
Cannabinoid Compounds as a Pharmacotherapeutic Option for the Treatment of Non-Cancer Skin Diseases


Abstract
:1. Introduction
2. The Endocannabinoid System: A Brief Overview
2.1. Classic Cannabinoid Receptors
2.2. Endocannabinoids
2.3. Enzymes Involved in Biosynthesis and Degradation of Endocannabinoids
2.4. Further Receptor Targets of Cannabinoids
3. The Endocannabinoid System in the Skin
3.1. Distribution of Components of the Endocannabinoid System in the Skin
3.1.1. Classic Endocannabinoid System
3.1.2. Extended Endocannabinoid System
3.2. The Endocannabinoid System as a Regulator of Skin Homeostasis
3.2.1. Influence on Melanogenesis
3.2.2. Influence on Wound Healing
Cannabinoid Receptor Knockout Models
Role of CB2 Receptor Activation—Results from In Vivo Experiments
Effects of ACEA and Adelmidrol
Role of FAAH—Results from In Vivo Experiments
Role of the Endocannabinoid System in Cell Migration—Results from In Vitro Experiments
On the Path to Clinical Use
3.2.3. Influence on Cutaneous Barrier Function
3.2.4. Influence on Sebocyte Biology
3.2.5. Influence on Hair Follicle Biology
3.2.6. Influence on the Photoexposed Epithelium
3.2.7. Influence on Cutaneous Pain
3.2.8. Influence on Keratinisation
3.2.9. Influence on Skin Ageing Processes
3.3. Regulation of the Endocannabinoid System in Skin Diseases
3.3.1. Endocannabinoids and Classic Cannabinoid Receptors
3.3.2. Extended Endocannabinoid System
TRP Channels
GPR55, GPR119 and GPR18
PPARs
4. Selected Skin Diseases—Pharmacotherapy and Effect of Cannabinoids
- A Brief Historical Overview on Anti-Inflammatory Effects of Cannabinoids
- A Brief Overview on Anti-Pruritic Effects of Cannabinoids
4.1. Androgenetic Alopecia
4.1.1. Current Therapies of Androgenetic Alopecia
4.1.2. Preclinical Findings on the Effect of Cannabinoids in Androgenetic Alopecia
4.2. Atopic Dermatitis
4.2.1. Current Therapies of Atopic Dermatitis
4.2.2. Preclinical Findings on the Effect of Cannabinoids in Atopic Dermatitis
4.3. Allergic Contact Dermatitis
4.3.1. Current Therapies of Allergic Contact Dermatitis
4.3.2. Preclinical Findings on the Effect of Cannabinoids in Allergic Contact Dermatitis
4.4. Psoriasis
4.4.1. Current Therapies of Psoriasis
4.4.2. Preclinical Findings on the Effect of Cannabinoids in Psoriasis
4.5. Acne
4.5.1. Current Therapies of Acne
4.5.2. Preclinical Findings on the Effect of Cannabinoids in Acne
4.6. Systemic Sclerosis
4.6.1. Current Therapies of Systemic Sclerosis
4.6.2. Preclinical Findings on Effects of Cannabinoids in Skin Fibrosis/Systemic Sclerosis
4.7. Dermatomyositis
4.7.1. Current Therapy of Dermatomyositis
4.7.2. Preclinical Findings on Effects of Cannabinoids in Dermatomyositis
4.8. Epidermolysis Bullosa
4.8.1. Current Therapies of Epidermolysis Bullosa
4.8.2. Case Reports on Cannabinoid Effects on Epidermolysis Bullosa
4.9. Pyoderma Gangrenosum
4.9.1. Current Therapies of Pyoderma Gangrenosum
4.9.2. Case Report on Cannabinoid Effects on Pyoderma Gangrenosum
4.10. Beneficial Effects of Cannabinoids in Other Disorders
4.10.1. Acute Inflammation
4.10.2. Keratin Disorders
4.10.3. Scars and Keloids
5. Clinical Trials with Cannabinoids
5.1. Acne Vulgaris
5.2. Acute Inflammation
5.3. Androgenetic Alopecia
5.4. Asteatotic Eczema
5.5. Atopic Dermatitis
5.6. Atopic Eczema
5.7. Dermatomyositis
5.8. Hidradenitis Suppurativa
5.9. Neurogenic Flare Reaction
5.10. Postherpetic Neuralgia
5.11. Pruritus (Chronic)
5.12. Pruritis (Uremic)
5.13. Psoriasis
5.14. Scalp Inflammation (Scalp Psoriasis/Seborrhoeic Dermatitis)
5.15. Systemic Sclerosis
5.16. Urticaria (Chronic Spontaneous)
5.17. Venous Leg Ulcers
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ramer, R.; Wendt, F.; Wittig, F.; Schäfer, M.; Boeckmann, L.; Emmert, S.; Hinz, B. Impact of Cannabinoid Compounds on Skin Cancer. Cancers 2022, 14, 1769. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.; Kirchhof, M.G. Dermatology-Related Uses of Medical Cannabis Promoted by Dispensaries in Canada, Europe, and the United States. J. Cutan. Med. Surg. 2019, 23, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Karsak, M.; Gaffal, E.; Date, R.; Wang-Eckhardt, L.; Rehnelt, J.; Petrosino, S.; Starowicz, K.; Steuder, R.; Schlicker, E.; Cravatt, B.; et al. Attenuation of allergic contact dermatitis through the endocannabinoid system. Science 2007, 316, 1494–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tóth, K.F.; Ádám, D.; Bíró, T.; Oláh, A. Cannabinoid Signaling in the Skin: Therapeutic Potential of the “C(ut)annabinoid” System. Molecules 2019, 24, 918. [Google Scholar] [CrossRef] [Green Version]
- Haruna, T.; Soga, M.; Morioka, Y.; Imura, K.; Furue, Y.; Yamamoto, M.; Hayakawa, J.; Deguchi, M.; Arimura, A.; Yasui, K. The Inhibitory Effect of S-777469, a Cannabinoid Type 2 Receptor Agonist, on Skin Inflammation in Mice. Pharmacology 2017, 99, 259–267. [Google Scholar] [CrossRef]
- Wilkinson, J.D.; Williamson, E.M. Cannabinoids inhibit human keratinocyte proliferation through a non-CB1/CB2 mechanism and have a potential therapeutic value in the treatment of psoriasis. J. Dermatol. Sci. 2007, 45, 87–92. [Google Scholar] [CrossRef]
- Jarocka-Karpowicz, I.; Biernacki, M.; Wroński, A.; Gęgotek, A.; Skrzydlewska, E. Cannabidiol Effects on Phospholipid Metabolism in Keratinocytes from Patients with Psoriasis Vulgaris. Biomolecules 2020, 10, 367. [Google Scholar] [CrossRef] [Green Version]
- Maddukuri, S.; Patel, J.; Diaz, D.A.; Chen, K.L.; Wysocka, M.; Bax, C.; Li, Y.; Ravishankar, A.; Grinnell, M.; Zeidi, M.; et al. Cannabinoid type 2 receptor (CB2R) distribution in dermatomyositis skin and peripheral blood mononuclear cells (PBMCs) and in vivo efects of LenabasumTM. Arthritis Res. Ther. 2022, 24, 12. [Google Scholar] [CrossRef]
- EU Designation Number: EU/3/18/2070. Available online: https://www.ema.europa.eu/en/medicines/human/orphan-designations/eu3182070 (accessed on 15 June 2022).
- EU Designation Number: EU/3/16/1808. Available online: https://www.ema.europa.eu/en/medicines/human/orphan-designations/eu3161808 (accessed on 15 June 2022).
- Vanti, G.; Grifoni, L.; Bergonzi, M.C.; Antiga, E.; Montefusco, F.; Caproni, M.; Bilia, A.R. Development and optimisation of biopharmaceutical properties of a new microemulgel of cannabidiol for locally-acting dermatological delivery. Int. J. Pharm. 2021, 607, 121036. [Google Scholar] [CrossRef]
- Del Río, C.; Navarrete, C.; Collado, J.A.; Bellido, M.L.; Gómez-Cañas, M.; Pazos, M.R.; Fernández-Ruiz, J.; Pollastro, F.; Appendino, G.; Calzado, M.A.; et al. The cannabinoid quinol VCE-004.8 alleviates bleomycin-induced scleroderma and exerts potent antifibrotic effects through peroxisome proliferator-activated receptor-γ and CB2 pathways. Sci. Rep. 2016, 6, 21703. [Google Scholar] [CrossRef] [Green Version]
- Palomares, B.; Ruiz-Pino, F.; Navarrete, C.; Velasco, I.; Sánchez-Garrido, M.A.; Jimenez-Jimenez, C.; Pavicic, C.; Vazquez, M.J.; Appendino, G.; Bellido, M.L.; et al. VCE-004.8, A Multitarget Cannabinoquinone, Attenuates Adipogenesis and Prevents Diet-Induced Obesity. Sci. Rep. 2018, 8, 16092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Martín, A.; Garrido-Rodríguez, M.; Navarrete, C.; Del Río, C.; Bellido, M.L.; Appendino, G.; Calzado, M.A.; Muñoz, E. EHP-101, an oral formulation of the cannabidiol aminoquinone VCE-004.8, alleviates bleomycin-induced skin and lung fibrosis. Biochem. Pharmacol. 2018, 157, 304–313. [Google Scholar] [CrossRef] [PubMed]
- García-Martín, A.; Garrido-Rodríguez, M.; Navarrete, C.; Caprioglio, D.; Palomares, B.; DeMesa, J.; Rollland, A.; Appendino, G.; Muñoz, E. Cannabinoid derivatives acting as dual PPARγ/CB2 agonists as therapeutic agents for systemic sclerosis. Biochem. Pharmacol. 2019, 163, 321–334. [Google Scholar] [CrossRef] [PubMed]
- U.S. National Library of Medicine, ClinicalTrials.gov. Study of the Safety, Tolerability and Efficacy of BTX 1204 in Patients with Moderate Atopic Dermatitis. ClinicalTrials.gov Identifier: NCT03824405. Available online: https://clinicaltrials.gov/ct2/show/NCT03824405?term=CBD&cond=Skin+Diseases&draw=1&rank=15 (accessed on 15 June 2022).
- Del Rosso, J.Q. Use of a palmitoylethanolamide-containing nonsteroidal cream for treating atopic dermatitis: Impact on the duration of response and time between flares. Cosmetic Dermatol. 2007, 20, 208–211. [Google Scholar]
- Gao, Y.; Li, Y.; Tan, Y.; Liu, W.; Ouaddi, S.; McCoy, J.; Kovacevic, M.; Situm, M.; Stanimirovic, A.; Li, M.; et al. Novel cannabidiol aspartame combination treatment (JW-100) significantly reduces ISGA score in atopic dermatitis: Results from a randomized double-blinded placebo-controlled interventional study. J. Cosmet. Dermatol. 2021, 21, 1647–1650. [Google Scholar] [CrossRef] [PubMed]
- Schräder, N.H.B.; Duipmans, J.C.; Molenbuur, B.; Wolff, A.P.; Jonkman, M.F. Combined tetrahydrocannabinol and cannabidiol to treat pain in epidermolysis bullosa: A report of three cases. Br. J. Dermatol. 2019, 180, 922–924. [Google Scholar] [CrossRef] [Green Version]
- Schräder, N.H.B.; Gorell, E.S.; Stewart, R.E.; Duipmans, J.C.; Harris, N.; Perez, V.A.; Tang, J.Y.; Wolff, A.P.; Bolling, M.C. Cannabinoid use and effects in patients with epidermolysis bullosa: An international cross-sectional survey study. Orphanet J. Rare Dis. 2021, 16, 377. [Google Scholar] [CrossRef] [PubMed]
- Maida, V.; Corban, J. Topical Medical Cannabis: A New Treatment for Wound Pain-Three Cases of Pyoderma Gangrenosum. J. Pain Symptom Manag. 2017, 54, 732–736. [Google Scholar] [CrossRef] [Green Version]
- Chelliah, M.P.; Zinn, Z.; Khuu, P.; Teng, J.M.C. Self-initiated use of topical cannabidiol oil for epidermolysis bullosa. Pediatr. Dermatol. 2018, 35, e224–e227. [Google Scholar] [CrossRef]
- Neff, G.W.; O’Brien, C.B.; Reddy, K.R.; Bergasa, N.V.; Regev, A.; Molina, E.; Amaro, R.; Rodriguez, M.J.; Chase, V.; Jeffers, L.; et al. Preliminary observation with dronabinol in patients with intractable pruritus secondary to cholestatic liver disease. Am. J. Gastroenterol. 2002, 97, 2117–2119. [Google Scholar] [CrossRef]
- Laborada, J.; Cohen, P.R. Cutaneous Squamous Cell Carcinoma and Lichen Simplex Chronicus Successfully Treated with Topical Cannabinoid Oil: A Case Report and Summary of Cannabinoids in Dermatology. Cureus 2022, 14, e23850. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Carlini, E.A. Toward drugs derived from cannabis. Naturwissenschaften 1978, 65, 174–179. [Google Scholar] [CrossRef]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, L.A.; Lolait, S.J.; Brownstein, M.J.; Young, A.C.; Bonner, T.I. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990, 346, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G. The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: Δ9-tetrahydrocannabinol, cannabidiol and Δ9-tetrahydrocannabivarin. Br. J. Pharmacol. 2008, 153, 199–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laprairie, R.B.; Bagher, A.M.; Kelly, M.E.M.; Denovan-Wright, E.M. Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor. Br. J. Pharmacol. 2015, 172, 4790–4805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechoulam, R.; Ben-Shabat, S.; Hanus, L.; Ligumsky, M.; Kaminski, N.E.; Schatz, A.R.; Gopher, A.; Almog, S.; Martin, B.R.; Compton, D.R. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol. 1995, 50, 83–90. [Google Scholar] [CrossRef]
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef]
- Mackie, K.; Devane, W.A.; Hille, B. Anandamide, an endogenous cannabinoid, inhibits calcium currents as a partial agonist in N18 neuroblastoma cells. Mol. Pharmacol. 1993, 44, 498–503. [Google Scholar]
- Sugiura, T.; Kodaka, T.; Nakane, S.; Miyashita, T.; Kondo, S.; Suhara, Y.; Takayama, H.; Waku, K.; Seki, C.; Baba, N.; et al. Evidence that the cannabinoid CB1 receptor is a 2-arachidonoylglycerol receptor. Structure-activity relationship of 2-arachidonoylglycerol, ether-linked analogues, and related compounds. J. Biol. Chem. 1999, 274, 2794–2801. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, T.; Kondo, S.; Kishimoto, S.; Miyashita, T.; Nakane, S.; Kodaka, T.; Suhara, Y.; Takayama, H.; Waku, K. Evidence that 2-arachidonoylglycerol but not N-palmitoylethanolamine or anandamide is the physiological ligand for the cannabinoid CB2 receptor. Comparison of the agonistic activities of various cannabinoid receptor ligands in HL-60 cells. J. Biol. Chem. 2000, 275, 605–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokorski, M.; Matysiak, Z. Fatty acid acylation of dopamine in the carotid body. Med. Hypotheses 1998, 50, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Bisogno, T.; Melck, D.; Bobrov, M.; Gretskaya, N.M.; Bezuglov, V.V.; De Petrocellis, L.; Di Marzo, V. N-acyl-dopamines: Novel synthetic CB1 cannabinoid-receptor ligands and inhibitors of anandamide inactivation with cannabimimetic activity in vitro and in vivo. Biochem. J. 2000, 351, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.M.; Bisogno, T.; Trevisani, M.; Al-Hayani, A.; De Petrocellis, L.; Fezza, F.; Tognetto, M.; Petros, T.J.; Krey, J.F.; Chu, C.J.; et al. An endogenous capsaicin-like substance with high potency at recombinant and native vanilloid VR1 receptors. Proc. Natl. Acad. Sci. USA 2002, 99, 8400–8405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanus, L.; Abu-Lafi, S.; Fride, E.; Breuer, A.; Vogel, Z.; Shalev, D.E.; Kustanovich, I.; Mechoulam, R. 2-arachidonyl glyceryl ether, an endogenous agonist of the cannabinoid CB1 receptor. Proc. Natl. Acad. Sci. USA 2001, 98, 3662–3665. [Google Scholar] [CrossRef] [Green Version]
- Porter, A.C.; Sauer, J.-M.; Knierman, M.D.; Becker, G.W.; Berna, M.J.; Bao, J.; Nomikos, G.G.; Carter, P.; Bymaster, F.P.; Leese, A.B.; et al. Characterization of a novel endocannabinoid, virodhamine, with antagonist activity at the CB1 receptor. J. Pharmacol. Exp. Ther. 2002, 301, 1020–1024. [Google Scholar] [CrossRef] [Green Version]
- Venn-Watson, S.; Reiner, J.; Jensen, E.D. Pentadecanoylcarnitine is a newly discovered endocannabinoid with pleiotropic activities relevant to supporting physical and mental health. Sci. Rep. 2022, 12, 13717. [Google Scholar] [CrossRef]
- Maccarrone, M.; Attinà, M.; Cartoni, A.; Bari, M.; Finazzi-Agrò, A. Gas chromatography-mass spectrometry analysis of endogenous cannabinoids in healthy and tumoral human brain and human cells in culture. J. Neurochem. 2001, 76, 594–601. [Google Scholar] [CrossRef]
- Schwarz, R.; Ramer, R.; Hinz, B. Targeting the endocannabinoid system as a potential anticancer approach. Drug Metab. Rev. 2018, 50, 26–53. [Google Scholar] [CrossRef]
- Di Marzo, V.; Melck, D.; Orlando, P.; Bisogno, T.; Zagoory, O.; Bifulco, M.; Vogel, Z.; De Petrocellis, L. Palmitoylethanolamide inhibits the expression of fatty acid amide hydrolase and enhances the anti-proliferative effect of anandamide in human breast cancer cells. Biochem. J. 2001, 358, 249–255. [Google Scholar] [CrossRef]
- De Petrocellis, L.; Bisogno, T.; Ligresti, A.; Bifulco, M.; Melck, D.; Di Marzo, V. Effect on cancer cell proliferation of palmitoylethanolamide, a fatty acid amide interacting with both the cannabinoid and vanilloid signalling systems. Fundam. Clin. Pharmacol. 2002, 16, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M.; Pauselli, R.; Di Rienzo, M.; Finazzi-Agrò, A. Binding, degradation and apoptotic activity of stearoylethanolamide in rat C6 glioma cells. Biochem. J. 2002, 366, 137–144. [Google Scholar] [CrossRef]
- Ben-Shabat, S.; Fride, E.; Sheskin, T.; Tamiri, T.; Rhee, M.-H.; Vogel, Z.; Bisogno, T.; De Petrocellis, L.; Di Marzo, V.; Mechoulam, R. An entourage effect: Inactive endogenous fatty acid glycerol esters enhance 2-arachidonoyl-glycerol cannabinoid activity. Eur. J. Pharmacol. 1998, 353, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V. The endocannabinoid system: Its general strategy of action, tools for its pharmacological manipulation and potential therapeutic exploitation. Pharmacol. Res. 2009, 60, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, D.G.; Chin, S.A. Enzymatic synthesis and degradation of anandamide, a cannabinoid receptor agonist. Biochem. Pharmacol. 1993, 46, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Blankman, J.L.; Simon, G.M.; Cravatt, B.F. A comprehensive profile of brain enzymes that hydrolyze the endocannabinoid 2-arachidonoylglycerol. Chem. Biol. 2007, 14, 1347–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuboi, K.; Sun, Y.-X.; Okamoto, Y.; Araki, N.; Tonai, T.; Ueda, N. Molecular characterization of N-acylethanolamine-hydrolyzing acid amidase, a novel member of the choloylglycine hydrolase family with structural and functional similarity to acid ceramidase. J. Biol. Chem. 2005, 280, 11082–11092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, A.; Pontis, S.; Mengatto, L.; Armirotti, A.; Chiurchiù, V.; Capurro, V.; Fiasella, A.; Nuzzi, A.; Romeo, E.; Moreno-Sanz, G.; et al. A Potent Systemically Active N-Acylethanolamine Acid Amidase Inhibitor that Suppresses Inflammation and Human Macrophage Activation. ACS Chem. Biol. 2015, 10, 1838–1846. [Google Scholar] [CrossRef] [Green Version]
