

Effect of Mechanical Loading of Senescent Myoblasts on Their Myogenic Lineage Progression and Survival
 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. C2C12 Cell Culture
2.2. C2C12 Myoblast Aging
2.3. C2C12 Cells Mechanical Loading
2.4. Cell lysis, RNA Extraction, Reverse Transcription, and Real-Time PCR
2.5. Protein Extraction and Immunoblotting Analysis
2.6. Statistics
3. Results
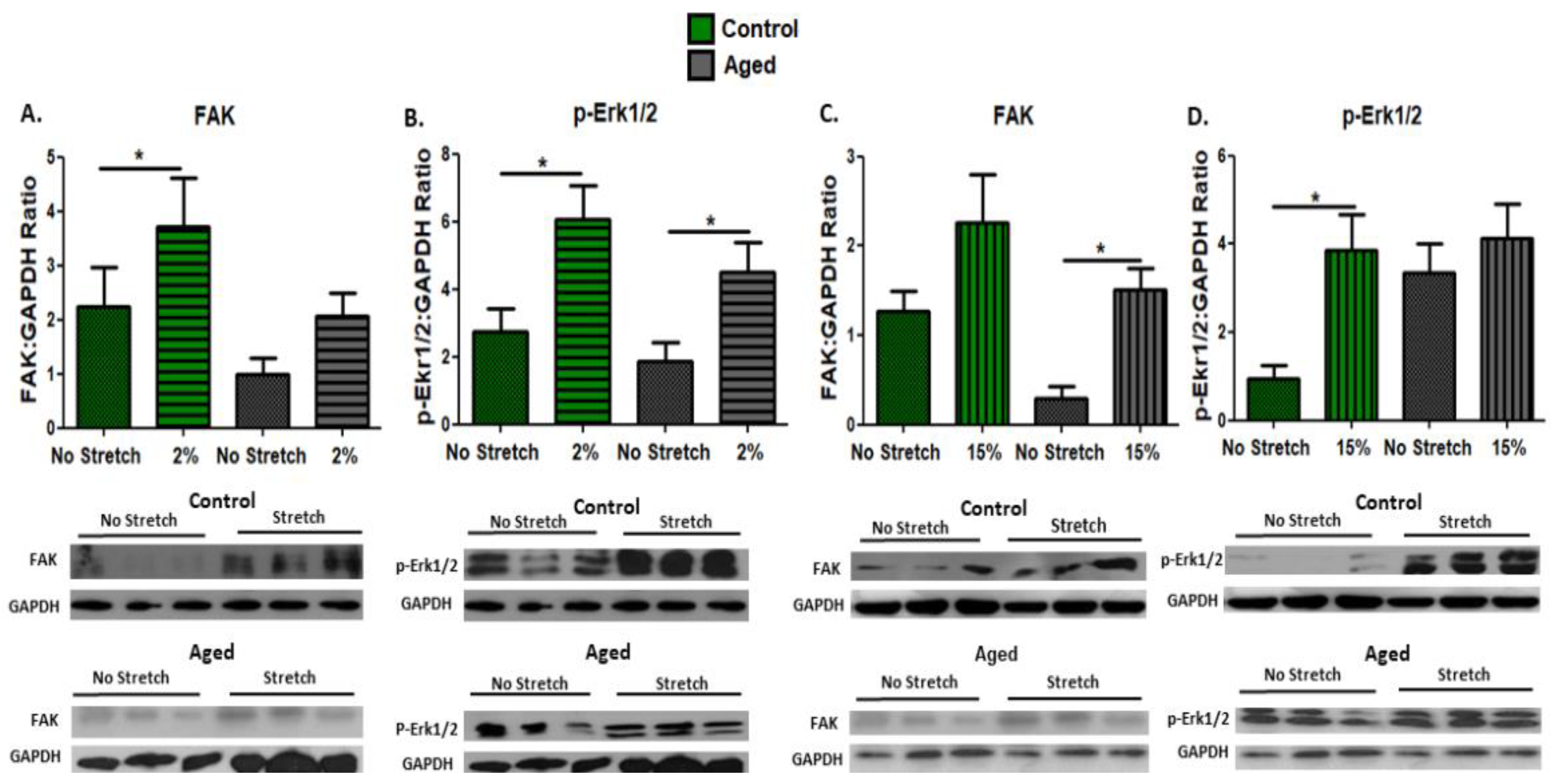
3.1. Effect of Mechanical Loading on Mechanosensitive and Signaling Proteins in C2C12 Myoblasts
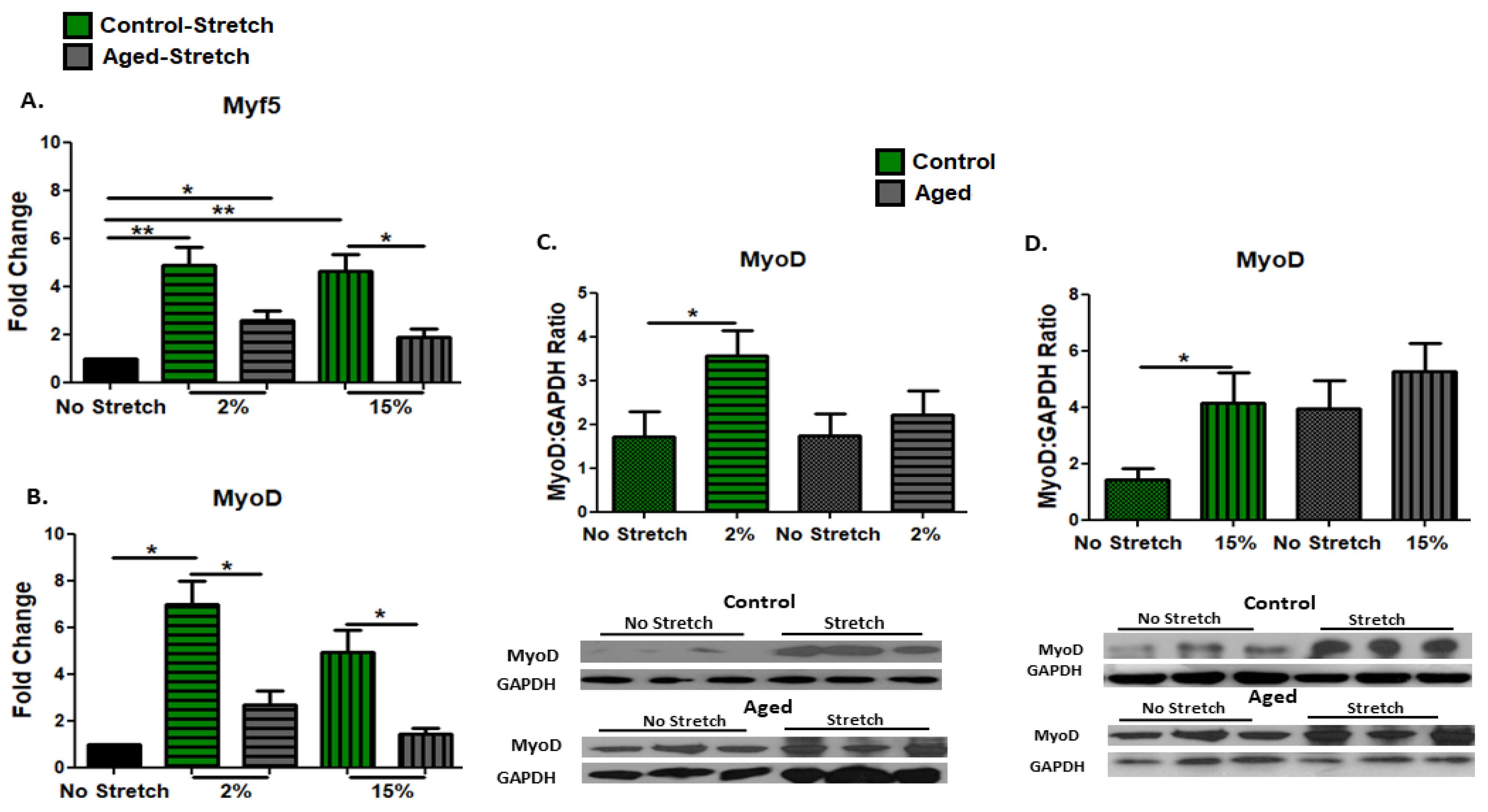
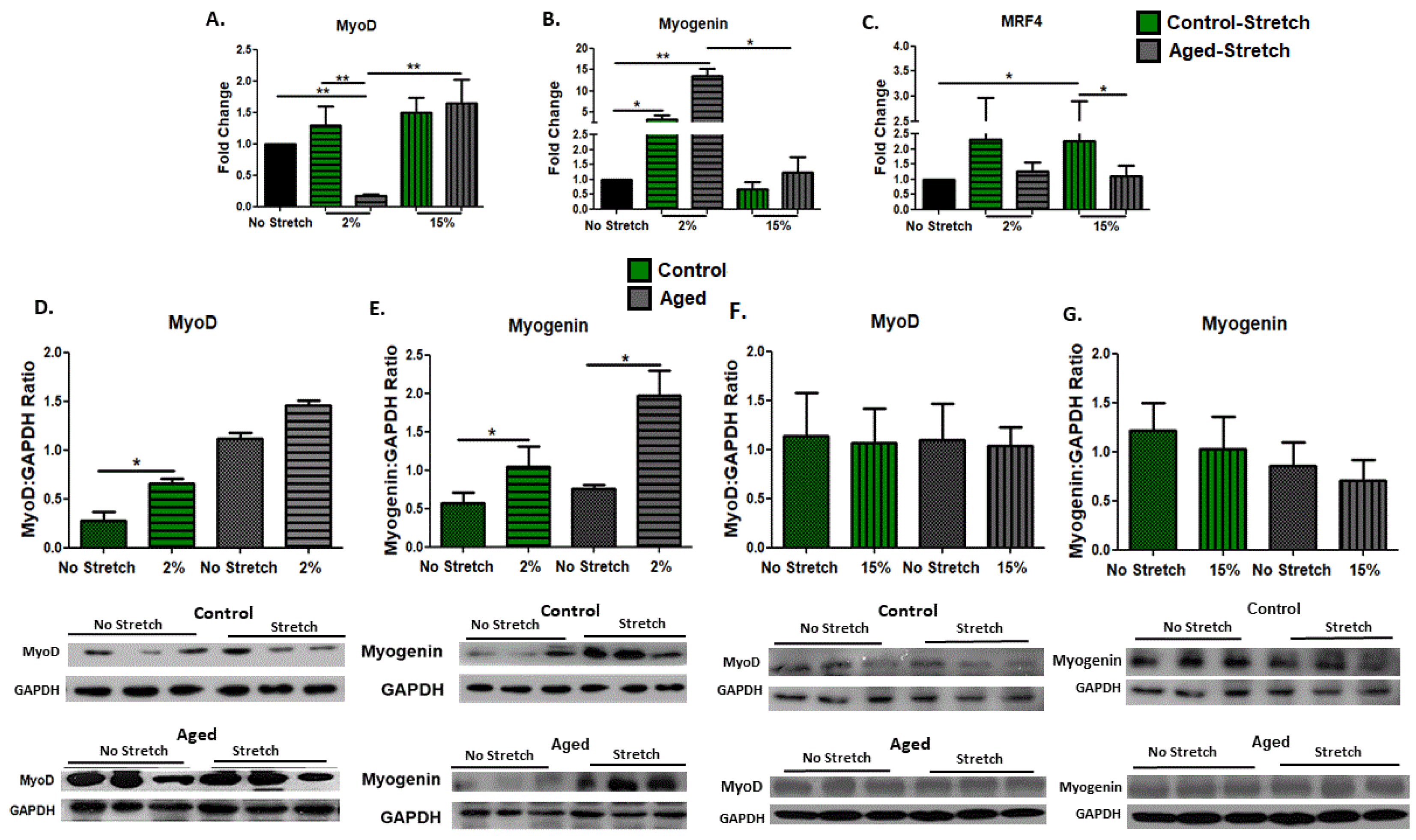
3.2. Effect of Mechanical Loading on MRFs in C2C12 Myoblasts
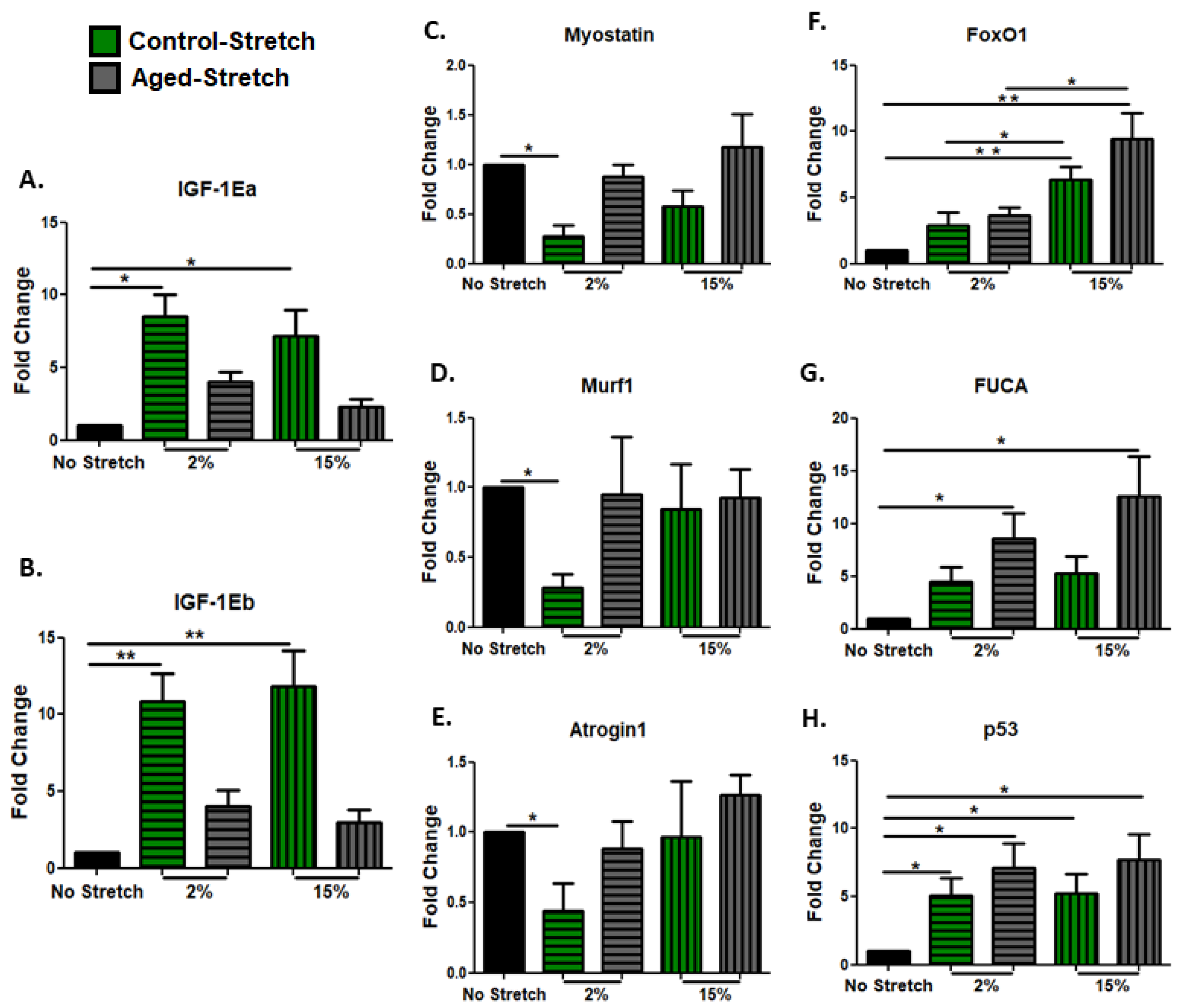
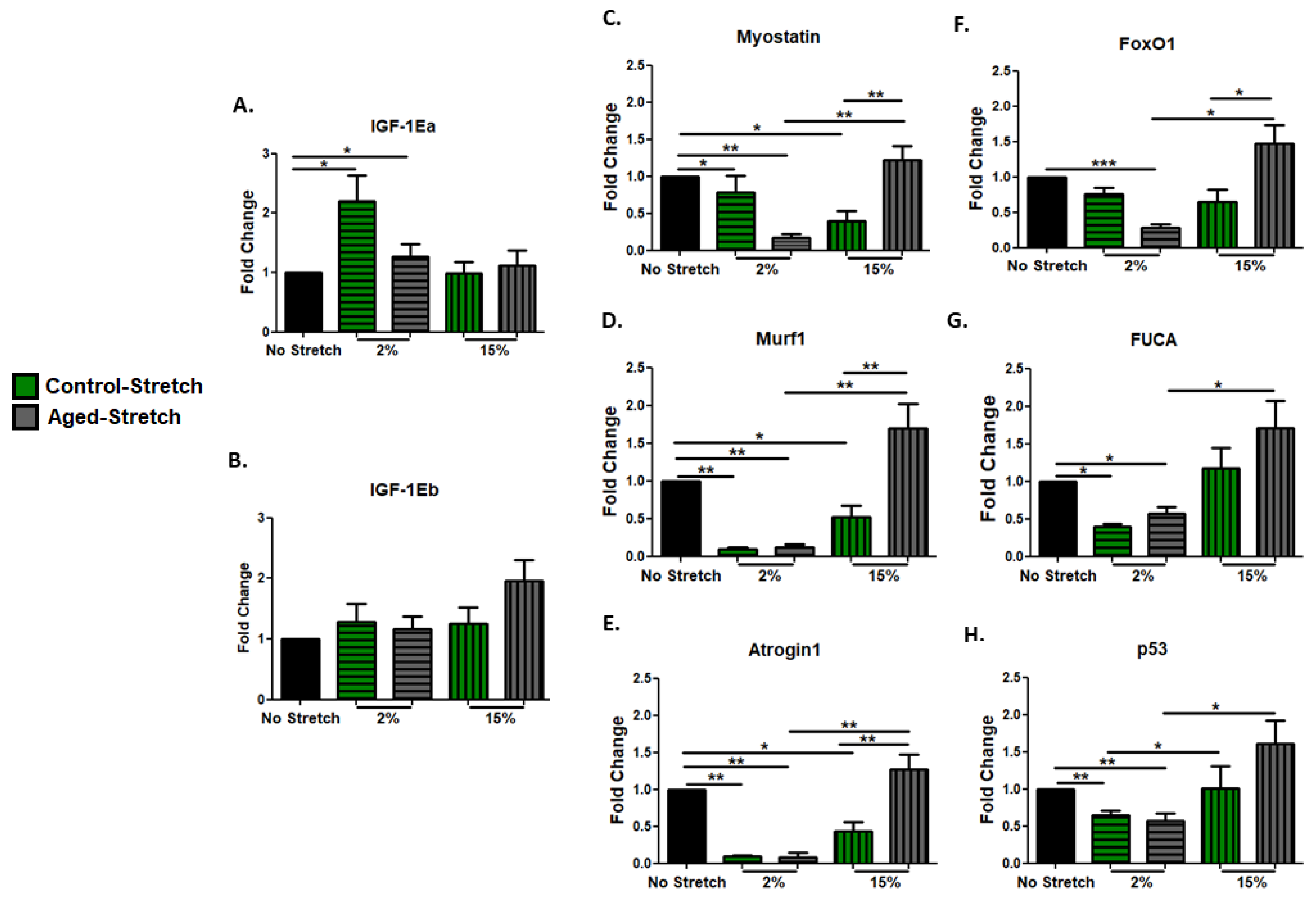
3.3. Effect of Mechanical Loading Protocols on Growth, Atrophy, and Pro-Apoptotic Factors in C2C12 Cells Myoblasts
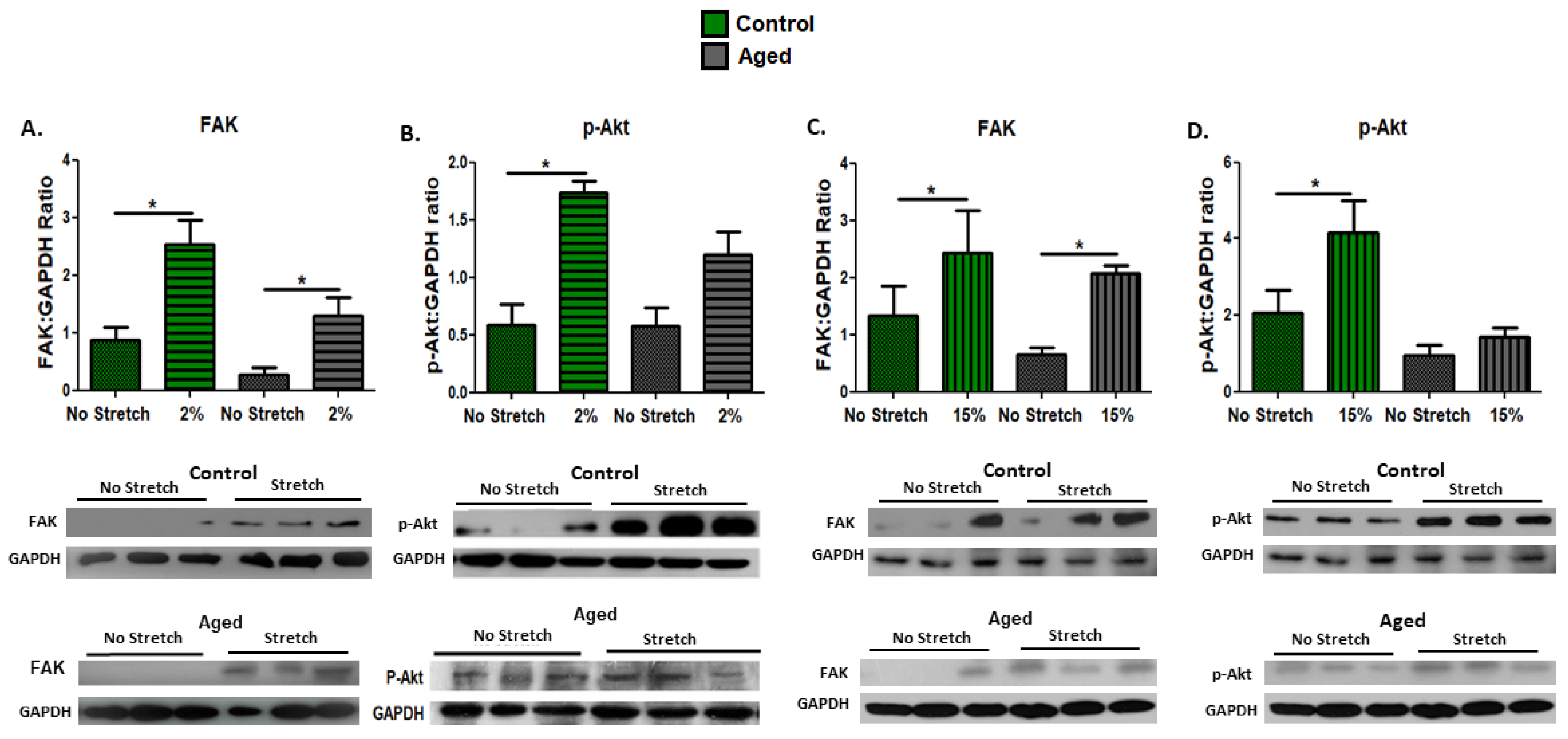
3.4. Effect of Mechanical Loading Protocols on Mechanosensitive and Signaling Proteins in C2C12 Myotubes
3.5. Effect of Mechanical Loading on Myogenic Regulatory Factors in C2C12 Myotubes
3.6. Effect of Mechanical Loading οn Growth, Muscle Atrophy, and Pro-Apoptotic Factors in C2C12 Myotubes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Combaret, L.; Dardevet, D.; Bechet, D.; Taillandier, D.; Mosoni, L.; Attaix, D. Skeletal muscle proteolysis in aging. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 37–41. [Google Scholar] [CrossRef]
- Brack, A.S.; Muñoz-Canoves, P. The ins and outs of muscle stem cell aging. Skelet. Muscle 2016, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Dijk, M.; Nagel, J.; Dijk, F.J.; Salles, J.; Verlaan, S.; Walrand, S.; van Norren, K.; Luiking, Y. Sarcopenia in older mice is characterized by a decreased anabolic response to a protein meal. Arch. Gerontol. Geriatr. 2017, 69, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Moustogiannis, A.; Philippou, A.; Taso, O.; Zevolis, E.; Pappa, M.; Chatzigeorgiou, A.; Koutsilieris, M. The Effects of Muscle Cell Aging on Myogenesis. Int. J. Mol. Sci. 2021, 22, 3721. [Google Scholar] [CrossRef] [PubMed]
- Hayflick, L.; Moorhead, P.S. The serial cultivation of human diploid cell strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef]
- Doherty, T.J. Invited Review: Aging and sarcopenia. J. Appl. Physiol. (1985) 2003, 95, 1717–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boers, H.E.; Haroon, M.; le Grand, F.; Bakker, A.D.; Klein-Nulend, J.; Jaspers, R.T. Mechanosensitivity of aged muscle stem cells. J. Orthop. Res. 2018, 36, 632–641. [Google Scholar]
