WNT-less wonder: WNT-independent β-catenin signaling. Pigment Cell Melanoma Res. 2016, 29, 524–540. [Google Scholar] [CrossRef] [Green Version]
- Yaccoby, S.; Ling, W.; Zhan, F.; Walker, R.; Barlogie, B.; Shaughnessy, J.D. Antibody-based inhibition of DKK1 suppresses tumor-induced bone resorption and multiple myeloma growth in vivo. Blood 2006, 109, 2106–2111. [Google Scholar] [CrossRef]
- Weilbaecher, K.N.; Guise, T.A.; McCauley, L.K. Cancer to bone: A fatal attraction. Nat. Rev. Cancer 2011, 11, 411–425. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Li, K.; Li, B.-Y.; Yokota, H. Wnt signaling: A double-edged sword in protecting bone from cancer. J. Bone Miner. Metab. 2022, 1–6. [Google Scholar] [CrossRef]
- Birk, R.Z.; Abramovitch-Gottlib, L.; Margalit, I.; Aviv, M.; Forti, E.; Geresh, S.; Vago, R. Conversion of adipogenic to osteogenic phenotype using crystalline porous biomatrices of marine origin. Tissue Eng. 2006, 12, 21–31. [Google Scholar] [CrossRef]
- Dang, C.V. c-Myc Target Genes Involved in Cell Growth, Apoptosis, and Metabolism. Mol. Cell. Biol. 1999, 19, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.; Hall, C.L.; Escara-Wilke, J.; Mizokami, A.; Keller, J.M.; Keller, E.T. Prostate Cancer Induces Bone Metastasis through Wnt-Induced Bone Morphogenetic Protein-Dependent and Independent Mechanisms. Cancer Res. 2008, 68, 5785–5794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-C.; Sosnoski, D.M.; Mastro, A.M. Breast cancer metastasis to the bone: Mechanisms of bone loss. Breast Cancer Res. 2010, 12, 215. [Google Scholar] [CrossRef] [Green Version]
- Salamanna, F.; Contartese, D.; Maglio, M.; Fini, M. A systematic review on in vitro 3D bone metastases models: A new horizon to recapitulate the native clinical scenario? Oncotarget 2016, 7, 44803. [Google Scholar] [CrossRef]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signaling in cancer. Oncogene 2017, 36, 1461. [Google Scholar] [CrossRef]
- McClung, M.R. Clinical utility of anti-sclerostin antibodies. Bone 2017, 96, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Anastas, J.N.; Moon, R.T. WNT signalling pathways as therapeutic targets in cancer. Nat. Rev. Cancer 2012, 13, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.L.; Kang, S.; MacDougald, O.; Keller, E.T. Role of wnts in prostate cancer bone metastases. J. Cell. Biochem. 2005, 97, 661–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James-Bhasin, M.; Siegel, P.M.; Nazhat, S.N. A Three-Dimensional Dense Collagen Hydrogel to Model Cancer Cell/Osteoblast Interactions. J. Funct. Biomater. 2018, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Cheng, G.; Hao, M.; Zheng, J.; Zhou, X.; Zhang, J.; Taichman, R.S.; Pienta, K.J.; Wang, J. CXCL12/CXCR4/CXCR7 chemokine axis and cancer pro-gression. Cancer Metastasis Rev. 2010, 29, 709–722. [Google Scholar] [CrossRef] [Green Version]
- Lepourcelet, M.; Chen, Y.-N.P.; France, D.S.; Wang, H.; Crews, P.; Petersen, F.; Bruseo, C.; Wood, A.W.; Shivdasani, R.A. Small-molecule antagonists of the oncogenic Tcf/β-catenin protein complex. Cancer Cell 2004, 5, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Buchholz, M.; Schatz, A.; Wagner, M.; Michl, P.; Linhart, T.; Adler, G.; Gress, T.M.; Ellenrieder, V. Overexpression of c-myc in pancreatic cancer caused by ectopic activation of NFATc1 and the Ca2 /calcineurin signaling pathway. EMBO J. 2006, 25, 3714–3724. [Google Scholar] [CrossRef] [Green Version]