- Zygmunt, P.M.; Petersson, J.; Andersson, D.A.; Chuang, H.; Sørgård, M.; Di Marzo, V.; Julius, D.; Högestätt, E.D. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 1999, 400, 452–457. [Google Scholar] [CrossRef]
- Bisogno, T.; Hanus, L.; De Petrocellis, L.; Tchilibon, S.; Ponde, D.E.; Brandi, I.; Moriello, A.S.; Davis, J.B.; Mechoulam, R.; Di Marzo, V. Molecular targets for cannabidiol and its synthetic analogues: Effect on vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide. Br. J. Pharmacol. 2001, 134, 845–852. [Google Scholar] [CrossRef]
- Duncan, M.; Millns, P.; Smart, D.; Wright, J.E.; Kendall, D.A.; Ralevic, V. Noladin ether, a putative endocannabinoid, attenuates sensory neurotransmission in the rat isolated mesenteric arterial bed via a non-CB1/CB2 Gi/o linked receptor. Br. J. Pharmacol. 2004, 142, 509–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Meo, C.; Tortolani, D.; Standoli, S.; Angelucci, C.B.; Fanti, F.; Leuti, A.; Sergi, M.; Kadhim, S.; Hsu, E.; Rapino, C.; et al. Effects of Rare Phytocannabinoids on the Endocannabinoid System of Human Keratinocytes. Int. J. Mol. Sci. 2022, 23, 5430. [Google Scholar] [CrossRef] [PubMed]
- Qin, N.; Neeper, M.P.; Liu, Y.; Hutchinson, T.L.; Lubin, M.L.; Flores, C.M. TRPV2 is activated by cannabidiol and mediates CGRP release in cultured rat dorsal root ganglion neurons. J. Neurosci. 2008, 28, 6231–6238. [Google Scholar] [CrossRef] [Green Version]
- De Petrocellis, L.; Ligresti, A.; Moriello, A.S.; Allarà, M.; Bisogno, T.; Petrosino, S.; Stott, C.G.; Di Marzo, V. Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Br. J. Pharmacol. 2011, 163, 1479–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Petrocellis, L.; Orlando, P.; Moriello, A.S.; Aviello, G.; Stott, C.; Izzo, A.A.; Di Marzo, V. Cannabinoid actions at TRPV channels: Effects on TRPV3 and TRPV4 and their potential relevance to gastrointestinal inflammation. Acta Physiol. 2012, 204, 255–266. [Google Scholar] [CrossRef]
- Yang, H.; Zhou, J.; Lehmann, C. GPR55—A putative “type 3” cannabinoid receptor in inflammation. J. Basic Clin. Physiol. Pharmacol. 2016, 27, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Gómez, E.; Andradas, C.; Flores, J.M.; Quintanilla, M.; Paramio, J.M.; Guzmán, M.; Sánchez, C. The orphan receptor GPR55 drives skin carcinogenesis and is upregulated in human squamous cell carcinomas. Oncogene 2013, 32, 2534–2542. [Google Scholar] [CrossRef] [Green Version]
- Ryberg, E.; Larsson, N.; Sjögren, S.; Hjorth, S.; Hermansson, N.-O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The orphan receptor GPR55 is a novel cannabinoid receptor. Br. J. Pharmacol. 2007, 152, 1092–1101. [Google Scholar] [CrossRef]
- Lauckner, J.E.; Jensen, J.B.; Chen, H.-Y.; Lu, H.-C.; Hille, B.; Mackie, K. GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc. Natl. Acad. Sci. USA 2008, 105, 2699–2704. [Google Scholar] [CrossRef] [Green Version]
- Kapur, A.; Zhao, P.; Sharir, H.; Bai, Y.; Caron, M.G.; Barak, L.S.; Abood, M.E. Atypical responsiveness of the orphan receptor GPR55 to cannabinoid ligands. J. Biol. Chem. 2009, 284, 29817–29827. [Google Scholar] [CrossRef] [Green Version]
- Overton, H.A.; Babbs, A.J.; Doel, S.M.; Fyfe, M.C.T.; Gardner, L.S.; Griffin, G.; Jackson, H.C.; Procter, M.J.; Rasamison, C.M.; Tang-Christensen, M.; et al. Deorphanization of a G protein-coupled receptor for oleoylethanolamide and its use in the discovery of small-molecule hypophagic agents. Cell Metab. 2006, 3, 167–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajaraman, G.; Simcocks, A.; Hryciw, D.H.; Hutchinson, D.S.; McAinch, A.J. G protein coupled receptor 18: A potential role for endocannabinoid signaling in metabolic dysfunction. Mol. Nutr. Food Res. 2016, 60, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Kohno, M.; Hasegawa, H.; Inoue, A.; Muraoka, M.; Miyazaki, T.; Oka, K.; Yasukawa, M. Identification of N-arachidonylglycine as the endogenous ligand for orphan G-protein-coupled receptor GPR18. Biochem. Biophys. Res. Commun. 2006, 347, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Rimmerman, N.; Bradshaw, H.B.; Hughes, H.V.; Chen, J.S.-C.; Hu, S.S.-J.; McHugh, D.; Vefring, E.; Jahnsen, J.A.; Thompson, E.L.; Masuda, K.; et al. N-palmitoyl glycine, a novel endogenous lipid that acts as a modulator of calcium influx and nitric oxide production in sensory neurons. Mol. Pharmacol. 2008, 74, 213–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHugh, D.; Page, J.; Dunn, E.; Bradshaw, H.B. Δ9-Tetrahydrocannabinol and N-arachidonyl glycine are full agonists at GPR18 receptors and induce migration in human endometrial HEC-1B cells. Br. J. Pharmacol. 2012, 165, 2414–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Console-Bram, L.; Brailoiu, E.; Brailoiu, G.C.; Sharir, H.; Abood, M.E. Activation of GPR18 by cannabinoid compounds: A tale of biased agonism. Br. J. Pharmacol. 2014, 171, 3908–3917. [Google Scholar] [CrossRef] [Green Version]
- Blunder, S.; Pavel, P.; Minzaghi, D.; Dubrac, S. PPARdelta in Affected Atopic Dermatitis and Psoriasis: A Possible Role in Metabolic Reprograming. Int. J. Mol. Sci. 2021, 22, 7354. [Google Scholar] [CrossRef]
- Fu, J.; Gaetani, S.; Oveisi, F.; Lo Verme, J.; Serrano, A.; Rodríguez de Fonseca, F.; Rosengarth, A.; Luecke, H.; Di Giacomo, B.; Tarzia, G.; et al. Oleylethanolamide regulates feeding and body weight through activation of the nuclear receptor PPAR-α. Nature 2003, 425, 90–93. [Google Scholar] [CrossRef]
- Lo Verme, J.; Fu, J.; Astarita, G.; La Rana, G.; Russo, R.; Calignano, A.; Piomelli, D. The nuclear receptor peroxisome proliferator-activated receptor-α mediates the anti-inflammatory actions of palmitoylethanolamide. Mol. Pharmacol. 2005, 67, 15–19. [Google Scholar] [CrossRef]
- Sarnelli, G.; Gigli, S.; Capoccia, E.; Iuvone, T.; Cirillo, C.; Seguella, L.; Nobile, N.; D’Alessandro, A.; Pesce, M.; Steardo, L.; et al. Palmitoylethanolamide Exerts Antiproliferative Effect and Downregulates VEGF Signaling in Caco-2 Human Colon Carcinoma Cell Line Through a Selective PPAR-α-Dependent Inhibition of Akt/mTOR Pathway. Phytother. Res. 2016, 30, 963–970. [Google Scholar] [CrossRef]
- Takeda, S.; Ikeda, E.; Su, S.; Harada, M.; Okazaki, H.; Yoshioka, Y.; Nishimura, H.; Ishii, H.; Kakizoe, K.; Taniguchi, A.; et al. Δ9-THC modulation of fatty acid 2-hydroxylase (FA2H) gene expression: Possible involvement of induced levels of PPARα in MDA-MB-231 breast cancer cells. Toxicology 2014, 326, 18–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Alexander, S.P.H.; Garle, M.J.; Gibson, C.L.; Hewitt, K.; Murphy, S.P.; Kendall, D.A.; Bennett, A.J. Cannabinoid activation of PPAR alpha; a novel neuroprotective mechanism. Br. J. Pharmacol. 2007, 152, 734–743. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.E.; Tarling, E.J.; Bennett, A.J.; Kendall, D.A.; Randall, M.D. Novel time-dependent vascular actions of Δ9-tetrahydrocannabinol mediated by peroxisome proliferator-activated receptor gamma. Biochem. Biophys. Res. Commun. 2005, 337, 824–831. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.E.; Sun, Y.; Bennett, A.J.; Randall, M.D.; Kendall, D.A. Time-dependent vascular actions of cannabidiol in the rat aorta. Eur. J. Pharmacol. 2009, 612, 61–68. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.E. Cannabinoids go nuclear: Evidence for activation of peroxisome proliferator-activated receptors. Br. J. Pharmacol. 2007, 152, 576–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Li, H.; Burstein, S.H.; Zurier, R.B.; Chen, J.D. Activation and binding of peroxisome proliferator-activated receptor γ by synthetic cannabinoid ajulemic acid. Mol. Pharmacol. 2003, 63, 983–992. [Google Scholar] [CrossRef] [Green Version]
- Vara, D.; Morell, C.; Rodríguez-Henche, N.; Diaz-Laviada, I. Involvement of PPARγ in the antitumoral action of cannabinoids on hepatocellular carcinoma. Cell Death Dis. 2013, 4, e618. [Google Scholar] [CrossRef] [Green Version]
- Hirao-Suzuki, M.; Takeda, S.; Koga, T.; Takiguchi, M.; Toda, A. Cannabidiolic acid dampens the expression of cyclooxygenase-2 in MDA-MB-231 breast cancer cells: Possible implication of the peroxisome proliferator-activated receptor β/δ abrogation. J. Toxicol. Sci. 2020, 45, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Hirao-Suzuki, M.; Takeda, S.; Watanabe, K.; Takiguchi, M.; Aramaki, H. Δ9-Tetrahydrocannabinol upregulates fatty acid 2-hydroxylase (FA2H) via PPARα induction: A possible evidence for the cancellation of PPARβ/δ-mediated inhibition of PPARα in MDA-MB-231 cells. Arch. Biochem. Biophys. 2019, 662, 219–225. [Google Scholar] [CrossRef]
- Felder, C.C.; Nielsen, A.; Briley, E.M.; Palkovits, M.; Priller, J.; Axelrod, J.; Nguyen, D.N.; Richardson, J.M.; Riggin, R.M.; Koppel, G.A.; et al. Isolation and measurement of the endogenous cannabinoid receptor agonist, anandamide, in brain and peripheral tissues of human and rat. FEBS Lett. 1996, 393, 231–235. [Google Scholar] [CrossRef] [Green Version]
- Calignano, A.; La Rana, G.; Giuffrida, A.; Piomelli, D. Control of pain initiation by endogenous cannabinoids. Nature 1998, 394, 277–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czifra, G.; Szöllősi, A.G.; Tóth, B.I.; Demaude, J.; Bouez, C.; Breton, L.; Bíró, T. Endocannabinoids regulate growth and survival of human eccrine sweat gland-derived epithelial cells. J. Investig. Dermatol. 2012, 132, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Telek, A.; Bíró, T.; Bodó, E.; Tóth, B.I.; Borbíró, I.; Kunos, G.; Paus, R. Inhibition of human hair follicle growth by endo- and exocannabinoids. FASEB J. 2007, 21, 3534–3541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ständer, S.; Schmelz, M.; Metze, D.; Luger, T.; Rukwied, R. Distribution of cannabinoid receptor 1 (CB1) and 2 (CB2) on sensory nerve fibers and adnexal structures in human skin. J. Dermatol. Sci. 2005, 38, 177–188. [Google Scholar] [CrossRef]
- Dobrosi, N.; Tóth, B.I.; Nagy, G.; Dózsa, A.; Géczy, T.; Nagy, L.; Zouboulis, C.C.; Paus, R.; Kovács, L.; Bíró, T. Endocannabinoids enhance lipid synthesis and apoptosis of human sebocytes via cannabinoid receptor-2-mediated signaling. FASEB J. 2008, 22, 3685–3695. [Google Scholar] [CrossRef] [Green Version]
- Pucci, M.; Pasquariello, N.; Battista, N.; Di Tommaso, M.; Rapino, C.; Fezza, F.; Zuccolo, M.; Jourdain, R.; Finazzi Agrò, A.; Breton, L.; et al. Endocannabinoids stimulate human melanogenesis via type-1 cannabinoid receptor. J. Biol. Chem. 2012, 287, 15466–15478. [Google Scholar] [CrossRef] [Green Version]
- Fujii, N.; Kenny, G.P.; Amano, T.; Honda, Y.; Kondo, N.; Nishiyasu, T. Evidence for TRPV4 channel induced skin vasodilatation through NOS, COX, and KCa channel mechanisms with no effect on sweat rate in humans. Eur. J. Pharmacol. 2019, 858, 172462. [Google Scholar] [CrossRef]
- Borbíró, I.; Lisztes, E.; Tóth, B.I.; Czifra, G.; Oláh, A.; Szöllosi, A.G.; Szentandrássy, N.; Nánási, P.P.; Péter, Z.; Paus, R.; et al. Activation of transient receptor potential vanilloid-3 inhibits human hair growth. J. Investig. Dermatol. 2011, 131, 1605–1614. [Google Scholar] [CrossRef]
- Miragliotta, V.; Ricci, P.L.; Albanese, F.; Pirone, A.; Tognotti, D.; Abramo, F. Cannabinoid receptor types 1 and 2 and peroxisome proliferator-activated receptor-α: Distribution in the skin of clinically healthy cats and cats with hypersensitivity dermatitis. Vet. Dermatol. 2018, 29, 316-e111. [Google Scholar] [CrossRef]
- Billoni, N.; Buan, B.; Gautier, B.; Collin, C.; Gaillard, O.; Mahé, Y.F.; Bernard, B.A. Expression of peroxisome proliferator activated receptors (PPARs) in human hair follicles and PPAR alpha involvement in hair growth. Acta Derm. Venereol. 2000, 80, 329–334. [Google Scholar]
- Westergaard, M.; Henningsen, J.; Svendsen, M.L.; Johansen, C.; Jensen, U.B.; Schrøder, H.D.; Kratchmarova, I.; Berge, R.K.; Iversen, L.; Bolund, L.; et al. Modulation of keratinocyte gene expression and differentiation by PPAR-selective ligands and tetradecylthioacetic acid. J. Investig. Dermatol. 2001, 116, 702–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmuth, M.; Moosbrugger-Martinz, V.; Blunder, S.; Dubrac, S. Role of PPAR, LXR, and PXR in epidermal homeostasis and inflammation. Biochim. Biophys. Acta 2014, 1841, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Alestas, T.; Ganceviciene, R.; Fimmel, S.; Müller-Decker, K.; Zouboulis, C.C. Enzymes involved in the biosynthesis of leukotriene B4 and prostaglandin E2 are active in sebaceous glands. J. Mol. Med. 2006, 84, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Wang, T.-T.; Quan, J.-H.; Li, S.-J.; Zhang, M.-F.; Liao, P.-Y.; Fan, Y.-M. Sox9 facilitates proliferation, differentiation and lipogenesis in primary cultured human sebocytes. J. Dermatol. Sci. 2017, 85, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Ghosh, A.K.; Sargent, J.L.; Komura, K.; Wu, M.; Huang, Q.-Q.; Jain, M.; Whitfield, M.L.; Feghali-Bostwick, C.; Varga, J. PPARγ downregulation by TGFß in fibroblast and impaired expression and function in systemic sclerosis: A novel mechanism for progressive fibrogenesis. PLoS ONE 2010, 5, e13778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markovics, A.; Angyal, Á.; Tóth, K.F.; Ádám, D.; Pénzes, Z.; Magi, J.; Pór, Á.; Kovács, I.; Törőcsik, D.; Zouboulis, C.C.; et al. GPR119 Is a Potent Regulator of Human Sebocyte Biology. J. Investig. Dermatol. 2020, 140, 1909–1918.e8. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.S.; Kim, Y.-J.; Kim, M.O.; Kang, M.; Oh, S.W.; Nho, Y.H.; Park, S.-H.; Lee, J. Cannabidiol upregulates melanogenesis through CB1 dependent pathway by activating p38 MAPK and p42/44 MAPK. Chem. Biol. Interact. 2017, 273, 107–114. [Google Scholar] [CrossRef]
- Magina, S.; Esteves-Pinto, C.; Moura, E.; Serrão, M.P.; Moura, D.; Petrosino, S.; Di Marzo, V.; Vieira-Coelho, M.A. Inhibition of basal and ultraviolet B-induced melanogenesis by cannabinoid CB1 receptors: A keratinocyte-dependent effect. Arch. Dermatol. Res. 2011, 303, 201–210. [Google Scholar] [CrossRef]
- Goenka, S. Comparative Study of Δ9-Tetrahydrocannabinol and Cannabidiol on Melanogenesis in Human Epidermal Melanocytes from Different Pigmentation Phototypes: A Pilot Study. J. Xenobiot. 2022, 12, 131–144. [Google Scholar] [CrossRef]
- Weigelt, M.A.; Sivamani, R.; Lev-Tov, H. The therapeutic potential of cannabinoids for integumentary wound management. Exp. Dermatol. 2021, 30, 201–211. [Google Scholar] [CrossRef]
- Ruhl, T.; Lippold, E.F.; Christer, T.; Schaefer, B.; Kim, B.-S.; Beier, J.P. Genetic deletion of the cannabinoid receptors CB1 and CB2 enhances inflammation with diverging effects on skin wound healing in mice. Life Sci. 2021, 285, 120018. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Ren, P.; Wang, Q.; Jiang, S.-K.; Zhang, M.; Li, J.-Y.; Wang, L.-L.; Guan, D.-W. Cannabinoid 2 receptor attenuates inflammation during skin wound healing by inhibiting M1 macrophages rather than activating M2 macrophages. J. Inflamm. 2018, 15, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.-L.; Zhao, R.; Li, J.-Y.; Li, S.-S.; Liu, M.; Wang, M.; Zhang, M.-Z.; Dong, W.-W.; Jiang, S.-K.; Zhang, M.; et al. Pharmacological activation of cannabinoid 2 receptor attenuates inflammation, fibrogenesis, and promotes re-epithelialization during skin wound healing. Eur. J. Pharmacol. 2016, 786, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-S.; Wang, L.-L.; Liu, M.; Jiang, S.-K.; Zhang, M.; Tian, Z.-L.; Wang, M.; Li, J.-Y.; Zhao, R.; Guan, D.-W. Cannabinoid CB₂ receptors are involved in the regulation of fibrogenesis during skin wound repair in mice. Mol. Med. Rep. 2016, 13, 3441–3450. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Shen, J.; Zhang, L.; Wang, L.; Xu, H.; Han, Y.; Jia, J.; Lu, Y.; Yu, R.; Liu, H. Injectable postoperative enzyme-responsive hydrogels for reversing temozolomide resistance and reducing local recurrence after glioma operation. Biomater. Sci. 2020, 8, 5306–5316. [Google Scholar] [CrossRef]
- Zhao, C.; Dong, Y.; Liu, J.; Cai, H.; Li, Z.; Sun, X.; Yin, W.; Ma, J.; Liu, H.; Li, S. An enzyme-responsive Gp1a-hydrogel for skin wound healing. J. Biomater. Appl. 2021, 36, 714–721. [Google Scholar] [CrossRef]
- Koyama, S.; Purk, A.; Kaur, M.; Soini, H.A.; Novotny, M.V.; Davis, K.; Kao, C.C.; Matsunami, H.; Mescher, A. Beta-caryophyllene enhances wound healing through multiple routes. PLoS ONE 2019, 14, e0216104. [Google Scholar] [CrossRef] [Green Version]
- Ramot, Y.; Sugawara, K.; Zákány, N.; Tóth, B.I.; Bíró, T.; Paus, R. A novel control of human keratin expression: Cannabinoid receptor 1-mediated signaling down-regulates the expression of keratins K6 and K16 in human keratinocytes in vitro and in situ. PeerJ 2013, 1, e40. [Google Scholar] [CrossRef] [Green Version]