- Chen, H.; Li, Y.; Tollefsbol, T.O. Cell Senescence Culturing Methods. Methods Mol. Biol. 2013, 1048, 1–10. [Google Scholar]
- Beck, J.; Horikawa, I.; Harris, C. Cellular Senescence: Mechanisms, Morphology, and Mouse Models. Vet. Pathol. 2020, 57, 747–757. [Google Scholar] [CrossRef]
- Asfour, H.A.; Allouh, M.Z.; Said, R.S. Myogenic regulatory factors: The orchestrators of myogenesis after 30 years of discovery. Exp. Biol. Med. 2018, 243, 118–128. [Google Scholar] [CrossRef]
- Karalaki, M.; Fili, S.; Philippou, A.; Koutsilieris, M. Muscle regeneration: Cellular and molecular events. In Vivo 2009, 23. [Google Scholar]
- Hernández-Hernández, J.M.; García-González, E.G.; Brun, C.E.; Rudnicki, M.A. The myogenic regulatory factors, determinants of muscle development, cell identity and regeneration. Semin. Cell Dev. Biol. 2017, 72, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Sharples, A.P.; Stewart, C. Myoblast models of skeletal muscle hypertrophy and atrophy. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 230–236. [Google Scholar] [CrossRef]
- Philippou, A.; Barton, E.R. Optimizing IGF-I for skeletal muscle therapeutics. Growth Horm. IGF Res. 2014, 24, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Philippou, A.; Halapas, A.; Maridaki, M.; Koutsilieris, M. Type I insulin-like growth factor receptor signaling in skeletal muscle regeneration and hypertrophy. J. Musculoskelet. Neuronal Interact. 2007, 7, 208–218. [Google Scholar]
- Philippou, A.; Maridaki, M.; Halapas, A.; Koutsilieris, M. The role of the insulin-like growth factor 1 (IGF-1) in skeletal muscle physiology. In Vivo 2007, 21, 45–54. [Google Scholar]
- Barton, E.R.; Park, S.; James, J.K.; Makarewich, C.A.; Philippou, A.; Eletto, D.; Lei, H.; Brisson, B.; Ostrovsky, O.; Li, Z.; et al. Deletion of muscle GRP94 impairs both muscle and body growth by inhibiting local IGF production. FASEB J. 2012, 26, 3691–3702. [Google Scholar] [CrossRef] [Green Version]
- Durzyńska, J.; Philippou, A.; Brisson, B.K.; Nguyen-McCarty, M.; Barton, E.R. The pro-Forms of Insulin-Like Growth Factor I (IGF-I) Are Predominant in Skeletal Muscle and Alter IGF-I Receptor Activation. Endocrinology 2013, 154, 1215–1224. [Google Scholar] [CrossRef] [Green Version]
- Papageorgiou, E.; Philippou, A.; Armakolas, A.; Christopoulos, P.F.; Dimakakos, A.; Koutsilieris, M. The human Ec peptide: The active core of a progression growth factor with species-specific mode of action. Hormones 2016, 15, 423–434. [Google Scholar] [CrossRef]