- Hall, C.L.; Bafico, A.; Dai, J.; Aaronson, S.A.; Keller, E.T. Prostate Cancer Cells Promote Osteoblastic Bone Metastases through Wnts. Cancer Res. 2005, 65, 7554–7560. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez Díaz, E.C.; Sinha, S.; Avedian, R.S.; Yang, F. Tissue-engineered 3D models for elucidating primary and metastatic bone cancer progression. Acta Biomater. 2019, 99, 18–32. [Google Scholar] [CrossRef]
- Abramovitch-Gottlib, L.; Geresh, S.; Vago, R. Biofabricated Marine Hydrozoan: A Bioactive Crystalline Material Promoting Ossification of Mesenchymal Stem Cells. Tissue Eng. 2006, 12, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Asulin, M.; Ghedalia-Peled, N.B.; Erez, I.C.; Ventura, Y.; Vago, R. Transition between canonical to non-canonical Wnt sig-naling during interactions between mesenchymal stem cells and osteosarcomas. Open J. Orthop. Rheumatol. 2020, 5, 037–048. [Google Scholar]
- A Guise, T. The vicious cycle of bone metastases. J. Musculoskelet. Neuronal Interact. 2002, 2, 570–572. [Google Scholar] [PubMed]
- Malladi, S.; Macalinao, D.G.; Jin, X.; He, L.; Basnet, H.; Zou, Y.; de Stanchina, E.; Massagué, J. Metastatic latency and immune evasion through au-tocrine inhibition of WNT. Cell 2016, 165, 45–60. [Google Scholar] [CrossRef] [Green Version]
- Orian-Rousseau, V. CD44 Acts as a Signaling Platform Controlling Tumor Progression and Metastasis. Front. Immunol. 2015, 6, 154. [Google Scholar] [CrossRef] [Green Version]
- Clines, K.L.; Clines, G.A. DKK1 and Kremen Expression Predicts the Osteoblastic Response to Bone Metastasis. Transl. Oncol. 2018, 11, 873–882. [Google Scholar] [CrossRef]
- Adjei, I.M.; Temples, M.N.; Brown, S.B.; Sharma, B. Targeted Nanomedicine to Treat Bone Metastasis. Pharmaceutics 2018, 10, 205. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Siegal, G.P. Metastatic Organotropism: An Intrinsic Property of Breast Cancer Molecular Subtypes. Adv. Anat. Pathol. 2017, 24, 78–81. [Google Scholar] [CrossRef]
- Lee, G.T.; I Kang, D.; Ha, Y.-S.; Jung, Y.S.; Chung, J.; Min, K.; Kim, T.H.; Moon, K.H.; Lee, D.H.; Kim, W.-J.; et al. Prostate cancer bone metastases acquire resistance to androgen deprivation via WNT5A-mediated BMP-6 induction. Br. J. Cancer 2014, 110, 1634–1644. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.C.; Chan, L.S.; Ip, J.C.Y.; Lo, C.; Yip, T.T.C.; Ngan, R.K.C.; Wong, R.N.S.; Lo, K.W.; Ng, W.T.; Lee, A.W.M.; et al. Therapeutic targeting of CBP/β-catenin signaling reduces cancer stem-like population and synergistically suppresses growth of EBV-positive nasopharyngeal carcinoma cells with cisplatin. Sci. Rep. 2015, 5, 9979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, B.-J.; Wang, Y.-Q.; Wei, X.-J.; Rong, L.-Q.; Wei, D.; Yan, C.-M.; Wang, D.-J.; Sun, J.-Y. Expression of WNT-5a and ROR2 correlates with disease severity in osteosarcoma. Mol. Med. Rep. 2012, 5, 1033–1036. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben-Ghedalia-Peled, N.; Vago, R. Wnt Signaling in the Development of Bone Metastasis. Cells 2022, 11, 3934. https://doi.org/10.3390/cells11233934
Ben-Ghedalia-Peled N, Vago R. Wnt Signaling in the Development of Bone Metastasis. Cells. 2022; 11(23):3934. https://doi.org/10.3390/cells11233934
Chicago/Turabian StyleBen-Ghedalia-Peled, Noa, and Razi Vago. 2022. "Wnt Signaling in the Development of Bone Metastasis" Cells 11, no. 23: 3934. https://doi.org/10.3390/cells11233934
APA StyleBen-Ghedalia-Peled, N., & Vago, R. (2022). Wnt Signaling in the Development of Bone Metastasis. Cells, 11(23), 3934. https://doi.org/10.3390/cells11233934