- Siracusa, R.; Impellizzeri, D.; Cordaro, M.; Gugliandolo, E.; Peritore, A.F.; Di Paola, R.; Cuzzocrea, S. Topical Application of Adelmidrol + Trans-Traumatic Acid Enhances Skin Wound Healing in a Streptozotocin-Induced Diabetic Mouse Model. Front. Pharmacol. 2018, 9, 871. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Di Paola, R.; Cordaro, M.; Gugliandolo, E.; Casili, G.; Morittu, V.M.; Britti, D.; Esposito, E.; Cuzzocrea, S. Adelmidrol, a palmitoylethanolamide analogue, as a new pharmacological treatment for the management of acute and chronic inflammation. Biochem. Pharmacol. 2016, 119, 27–41. [Google Scholar] [CrossRef]
- Sasso, O.; Pontis, S.; Armirotti, A.; Cardinali, G.; Kovacs, D.; Migliore, M.; Summa, M.; Moreno-Sanz, G.; Picardo, M.; Piomelli, D. Endogenous N-acyl taurines regulate skin wound healing. Proc. Natl. Acad. Sci. USA 2016, 113, E4397–E4406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lüder, E.; Ramer, R.; Peters, K.; Hinz, B. Decisive role of P42/44 mitogen-activated protein kinase in Δ9-tetrahydrocannabinol-induced migration of human mesenchymal stem cells. Oncotarget 2017, 8, 105984–105994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmuhl, E.; Ramer, R.; Salamon, A.; Peters, K.; Hinz, B. Increase of mesenchymal stem cell migration by cannabidiol via activation of p42/44 MAPK. Biochem. Pharmacol. 2014, 87, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Wollank, Y.; Ramer, R.; Ivanov, I.; Salamon, A.; Peters, K.; Hinz, B. Inhibition of FAAH confers increased stem cell migration via PPARα. J. Lipid Res. 2015, 56, 1947–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Styrczewska, M.; Kostyn, A.; Kulma, A.; Majkowska-Skrobek, G.; Augustyniak, D.; Prescha, A.; Czuj, T.; Szopa, J. Flax Fiber Hydrophobic Extract Inhibits Human Skin Cells Inflammation and Causes Remodeling of Extracellular Matrix and Wound Closure Activation. Biomed Res. Int. 2015, 2015, 862391. [Google Scholar] [CrossRef] [Green Version]
- Haskó, J.; Fazakas, C.; Molnár, J.; Nyúl-Tóth, Á.; Herman, H.; Hermenean, A.; Wilhelm, I.; Persidsky, Y.; Krizbai, I.A. CB2 receptor activation inhibits melanoma cell transmigration through the blood-brain barrier. Int. J. Mol. Sci. 2014, 15, 8063–8074. [Google Scholar] [CrossRef] [Green Version]
- Vaseghi, G.; Taki, M.J.; Javanmard, S.H. Standardized Cannabis sativa extract attenuates tau and stathmin gene expression in the melanoma cell line. Iran. J. Basic Med. Sci. 2017, 20, 1178–1181. [Google Scholar]
- Zhao, Z.; Yang, J.; Zhao, H.; Fang, X.; Li, H. Cannabinoid receptor 2 is upregulated in melanoma. J. Cancer Res. Ther. 2012, 8, 549–554. [Google Scholar] [CrossRef]
- Sailler, S.; Schmitz, K.; Jäger, E.; Ferreiros, N.; Wicker, S.; Zschiebsch, K.; Pickert, G.; Geisslinger, G.; Walter, C.; Tegeder, I.; et al. Regulation of circulating endocannabinoids associated with cancer and metastases in mice and humans. Oncoscience 2014, 1, 272–282. [Google Scholar] [CrossRef] [Green Version]
- Hohmann, U.; Walsleben, C.; Ghadban, C.; Kirchhoff, F.; Dehghani, F.; Hohmann, T. Interaction of Glia Cells with Glioblastoma and Melanoma Cells under the Influence of Phytocannabinoids. Cells 2022, 11, 147. [Google Scholar] [CrossRef]
- Carpi, S.; Fogli, S.; Polini, B.; Montagnani, V.; Podestà, A.; Breschi, M.C.; Romanini, A.; Stecca, B.; Nieri, P. Tumor-promoting effects of cannabinoid receptor type 1 in human melanoma cells. Toxicol. Vitro 2017, 40, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Qi, J.; Hu, L.; Ouyang, D.; Wang, H.; Sun, Q.; Lin, L.; You, L.; Tang, B. A cannabidiol-containing alginate based hydrogel as novel multifunctional wound dressing for promoting wound healing. Biomater. Adv. 2022, 134, 112560. [Google Scholar] [CrossRef] [PubMed]
- Maida, V.; Shi, R.B.; Fazzari, F.G.T.; Zomparelli, L. Topical cannabis-based medicines—A novel adjuvant treatment for venous leg ulcers: An open-label trial. Exp. Dermatol. 2021, 30, 1258–1267. [Google Scholar] [CrossRef] [PubMed]
- Roelandt, T.; Heughebaert, C.; Bredif, S.; Giddelo, C.; Baudouin, C.; Msika, P.; Roseeuw, D.; Uchida, Y.; Elias, P.M.; Hachem, J.-P. Cannabinoid receptors 1 and 2 oppositely regulate epidermal permeability barrier status and differentiation. Exp. Dermatol. 2012, 21, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, B.; Park, B.M.; Jeon, J.E.; Lee, S.H.; Mann, S.; Ahn, S.K.; Hong, S.-P.; Jeong, S.K. Topical cannabinoid receptor 1 agonist attenuates the cutaneous inflammatory responses in oxazolone-induced atopic dermatitis model. Int. J. Dermatol. 2015, 54, e401–e408. [Google Scholar] [CrossRef]
- Proksch, E.; Soeberdt, M.; Neumann, C.; Kilic, A.; Abels, C. Modulators of the endocannabinoid system influence skin barrier repair, epidermal proliferation, differentiation and inflammation in a mouse model. Exp. Dermatol. 2019, 28, 1058–1065. [Google Scholar] [CrossRef] [PubMed]
- Mehling, A.; Chkarnat, C.; Degwert, J.; Ennen, J.; Fink, E.; Matthies, W.; Roethlisberger, R.; Rossow, U.; Schnitker, J.; Tronnier, H.; et al. Interlaboratory studies with a proposed patch test design to evaluate the irritation potential of surfactants. Contact Dermat. 2010, 62, 157–164. [Google Scholar] [CrossRef]
- Hinrichsen, K.; Podschun, R.; Schubert, S.; Schröder, J.M.; Harder, J.; Proksch, E. Mouse beta-defensin-14, an antimicrobial ortholog of human beta-defensin-3. Antimicrob. Agents Chemother. 2008, 52, 1876–1879. [Google Scholar] [CrossRef] [Green Version]
- Shin, K.-O.; Kim, S.; Park, B.D.; Uchida, Y.; Park, K. N-Palmitoyl Serinol Stimulates Ceramide Production through a CB1-Dependent Mechanism in In Vitro Model of Skin Inflammation. Int. J. Mol. Sci. 2021, 22, 8302. [Google Scholar] [CrossRef]
- Ramer, R.; Weinzierl, U.; Schwind, B.; Brune, K.; Hinz, B. Ceramide is involved in R(+)-methanandamide-induced cyclooxygenase-2 expression in human neuroglioma cells. Mol. Pharmacol. 2003, 64, 1189–1198. [Google Scholar] [CrossRef] [Green Version]
- Galve-Roperh, I.; Sánchez, C.; Cortés, M.L.; Del Gómez Pulgar, T.; Izquierdo, M.; Guzmán, M. Anti-tumoral action of cannabinoids: Involvement of sustained ceramide accumulation and extracellular signal-regulated kinase activation. Nat. Med. 2000, 6, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Ikarashi, N.; Shiseki, M.; Yoshida, R.; Tabata, K.; Kimura, R.; Watanabe, T.; Kon, R.; Sakai, H.; Kamei, J. Cannabidiol Application Increases Cutaneous Aquaporin-3 and Exerts a Skin Moisturizing Effect. Pharmaceuticals 2021, 14, 879. [Google Scholar] [CrossRef] [PubMed]
- Konger, R.L.; Derr-Yellin, E.; Zimmers, T.A.; Katona, T.; Xuei, X.; Liu, Y.; Zhou, H.-M.; Simpson, E.R.; Turner, M.J. Epidermal PPARγ Is a Key Homeostatic Regulator of Cutaneous Inflammation and Barrier Function in Mouse Skin. Int. J. Mol. Sci. 2021, 22, 8634. [Google Scholar] [CrossRef] [PubMed]
- de Guzman Strong, C.; Wertz, P.W.; Wang, C.; Yang, F.; Meltzer, P.S.; Andl, T.; Millar, S.E.; Ho, I.-C.; Pai, S.-Y.; Segre, J.A. Lipid defect underlies selective skin barrier impairment of an epidermal-specific deletion of Gata-3. J. Cell Biol. 2006, 175, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Pappas, A. Epidermal surface lipids. Dermatoendocrinology 2009, 1, 72–76. [Google Scholar] [CrossRef] [Green Version]
- Furue, K.; Mitoma, C.; Tsuji, G.; Furue, M. Protective role of peroxisome proliferator-activated receptor α agonists in skin barrier and inflammation. Immunobiology 2018, 223, 327–330. [Google Scholar] [CrossRef]
- Zákány, N.; Oláh, A.; Markovics, A.; Takács, E.; Aranyász, A.; Nicolussi, S.; Piscitelli, F.; Allarà, M.; Pór, Á.; Kovács, I.; et al. Endocannabinoid Tone Regulates Human Sebocyte Biology. J. Investig. Dermatol. 2018, 138, 1699–1706. [Google Scholar] [CrossRef] [Green Version]
- Tóth, B.I.; Oláh, A.; Szöllosi, A.G.; Czifra, G.; Bíró, T. “Sebocytes’ makeup”: Novel mechanisms and concepts in the physiology of the human sebaceous glands. Pflugers Arch. 2011, 461, 593–606. [Google Scholar] [CrossRef]
- Trivedi, N.R.; Cong, Z.; Nelson, A.M.; Albert, A.J.; Rosamilia, L.L.; Sivarajah, S.; Gilliland, K.L.; Liu, W.; Mauger, D.T.; Gabbay, R.A.; et al. Peroxisome proliferator-activated receptors increase human sebum production. J. Investig. Dermatol. 2006, 126, 2002–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, M.; Zouboulis, C.C.; Ochsendorf, F.; Müller, J.; Thaçi, D.; Bernd, A.; Kaufmann, R.; Kippenberger, S. Peroxisome proliferator-activated receptor activators protect sebocytes from apoptosis: A new treatment modality for acne? Br. J. Dermatol. 2011, 164, 182–186. [Google Scholar] [CrossRef]
- Sardella, C.; Winkler, C.; Quignodon, L.; Hardman, J.A.; Toffoli, B.; Giordano Attianese, G.M.P.; Hundt, J.E.; Michalik, L.; Vinson, C.R.; Paus, R.; et al. Delayed Hair Follicle Morphogenesis and Hair Follicle Dystrophy in a Lipoatrophy Mouse Model of Pparg Total Deletion. J. Investig. Dermatol. 2018, 138, 500–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szántó, M.; Oláh, A.; Szöllősi, A.G.; Tóth, K.F.; Páyer, E.; Czakó, N.; Pór, Á.; Kovács, I.; Zouboulis, C.C.; Kemény, L.; et al. Activation of TRPV3 Inhibits Lipogenesis and Stimulates Production of Inflammatory Mediators in Human Sebocytes-A Putative Contributor to Dry Skin Dermatoses. J. Investig. Dermatol. 2019, 139, 250–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szöllősi, A.G.; Vasas, N.; Angyal, Á.; Kistamás, K.; Nánási, P.P.; Mihály, J.; Béke, G.; Herczeg-Lisztes, E.; Szegedi, A.; Kawada, N.; et al. Activation of TRPV3 Regulates Inflammatory Actions of Human Epidermal Keratinocytes. J. Investig. Dermatol. 2018, 138, 365–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rietcheck, H.R.; Maghfour, J.; Rundle, C.W.; Husayn, S.S.; Presley, C.L.; Sillau, S.H.; Liu, Y.; Leehey, M.A.; Dunnick, C.A.; Dellavalle, R.P. A Review of the Current Evidence Connecting Seborrheic Dermatitis and Parkinson’s Disease and the Potential Role of Oral Cannabinoids. Dermatology 2020, 237, 872–877. [Google Scholar] [CrossRef]
- Sugawara, K.; Zákány, N.; Tiede, S.; Purba, T.; Harries, M.; Tsuruta, D.; Bíró, T.; Paus, R. Human epithelial stem cell survival within their niche requires “tonic” cannabinoid receptor 1-signalling-Lessons from the hair follicle. Exp. Dermatol. 2021, 30, 479–493. [Google Scholar] [CrossRef]
- Karnik, P.; Tekeste, Z.; McCormick, T.S.; Gilliam, A.C.; Price, V.H.; Cooper, K.D.; Mirmirani, P. Hair follicle stem cell-specific PPARγ deletion causes scarring alopecia. J. Investig. Dermatol. 2009, 129, 1243–1257. [Google Scholar] [CrossRef] [Green Version]
- Heiland, I.; Erdmann, R. Biogenesis of peroxisomes. Topogenesis of the peroxisomal membrane and matrix proteins. FEBS J. 2005, 272, 2362–2372. [Google Scholar] [CrossRef]
- Zheng, D.; Bode, A.M.; Zhao, Q.; Cho, Y.-Y.; Zhu, F.; Ma, W.-Y.; Dong, Z. The cannabinoid receptors are required for ultraviolet-induced inflammation and skin cancer development. Cancer Res. 2008, 68, 3992–3998. [Google Scholar] [CrossRef] [Green Version]
- Atalay, S.; Gęgotek, A.; Wroński, A.; Domigues, P.; Skrzydlewska, E. Therapeutic application of cannabidiol on UVA and UVB irradiated rat skin. A proteomic study. J. Pharm. Biomed. Anal. 2021, 192, 113656. [Google Scholar] [CrossRef]
- Jastrząb, A.; Gęgotek, A.; Skrzydlewska, E. Cannabidiol Regulates the Expression of Keratinocyte Proteins Involved in the Inflammation Process through Transcriptional Regulation. Cells 2019, 8, 827. [Google Scholar] [CrossRef] [Green Version]
- Perez, E.; Fernandez, J.R.; Fitzgerald, C.; Rouzard, K.; Tamura, M.; Savile, C. In Vitro and Clinical Evaluation of Cannabigerol (CBG) Produced via Yeast Biosynthesis: A Cannabinoid with a Broad Range of Anti-Inflammatory and Skin Health-Boosting Properties. Molecules 2022, 27, 491. [Google Scholar] [CrossRef] [PubMed]
- Arlen, P.A.; Wu, X. The expression of cannabinoid-related genes in multiple disease cell lines. Appl. Cell Biol. 2020, 8, 11–20. [Google Scholar]
- Yamanaka-Takaichi, M.; Sugawara, K.; Sumitomo, R.; Tsuruta, D. The Mast Cell-SCF-CB1 Interaction Is a Key Player in Seborrheic Keratosis. J. Histochem. Cytochem. 2020, 68, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, J.V.; Miot, H.A. Actinic keratosis: A clinical and epidemiological revision. An. Bras. Dermatol. 2012, 87, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, K.; Bíró, T.; Tsuruta, D.; Tóth, B.I.; Kromminga, A.; Zákány, N.; Zimmer, A.; Funk, W.; Gibbs, B.F.; Zimmer, A.; et al. Endocannabinoids limit excessive mast cell maturation and activation in human skin. J. Allergy Clin. Immunol. 2012, 129, 726–738.e8. [Google Scholar] [CrossRef]
- Piquero-Casals, J.; Morgado-Carrasco, D.; Gilaberte, Y.; Del Rio, R.; Macaya-Pascual, A.; Granger, C.; López-Estebaranz, J.L. Management Pearls on the Treatment of Actinic Keratoses and Field Cancerization. Dermatol. Ther. 2020, 10, 903–915. [Google Scholar] [CrossRef]
- Ramot, Y.; Paus, R. Harnessing neuroendocrine controls of keratin expression: A new therapeutic strategy for skin diseases? Bioessays 2014, 36, 672–686. [Google Scholar] [CrossRef]
- Casares, L.; García, V.; Garrido-Rodríguez, M.; Millán, E.; Collado, J.A.; García-Martín, A.; Peñarando, J.; Calzado, M.A.; de La Vega, L.; Muñoz, E. Cannabidiol induces antioxidant pathways in keratinocytes by targeting BACH1. Redox Biol. 2020, 28, 101321. [Google Scholar] [CrossRef]
- Pucci, M.; Rapino, C.; Di Francesco, A.; Dainese, E.; D’Addario, C.; Maccarrone, M. Epigenetic control of skin differentiation genes by phytocannabinoids. Br. J. Pharmacol. 2013, 170, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Paradisi, A.; Pasquariello, N.; Barcaroli, D.; Maccarrone, M. Anandamide regulates keratinocyte differentiation by inducing DNA methylation in a CB1 receptor-dependent manner. J. Biol. Chem. 2008, 283, 6005–6012. [Google Scholar] [CrossRef] [Green Version]
- Maccarrone, M.; Di Rienzo, M.; Battista, N.; Gasperi, V.; Guerrieri, P.; Rossi, A.; Finazzi-Agrò, A. The endocannabinoid system in human keratinocytes. Evidence that anandamide inhibits epidermal differentiation through CB1 receptor-dependent inhibition of protein kinase C, activation protein-1, and transglutaminase. J. Biol. Chem. 2003, 278, 33896–33903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilkei-Gorzo, A.; Drews, E.; Albayram, Ö.; Piyanova, A.; Gaffal, E.; Tueting, T.; Michel, K.; Mauer, D.; Maier, W.; Zimmer, A. Early onset of aging-like changes is restricted to cognitive abilities and skin structure in Cnr1⁻/⁻ mice. Neurobiol. Aging 2012, 33, 200.e11–200.e22. [Google Scholar] [CrossRef] [PubMed]
- Leal, E.C.; Moura, L.I.F.; Pirzgalska, R.M.; Marques-da-Silva, D.; Ledent, C.; Köfalvi, A.; Carvalho, E. Diabetes and Cannabinoid CB1 receptor deficiency promote similar early onset aging-like changes in the skin. Exp. Gerontol. 2021, 154, 111528. [Google Scholar] [CrossRef] [PubMed]
- Oláh, A.; Alam, M.; Chéret, J.; Kis, N.G.; Hegyi, Z.; Szöllősi, A.G.; Vidali, S.; Bíró, T.; Paus, R. Mitochondrial energy metabolism is negatively regulated by cannabinoid receptor 1 in intact human epidermis. Exp. Dermatol. 2020, 29, 616–622. [Google Scholar] [CrossRef]
- Zagórska-Dziok, M.; Bujak, T.; Ziemlewska, A.; Nizioł-Łukaszewska, Z. Positive Effect of Cannabis sativa L. Herb Extracts on Skin Cells and Assessment of Cannabinoid-Based Hydrogels Properties. Molecules 2021, 26, 802. [Google Scholar] [CrossRef]
- Zawatsky, C.N.; Park, J.K.; Abdalla, J.; Kunos, G.; Iyer, M.R.; Cinar, R. Peripheral Hybrid CB1R and iNOS Antagonist MRI-1867 Displays Anti-Fibrotic Efficacy in Bleomycin-Induced Skin Fibrosis. Front. Endocrinol. 2021, 12, 744857. [Google Scholar] [CrossRef]
- Oka, S.; Yanagimoto, S.; Ikeda, S.; Gokoh, M.; Kishimoto, S.; Waku, K.; Ishima, Y.; Sugiura, T. Evidence for the involvement of the cannabinoid CB2 receptor and its endogenous ligand 2-arachidonoylglycerol in 12-O-tetradecanoylphorbol-13-acetate-induced acute inflammation in mouse ear. J. Biol. Chem. 2005, 280, 18488–18497. [Google Scholar] [CrossRef] [Green Version]
- Oka, S.; Wakui, J.; Ikeda, S.; Yanagimoto, S.; Kishimoto, S.; Gokoh, M.; Nasui, M.; Sugiura, T. Involvement of the cannabinoid CB2 receptor and its endogenous ligand 2-arachidonoylglycerol in oxazolone-induced contact dermatitis in mice. J. Immunol. 2006, 177, 8796–8805. [Google Scholar] [CrossRef] [Green Version]
- Ambrożewicz, E.; Wójcik, P.; Wroński, A.; Łuczaj, W.; Jastrząb, A.; Žarković, N.; Skrzydlewska, E. Pathophysiological Alterations of Redox Signaling and Endocannabinoid System in Granulocytes and Plasma of Psoriatic Patients. Cells 2018, 7, 159. [Google Scholar] [CrossRef] [Green Version]
- Nattkemper, L.A.; Tey, H.L.; Valdes-Rodriguez, R.; Lee, H.; Mollanazar, N.K.; Albornoz, C.; Sanders, K.M.; Yosipovitch, G. The Genetics of Chronic Itch: Gene Expression in the Skin of Patients with Atopic Dermatitis and Psoriasis with Severe Itch. J. Investig. Dermatol. 2018, 138, 1311–1317. [Google Scholar] [CrossRef] [Green Version]