- Philippou, A.; Papageorgiou, E.; Bogdanis, G.; Halapas, A.; Sourla, A.; Maridaki, M.; Pissimissis, N.; Koutsilieris, M. Expression of IGF-1 isoforms after exercise-induced muscle damage in humans: Characterization of the MGF E peptide actions in vitro. In Vivo 2009, 23, 567–575. [Google Scholar] [PubMed]
- Vassilakos, G.; Philippou, A.; Koutsilieris, M. Identification of the IGF-1 processing product human Ec/rodent Eb peptide in various tissues: Evidence for its differential regulation after exercise-induced muscle damage in humans. Growth Horm. IGF Res. 2017, 32, 22–28. [Google Scholar] [CrossRef]
- Lin, Y.-A.; Li, Y.-R.; Chang, Y.-C.; Hsu, M.-C.; Chen, S.-T. Activation of IGF-1 pathway and suppression of atrophy related genes are involved in Epimedium extract (icariin) promoted C2C12 myotube hypertrophy. Sci. Rep. 2021, 11, 1–16. [Google Scholar]
- Sacheck, J.M.; Ohtsuka, A.; McLary, S.C.; Goldberg, A.L. IGF-I stimulates muscle growth by suppressing protein breakdown and expression of atrophy-related ubiquitin ligases, atrogin-1 and MuRF1. Am. J. Physiol. Metab. 2004, 287, E591–E601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stitt, T.N.; Drujan, D.; Clarke, B.A.; Panaro, F.; Timofeyva, Y.; Kline, W.O.; Gonzalez, M.; Yancopoulos, G.D.; Glass, D.J. The IGF-1/PI3K/Akt pathway prevents expression of muscle atrophy-induced ubiquitin ligases by inhibiting FOXO transcription factors. Mol. Cell 2004, 14, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Bian, A.; Ma, Y.; Zhou, X.; Guo, Y.; Wang, W.; Zhang, Y.; Wang, X. Association between sarcopenia and levels of growth hormone and insulin-like growth factor-1 in the elderly. BMC Musculoskelet. Disord. 2020, 21, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mancinelli, R.; Checcaglini, F.; Coscia, F.; Gigliotti, P.; Fulle, S.; Fanò-Illic, G. Biological Aspects of Selected Myokines in Skeletal Muscle: Focus on Aging. Int. J. Mol. Sci. 2021, 22, 8520. [Google Scholar] [CrossRef]
- Philippou, A.; Minozzo, F.C.; Spinazzola, J.M.; Smith, L.R.; Lei, H.; Rassier, D.E.; Barton, E.R. Masticatory muscles of mouse do not undergo atrophy in space. FASEB J. 2015, 29, 2769–2779. [Google Scholar] [CrossRef] [Green Version]
- Dirks, A.J.; Leeuwenburgh, C. The Role of Apoptosis in Age-Related Skeletal Muscle Atrophy. Sports Med. 2005, 35, 473–483. [Google Scholar] [CrossRef]
- Baczek, J.; Silkiewicz, M.; Wojszel, Z.B. Myostatin as a Biomarker of Muscle Wasting and other Pathologies-State of the Art and Knowledge Gaps. Nutrients 2020, 12, 2401. [Google Scholar] [CrossRef]
- Kawada, S.; Tachi, C.; Ishii, N. Content and localization of myostatin in mouse skeletal muscles during aging, mechanical unloading and reloading. J. Muscle Res. Cell Motil. 2001, 22, 627–633. [Google Scholar] [CrossRef]