- Campora, L.; Miragliotta, V.; Ricci, E.; Cristino, L.; Di Marzo, V.; Albanese, F.; Della Federica Valle, M.; Abramo, F. Cannabinoid receptor type 1 and 2 expression in the skin of healthy dogs and dogs with atopic dermatitis. Am. J. Vet. Res. 2012, 73, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.-L.; Yu, T.-S.; Li, X.-N.; Fan, Y.-Y.; Ma, W.-X.; Du, Y.; Zhao, R.; Guan, D.-W. Cannabinoid receptor type 2 is time-dependently expressed during skin wound healing in mice. Int. J. Legal Med. 2012, 126, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Correia-Sá, I.B.; Carvalho, C.M.; Serrão, P.V.; Machado, V.A.; Carvalho, S.O.; Marques, M.; Vieira-Coelho, M.A. AM251, a cannabinoid receptor 1 antagonist, prevents human fibroblasts differentiation and collagen deposition induced by TGF-β—An in vitro study. Eur. J. Pharmacol. 2021, 892, 173738. [Google Scholar] [CrossRef] [PubMed]
- Correia-Sá, I.; Carvalho, C.; Machado, V.A.; Carvalho, S.; Serrão, P.; Marques, M.; Vieira-Coelho, M.A. Targeting cannabinoid receptor 2 (CB2) limits collagen production-An in vitro study in a primary culture of human fibroblasts. Fundam. Clin. Pharmacol. 2022, 36, 89–99. [Google Scholar] [CrossRef]
- Li, L.; Liu, X.; Ge, W.; Chen, C.; Huang, Y.; Jin, Z.; Zhan, M.; Duan, X.; Liu, X.; Kong, Y.; et al. CB2R Deficiency Exacerbates Imiquimod-Induced Psoriasiform Dermatitis and Itch Through the Neuro-Immune Pathway. Front. Pharmacol. 2022, 13, 790712. [Google Scholar] [CrossRef]
- Sulk, M.; Seeliger, S.; Aubert, J.; Schwab, V.D.; Cevikbas, F.; Rivier, M.; Nowak, P.; Voegel, J.J.; Buddenkotte, J.; Steinhoff, M. Distribution and expression of non-neuronal transient receptor potential (TRPV) ion channels in rosacea. J. Investig. Dermatol. 2012, 132, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Feng, J.; Yu, G.; Yang, P.; Mack, M.R.; Du, J.; Yu, W.; Qian, A.; Zhang, Y.; Liu, S.; et al. Transient receptor potential vanilloid 4-expressing macrophages and keratinocytes contribute differentially to allergic and nonallergic chronic itch. J. Allergy Clin. Immunol. 2018, 141, 608–619.e7. [Google Scholar] [CrossRef] [Green Version]
- Nijsten, T.; Geluyckens, E.; Colpaert, C.; Lambert, J. Peroxisome proliferator-activated receptors in squamous cell carcinoma and its precursors. J. Cutan. Pathol. 2005, 32, 340–347. [Google Scholar] [CrossRef]
- Staumont-Sallé, D.; Abboud, G.; Brénuchon, C.; Kanda, A.; Roumier, T.; Lavogiez, C.; Fleury, S.; Rémy, P.; Papin, J.-P.; Bertrand-Michel, J.; et al. Peroxisome proliferator-activated receptor alpha regulates skin inflammation and humoral response in atopic dermatitis. J. Allergy Clin. Immunol. 2008, 121, 962–968.e6. [Google Scholar] [CrossRef]
- Lee, D.J.; Lee, J.; Ha, J.; Park, K.-C.; Ortonne, J.-P.; Kang, H.Y. Defective barrier function in melasma skin. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 1533–1537. [Google Scholar] [CrossRef]
- Ruzehaji, N.; Frantz, C.; Ponsoye, M.; Avouac, J.; Pezet, S.; Guilbert, T.; Luccarini, J.-M.; Broqua, P.; Junien, J.-L.; Allanore, Y. Pan PPAR agonist IVA337 is effective in prevention and treatment of experimental skin fibrosis. Ann. Rheum. Dis. 2016, 75, 2175–2183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.J.; Jin, X.-J.; Kim, Y.K.; Oh, I.K.; Kim, J.E.; Park, C.-H.; Chung, J.H. UV decreases the synthesis of free fatty acids and triglycerides in the epidermis of human skin in vivo, contributing to development of skin photoaging. J. Dermatol. Sci. 2010, 57, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Mahgoub, D.; El Tawdy, A.M.; Metwally, D.; Manar, A.; Rashed, L. Estimation of peroxisome proliferators—Activated receptor γ gene expression in inflammatory skin diseases: Atopic dermatitis and psoriasis. Our Dermatol. Online 2014, 5, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Luo, S.; Zhan, Y.; Wang, J.; Zhao, R.; Li, Y.; Zeng, J.; Lu, Q. Increased Expression of PPAR-γ Modulates Monocytes Into a M2-Like Phenotype in SLE Patients: An Implicative Protective Mechanism and Potential Therapeutic Strategy of Systemic Lupus Erythematosus. Front. Immunol. 2020, 11, 579372. [Google Scholar] [CrossRef] [PubMed]
- Westergaard, M.; Henningsen, J.; Johansen, C.; Rasmussen, S.; Svendsen, M.L.; Jensen, U.B.; Schrøder, H.D.; Staels, B.; Iversen, L.; Bolund, L.; et al. Expression and localization of peroxisome proliferator-activated receptors and nuclear factor κB in normal and lesional psoriatic skin. J. Investig. Dermatol. 2003, 121, 1104–1117. [Google Scholar] [CrossRef] [PubMed]
- Babes, A.; Kichko, T.I.; Selescu, T.; Manolache, A.; Neacsu, C.; Gebhardt, L.; Reeh, P.W. Psoralens activate and photosensitize Transient Receptor Potential channels Ankyrin type 1 (TRPA1) and Vanilloid type 1 (TRPV1). Eur. J. Pain 2021, 25, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Garbutcheon-Singh, K.B.; Smith, S.D. Cannabinoids interaction with transient receptor potential family and implications in the treatment of rosacea. Dermatol. Ther. 2021, 34, e15162. [Google Scholar] [CrossRef]
- Yun, J.-W.; Seo, J.A.; Jang, W.-H.; Koh, H.J.; Bae, I.-H.; Park, Y.-H.; Lim, K.-M. Antipruritic effects of TRPV1 antagonist in murine atopic dermatitis and itching models. J. Investig. Dermatol. 2011, 131, 1576–1579. [Google Scholar] [CrossRef] [Green Version]
- Denda, M.; Sokabe, T.; Fukumi-Tominaga, T.; Tominaga, M. Effects of skin surface temperature on epidermal permeability barrier homeostasis. J. Investig. Dermatol. 2007, 127, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.-J.; Shim, W.-S. Cutaneous Neuroimmune Interactions of TSLP and TRPV4 Play Pivotal Roles in Dry Skin-Induced Pruritus. Front. Immunol. 2021, 12, 772941. [Google Scholar] [CrossRef]
- Do, B.H.; Koizumi, H.; Ohbuchi, T.; Kawaguchi, R.; Suzuki, H. Expressions of TRPVs in the cholesteatoma epithelium. Acta Otolaryngol. 2017, 137, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Motwani, M.P.; Colas, R.A.; George, M.J.; Flint, J.D.; Dalli, J.; Richard-Loendt, A.; de Maeyer, R.P.; Serhan, C.N.; Gilroy, D.W. Pro-resolving mediators promote resolution in a human skin model of UV-killed Escherichia coli-driven acute inflammation. JCI Insight 2018, 3, e94463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amr, K.; Abdel-Hameed, M.; Sayed, K.; Nour-Edin, F.; Abdel Hay, R. The Pro12Ala polymorphism of the gene for peroxisome proliferator activated receptor-gamma is associated with a lower Global Acne Grading System score in patients with acne vulgaris. Clin. Exp. Dermatol. 2014, 39, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Zhang, Y.L.; Han, D.H.; Kim, D.Y.; Rhee, C.S. Antiallergic Function of KR62980, a Peroxisome Proliferator-Activated Receptor-γ Agonist, in a Mouse Allergic Rhinitis Model. Allergy Asthma Immunol. Res. 2015, 7, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Angelina, A.; Pérez-Diego, M.; López-Abente, J.; Palomares, O. The Role of Cannabinoids in Allergic Diseases: Collegium Internationale Allergologicum (CIA) Update 2020. Int. Arch. Allergy Immunol. 2020, 181, 565–584. [Google Scholar] [CrossRef]
- Russo, E.B. History of cannabis and its preparations in saga, science, and sobriquet. Chem. Biodivers. 2007, 4, 1614–1648. [Google Scholar] [CrossRef]
- Russo, E.B. The Pharmacological History of Cannabis. In Handbook of Cannabis, 1st ed.; Pertwee, R.G., Ed.; Oxford University Press: Oxford, UK, 2014; pp. 23–43. ISBN 9780199662685. [Google Scholar]
- Lemery, N. Dictionaire ou Traite Universel Des Drogues Simples; Jean Hofhout: Rotterdam, The Netherlands, 1727. [Google Scholar]
- Johanek, L.M.; Simone, D.A. Activation of peripheral cannabinoid receptors attenuates cutaneous hyperalgesia produced by a heat injury. Pain 2004, 109, 432–442. [Google Scholar] [CrossRef]
- Sofia, R.D.; Nalepa, S.D.; Harakal, J.J.; Vassar, H.B. Anti-edema and analgesic properties of Δ9-tetrahydrocannabinol (THC). J. Pharmacol. Exp. Ther. 1973, 186, 646–655. [Google Scholar]
- Duane Sofia, R.; Nalepa, S.D.; Vassar, H.B.; Knobloch, L.C. Comparative anti-phlogistic activity of Δ9-tetrahydrocannabinol, hydrocortisone and aspirin in various rat paw edema models. Life Sci. 1974, 15, 251–260. [Google Scholar] [CrossRef]
- Kehl, L.J.; Hamamoto, D.T.; Wacnik, P.W.; Croft, D.L.; Norsted, B.D.; Wilcox, G.L.; Simone, D.A. A cannabinoid agonist differentially attenuates deep tissue hyperalgesia in animal models of cancer and inflammatory muscle pain. Pain 2003, 103, 175–186. [Google Scholar] [CrossRef]
- Clayton, N.; Marshall, F.H.; Bountra, C.; O’Shaughnessy, C.T. CB1 and CB2 cannabinoid receptors are implicated in inflammatory pain. Pain 2002, 96, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Hanus, L.; Breuer, A.; Tchilibon, S.; Shiloah, S.; Goldenberg, D.; Horowitz, M.; Pertwee, R.G.; Ross, R.A.; Mechoulam, R.; Fride, E. HU-308: A specific agonist for CB2, a peripheral cannabinoid receptor. Proc. Natl. Acad. Sci. USA 1999, 96, 14228–14233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuehl, F.A.; Jacob, T.A.; Ganley, O.H.; Ormond, R.E.; Meisinger, M.A.P. The identification of N-(2-hydroxyethyl)-palmitamide as a naturally occurring anti-inflammatory agent. J. Am. Chem. Soc. 1957, 79, 5577–5578. [Google Scholar] [CrossRef]
- Costa, B.; Conti, S.; Giagnoni, G.; Colleoni, M. Therapeutic effect of the endogenous fatty acid amide, palmitoylethanolamide, in rat acute inflammation: Inhibition of nitric oxide and cyclo-oxygenase systems. Br. J. Pharmacol. 2002, 137, 413–420. [Google Scholar] [CrossRef]
- Benvenuti, F.; Lattanzi, F.; de Gori, A.; Tarli, P. Attivita’ di alcuni derivati della palmitoiletanolamide sull’edema da carragenina nella zampa di ratto [Activity of some derivatives of palmitoylethanolamide on carragenine-induced edema in the rat paw]. Boll. Soc. Ital. Biol. Sper. 1968, 44, 809–813. [Google Scholar]
- Mazzari, S.; Canella, R.; Petrelli, L.; Marcolongo, G.; Leon, A. N-(2-Hydroxyethyl)hexadecanamide is orally active in reducing edema formation and inflammatory hyperalgesia by down-modulating mast cell activation. Eur. J. Pharmacol. 1996, 300, 227–236. [Google Scholar] [CrossRef]
- Marini, I.; Bartolucci, M.L.; Bortolotti, F.; Gatto, M.R.; Bonetti, G.A. Palmitoylethanolamide versus a nonsteroidal anti-inflammatory drug in the treatment of temporomandibular joint inflammatory pain. J. Orofac. Pain 2012, 26, 99–104. [Google Scholar]
- Artukoglu, B.B.; Beyer, C.; Zuloff-Shani, A.; Brener, E.; Bloch, M.H. Efficacy of Palmitoylethanolamide for Pain: A Meta-Analysis. Pain Physician 2017, 20, 353–362. [Google Scholar]
- Robinson, R.H.; Meissler, J.J.; Fan, X.; Yu, D.; Adler, M.W.; Eisenstein, T.K. A CB2-Selective Cannabinoid Suppresses T-Cell Activities and Increases Tregs and IL-10. J. Neuroimmune Pharmacol. 2015, 10, 318–332. [Google Scholar] [CrossRef] [Green Version]
- Ke, P.; Shao, B.-Z.; Xu, Z.-Q.; Wei, W.; Han, B.-Z.; Chen, X.-W.; Su, D.-F.; Liu, C. Activation of Cannabinoid Receptor 2 Ameliorates DSS-Induced Colitis through Inhibiting NLRP3 Inflammasome in Macrophages. PLoS ONE 2016, 11, e0155076. [Google Scholar] [CrossRef] [Green Version]
- Tomar, S.; Zumbrun, E.E.; Nagarkatti, M.; Nagarkatti, P.S. Protective role of cannabinoid receptor 2 activation in galactosamine/lipopolysaccharide-induced acute liver failure through regulation of macrophage polarization and microRNAs. J. Pharmacol. Exp. Ther. 2015, 353, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurier, R.B.; Rossetti, R.G.; Burstein, S.H.; Bidinger, B. Suppression of human monocyte interleukin-1β production by ajulemic acid, a nonpsychoactive cannabinoid. Biochem. Pharmacol. 2003, 65, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, V.A.; Aslan, S.; Safaei, R.; Vaughan, C.W. Effect of the cannabinoid ajulemic acid on rat models of neuropathic and inflammatory pain. Neurosci. Lett. 2005, 382, 231–235. [Google Scholar] [CrossRef]
- Gonzalez, E.G.; Selvi, E.; Balistreri, E.; Akhmetshina, A.; Palumbo, K.; Lorenzini, S.; Lazzerini, P.E.; Montilli, C.; Capecchi, P.L.; Lucattelli, M.; et al. Synthetic cannabinoid ajulemic acid exerts potent antifibrotic effects in experimental models of systemic sclerosis. Ann. Rheum. Dis. 2012, 71, 1545–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, J.; Atez, F.; Rossetti, R.G.; Skulas, A.; Patel, R.; Zurier, R.B. Suppression of human macrophage interleukin-6 by a nonpsychoactive cannabinoid acid. Rheumatol. Int. 2008, 28, 631–635. [Google Scholar] [CrossRef]
- Motwani, M.P.; Bennett, F.; Norris, P.C.; Maini, A.A.; George, M.J.; Newson, J.; Henderson, A.; Hobbs, A.J.; Tepper, M.; White, B.; et al. Potent Anti-Inflammatory and Pro-Resolving Effects of Anabasum in a Human Model of Self-Resolving Acute Inflammation. Clin. Pharmacol. Ther. 2018, 104, 675–686. [Google Scholar] [CrossRef] [Green Version]
- Burstein, S. Molecular Mechanisms for the Inflammation-Resolving Actions of Lenabasum. Mol. Pharmacol. 2021, 99, 125–132. [Google Scholar] [CrossRef]
- Chiang, N.; Dalli, J.; Colas, R.A.; Serhan, C.N. Identification of resolvin D2 receptor mediating resolution of infections and organ protection. J. Exp. Med. 2015, 212, 1203–1217. [Google Scholar] [CrossRef] [Green Version]
- Schlosburg, J.E.; Boger, D.L.; Cravatt, B.F.; Lichtman, A.H. Endocannabinoid modulation of scratching response in an acute allergenic model: A new prospective neural therapeutic target for pruritus. J. Pharmacol. Exp. Ther. 2009, 329, 314–323. [Google Scholar] [CrossRef]
- Gercek, O.Z.; Oflaz, B.; Oguz, N.; Demirci, K.; Gunduz, O.; Ulugol, A. Role of Nitric Oxide in the Antipruritic Effect of WIN 55,212-2, a Cannabinoid Agonist. Basic Clin. Neurosci. 2020, 11, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Odan, M.; Ishizuka, N.; Hiramatsu, Y.; Inagaki, M.; Hashizume, H.; Fujii, Y.; Mitsumori, S.; Morioka, Y.; Soga, M.; Deguchi, M.; et al. Discovery of S-777469: An orally available CB2 agonist as an antipruritic agent. Bioorg. Med. Chem. Lett. 2012, 22, 2803–2806. [Google Scholar] [CrossRef] [PubMed]
- Haruna, T.; Soga, M.; Morioka, Y.; Hikita, I.; Imura, K.; Furue, Y.; Yamamoto, M.; Imura, C.; Ikeda, M.; Yamauchi, A.; et al. S-777469, a novel cannabinoid type 2 receptor agonist, suppresses itch-associated scratching behavior in rodents through inhibition of itch signal transmission. Pharmacology 2015, 95, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Marsella, R.; Ahrens, K.; Sanford, R.; Trujillo, A.; Massre, D.; Soeberdt, M.; Abels, C. Double blinded, vehicle controlled, crossover study on the efficacy of a topical endocannabinoid membrane transporter inhibitor in atopic Beagles. Arch. Dermatol. Res. 2019, 311, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Lucaciu, O.C.; Connell, G.P. Itch sensation through transient receptor potential channels: A systematic review and relevance to manual therapy. J. Manip. Physiol. Ther. 2013, 36, 385–393. [Google Scholar] [CrossRef]
- Sanjel, B.; Kim, B.-H.; Song, M.-H.; Carstens, E.; Shim, W.-S. Glucosylsphingosine evokes pruritus via activation of 5-HT2A receptor and TRPV4 in sensory neurons. Br. J. Pharmacol. 2022, 179, 2193–2207. [Google Scholar] [CrossRef]
- Yan, J.; Ye, F.; Ju, Y.; Wang, D.; Chen, J.; Zhang, X.; Yin, Z.; Wang, C.; Yang, Y.; Zhu, C.; et al. Cimifugin relieves pruritus in psoriasis by inhibiting TRPV4. Cell Calcium 2021, 97, 102429. [Google Scholar] [CrossRef]
- Szabó, I.L.; Lisztes, E.; Béke, G.; Tóth, K.F.; Paus, R.; Oláh, A.; Bíró, T. The Phytocannabinoid (-)-Cannabidiol Operates as a Complex, Differential Modulator of Human Hair Growth: Anti-Inflammatory Submicromolar versus Hair Growth Inhibitory Micromolar Effects. J. Investig. Dermatol. 2020, 140, 484–488.e5. [Google Scholar] [CrossRef]
- Küster, W.; Happle, R. The inheritance of common baldness: Two B or not two B? J. Am. Acad. Dermatol. 1984, 11, 921–926. [Google Scholar] [CrossRef]
- Thiers, B.H.; Dobson, R.L. (Eds.) Pathogenesis of Skin Disease; Churchill Livingstone: New York, NY, USA, 1986. [Google Scholar]
- Hamilton, J.B. Male hormone stimulation is prerequisite and an incitant in common baldness. Am. J. Anat. 1942, 71, 451–480. [Google Scholar] [CrossRef]
- Hamilton, J.B. Patterned loss of hair in man; types and incidence. Ann. N. Y. Acad. Sci. 1951, 53, 708–728. [Google Scholar] [CrossRef]
- Gupta, A.K.; Mays, R.R.; Dotzert, M.S.; Versteeg, S.G.; Shear, N.H.; Piguet, V. Efficacy of non-surgical treatments for androgenetic alopecia: A systematic review and network meta-analysis. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 2112–2125. [Google Scholar] [CrossRef]
- Motofei, I.G.; Rowland, D.L.; Baconi, D.L.; Tampa, M.; Sârbu, M.-I.; Păunică, S.; Constantin, V.D.; Bălălău, C.; Păunică, I.; Georgescu, S.R. Androgenetic alopecia; drug safety and therapeutic strategies. Expert Opin. Drug Saf. 2018, 17, 407–412. [Google Scholar] [CrossRef]
- Gentile, P.; Garcovich, S. Systematic Review of Platelet-Rich Plasma Use in Androgenetic Alopecia Compared with Minoxidil®, Finasteride®, and Adult Stem Cell-Based Therapy. Int. J. Mol. Sci. 2020, 21, 2702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiting, D.A. Diagnostic and predictive value of horizontal sections of scalp biopsy specimens in male pattern androgenetic alopecia. J. Am. Acad. Dermatol. 1993, 28, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Nirmal, B.; Somiah, S.; Sacchidanand, S.A.; Biligi, D.S.; Palo, S. Evaluation of Perifollicular Inflammation of Donor Area during Hair Transplantation in Androgenetic Alopecia and its Comparison with Controls. Int. J. Trichology 2013, 5, 73–76. [Google Scholar] [PubMed]