- Trendelenburg, A.U.; Meyer, A.; Rohner, D.; Boyle, J.; Hatakeyama, S.; Glass, D.J. Myostatin reduces Akt/TORC1/p70S6K signaling, inhibiting myoblast differentiation and myotube size. Am. J. Physiol. Physiol. 2009, 296, C1258–C1270. [Google Scholar] [CrossRef] [Green Version]
- Morissette, M.R.; Cook, S.A.; Buranasombati, C.; Rosenberg, M.A.; Rosenzweig, A. Myostatin inhibits IGF-I-induced myotube hypertrophy through Akt. Am. J. Physiol. Physiol. 2009, 297, 1124–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumucio, J.P.; Mendias, C.L. Atrogin-1, MuRF-1, and sarcopenia. Endocrine 2013, 43, 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodine, S.C.; Latres, E.; Baumhueter, S.; Lai, V.K.-M.; Nunez, L.; Clarke, B.A.; Poueymirou, W.T.; Panaro, F.J.; Na, E.; Dharmarajan, K.; et al. Identification of Ubiquitin Ligases Required for Skeletal Muscle Atrophy. Science 2001, 294, 1704–1708. [Google Scholar] [CrossRef]
- Kitamura, T.; Kitamura, Y.I.; Funahashi, Y.; Shawber, C.J.; Castrillon, D.H.; Kollipara, R.; DePinho, R.; Kitajewski, J.; Accili, D. A Foxo/Notch pathway controls myogenic differentiation and fiber type specification. J. Clin. Investig. 2007, 117, 2477–2485. [Google Scholar] [CrossRef] [Green Version]
- Bourgeois, B.; Madl, T. Regulation of cellular senescence via the FOXO4-p53 axis. FEBS Lett. 2018, 592, 2083–2097. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, L.M. Atrophy and programmed cell death of skeletal muscle. Cell Death Differ. 2008, 15, 1163–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron-Smith, D. Exercise And Skeletal Muscle Gene Expression. Clin. Exp. Pharmacol. Physiol. 2002, 29, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Egan, B.; Zierath, J.R. Exercise Metabolism and the Molecular Regulation of Skeletal Muscle Adaptation. Cell Metab. 2013, 17, 162–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkholder, T.J. Mechanotransduction in skeletal muscle. Front. Biosci. 2007, 12, 174–191. [Google Scholar] [CrossRef] [Green Version]
- Passey, S.; Martin, N.; Player, D.; Lewis, M.P. Stretching skeletal muscle in vitro: Does it replicate in vivo physiology? Biotechnol. Lett. 2011, 33, 1513–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.