- Gupta, A.K.; Talukder, M. Cannabinoids for skin diseases and hair regrowth. J. Cosmet. Dermatol. 2021, 20, 2703–2711. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-J.; Ryu, J.-M.; Na, H.-H.; Jung, H.-S.; Kim, B.; Park, J.-S.; Ahn, B.-S.; Kim, K.-C. Regulatory Effect of Cannabidiol (CBD) on Decreased β-Catenin Expression in Alopecia Models by Testosterone and PMA Treatment in Dermal Papilla Cells. J. Pharmacopunct. 2021, 24, 68–75. [Google Scholar] [CrossRef]
- Srivastava, B.K.; Soni, R.; Patel, J.Z.; Joharapurkar, A.; Sadhwani, N.; Kshirsagar, S.; Mishra, B.; Takale, V.; Gupta, S.; Pandya, P.; et al. Hair growth stimulator property of thienyl substituted pyrazole carboxamide derivatives as a CB1 receptor antagonist with in vivo antiobesity effect. Bioorg. Med. Chem. Lett. 2009, 19, 2546–2550. [Google Scholar] [CrossRef]
- Weidinger, S.; Beck, L.A.; Bieber, T.; Kabashima, K.; Irvine, A.D. Atopic dermatitis. Nat. Rev. Dis. Prim. 2018, 4, 1. [Google Scholar] [CrossRef]
- Tobin, D.; Nabarro, G.; de La Faute, H.B.; van Vloten, W.A.; van der Putte, S.C.; Schuurman, H.-J. Increased number of immunoreactive nerve fibers in atopic dermatitis. J. Allergy Clin. Immunol. 1992, 90, 613–622. [Google Scholar] [CrossRef]
- Kay, A.B. Calcitonin gene-related peptide– and vascular endothelial growth factor-positive inflammatory cells in late-phase allergic skin reactions in atopic subjects. J. Allergy Clin. Immunol. 2011, 127, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Gutzmer, R.; Mommert, S.; Gschwandtner, M.; Zwingmann, K.; Stark, H.; Werfel, T. The histamine H4 receptor is functionally expressed on TH2 cells. J. Allergy Clin. Immunol. 2009, 123, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Chun, P.I.F.; Lehman, H. Current and Future Monoclonal Antibodies in the Treatment of Atopic Dermatitis. Clin. Rev. Allergy Immunol. 2020, 59, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Meggitt, S.J.; Gray, J.C.; Reynolds, N.J. Azathioprine dosed by thiopurine methyltransferase activity for moderate-to-severe atopic eczema: A double-blind, randomised controlled trial. Lancet 2006, 367, 839–846. [Google Scholar] [CrossRef]
- Roekevisch, E.; Schram, M.E.; Leeflang, M.M.G.; Brouwer, M.W.D.; Gerbens, L.A.A.; Bos, J.D.; Spuls, P.I. Methotrexate versus azathioprine in patients with atopic dermatitis: 2-year follow-up data. J. Allergy Clin. Immunol. 2018, 141, 825–827.e10. [Google Scholar] [CrossRef] [Green Version]
- Gerbens, L.A.A.; Hamann, S.A.S.; Brouwer, M.W.D.; Roekevisch, E.; Leeflang, M.M.G.; Spuls, P.I. Methotrexate and azathioprine for severe atopic dermatitis: A 5-year follow-up study of a randomized controlled trial. Br. J. Dermatol. 2018, 178, 1288–1296. [Google Scholar] [CrossRef]
- Hong, C.-H.; Gooderham, M.; Bissonnette, R. Evidence Review of Topical Calcineurin Inhibitors for the Treatment of Adult Atopic Dermatitis. J. Cutan. Med. Surg. 2019, 23, 5S–10S. [Google Scholar] [CrossRef]
- Nogueira, M.; Torres, T. Janus Kinase Inhibitors for the Treatment of Atopic Dermatitis: Focus on Abrocitinib, Baricitinib, and Upadacitinib. Dermatol. Pract. Concept. 2021, 11, e2021145. [Google Scholar] [CrossRef]
- Chiricozzi, A.; Gori, N.; Maurelli, M.; Gisondi, P.; Caldarola, G.; De Simone, C.; Peris, K.; Girolomoni, G. Biological agents targeting interleukin-13 for atopic dermatitis. Expert Opin. Biol. Ther. 2022, 22, 651–659. [Google Scholar] [CrossRef]
- Sidbury, R.; Alpizar, S.; Laquer, V.; Dhawan, S.; Abramovits, W.; Loprete, L.; Krishnaswamy, J.K.; Ahmad, F.; Jabbar-Lopez, Z.; Piketty, C. Pharmacokinetics, Safety, Efficacy, and Biomarker Profiles During Nemolizumab Treatment of Atopic Dermatitis in Adolescents. Dermatol. Ther. 2022, 12, 631–642. [Google Scholar] [CrossRef]
- Schneider, S.; Li, L.; Zink, A. The New Era of Biologics in Atopic Dermatitis: A Review. Dermatol. Pract. Concept. 2021, 11, e2021144. [Google Scholar] [CrossRef] [PubMed]
- Paller, A.S.; Tom, W.L.; Lebwohl, M.G.; Blumenthal, R.L.; Boguniewicz, M.; Call, R.S.; Eichenfield, L.F.; Forsha, D.W.; Rees, W.C.; Simpson, E.L.; et al. Efficacy and safety of crisaborole ointment, a novel, nonsteroidal phosphodiesterase 4 (PDE4) inhibitor for the topical treatment of atopic dermatitis (AD) in children and adults. J. Am. Acad. Dermatol. 2016, 75, 494–503.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thom, H.; Cheng, V.; Keeney, E.; Neary, M.P.; Eccleston, A.; Zang, C.; Cappelleri, J.C.; Cha, A.; Thyssen, J.P. Matching-Adjusted Indirect Comparison of Crisaborole Ointment 2% vs. Topical Calcineurin Inhibitors in the Treatment of Patients with Mild-to-Moderate Atopic Dermatitis. Dermatol. Ther. 2022, 12, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Blauvelt, A.; Teixeira, H.D.; Simpson, E.L.; Costanzo, A.; de Bruin-Weller, M.; Barbarot, S.; Prajapati, V.H.; Lio, P.; Hu, X.; Wu, T.; et al. Efficacy and Safety of Upadacitinib vs Dupilumab in Adults with Moderate-to-Severe Atopic Dermatitis: A Randomized Clinical Trial. JAMA Dermatol. 2021, 157, 1047–1055. [Google Scholar] [CrossRef]
- Dhillon, S. Delgocitinib: First Approval. Drugs 2020, 80, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.; Molin, S. Delgocitinib in atopic dermatitis. Drugs Today 2021, 57, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Sher, L.G.; Chang, J.; Patel, I.B.; Balkrishnan, R.; Fleischer, A.B. Relieving the pruritus of atopic dermatitis: A meta-analysis. Acta Derm. Venereol. 2012, 92, 455–461. [Google Scholar] [CrossRef]
- Gaffal, E.; Glodde, N.; Jakobs, M.; Bald, T.; Tüting, T. Cannabinoid 1 receptors in keratinocytes attenuate fluorescein isothiocyanate-induced mouse atopic-like dermatitis. Exp. Dermatol. 2014, 23, 401–406. [Google Scholar] [CrossRef]
- Maekawa, T.; Nojima, H.; Kuraishi, Y.; Aisaka, K. The cannabinoid CB2 receptor inverse agonist JTE-907 suppresses spontaneous itch-associated responses of NC mice, a model of atopic dermatitis. Eur. J. Pharmacol. 2006, 542, 179–183. [Google Scholar] [CrossRef]
- Engel, M.A.; Izydorczyk, I.; Mueller-Tribbensee, S.M.; Becker, C.; Neurath, M.F.; Reeh, P.W. Inhibitory CB1 and activating/desensitizing TRPV1-mediated cannabinoid actions on CGRP release in rodent skin. Neuropeptides 2011, 45, 229–237. [Google Scholar] [CrossRef]
- Nam, G.; Jeong, S.K.; Park, B.M.; Lee, S.H.; Kim, H.J.; Hong, S.-P.; Kim, B.; Kim, B.-W. Selective Cannabinoid Receptor-1 Agonists Regulate Mast Cell Activation in an Oxazolone-Induced Atopic Dermatitis Model. Ann. Dermatol. 2016, 28, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loewinger, M.; Wakshlag, J.J.; Bowden, D.; Peters-Kennedy, J.; Rosenberg, A. The effect of a mixed cannabidiol and cannabidiolic acid based oil on client-owned dogs with atopic dermatitis. Vet. Dermatol. 2022, 33, 329-e77. [Google Scholar] [CrossRef] [PubMed]
- Jakasa, I.; Thyssen, J.P.; Kezic, S. The role of skin barrier in occupational contact dermatitis. Exp. Dermatol. 2018, 27, 909–914. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, L. Contact Dermatitis: Classifications and Management. Clin. Rev. Allergy Immunol. 2021, 61, 245–281. [Google Scholar] [CrossRef] [PubMed]
- Hamann, C.R.; Hamann, D.; Egeberg, A.; Johansen, J.D.; Silverberg, J.; Thyssen, J.P. Association between atopic dermatitis and contact sensitization: A systematic review and meta-analysis. J. Am. Acad. Dermatol. 2017, 77, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Smit, H.A.; Burdorf, A.; Coenraads, P.J. Prevalence of hand dermatitis in different occupations. Int. J. Epidemiol. 1993, 22, 288–293. [Google Scholar] [CrossRef]
- Ruzicka, T.; Larsen, F.G.; Galewicz, D.; Horváth, A.; Coenraads, P.J.; Thestrup-Pedersen, K.; Ortonne, J.P.; Zouboulis, C.C.; Harsch, M.; Brown, T.C.; et al. Oral alitretinoin (9-cis-retinoic acid) therapy for chronic hand dermatitis in patients refractory to standard therapy: Results of a randomized, double-blind, placebo-controlled, multicenter trial. Arch. Dermatol. 2004, 140, 1453–1459. [Google Scholar] [CrossRef] [Green Version]
- Murphy, G.M.; Maurice, P.D.; Norris, P.G.; Morris, R.W.; Hawk, J.L. Azathioprine treatment in chronic actinic dermatitis: A double-blind controlled trial with monitoring of exposure to ultraviolet radiation. Br. J. Dermatol. 1989, 121, 639–646. [Google Scholar] [CrossRef]
- Petrosino, S.; Cristino, L.; Karsak, M.; Gaffal, E.; Ueda, N.; Tüting, T.; Bisogno, T.; de Filippis, D.; D’Amico, A.; Saturnino, C.; et al. Protective role of palmitoylethanolamide in contact allergic dermatitis. Allergy 2010, 65, 698–711. [Google Scholar] [CrossRef]
- Petrosino, S.; Verde, R.; Vaia, M.; Allarà, M.; Iuvone, T.; Di Marzo, V. Anti-inflammatory Properties of Cannabidiol, a Nonpsychotropic Cannabinoid, in Experimental Allergic Contact Dermatitis. J. Pharmacol. Exp. Ther. 2018, 365, 652–663. [Google Scholar] [CrossRef]
- Vaia, M.; Petrosino, S.; de Filippis, D.; Negro, L.; Guarino, A.; Carnuccio, R.; Di Marzo, V.; Iuvone, T. Palmitoylethanolamide reduces inflammation and itch in a mouse model of contact allergic dermatitis. Eur. J. Pharmacol. 2016, 791, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Kircik, L. A nonsteroidal lamellar matrix cream containing palmitoylethanolamide for the treatment of atopic dermatitis. J. Drugs Dermatol. 2010, 9, 334–338. [Google Scholar] [PubMed]
- Mugnaini, C.; Rabbito, A.; Brizzi, A.; Palombi, N.; Petrosino, S.; Verde, R.; Di Marzo, V.; Ligresti, A.; Corelli, F. Synthesis of novel 2-(1-adamantanylcarboxamido)thiophene derivatives. Selective cannabinoid type 2 (CB2) receptor agonists as potential agents for the treatment of skin inflammatory disease. Eur. J. Med. Chem. 2019, 161, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Iwamura, H.; Suzuki, H.; Ueda, Y.; Kaya, T.; Inaba, T. In vitro and in vivo pharmacological characterization of JTE-907, a novel selective ligand for cannabinoid CB2 receptor. J. Pharmacol. Exp. Ther. 2001, 296, 420–425. [Google Scholar] [PubMed]
- Ueda, Y.; Miyagawa, N.; Wakitani, K. Involvement of cannabinoid CB2 receptors in the IgE-mediated triphasic cutaneous reaction in mice. Life Sci. 2007, 80, 414–419. [Google Scholar] [CrossRef]
- Ueda, Y.; Miyagawa, N.; Matsui, T.; Kaya, T.; Iwamura, H. Involvement of cannabinoid CB2 receptor-mediated response and efficacy of cannabinoid CB2 receptor inverse agonist, JTE-907, in cutaneous inflammation in mice. Eur. J. Pharmacol. 2005, 520, 164–171. [Google Scholar] [CrossRef]
- Sheu, M.Y.; Fowler, A.J.; Kao, J.; Schmuth, M.; Schoonjans, K.; Auwerx, J.; Fluhr, J.W.; Man, M.-Q.; Elias, P.M.; Feingold, K.R. Topical peroxisome proliferator activated receptor-α activators reduce inflammation in irritant and allergic contact dermatitis models. J. Investig. Dermatol. 2002, 118, 94–101. [Google Scholar] [CrossRef]
- Sasso, O.; Summa, M.; Armirotti, A.; Pontis, S.; de Mei, C.; Piomelli, D. The N-Acylethanolamine Acid Amidase Inhibitor ARN077 Suppresses Inflammation and Pruritus in a Mouse Model of Allergic Dermatitis. J. Investig. Dermatol. 2018, 138, 562–569. [Google Scholar] [CrossRef] [Green Version]
- Wen, S.; Ye, L.; Liu, D.; Yang, B.; Man, M.-Q. Topical N-palmitoyl serinol, a commensal bacterial metabolite, prevents the development of epidermal permeability barrier dysfunction in a murine model of atopic dermatitis-like skin. Can. J. Vet. Res. 2021, 85, 201–204. [Google Scholar]
- Timis, T.-L.; Florian, I.-A.; Vesa, S.-C.; Mitrea, D.R.; Orasan, R.-I. An updated guide in the management of psoriasis for every practitioner. Int. J. Clin. Pract. 2021, 75, e14290. [Google Scholar] [CrossRef]
- Ogawa, E.; Sato, Y.; Minagawa, A.; Okuyama, R. Pathogenesis of psoriasis and development of treatment. J. Dermatol. 2018, 45, 264–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, P.; Liu, T.; Zhou, W.-Y.; Zhuang, Y.; Peng, L.; Zhang, J.-Y.; Yin, Z.-N.; Mao, X.; Guo, G.; Shi, Y.; et al. Role of gamma-delta T cells in host response against Staphylococcus aureus-induced pneumonia. BMC Immunol. 2012, 13, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teunissen, M.B.; Koomen, C.W.; de Waal Malefyt, R.; Wierenga, E.A.; Bos, J.D. Interleukin-17 and interferon-gamma synergize in the enhancement of proinflammatory cytokine production by human keratinocytes. J. Investig. Dermatol. 1998, 111, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Elmets, C.A.; Korman, N.J.; Prater, E.F.; Wong, E.B.; Rupani, R.N.; Kivelevitch, D.; Armstrong, A.W.; Connor, C.; Cordoro, K.M.; Davis, D.M.R.; et al. Joint AAD-NPF Guidelines of care for the management and treatment of psoriasis with topical therapy and alternative medicine modalities for psoriasis severity measures. J. Am. Acad. Dermatol. 2021, 84, 432–470. [Google Scholar] [CrossRef] [PubMed]
- Katz, H.I.; Prawer, S.E.; Medansky, R.S.; Krueger, G.G.; Mooney, J.J.; Jones, M.L.; Samson, C.R. Intermittent corticosteroid maintenance treatment of psoriasis: A double-blind multicenter trial of augmented betamethasone dipropionate ointment in a pulse dose treatment regimen. Dermatologica 1991, 183, 269–274. [Google Scholar] [CrossRef]
- Menter, A.; Korman, N.J.; Elmets, C.A.; Feldman, S.R.; Gelfand, J.M.; Gordon, K.B.; Gottlieb, A.; Koo, J.Y.M.; Lebwohl, M.; Lim, H.W.; et al. Guidelines of care for the management of psoriasis and psoriatic arthritis. Section 3. Guidelines of care for the management and treatment of psoriasis with topical therapies. J. Am. Acad. Dermatol. 2009, 60, 643–659. [Google Scholar] [CrossRef]
- Benezeder, T.; Painsi, C.; Patra, V.; Dey, S.; Holcmann, M.; Lange-Asschenfeldt, B.; Sibilia, M.; Wolf, P. Dithranol targets keratinocytes, their crosstalk with neutrophils and inhibits the IL-36 inflammatory loop in psoriasis. eLife 2020, 9, e56991. [Google Scholar] [CrossRef]
- Weinstein, G.D.; Koo, J.Y.M.; Krueger, G.G.; Lebwohl, M.G.; Lowe, N.J.; Menter, M.A.; Lew-Kaya, D.A.; Sefton, J.; Gibson, J.R.; Walker, P.S. Tazarotene cream in the treatment of psoriasis: Two multicenter, double-blind, randomized, vehicle-controlled studies of the safety and efficacy of tazarotene creams 0.05% and 0.1% applied once daily for 12 weeks. J. Am. Acad. Dermatol. 2003, 48, 760–767. [Google Scholar] [CrossRef]
- Lan, J.; Li, Y.; Wen, J.; Chen, Y.; Yang, J.; Zhao, L.; Xia, Y.; Du, H.; Tao, J.; Li, Y.; et al. Acitretin-Conjugated Dextran Nanoparticles Ameliorate Psoriasis-like Skin Disease at Low Dosages. Front. Bioeng. Biotechnol. 2021, 9, 816757. [Google Scholar] [CrossRef]
- Chiricozzi, A.; Panduri, S.; Dini, V.; Tonini, A.; Gualtieri, B.; Romanelli, M. Optimizing acitretin use in patients with plaque psoriasis. Dermatol. Ther. 2017, 30, e12453. [Google Scholar] [CrossRef]
- Atwan, A.; Ingram, J.R.; Abbott, R.; Kelson, M.J.; Pickles, T.; Bauer, A.; Piguet, V. Oral fumaric acid esters for psoriasis: Abridged Cochrane systematic review including GRADE assessments. Br. J. Dermatol. 2016, 175, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Afra, T.P.; Razmi, T.M.; Dogra, S. Apremilast in Psoriasis and Beyond: Big Hopes on a Small Molecule. Indian Dermatol. Online J. 2019, 10, 1–12. [Google Scholar] [PubMed]
- Bachelez, H.; van de Kerkhof, P.C.M.; Strohal, R.; Kubanov, A.; Valenzuela, F.; Lee, J.-H.; Yakusevich, V.; Chimenti, S.; Papacharalambous, J.; Proulx, J.; et al. Tofacitinib versus etanercept or placebo in moderate-to-severe chronic plaque psoriasis: A phase 3 randomised non-inferiority trial. Lancet 2015, 386, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Menter, A.; Gelfand, J.M.; Connor, C.; Armstrong, A.W.; Cordoro, K.M.; Davis, D.M.R.; Elewski, B.E.; Gordon, K.B.; Gottlieb, A.B.; Kaplan, D.H.; et al. Joint American Academy of Dermatology-National Psoriasis Foundation guidelines of care for the management of psoriasis with systemic nonbiologic therapies. J. Am. Acad. Dermatol. 2020, 82, 1445–1486. [Google Scholar] [CrossRef] [PubMed]
- Akhyani, M.; Chams-Davatchi, C.; Hemami, M.R.; Fateh, S. Efficacy and safety of mycophenolate mofetil vs. methotrexate for the treatment of chronic plaque psoriasis. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 1447–1451. [Google Scholar] [CrossRef]
- Armstrong, A.W.; Soliman, A.M.; Betts, K.A.; Wang, Y.; Gao, Y.; Puig, L.; Augustin, M. Comparative Efficacy and Relative Ranking of Biologics and Oral Therapies for Moderate-to-Severe Plaque Psoriasis: A Network Meta-analysis. Dermatol. Ther. 2021, 11, 885–905. [Google Scholar] [CrossRef]
- Gelfand, J.M.; Shin, D.B.; Alavi, A.; Torigian, D.A.; Werner, T.; Papadopoulos, M.; Takeshita, J.; Noe, M.H.; Dey, A.K.; Playford, M.P.; et al. A Phase IV, Randomized, Double-Blind, Placebo-Controlled Crossover Study of the Effects of Ustekinumab on Vascular Inflammation in Psoriasis (the VIP-U Trial). J. Investig. Dermatol. 2020, 140, 85–93.e2. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, M.; Torres, T. Guselkumab for the treatment of psoriasis—Evidence to date. Drugs Context 2019, 8, 212594. [Google Scholar] [CrossRef]
- Drerup, K.A.; Seemann, C.; Gerdes, S.; Mrowietz, U. Effective and Safe Treatment of Psoriatic Disease with the Anti-IL-23p19 Biologic Tildrakizumab: Results of a Real-World Prospective Cohort Study in Nonselected Patients. Dermatology 2021, 238, 615–619. [Google Scholar] [CrossRef]
- Gooderham, M.; Pinter, A.; Ferris, L.K.; Warren, R.B.; Zhan, T.; Zeng, J.; Soliman, A.M.; Kaufmann, C.; Kaplan, B.; Photowala, H.; et al. Long-term, durable, absolute Psoriasis Area and Severity Index and health-related quality of life improvements with risankizumab treatment: A post hoc integrated analysis of patients with moderate-to-severe plaque psoriasis. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 855–865. [Google Scholar] [CrossRef]
- Giunta, A.; Ventura, A.; Chimenti, M.S.; Bianchi, L.; Esposito, M. Spotlight on ixekizumab for the treatment of moderate-to-severe plaque psoriasis: Design, development, and use in therapy. Drug Des. Devel. Ther. 2017, 11, 1643–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruin, G.; Loesche, C.; Nyirady, J.; Sander, O. Population Pharmacokinetic Modeling of Secukinumab in Patients with Moderate to Severe Psoriasis. J. Clin. Pharmacol. 2017, 57, 876–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foulkes, A.C.; Warren, R.B. Brodalumab in psoriasis: Evidence to date and clinical potential. Drugs Context 2019, 8, 212570. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ma, P.; Cole, R.B.; Wang, G. In vitro metabolism of indomethacin morpholinylamide (BML-190), an inverse agonist for the peripheral cannabinoid receptor (CB2) in rat liver microsomes. Eur. J. Pharm. Sci. 2010, 41, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Maybruck, B.T.; Lam, W.C.; Specht, C.A.; Ilagan, M.X.G.; Donlin, M.J.; Lodge, J.K. The Aminoalkylindole BML-190 Negatively Regulates Chitosan Synthesis via the Cyclic AMP/Protein Kinase A1 Pathway in Cryptococcus neoformans. mBio 2019, 10, e02264-19. [Google Scholar] [CrossRef] [Green Version]
- Urasaki, Y.; Beaumont, C.; Workman, M.; Talbot, J.N.; Hill, D.K.; Le, T.T. Fast-Acting and Receptor-Mediated Regulation of Neuronal Signaling Pathways by Copaiba Essential Oil. Int. J. Mol. Sci. 2020, 21, 2259. [Google Scholar] [CrossRef] [Green Version]
- Tóth, B.I.; Dobrosi, N.; Dajnoki, A.; Czifra, G.; Oláh, A.; Szöllosi, A.G.; Juhász, I.; Sugawara, K.; Paus, R.; Bíró, T. Endocannabinoids modulate human epidermal keratinocyte proliferation and survival via the sequential engagement of cannabinoid receptor-1 and transient receptor potential vanilloid-1. J. Investig. Dermatol. 2011, 131, 1095–1104. [Google Scholar] [CrossRef]
- Chiurchiù, V.; Rapino, C.; Talamonti, E.; Leuti, A.; Lanuti, M.; Gueniche, A.; Jourdain, R.; Breton, L.; Maccarrone, M. Anandamide Suppresses Proinflammatory T Cell Responses In Vitro through Type-1 Cannabinoid Receptor-Mediated mTOR Inhibition in Human Keratinocytes. J. Immunol. 2016, 197, 3545–3553. [Google Scholar] [CrossRef] [Green Version]
- Łuczaj, W.; Dobrzyńska, I.; Wroński, A.; Domingues, M.R.; Domingues, P.; Skrzydlewska, E. Cannabidiol-Mediated Changes to the Phospholipid Profile of UVB-Irradiated Keratinocytes from Psoriatic Patients. Int. J. Mol. Sci. 2020, 21, 6592. [Google Scholar] [CrossRef]
- Degitz, K.; Placzek, M.; Borelli, C.; Plewig, G. Pathophysiology of acne. J. Dtsch. Dermatol. Ges. 2007, 5, 316–323. [Google Scholar] [CrossRef]
- Vos, T.; Flaxman, A.D.; Naghavi, M.; Lozano, R.; Michaud, C.; Ezzati, M.; Shibuya, K.; Salomon, J.A.; Abdalla, S.; Aboyans, V.; et al. Years lived with disability (YLDs) for 1160 sequelae of 289 diseases and injuries 1990–2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2163–2196. [Google Scholar] [CrossRef] [PubMed]
- Zaenglein, A.L. Acne Vulgaris. N. Engl. J. Med. 2018, 379, 1343–1352. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, H. Acne, the Skin Microbiome, and Antibiotic Treatment. Am. J. Clin. Dermatol. 2019, 20, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Sutaria, A.H.; Masood, S.; Schlessinger, J. Acne Vulgaris; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Heng, A.H.S.; Say, Y.-H.; Sio, Y.Y.; Ng, Y.T.; Chew, F.T. Gene variants associated with acne vulgaris presentation and severity: A systematic review and meta-analysis. BMC Med. Genom. 2021, 14, 103. [Google Scholar] [CrossRef] [PubMed]
- See, J.-A.; Goh, C.L.; Hayashi, N.; Suh, D.H.; Casintahan, F.A. Optimizing the use of topical retinoids in Asian acne patients. J. Dermatol. 2018, 45, 522–528. [Google Scholar] [CrossRef]
- Stamatiadis, D.; Bulteau-Portois, M.C.; Mowszowicz, I. Inhibition of 5 alpha-reductase activity in human skin by zinc and azelaic acid. Br. J. Dermatol. 1988, 119, 627–632. [Google Scholar] [CrossRef]
- Sieber, M.A.; Hegel, J.K.E. Azelaic acid: Properties and mode of action. Skin Pharmacol. Physiol. 2014, 27 (Suppl. 1), 9–17. [Google Scholar] [CrossRef]
- Liu, H.; Yu, H.; Xia, J.; Liu, L.; Liu, G.J.; Sang, H.; Peinemann, F. Topical azelaic acid, salicylic acid, nicotinamide, sulphur, zinc and fruit acid (alpha-hydroxy acid) for acne. Cochrane Database Syst. Rev. 2020, 5, CD011368. [Google Scholar]
- Lu, J.; Cong, T.; Wen, X.; Li, X.; Du, D.; He, G.; Jiang, X. Salicylic acid treats acne vulgaris by suppressing AMPK/SREBP1 pathway in sebocytes. Exp. Dermatol. 2019, 28, 786–794. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Z.; Sun, L.; Liu, H.; Zhang, F. Efficacy and safety of dapsone gel for acne: A systematic review and meta-analysis. Ann. Palliat. Med. 2022, 11, 611–620. [Google Scholar] [CrossRef]
- Rogers, L.R.; Oppelt, P.; Nayak, L. Hemolytic anemia associated with dapsone PCP prophylaxis in GBM patients with normal G6PD activity. Neuro Oncol. 2020, 22, 892–893. [Google Scholar] [CrossRef] [PubMed]
- Rademaker, M. Making sense of the effects of the cumulative dose of isotretinoin in acne vulgaris. Int. J. Dermatol. 2016, 55, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Arowojolu, A.O.; Gallo, M.F.; Lopez, L.M.; Grimes, D.A. Combined oral contraceptive pills for treatment of acne. Cochrane Database Syst. Rev. 2012, 6, CD004425. [Google Scholar]
- Han, J.J.; Faletsky, A.; Barbieri, J.S.; Mostaghimi, A. New Acne Therapies and Updates on Use of Spironolactone and Isotretinoin: A Narrative Review. Dermatol. Ther. 2021, 11, 79–91. [Google Scholar] [CrossRef]
- Oláh, A.; Tóth, B.I.; Borbíró, I.; Sugawara, K.; Szöllõsi, A.G.; Czifra, G.; Pál, B.; Ambrus, L.; Kloepper, J.; Camera, E.; et al. Cannabidiol exerts sebostatic and antiinflammatory effects on human sebocytes. J. Clin. Investig. 2014, 124, 3713–3724. [Google Scholar] [CrossRef] [Green Version]
- Oláh, A.; Markovics, A.; Szabó-Papp, J.; Szabó, P.T.; Stott, C.; Zouboulis, C.C.; Bíró, T. Differential effectiveness of selected non-psychotropic phytocannabinoids on human sebocyte functions implicates their introduction in dry/seborrhoeic skin and acne treatment. Exp. Dermatol. 2016, 25, 701–707. [Google Scholar] [CrossRef] [Green Version]
- Tubaro, A.; Giangaspero, A.; Sosa, S.; Negri, R.; Grassi, G.; Casano, S.; Della Loggia, R.; Appendino, G. Comparative topical anti-inflammatory activity of cannabinoids and cannabivarins. Fitoterapia 2010, 81, 816–819. [Google Scholar] [CrossRef]
- Jin, S.; Lee, M.-Y. The ameliorative effect of hemp seed hexane extracts on the Propionibacterium acnes-induced inflammation and lipogenesis in sebocytes. PLoS ONE 2018, 13, e0202933. [Google Scholar] [CrossRef]
- Jiang, Z.; Jin, S.; Fan, X.; Cao, K.; Liu, Y.; Wang, X.; Ma, Y.; Xiang, L. Cannabidiol Inhibits Inflammation Induced by Cutibacterium acnes-Derived Extracellular Vesicles via Activation of CB2 Receptor in Keratinocytes. J. Inflamm. Res. 2022, 15, 4573–4583. [Google Scholar] [CrossRef]
- Blaskovich, M.A.T.; Kavanagh, A.M.; Elliott, A.G.; Zhang, B.; Ramu, S.; Amado, M.; Lowe, G.J.; Hinton, A.O.; Pham, D.M.T.; Zuegg, J.; et al. The antimicrobial potential of cannabidiol. Commun. Biol. 2021, 4, 7. [Google Scholar] [CrossRef]
- Denton, C.P.; Khanna, D. Systemic sclerosis. Lancet 2017, 390, 1685–1699. [Google Scholar] [CrossRef] [PubMed]
- Dowson, C.; Simpson, N.; Duffy, L.; O’Reilly, S. Innate Immunity in Systemic Sclerosis. Curr. Rheumatol. Rep. 2017, 19, 2. [Google Scholar] [CrossRef] [PubMed]
- Bobeica, C.; Niculet, E.; Tatu, A.L.; Craescu, M.; Vata, D.; Statescu, L.; Iancu, A.V.; Musat, C.L.; Draganescu, M.L.; Onisor, C.; et al. Old and new therapeutic strategies in systemic sclerosis (Review). Exp. Ther. Med. 2022, 23, 134. [Google Scholar] [CrossRef] [PubMed]
- Denton, C.P. Advances in pathogenesis and treatment of systemic sclerosis. Clin. Med. 2016, 16, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barsotti, S.; Orlandi, M.; Codullo, V.; Di Battista, M.; Lepri, G.; Della Rossa, A.; Guiducci, S. One year in review 2019: Systemic sclerosis. Clin. Exp. Rheumatol. 2019, 37, 3–14. [Google Scholar]
- Hilberg, F.; Roth, G.J.; Krssak, M.; Kautschitsch, S.; Sommergruber, W.; Tontsch-Grunt, U.; Garin-Chesa, P.; Bader, G.; Zoephel, A.; Quant, J.; et al. BIBF 1120: Triple angiokinase inhibitor with sustained receptor blockade and good antitumor efficacy. Cancer Res. 2008, 68, 4774–4782. [Google Scholar] [CrossRef] [Green Version]
- Bütikofer, L.; Varisco, P.A.; Distler, O.; Kowal-Bielecka, O.; Allanore, Y.; Riemekasten, G.; Villiger, P.M.; Adler, S. ACE inhibitors in SSc patients display a risk factor for scleroderma renal crisis-a EUSTAR analysis. Arthritis Res. Ther. 2020, 22, 59. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.K. Pharmacological activation of PPAR-γ: A potential therapy for skin fibrosis. Int. J. Dermatol. 2021, 60, 376–383. [Google Scholar] [CrossRef]
- Liu, T.; de Los Santos, F.G.; Phan, S.H. The Bleomycin Model of Pulmonary Fibrosis. Methods Mol. Biol. 2017, 1627, 27–42. [Google Scholar]
- Servettaz, A.; Kavian, N.; Nicco, C.; Deveaux, V.; Chéreau, C.; Wang, A.; Zimmer, A.; Lotersztajn, S.; Weill, B.; Batteux, F. Targeting the cannabinoid pathway limits the development of fibrosis and autoimmunity in a mouse model of systemic sclerosis. Am. J. Pathol. 2010, 177, 187–196. [Google Scholar] [CrossRef]
- Balistreri, E.; Garcia-Gonzalez, E.; Selvi, E.; Akhmetshina, A.; Palumbo, K.; Lorenzini, S.; Maggio, R.; Lucattelli, M.; Galeazzi, M.; Distler, J.W.H. The cannabinoid WIN55, 212–2 abrogates dermal fibrosis in scleroderma bleomycin model. Ann. Rheum. Dis. 2011, 70, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Akhmetshina, A.; Dees, C.; Busch, N.; Beer, J.; Sarter, K.; Zwerina, J.; Zimmer, A.; Distler, O.; Schett, G.; Distler, J.H.W. The cannabinoid receptor CB2 exerts antifibrotic effects in experimental dermal fibrosis. Arthritis Rheum. 2009, 60, 1129–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Martín, A.; Navarrete, C.; Garrido-Rodríguez, M.; Prados, M.E.; Caprioglio, D.; Appendino, G.; Muñoz, E. EHP-101 alleviates angiotensin II-induced fibrosis and inflammation in mice. Biomed. Pharmacother. 2021, 142, 112007. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Chen, S.; Zhang, Q.; Yi, C.; He, J.; Ye, X.; Liu, M.; Lu, W. Celastrol is a novel selective agonist of cannabinoid receptor 2 with anti-inflammatory and anti-fibrotic activity in a mouse model of systemic sclerosis. Phytomedicine 2020, 67, 153160. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gonzalez, E.; Selvi, E.; Balistreri, E.; Lorenzini, S.; Maggio, R.; Natale, M.-R.; Capecchi, P.-L.; Lazzerini, P.-E.; Bardelli, M.; Laghi-Pasini, F.; et al. Cannabinoids inhibit fibrogenesis in diffuse systemic sclerosis fibroblasts. Rheumatology 2009, 48, 1050–1056. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, M.; McCann, M.; Liu, S.; Huh, K.; Denton, C.P.; Abraham, D.J.; Leask, A. Loss of peroxisome proliferator-activated receptor gamma in mouse fibroblasts results in increased susceptibility to bleomycin-induced skin fibrosis. Arthritis Rheum. 2009, 60, 2822–2829. [Google Scholar] [CrossRef]
- Cinar, R.; Park, J.K.; Zawatsky, C.N.; Coffey, N.J.; Bodine, S.P.; Abdalla, J.; Yokoyama, T.; Jourdan, T.; Jay, L.; Zuo, M.X.G.; et al. CB1 R and iNOS are distinct players promoting pulmonary fibrosis in Hermansky-Pudlak syndrome. Clin. Transl. Med. 2021, 11, e471. [Google Scholar] [CrossRef]
- Cinar, R.; Gochuico, B.R.; Iyer, M.R.; Jourdan, T.; Yokoyama, T.; Park, J.K.; Coffey, N.J.; Pri-Chen, H.; Szanda, G.; Liu, Z.; et al. Cannabinoid CB1 receptor overactivity contributes to the pathogenesis of idiopathic pulmonary fibrosis. JCI Insight 2017, 2, e92281. [Google Scholar] [CrossRef]
- Bronova, I.; Smith, B.; Aydogan, B.; Weichselbaum, R.R.; Vemuri, K.; Erdelyi, K.; Makriyannis, A.; Pacher, P.; Berdyshev, E.V. Protection from Radiation-Induced Pulmonary Fibrosis by Peripheral Targeting of Cannabinoid Receptor-1. Am. J. Respir. Cell Mol. Biol. 2015, 53, 555–562. [Google Scholar] [CrossRef] [Green Version]
- Palumbo-Zerr, K.; Horn, A.; Distler, A.; Zerr, P.; Dees, C.; Beyer, C.; Selvi, E.; Cravatt, B.F.; Distler, O.; Schett, G.; et al. Inactivation of fatty acid amide hydrolase exacerbates experimental fibrosis by enhanced endocannabinoid-mediated activation of CB1. Ann. Rheum. Dis. 2012, 71, 2051–2054. [Google Scholar] [CrossRef]
- Marquart, S.; Zerr, P.; Akhmetshina, A.; Palumbo, K.; Reich, N.; Tomcik, M.; Horn, A.; Dees, C.; Engel, M.; Zwerina, J.; et al. Inactivation of the cannabinoid receptor CB1 prevents leukocyte infiltration and experimental fibrosis. Arthritis Rheum. 2010, 62, 3467–3476. [Google Scholar] [CrossRef]
- Okogbaa, J.; Batiste, L. Dermatomyositis: An Acute Flare and Current Treatments. Clin. Med. Insights Case Rep. 2019, 12, 1179547619855370. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, T. Management of Myositis-Associated Interstitial Lung Disease. Medicina 2021, 57, 347. [Google Scholar] [CrossRef]
- Dressler, F.; Maurer, B. Dermatomyositis und juvenile Dermatomyositis. Z. Rheumatol. 2022; epub ahead of print. [Google Scholar] [CrossRef]
- Rodríguez-Tejero, A.; López-Espadafor, B.; Montero-Vílchez, T.; Sánchez-Díaz, M.; Arias-Santiago, S.; Molina-Leyva, A. Treatment challenges in clinically amyopathic dermatomyositis: A case series and review of new therapeutic options for skin involvement. Dermatol. Ther. 2021, 34, e14942. [Google Scholar] [CrossRef]
- Robinson, E.S.; Alves, P.; Bashir, M.M.; Zeidi, M.; Feng, R.; Werth, V.P. Cannabinoid Reduces Inflammatory Cytokines, Tumor Necrosis Factor-α, and Type I Interferons in Dermatomyositis In Vitro. J. Investig. Dermatol. 2017, 137, 2445–2447. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Zeidi, M.; Bonciani, D.; Pena, S.M.; Tiao, J.; Sahu, S.; Werth, V.P. Itch in dermatomyositis: The role of increased skin interleukin-31. Br. J. Dermatol. 2018, 179, 669–678. [Google Scholar] [CrossRef]
- Atkinson, S.D.; McGilligan, V.E.; Liao, H.; Szeverenyi, I.; Smith, F.J.D.; Moore, C.B.T.; McLean, W.H.I. Development of allele-specific therapeutic siRNA for keratin 5 mutations in epidermolysis bullosa simplex. J. Investig. Dermatol. 2011, 131, 2079–2086. [Google Scholar] [CrossRef] [Green Version]
- Baardman, R.; Yenamandra, V.K.; Duipmans, J.C.; Pasmooij, A.M.G.; Jonkman, M.F.; van den Akker, P.C.; Bolling, M.C. Novel insights into the epidemiology of epidermolysis bullosa (EB) from the Dutch EB Registry: EB more common than previously assumed? J. Eur. Acad. Dermatol. Venereol. 2021, 35, 995–1006. [Google Scholar] [CrossRef]
- Hou, P.-C.; Wang, H.-T.; Abhee, S.; Tu, W.-T.; McGrath, J.A.; Hsu, C.-K. Investigational Treatments for Epidermolysis Bullosa. Am. J. Clin. Dermatol. 2021, 22, 801–817. [Google Scholar] [CrossRef]
- Wally, V.; Lettner, T.; Peking, P.; Peckl-Schmid, D.; Murauer, E.M.; Hainzl, S.; Hintner, H.; Bauer, J.W. The pathogenetic role of IL-1β in severe epidermolysis bullosa simplex. J. Investig. Dermatol. 2013, 133, 1901–1903. [Google Scholar] [CrossRef] [Green Version]
- Castela, E.; Tulic, M.K.; Rozières, A.; Bourrat, E.; Nicolas, J.-F.; Kanitakis, J.; Vabres, P.; Bessis, D.; Mazereeuw, J.; Morice-Picard, F.; et al. Epidermolysis bullosa simplex generalized severe induces a T helper 17 response and is improved by apremilast treatment. Br. J. Dermatol. 2019, 180, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Shehadeh, W.; Sarig, O.; Bar, J.; Sprecher, E.; Samuelov, L. Treatment of epidermolysis bullosa pruriginosa-associated pruritus with dupilumab. Br. J. Dermatol. 2020, 182, 1495–1497. [Google Scholar] [CrossRef]
- Welponer, T.; Diem, A.; Nahler, G.; Laimer, M. Purified oral cannabidiol for pain management in severe recessive dystrophic epidermolysis bullosa. Indian J. Dermatol. Venereol. Leprol. 2022, 88, 551–552. [Google Scholar] [CrossRef]
- U.S. National Library of Medicine, ClinicalTrials.gov. The Efficacy and Safety of 3% Cannabidiol (CBD) Cream in Patients with Epidermolysis Bullosa: A Phase II/III Trial. Available online: https://clinicaltrials.gov/ct2/show/NCT04613102?term=CBD&cond=Skin+Diseases&draw=1&rank=6 (accessed on 15 June 2022).