-H.; Ushida, T.; Akimoto, T. Control of cell differentiation by mechanical stress. J. Phys. Fit. Sports Med. 2013, 2, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Zevolis, E.; Philippou, A.; Moustogiannis, A.; Chatzigeorgiou, A.; Koutsilieris, M. Optimizing mechanical stretching protocols for hypertrophic and anti-apoptotic responses in cardiomyocyte-like H9C2 cells. Mol. Biol. Rep. 2021, 48, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Moustogiannis, A.; Philippou, A.; Zevolis, E.; Taso, O.; Chatzigeorgiou, A.; Koutsilieris, M. Characterization of Optimal Strain, Frequency and Duration of Mechanical Loading on Skeletal Myotubes' Biological Responses. In Vivo 2020, 34, 1779–1788. [Google Scholar] [CrossRef] [PubMed]
- Zevolis, E.; Philippou, A.; Moustogiannis, A.; Chatzigeorgiou, A.; Koutsilieris, M. The Effects of Mechanical Loading Variations on the Hypertrophic, Anti-Apoptotic, and Anti-Inflammatory Responses of Differentiated Cardiomyocyte-like H9C2 Cells. Cells 2022, 11, 473. [Google Scholar] [CrossRef] [PubMed]
- Ingber, D.E. Cellular mechanotransduction: Putting all the pieces together again. FASEB J. 2006, 20, 811–827. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, P.G.; Walker, R.G. Molecular basis of mechanosensory transduction. Nature 2001, 413, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Vining, K.H.; Mooney, D.J. Mechanical forces direct stem cell behaviour in development and regeneration. Nat. Rev. Mol. Cell Biol. 2017, 18, 728–742. [Google Scholar] [CrossRef]
- A Schwartz, M.; DeSimone, D.W. Cell adhesion receptors in mechanotransduction. Curr. Opin. Cell Biol. 2008, 20, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Zebda, N.; Dubrovskyi, O.; Birukov, K.G. Focal Adhesion Kinase Regulation of Mechanotransduction and its Impact on Endothelial Cell Functions. Microvasc. Res. 2012, 83, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Lassiter, D.G.; Nylén, C.; Sjögren, R.J.O.; Chibalin, A.V.; Wallberg-Henriksson, H.; Näslund, E.; Krook, A.; Zierath, J.R. FAK tyrosine phosphorylation is regulated by AMPK and controls metabolism in human skeletal muscle. Diabetologia 2017, 61, 424–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisht, B.; Goel, H.L.; Dey, C.S. Focal adhesion kinase regulates insulin resistance in skeletal muscle. Diabetologia 2007, 50, 1058–1069. [Google Scholar] [CrossRef] [Green Version]
- Graham, Z.A.; Gallagher, P.M.; Cardozo, C.P. Focal adhesion kinase and its role in skeletal muscle. J. Muscle Res. Cell Motil. 2015, 36, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Graham, Z.A.; Qin, W.; Harlow, L.C.; Ross, N.H.; A Bauman, W.; Gallagher, P.M.; Cardozo, C.P. Focal adhesion kinase signaling is decreased 56 days following spinal cord injury in rat gastrocnemius. Spinal Cord 2015, 54, 502–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crossland, H.; Kazi, A.A.; Lang, C.H.; A Timmons, J.; Pierre, P.; Wilkinson, D.J.; Smith, K.; Szewczyk, N.; Atherton, P.J. Focal adhesion kinase is required for IGF-I-mediated growth of skeletal muscle cells via a TSC2/mTOR/S6K1-associated pathway. Am. J. Physiol. Metab. 2013, 305, E183–E193. [Google Scholar] [CrossRef] [PubMed]