- Martinez, A.E. Time to drop the stigma: Cannabinoids are drugs that may alleviate pain in people with epidermolysis bullosa. Br. J. Dermatol. 2019, 180, 711–712. [Google Scholar] [CrossRef]
- DeFilippis, E.M.; Feldman, S.R.; Huang, W.W. The genetics of pyoderma gangrenosum and implications for treatment: A systematic review. Br. J. Dermatol. 2015, 172, 1487–1497. [Google Scholar] [CrossRef]
- Alavi, A.; French, L.E.; Davis, M.D.; Brassard, A.; Kirsner, R.S. Pyoderma Gangrenosum: An Update on Pathophysiology, Diagnosis and Treatment. Am. J. Clin. Dermatol. 2017, 18, 355–372. [Google Scholar] [CrossRef]
- Shanmugam, V.K.; Couch, K.S.; McNish, S.; Amdur, R.L. Relationship between opioid treatment and rate of healing in chronic wounds. Wound Repair Regen. 2017, 25, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Stanley, P.L.; Steiner, S.; Havens, M.; Tramposch, K.M. Mouse skin inflammation induced by multiple topical applications of 12-O-tetradecanoylphorbol-13-acetate. Skin Pharmacol. 1991, 4, 262–271. [Google Scholar] [CrossRef]
- Nakajima, J.; Nakae, D.; Yasukawa, K. Structure-dependent inhibitory effects of synthetic cannabinoids against 12-O-tetradecanoylphorbol-13-acetate-induced inflammation and skin tumour promotion in mice. J. Pharm. Pharmacol. 2013, 65, 1223–1230. [Google Scholar] [CrossRef]
- Rundle, C.W.; Rietcheck, H.R.; Maghfour, J.; Dercon, S.; Fernandez, J.; Lio, P.; Dellavalle, R.P.; Fujita, M.; Yardley, H. Anti-inflammatory Effect of Cannabidiol and Palmitoylethanolamide Containing Topical Formulation on Skin in a 12-O-Tetradecanoylphorbol-13-Acetate-Induced Dermatitis Model in Mice. Dermatitis 2021, 33, 277–281. [Google Scholar] [CrossRef]
- Capozzi, A.; Caissutti, D.; Mattei, V.; Gado, F.; Martellucci, S.; Longo, A.; Recalchi, S.; Manganelli, V.; Riitano, G.; Garofalo, T.; et al. Anti-Inflammatory Activity of a CB2 Selective Cannabinoid Receptor Agonist: Signaling and Cytokines Release in Blood Mononuclear Cells. Molecules 2021, 27, 64. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.J.D.; Liao, H.; Cassidy, A.J.; Stewart, A.; Hamill, K.J.; Wood, P.; Joval, I.; van Steensel, M.A.M.; Björck, E.; Callif-Daley, F.; et al. The genetic basis of pachyonychia congenita. J. Investig. Dermatol. Symp. Proc. 2005, 10, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harries, M.J.; Paus, R. The pathogenesis of primary cicatricial alopecias. Am. J. Pathol. 2010, 177, 2152–2162. [Google Scholar] [CrossRef]
- Ramot, Y.; Oláh, A.; Paus, R. Cover Image: Neuroendocrine treatment of inherited keratin disorders by cannabinoids? Br. J. Dermatol. 2018, 178, 1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, B.; Laurino, C.; Vadalà, M. A therapeutic effect of cbd-enriched ointment in inflammatory skin diseases and cutaneous scars. Clin. Ter. 2019, 170, e93–e99. [Google Scholar] [PubMed]
- Scheffer, I.E.; Hulihan, J.; Messenheimer, J.; Ali, S.; Keenan, N.; Griesser, J.; Gutterman, D.L.; Sebree, T.; Sadleir, L.G. Safety and Tolerability of Transdermal Cannabidiol Gel in Children with Developmental and Epileptic Encephalopathies: A Nonrandomized Controlled Trial. JAMA Netw. Open 2021, 4, e2123930. [Google Scholar] [CrossRef] [PubMed]
- U.S. National Library of Medicine, ClinicalTrials.gov. The Pharmacokinetics and Pharmacodynamics of Hemp-based Topical Cannabinoid Products. ClinicalTrials.gov Identifier: NCT04741477. Available online: https://clinicaltrials.gov/ct2/show/NCT04741477?term=cannabinoid&draw=4&rank=7 (accessed on 15 June 2022).
- Peyravian, N.; Deo, S.; Daunert, S.; Jimenez, J.J. The Anti-Inflammatory Effects of Cannabidiol (CBD) on Acne. J. Inflamm. Res. 2022, 15, 2795–2801. [Google Scholar] [CrossRef]
- Spleman, L.; Sinclair, R.; Freeman, M.; Davis, M.; Gebauer, K. 1061 The safety of topical cannabidiol (CBD) for the treatment of acne. J. Investig. Dermatol. 2018, 138, S180. [Google Scholar] [CrossRef]
- U.S. National Library of Medicine, ClinicalTrials.gov. Evaluation of BTX 1503 in Patients with Moderate to Severe Acne Vulgaris”. ClinicalTrials.gov Identifier: NCT03573518. Available online: https://clinicaltrials.gov/ct2/show/NCT03573518?term=CBD&cond=Skin+Diseases&draw=1&rank=10 (accessed on 15 June 2022).
- Ali, A.; Akhtar, N. The safety and efficacy of 3% Cannabis seeds extract cream for reduction of human cheek skin sebum and erythema content. Pak. J. Pharm. Sci. 2015, 28, 1389–1395. [Google Scholar]
- Kurokawa, I.; Danby, F.W.; Ju, Q.; Wang, X.; Xiang, L.F.; Xia, L.; Chen, W.; Nagy, I.; Picardo, M.; Suh, D.H.; et al. New developments in our understanding of acne pathogenesis and treatment. Exp. Dermatol. 2009, 18, 821–832. [Google Scholar] [CrossRef]
- Ellis, C.N.; Krach, K.J. Uses and complications of isotretinoin therapy. J. Am. Acad. Dermatol. 2001, 45, S150–S157. [Google Scholar] [CrossRef] [PubMed]
- Rigopoulos, D.; Larios, G.; Katsambas, A.D. The role of isotretinoin in acne therapy: Why not as first-line therapy? facts and controversies. Clin. Dermatol. 2010, 28, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.; Satino, J. Hair Regrowth with Cannabidiol (CBD)-rich Hemp Extract—A Case Series. Cannabis 2021, 4, 53–59. [Google Scholar] [CrossRef]
- U.S. National Library of Medicine, ClinicalTrials.gov. Androgenetic Alopecia Treatment Using Varin and Cannabidiol Rich Topical Hemp Oil: A Case Series (Hair Regrowth). ClinicalTrials.gov Identifier: NCT04842383. Available online: https://clinicaltrials.gov/ct2/show/study/NCT04842383 (accessed on 15 June 2022).
- Buranakarn, V. Alopecia Areata Treatment with Extracted Cannabis Oil: Case Study 52-year-old Asian Male. Int. Jounal Sci. Innov. Technol. 2019, 2, 51–56. [Google Scholar]
- Specht, S.; Persaud, Y. Asteatotic Eczema; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Polat, M.; İlhan, M.N. Dermatological Complaints of the Elderly Attending a Dermatology Outpatient Clinic in Turkey: A Prospective Study over a One-year Period. Acta Dermatovenerol. Croat. 2015, 23, 277–281. [Google Scholar]
- Schulz, P.; Bunselmeyer, B.; Bräutigam, M.; Luger, T.A. Pimecrolimus cream 1% is effective in asteatotic eczema: Results of a randomized, double-blind, vehicle-controlled study in 40 patients. J. Eur. Acad. Dermatol. Venereol. 2007, 21, 90–94. [Google Scholar] [CrossRef]
- Yuan, C.; Wang, X.-M.; Guichard, A.; Tan, Y.-M.; Qian, C.-Y.; Yang, L.-J.; Humbert, P. N-palmitoylethanolamine and N-acetylethanolamine are effective in asteatotic eczema: Results of a randomized, double-blind, controlled study in 60 patients. Clin. Interv. Aging 2014, 9, 1163–1169. [Google Scholar] [CrossRef] [Green Version]
- Pulvirenti, N.; Nasca, M.R.; Micali, G. Topical adelmidrol 2% emulsion, a novel aliamide, in the treatment of mild atopic dermatitis in pediatric subjects: A pilot study. Acta Dermatovenerol. Croat. 2007, 15, 80–83. [Google Scholar]
- Reuters. Available online: https://www.reuters.com/article/brief-botanix-pharmaceuticals-says-btx-1-idUSFWN2BH1HT (accessed on 15 June 2022).
- Maghfour, J.; Rietcheck, H.R.; Rundle, C.W.; Runion, T.M.; Jafri, Z.A.; Dercon, S.; Lio, P.; Fernandez, J.; Fujita, M.; Dellavalle, R.P.; et al. An Observational Study of the Application of a Topical Cannabinoid Gel on Sensitive Dry Skin. J. Drugs Dermatol. 2020, 19, 1204–1208. [Google Scholar] [CrossRef]
- Callaway, J.; Schwab, U.; Harvima, I.; Halonen, P.; Mykkänen, O.; Hyvönen, P.; Järvinen, T. Efficacy of dietary hempseed oil in patients with atopic dermatitis. J. Dermatolog. Treat. 2005, 16, 87–94. [Google Scholar] [CrossRef]
- U.S. National Library of Medicine, ClinicalTrials.gov. A Phase Ib/IIa, Double-Blind, Randomized Study to Assess the Safety, Tolerability, Pharmacokinetics, and Pharmacodynamics of S-777469 in Subjects with Atopic Dermatitis. ClinicalTrials.gov Identifier: NCT00697710. Available online: https://clinicaltrials.gov/ct2/show/NCT00697710?term=cannabinoid&cond=skin&draw=1&rank=6 (accessed on 15 June 2022).
- Eberlein, B.; Eicke, C.; Reinhardt, H.-W.; Ring, J. Adjuvant treatment of atopic eczema: Assessment of an emollient containing N-palmitoylethanolamine (ATOPA study). J. Eur. Acad. Dermatol. Venereol. 2008, 22, 73–82. [Google Scholar] [CrossRef]
- U.S. National Library of Medicine, ClinicalTrials.gov. The Effectiveness of a Topical Palmitoylethanolamide (PEA) Formulation (Levagen+) for Reducing Symptoms of Eczema. ClinicalTrials.gov Identifier: NCT05003453. Available online: https://clinicaltrials.gov/ct2/show/NCT05003453?term=palmitoylethanolamide&draw=2&rank=7 (accessed on 15 June 2022).
- U.S. National Library of Medicine, ClinicalTrials.gov. Safety, Tolerability, and Efficacy of JBT-101 in Subjects with Dermatomyositis. ClinicalTrials.gov Identifier: NCT02466243. Available online: https://clinicaltrials.gov/ct2/show/NCT02466243?term=lenabasum&draw=1&rank=5 (accessed on 15 June 2022).
- Werth, V.P.; Hejazi, E.; Pena, S.M.; Haber, J.; Zeidi, M.; Reddy, N.; Okawa, J.; Feng, R.; Bashir, M.M.; Gebre, K.; et al. Safety and Efficacy of Lenabasum, a Cannabinoid Receptor Type 2 Agonist, in Patients with Dermatomyositis with Refractory Skin Disease: A Randomized Clinical Trial. J. Investig. Dermatol. 2022, 142, 2651–2659.e1. [Google Scholar] [CrossRef] [PubMed]
- U.S. National Library of Medicine, ClinicalTrials.gov. A Multicenter, Randomized, Double-Blind, Placebo-Controlled Phase 3 Trial to Evaluate Efficacy and Safety of Lenabasum in Dermatomyositis (DETERMINE). ClinicalTrials.gov Identifier: NCT03813160. Available online: https://clinicaltrials.gov/ct2/show/NCT03813160?cond=Dermatomyositis&draw=2&rank=6 (accessed on 15 June 2022).
- Werth, V.; White, B.; Dgetluck, N.; Hally, K.; Constantine, S.; Aggarwal, R.; Fiorentino, D.; Lundberg, I.E.; Oddis, C.V. OP0162 EFFICACY AND SAFETY OF LENABASUM IN THE PHASE 3 DETERMINE TRIAL IN DERMATOMYOSITIS. Ann. Rheum. Dis. 2022, 81, 106–107. [Google Scholar] [CrossRef]
- Amat-Samaranch, V.; Agut-Busquet, E.; Vilarrasa, E.; Puig, L. New perspectives on the treatment of hidradenitis suppurativa. Ther. Adv. Chronic Dis. 2021, 12, 20406223211055920. [Google Scholar] [CrossRef] [PubMed]
- Ingram, J.R.; Collier, F.; Brown, D.; Burton, T.; Burton, J.; Chin, M.F.; Desai, N.; Goodacre, T.E.E.; Piguet, V.; Pink, A.E.; et al. British Association of Dermatologists guidelines for the management of hidradenitis suppurativa (acne inversa) 2018. Br. J. Dermatol. 2019, 180, 1009–1017. [Google Scholar] [CrossRef] [Green Version]
- U.S. National Library of Medicine. Study to Investigate the Efficacy and Safety of Cannabis Oil for the Treatment of Subjects with Hidradenitis Suppurativa. ClinicalTrials.gov Identifer: NCT03929835. Available online: https://clinicaltrials.gov/ct2/show/NCT03929835 (accessed on 15 June 2022).
- Scheinfeld, N. Topical treatments of skin pain: A general review with a focus on hidradenitis suppurativa with topical agents. Dermatol. Online J. 2014, 20, 13030. [Google Scholar] [CrossRef]
- Dvorak, M.; Watkinson, A.; McGlone, F.; Rukwied, R. Histamine induced responses are attenuated by a cannabinoid receptor agonist in human skin. Inflamm. Res. 2003, 52, 238–245. [Google Scholar] [CrossRef]
- Wassilew, S.W. Zoster-associated neuralgias. J. Dtsch. Dermatol. Ges. 2006, 4, 871–879, quiz 880-1. [Google Scholar] [CrossRef]
- Phan, N.Q.; Siepmann, D.; Gralow, I.; Ständer, S. Adjuvant topical therapy with a cannabinoid receptor agonist in facial postherpetic neuralgia. J. Dtsch. Dermatol. Ges. 2010, 8, 88–91. [Google Scholar] [CrossRef]
- Visse, K.; Blome, C.; Phan, N.Q.; Augustin, M.; Ständer, S. Efficacy of Body Lotion Containing N-palmitoylethanolamine in Subjects with Chronic Pruritus due to Dry Skin: A Dermatocosmetic Study. Acta Derm. Venereol. 2017, 97, 639–641. [Google Scholar] [CrossRef] [Green Version]
- Szepietowski, J.C.; Szepietowski, T.; Reich, A. Efficacy and tolerance of the cream containing structured physiological lipids with endocannabinoids in the treatment of uremic pruritus: A preliminary study. Acta Dermatovenerol. Croat. 2005, 13, 97–103. [Google Scholar] [PubMed]
- Vela, J.; Dreyer, L.; Petersen, K.K.; Arendt-Nielsen, L.; Duch, K.S.; Kristensen, S. Cannabidiol treatment in hand osteoarthritis and psoriatic arthritis: A randomized, double-blind, placebo-controlled trial. Pain 2022, 163, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Friedman, A.J.; Momeni, K.; Kogan, M. Topical Cannabinoids for the Management of Psoriasis Vulgaris: Report of a Case and Review of the Literature. J. Drugs Dermatol. 2020, 19, 795. [Google Scholar] [CrossRef] [PubMed]
- Vincenzi, C.; Tosti, A. Efficacy and Tolerability of a Shampoo Containing Broad-Spectrum Cannabidiol in the Treatment of Scalp Inflammation in Patients with Mild to Moderate Scalp Psoriasis or Seborrheic Dermatitis. Skin Appendage Disord. 2020, 6, 355–361. [Google Scholar] [CrossRef]
- U.S. National Library of Medicine, ClinicalTrials.gov. Trial to Evaluate Efficacy and Safety of Lenabasum in Diffuse Cutaneous Systemic Sclerosis (RESOLVE-1). ClinicalTrials.gov Identifier: NCT03398837. Available online: https://clinicaltrials.gov/ct2/show/NCT03398837?term=cannabinoid&cond=skin&draw=1&rank=8 (accessed on 15 June 2022).