- Rozo, M.; Li, L.; Fan, M.R.L.L.C.-M. Targeting β1-integrin signaling enhances regeneration in aged and dystrophic muscle in mice. Nat. Med. 2016, 22, 889–896. [Google Scholar] [CrossRef]
- Moorwood, C.; Philippou, A.; Spinazzola, J.; Keyser, B.; Macarak, E.J.; Barton, E.R. Absence of γ-sarcoglycan alters the response of p70S6 kinase to mechanical perturbation in murine skeletal muscle. Skelet. Muscle 2014, 4, 13. [Google Scholar] [CrossRef] [Green Version]
- Stavropoulou, A.; Halapas, A.; Sourla, A.; Philippou, A.; Papageorgiou, E.; Papalois, A.; Koutsilieris, M. IGF-1 Expression in Infarcted Myocardium and MGF E Peptide Actions in Rat Cardiomyocytes in Vitro. Mol. Med. 2009, 15, 127–135. [Google Scholar] [CrossRef]
- Philippou, A.; Armakolas, A.; Koutsilieris, M. Evidence for the Possible Biological Significance of the igf-1 Gene Alternative Splicing in Prostate Cancer. Front. Endocrinol. 2013, 4, 31. [Google Scholar] [CrossRef] [Green Version]
- Soltow, Q.A.; Zeanah, E.H.; Lira, V.A.; Criswell, D.S. Cessation of cyclic stretch induces atrophy of C2C12 myotubes. Biochem. Biophys. Res. Commun. 2013, 434, 316–321. [Google Scholar] [CrossRef]
- Atherton, P.J.; Szewczyk, N.; Selby, A.; Rankin, D.; Hillier, K.; Smith, K.; Rennie, M.J.; Loughna, P. Cyclic stretch reduces myofibrillar protein synthesis despite increases in FAK and anabolic signalling in L6 cells. J. Physiol. 2009, 587, 3719–3727. [Google Scholar] [CrossRef] [PubMed]
- Sulzmaier, F.J.; Jean, C.; Schlaepfer, D.D. FAK in cancer: Mechanistic findings and clinical applications. Nat. Rev. Cancer 2014, 14, 598–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Chen, Q.; Xing, D. Focal adhesion kinase activates NF-κB via the ERK1/2 and p38MAPK Pathways in amyloid-β25-35-induced apoptosis in PC12 cells. J. Alzheimers Dis. 2012, 32, 77–94. [Google Scholar] [CrossRef] [Green Version]
- Knight, J.; Kothary, R. The myogenic kinome: Protein kinases critical to mammalian skeletal myogenesis. Skelet. Muscle 2011, 1, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Woodring, P.J.; Bhakta, K.S.; Tamura, K.; Wen, F.; Feramisco, J.R.; Karin, M.; Wang, J.Y.J.; Puri, P.L. p38 and Extracellular Signal-Regulated Kinases Regulate the Myogenic Program at Multiple Steps. Mol. Cell. Biol. 2000, 20, 3951–3964. [Google Scholar] [CrossRef] [Green Version]