- Spiera, R.; Hummers, L.; Chung, L.; Frech, T.M.; Domsic, R.; Hsu, V.; Furst, D.E.; Gordon, J.; Mayes, M.; Simms, R.; et al. Safety and Efficacy of Lenabasum in a Phase II, Randomized, Placebo-Controlled Trial in Adults with Systemic Sclerosis. Arthritis Rheumatol. 2020, 72, 1350–1360. [Google Scholar] [CrossRef]
- Hinchcliff, M. Lenabasum for Skin Disease in Patients with Diffuse Cutaneous Systemic Sclerosis. Arthritis Rheumatol. 2020, 72, 1237–1240. [Google Scholar] [CrossRef]
- U.S. National Library of Medicine, ClinicalTrials.gov. Study to Evaluate the Safety, Tolerability and Efficacy of Cannabidiol (CBD) as a Steroid-sparing Therapy in Chronic Spontaneous Urticaria (CSU) Patients. ClinicalTrials.gov Identifier: NCT04439955. Available online: https://clinicaltrials.gov/ct2/show/NCT04439955?term=CBD&cond=Skin+Diseases&draw=1&rank=4 (accessed on 15 June 2022).
- Mallard, A.; Briskey, D.; Richards, A.; Mills, D.; Rao, A. The Effect of Orally Dosed Levagen+™ (palmitoylethanolamide) on Exercise Recovery in Healthy Males-A Double-Blind, Randomized, Placebo-Controlled Study. Nutrients 2020, 12, 596. [Google Scholar] [CrossRef]
- AminiLari, M.; Wang, L.; Neumark, S.; Adli, T.; Couban, R.J.; Giangregorio, A.; Carney, C.E.; Busse, J.W. Medical cannabis and cannabinoids for impaired sleep: A systematic review and meta-analysis of randomized clinical trials. Sleep 2022, 45, 234. [Google Scholar] [CrossRef]
- Moltke, J.; Hindocha, C. Reasons for cannabidiol use: A cross-sectional study of CBD users, focusing on self-perceived stress, anxiety, and sleep problems. J. Cannabis Res. 2021, 3, 5. [Google Scholar] [CrossRef]



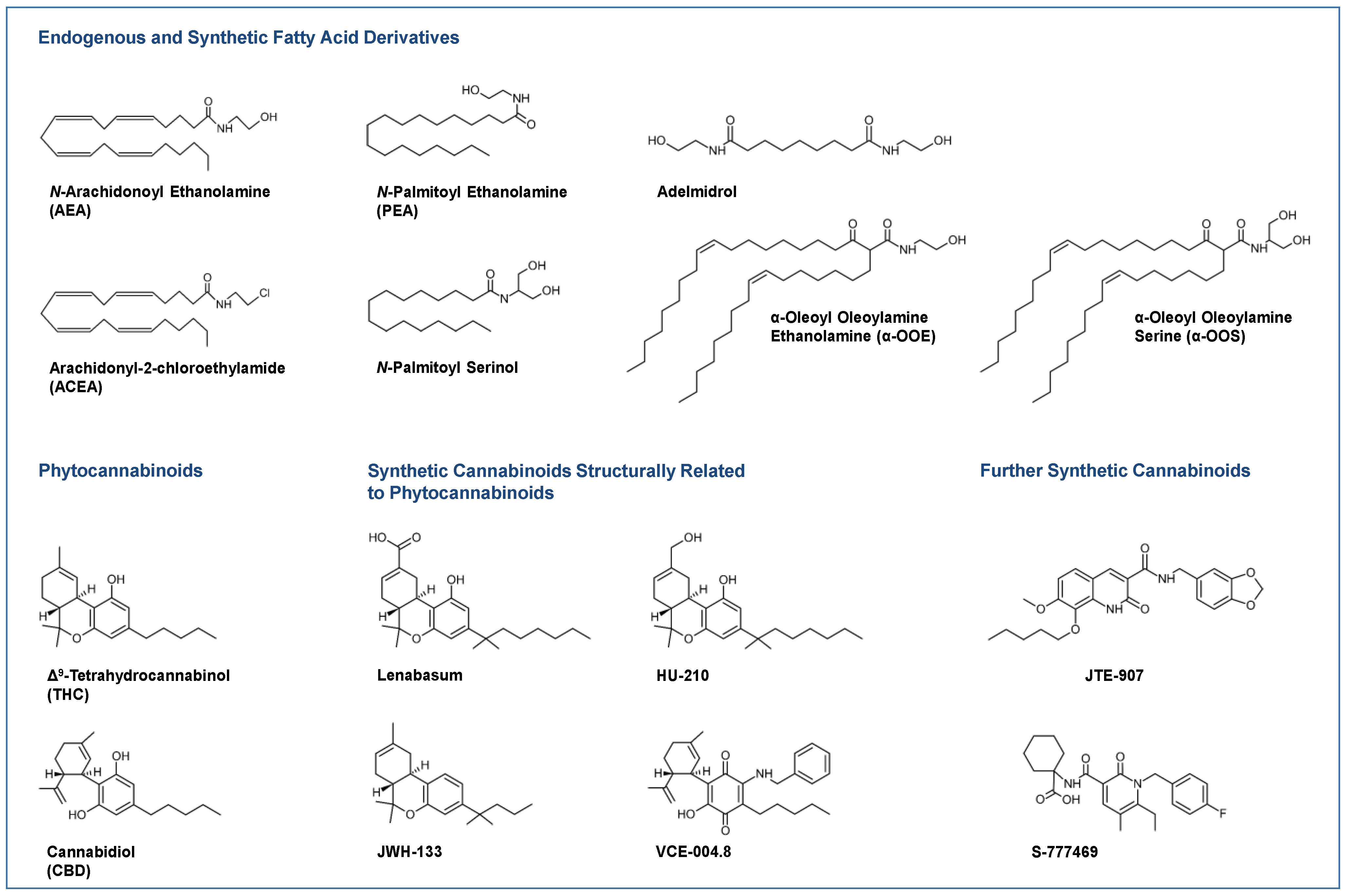{kind=link}
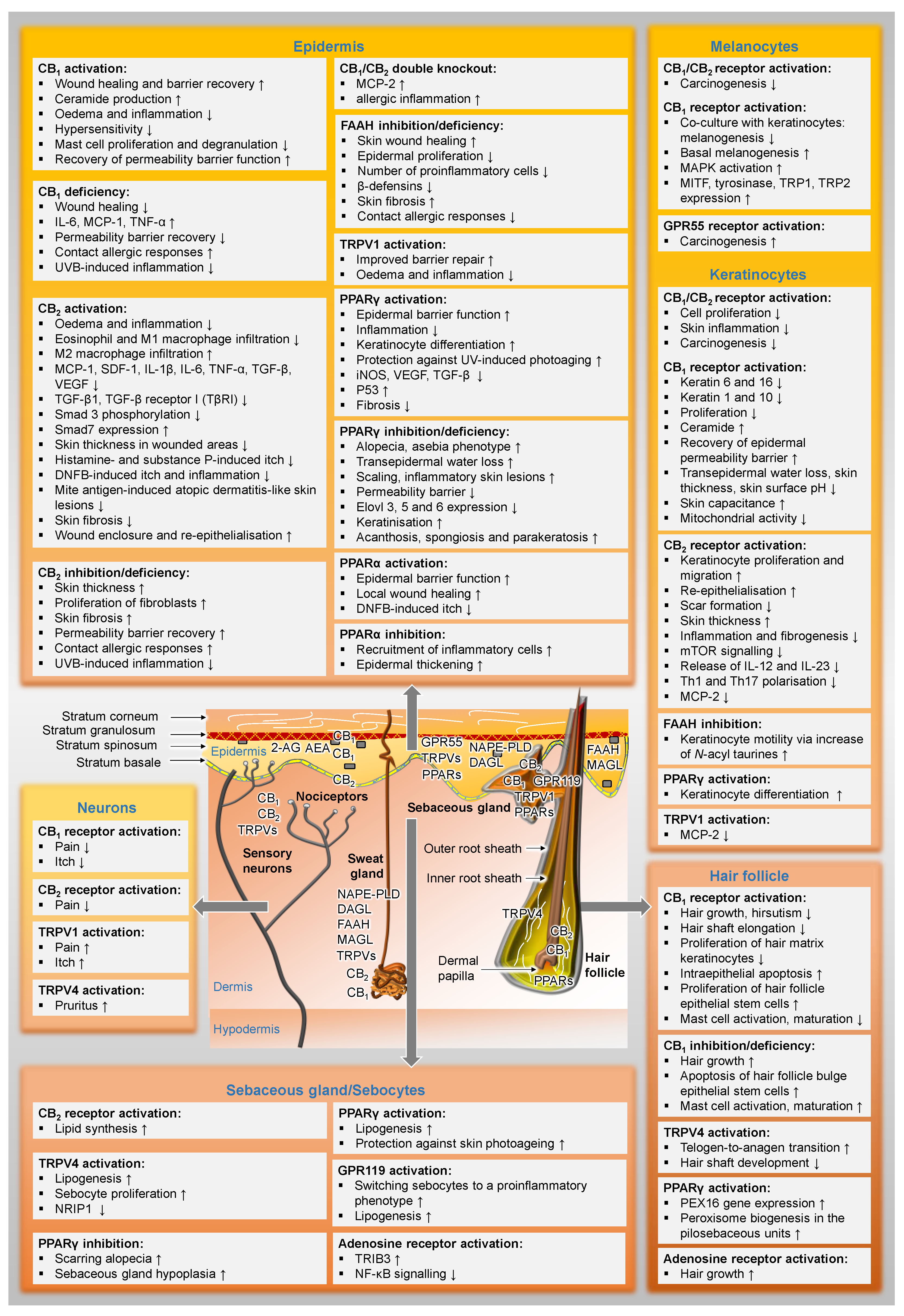{kind=link}
| Endocannabinoid/ Receptor | Disease | Regulation | Species | Cells/Tissue | Ref. |
|---|---|---|---|---|---|
| AEA | TPA-induced acute ear inflammation | ↔ | mouse | inflamed ear versus control ear | [170] |
| oxazolone-induced contact dermatitis | ↔ | mouse | oxazolone-treated ear versus control ear | [171] | |
| DNFB-induced contact dermatitis | ↑ | mouse | DNFB-exposed ear samples | [3] | |
| psoriasis vulgaris and psoriatic arthritis | ↑ | human | human plasma | [172] | |
| psoriasis | ↓ | human | keratinocytes from patients | [7] | |
| bleomycin-induced skin fibrosis | ↑ | mouse | fibrotic versus control skin | [169] | |
| 2-AG | TPA-induced acute ear inflammation | ↑ | mouse | inflamed ear versus control ear | [170] |
| oxazolone-induced contact dermatitis | ↑ | mouse | oxazolone-treated ear versus control ear | [171] | |
| DNFB-induced contact dermatitis | ↑ | mouse | DNFB-exposed ear samples | [3] | |
| mite antigen-induced dermatitis | ↑ | mouse | skin lesions versus healthy skin | [5] | |
| psoriasis vulgaris and psoriatic arthritis | ↑ | human | human plasma | [172] | |
| psoriasis | ↓ | human | keratinocytes from patients | [7] | |
| bleomycin-induced skin fibrosis | ↑ | mouse | fibrotic versus control skin | [169] | |
| CB1 | DNFB-induced contact dermatitis | ↓ | mouse | DNFB-exposed ear samples | [3] |
| psoriatic arthritis | ↑ | human | granulocytes from patients | [172] | |
| psoriasis vulgaris | ↔ | human | granulocytes from patients | ||
| atopic dermatitis | ↓ | human | itchy lesional skin | [173] | |
| psoriasis | ↓ | human | itchy lesional skin | ||
| seborrheic keratosis | ↓ | human | seborrhoeic keratosis affected tissue | [156] | |
| streptozotocin-induced type 1 diabetes | ↓ | mouse | skin tissue | [166] | |
| CB2 | DNFB-induced contact dermatitis | ↑ | mouse | DNFB-exposed ear samples | [3] |
| incised wounds | ↑ | mouse | various cell types | [175] | |
| psoriatic arthritis | ↔ | human | granulocytes from patients | [172] | |
| psoriasis vulgaris | ↑ | human | granulocytes from patients | ||
| atopic dermatitis | ↓ | human | itchy lesional skin | [173] | |
| psoriasis | ↓ | human | itchy lesional skin | ||
| dermatomyositis | ↑ | human | lesional skin and PBMCs | [8] | |
| imiquimod-induced psoriasis | ↑ | mouse | psoriatic skin lesion | [178] |
| Receptor | Disease | Regulation | Cells/Tissue | Ref. |
|---|---|---|---|---|
| TRPV1 | erythematotelangiectatic rosacea | ↑ | affected skin | [179] |
| phymatous rosacea | ↑ | affected skin | ||
| atopic dermatitis | ↑ | pruritic lesional skin | [173] | |
| psoriasis | ↑ | pruritic lesional skin | ||
| TRPV2 | erythematotelangiectatic rosacea | ↑ | affected skin | [179] |
| papulopustular rosacea | ↑ | affected skin | ||
| phymatous rosacea | ↓ | affected skin | ||
| atopic dermatitis | ↑ | pruritic lesional skin | [173] | |
| psoriasis | ↓ | pruritic lesional skin | ||
| TRPV3 | erythematotelangiectatic rosacea | ↑ | affected skin | [179] |
| papulopustular rosacea | ↑ | affected skin | ||
| phymatous rosacea | ↑ | affected skin | ||
| atopic dermatitis | ↓ | pruritic lesional skin | [173] | |
| psoriasis | ↑ | pruritic lesional skin | ||
| TRPV4 | phymatous rosacea | ↑ | affected skin | [179] |
| chronic idiopathic pruritus | ↑ | pruritic skin | [180] | |
| GPR55 | psoriasis vulgaris, psoriatic arthritis | ↑ | granulocytes from patients | [172] |
| GPR119 | acne | ↓ | sebaceous glands | [99] |
| PPARα | actinic keratosis | ↓ | affected skin | [181] |
| lichen planopilaris | ↔ | lymphocytic scarring alopecias | [149] | |
| atopic dermatitis | ↓ | eczematous skin | [182] | |
| melasma | ↓ | affected skin | [183] | |
| psoriasis | ↓ | pruritic lesional skin | [173] | |
| systemic sclerosis | ↓ | fibrotic skin tissue | [184] | |
| PPARγ | actinic keratosis | ↔ | affected skin | [181] |
| lichen planopilaris | ↓ | lymphocytic scarring alopecias | [149] | |
| photoageing in human skin | ↓ | photoaged or acutely UV-irradiated human skin | [185] | |
| atopic dermatitis | ↓ | atopic lesions | [186] | |
| psoriasis | ↓ | psoriatic lesions | ||
| psoriasis | ↓ | pruritic lesional skin | [173] | |
| systemic lupus erythematosus | ↓ | circulating CD14+ monocytes | [187] | |
| systemic sclerosis | ↓ | lesional skin fibroblasts | [98] | |
| systemic sclerosis | ↓ | fibrotic skin tissue | [184] | |
| PPARδ | psoriasis | ↑ | psoriatic lesions | [188] |
| lichen planopilaris | ↔ | lymphocytic scarring alopecias | [149] |
| Substance | Disease/ Condition | Study Design | Formulation | Outcome | Ref. |
|---|---|---|---|---|---|
| PEA | asteatotic eczema | randomised, monocentric, double-blind, comparative trial | Physiogel® A.I. Cream 1; combination with AEA | higher capacitance of skin surface; improved skin barrier function; reduction of pruritus | [400] |
| atopic dermatitis | randomised, investigator-blinded, split-body trial | Physiogel® A.I. Cream 1 | faster healing; reduction of flares | [17] | |
| atopic eczema | multinational, multicentre, observational, non-controlled, prospective cohort study | Physiogel® A.I. Cream 1 | reduction of skin dryness, excoriation, lichenification, scaling, erythema and pruritus | [406] | |
| randomised, interventional, double-blind trial | Levagen®+ 2 | no results posted yet | [407] | ||
| chronic pruritus | randomised, prospective, controlled, open-label, noninterventional study | Physiogel® Calming Relief A.I. Body Lotion | no significant improvement of pruritus and quality of life | [419] | |
| postherpetic neuralgia | open observational study; no placebo control | Physiogel® A.I. Cream 1 | reduction of pain and pruritus | [418] | |
| uremic pruritis | preliminary observational study; no placebo control | Derma Membrane Structure®; combination with AEA | reduction of pruritus and xerosis | [420] | |
| CBD | acne vulgaris | randomised, double-blind, placebo-controlled phase II study | BTX 1503 liquid formulation | no results posted yet | [389] |
| andro- genetic alopecia | case series study | CBD-rich hemp oil extract | enhanced hair regrowth | [394] | |
| atopic dermatitis | randomised, double-blind, placebo-controlled, phase II study | BTX 1204 liquid formulation | no significant improvement | [16,402] | |
| double-blind, placebo-controlled interventional study | JW-100 (topical formulation containing CBD and aspartame) | improvement in ISGA 3 scale | [18] | ||
| observational study | topical CBD | decrease in itch; improvement in eczema | [403] | ||
| chronic spon- taneous urticaria | placebo-controlled, open- label, single center, phase IIa study | oral CBD | no results posted yet | [427] | |
| psoriatic arthritis | randomised, double-blind, placebo-controlled trial | oral CBD | pain reduction | [421] | |
| scalp psoriasis/ seborrheic dermatitis | observational study | REVITA.CBD® SHAMPOO 4 | reduction of arborizing vessels, twisted capillaries, scales, erythema, itching and burning | [423] | |
| Lenabasum | dermato- myositis | randomised, single-center, double-blind, placebo-controlled phase II study | oral JBT-101 | reduction of CDASI 5 | [408,409] |
| randomised, multicenter, double-blind, placebo-controlled, phase III study | oral JBT-101 | reduction of CDASI 5 (patients without muscle weakness); improvement in TIS 6 (patients with muscle weakness) | [410,411] | ||
| healthy volunteers 7 | randomised, open-label, placebo-controlled study | oral anabasum | inhibition of neutrophil infiltration; resolution of inflammation | [221] | |
| systemic sclerosis | randomised, placebo- controlled phase II trial | oral lenabasum | improvement in CRISS 8 | [424,425] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramer, R.; Hinz, B. Cannabinoid Compounds as a Pharmacotherapeutic Option for the Treatment of Non-Cancer Skin Diseases. Cells 2022, 11, 4102. https://doi.org/10.3390/cells11244102
Ramer R, Hinz B. Cannabinoid Compounds as a Pharmacotherapeutic Option for the Treatment of Non-Cancer Skin Diseases. Cells. 2022; 11(24):4102. https://doi.org/10.3390/cells11244102
Chicago/Turabian StyleRamer, Robert, and Burkhard Hinz. 2022. "Cannabinoid Compounds as a Pharmacotherapeutic Option for the Treatment of Non-Cancer Skin Diseases" Cells 11, no. 24: 4102. https://doi.org/10.3390/cells11244102
APA StyleRamer, R., & Hinz, B. (2022). Cannabinoid Compounds as a Pharmacotherapeutic Option for the Treatment of Non-Cancer Skin Diseases. Cells, 11(24), 4102. https://doi.org/10.3390/cells11244102