- Ostrovsky, O.; Bengal, E. The Mitogen-activated Protein Kinase Cascade Promotes Myoblast Cell Survival by Stabilizing the Cyclin-dependent Kinase Inhibitor, p21WAF1 Protein. J. Biol. Chem. 2003, 278, 21221–21231. [Google Scholar] [CrossRef]
- Sonoda, Y.; Watanabe, S.; Matsumoto, Y.; Aizu-Yokota, E.; Kasahara, T. FAK Is the Upstream Signal Protein of the Phosphatidylinositol 3-Kinase-Akt Survival Pathway in Hydrogen Peroxide-induced Apoptosis of a Human Glioblastoma Cell Line. J. Biol. Chem. 1999, 274, 10566–10570. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Luo, Q.; Chen, Z.; Shi, Y.; Ju, Y.; Yang, L.; Song, G. Increased nuclear stiffness via FAK-ERK1/2 signaling is necessary for synthetic mechano-growth factor E peptide-induced tenocyte migration. Sci. Rep. 2016, 6, 18809. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Peng, C.; Wei, X.; Luo, D.; Lin, Y.; Yang, T.; Jin, X.; Gong, L.; Li, H.; Wang, K. Simulated physiological stretch increases expression of extracellular matrix proteins in human bladder smooth muscle cells via integrin α4/αv-FAK-ERK1/2 signaling pathway. World J. Urol. 2017, 35, 1247–1254. [Google Scholar] [CrossRef]
- Klossner, S.; Durieux, A.-C.; Freyssenet, D.; Flueck, M. Mechano-transduction to muscle protein synthesis is modulated by FAK. Eur. J. Appl. Physiol. 2009, 106, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Chargé, S.B.P.; Rudnicki, M. Cellular and Molecular Regulation of Muscle Regeneration. Physiol. Rev. 2004, 84, 209–238. [Google Scholar] [CrossRef] [PubMed]
- Glass, D.J. Signalling pathways that mediate skeletal muscle hypertrophy and atrophy. Nature 2003, 5, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Fannin, J.; Rice, K.M.; Wang, B.; Blough, E.R. Effect of aging on cellular mechanotransduction. Ageing Res. Rev. 2011, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Katta, A.; Gadde, M.K.; Liu, H.; Kakarla, S.K.; Fannin, J.; Paturi, S.; Arvapalli, R.K.; Rice, K.M.; Wang, Y.; et al. Aging-Associated Dysfunction of Akt/Protein Kinase B: S-Nitrosylation and Acetaminophen Intervention. PLoS ONE 2009, 4, e6430. [Google Scholar] [CrossRef]
- Ascenzi, F.; Barberi, L.; Dobrowolny, G.; Villa Nova Bacurau, A.; Nicoletti, C.; Rizzuto, E.; Rosenthal, N.; Scicchitano, B.M.; Musarò, A. Effects of IGF-1 isoforms on muscle growth and sarcopenia. Aging Cell 2019, 18, e12954. [Google Scholar] [CrossRef] [PubMed]
- Langley, B.; Thomas, M.; Bishop, A.; Sharma, M.; Gilmour, S.; Kambadur, R. Myostatin Inhibits Myoblast Differentiation by Down-regulating MyoD Expression. J. Biol. Chem. 2002, 277, 49831–49840. [Google Scholar] [CrossRef]
- Jogo, M.; Shiraishi, S.; Tamura, T.-A. Identification of MAFbx as a myogenin-engaged F-box protein in SCF ubiquitin ligase. FEBS Lett. 2009, 583, 2715–2719. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | 5′-3′ forward Primer Sequence | 5′-3′ Reverse Primer Sequence | Product Length |
|---|---|---|---|
| GAPDH | CAA CTC CCT CAA GAT TGT CAG CAA | GGC ATG GAC TGT GGT CAT GA | 118 |
| Myf5 | CTA TTA CAG CCT GCC GGG AC | CTC GGA TGG CTC TGT AGA CG | 232 |
| MyoD | TGC TCC TTT GAG ACA GCA GA | AGT AGG GAA GTG TGC GTG CT | 141 |
| Myogenin | AGG AGA GAA AGA TGG AGT CCA GAG | TAA CAA AAG AAG TCA CCC CAA GAG | 430 |
| MRF4 | AGG GCT CTC CTT TGT ATC CAG | TGG AAG AAA GGC GCT GAA GA | 579 |
| IGF-1Ea | GTG GAC GCT CTT CAG TTC GT | GCT TCC TTT TCT TGT GTG TCG ATA G | 262 |
| IGF-1Eb | GTC CCC AGC ACA CAT CGC G | TCT TTT GTG CAA AAT AAG GCG TA | 259 |
| FUCA | TTT GGT CGG TGA GTT GGG AG | CCA TTC CAA GAG CGA GTG GT | 76 |
| FoxO1 | AGT GGA TGG TGA AGA GCG TG | GAA GGG ACA GAT TGT GGC GA | 96 |
| p53 | GAG AGA CCG CCG TAC AGA AG | AGC AGT TTG GGC TTT CCT CC | 317 |
| Myostatin | CTG TAA CCT TCC CAG GAC CA | GCA GTC AAG CCC AAA GTC TC | 104 |
| MuRF1 | AGG GCT CCC CAC CAC CTG TGT | TGC CCT CTC TAG GCC ACC G | 310 |
| Atrogin1 (MAFbx) | AAC AAG GAG GTA TAC AGT AAG G | AAT TGT TCA TGA AGT TCT TTT G | 322 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moustogiannis, A.; Philippou, A.; Zevolis, E.; Taso, O.S.; Giannopoulos, A.; Chatzigeorgiou, A.; Koutsilieris, M. Effect of Mechanical Loading of Senescent Myoblasts on Their Myogenic Lineage Progression and Survival. Cells 2022, 11, 3979. https://doi.org/10.3390/cells11243979
Moustogiannis A, Philippou A, Zevolis E, Taso OS, Giannopoulos A, Chatzigeorgiou A, Koutsilieris M. Effect of Mechanical Loading of Senescent Myoblasts on Their Myogenic Lineage Progression and Survival. Cells. 2022; 11(24):3979. https://doi.org/10.3390/cells11243979
Chicago/Turabian StyleMoustogiannis, Athanasios, Anastassios Philippou, Evangelos Zevolis, Orjona S. Taso, Antonios Giannopoulos, Antonios Chatzigeorgiou, and Michael Koutsilieris. 2022. "Effect of Mechanical Loading of Senescent Myoblasts on Their Myogenic Lineage Progression and Survival" Cells 11, no. 24: 3979. https://doi.org/10.3390/cells11243979
APA StyleMoustogiannis, A., Philippou, A., Zevolis, E., Taso, O. S., Giannopoulos, A., Chatzigeorgiou, A., & Koutsilieris, M. (2022). Effect of Mechanical Loading of Senescent Myoblasts on Their Myogenic Lineage Progression and Survival. Cells, 11(24), 3979. https://doi.org/10.3390/cells11243979