
The Role of LIM Kinases during Development: A Lens to Get a Glimpse of Their Implication in Pathologies


Abstract
:1. Introduction
2. Expression of LIMKs during Development

{kind=link}
| Publications | LIMK Isoform Species | Embryonic/Adult Tissues—Cell Lines | Experimental Procedures | Main Observations |
|---|---|---|---|---|
| Mizuno et al. Oncogene, 1994 [1] | LIMK human, rat | adult rat brain, epithelial and hematopoietic cell lines | Northern blot | High level in the rat brain. Expressed in human epithelial and hematopoietic cell lines |
| Bernard et al. Cell Growth and Differentiation, 1994 [2] | LIMK human, mouse | E13, E14, E15, E16, E18, P0 mouse brain, adult human brain, mouse brain, heart, liver, muscle mouse olfactory epithelial cell lines | Northern blot, RNase protection assay in situ hybridization, immunohistochemistry | Identification of LIMK. Expressed in human and mouse brain and olfactory epithelial cell lines |
| Ohashi et al. Journal of Biochemistry, 1994 [42] | LIMK chicken | adult lung, brain, kidney, liver, gizzard, intestine, spleen | Northern blot | Expressed in lung, brain, kidney, liver, spleen, gizzard and intestine |
| Cheng and Robertson Mechanisms of Development, 1995 [43] | LIMK mouse | E8.5, E11.5, E15.5 brain, olfactory system, gut, trophoblast giant cells adult brain, ovary, testis, skin, lung | Northern blot, in situ hybridization | Variable expression rates depending on the stage of development and the tissue |
| Okano et al. Journal of Biological Chemistry, 1995 [34] | LIMK1 LIMK2 human | adult brain, skeletal muscle, heart, placenta | Northern blot | LIMK1 expressed in all tissues, with highest amounts in the brain. Two LIMK2 isoforms: longer in all tissues, smaller only in skeletal muscle and heart |
| Pröschel et al. Oncogene, 1995 [44] | LIMK1 mouse | adult spinal cord, brain, cranial nerve, dorsal root ganglia | Northern blot, in situ hybridization | Nervous system expression of LIMK1 |
| Nunoue et al. Oncogene, 1995 [3] | LIMK1 LIMK2 rat | adult brain, various tissues | Northern blot | LIMK1 in the brain, LIMK2 in various tissues |
| Ikebe et al. Genomics, 1997 [45] | LIMK2 mouse | adult brain, thymus, lung, heart, stomach, spleen, kidney, intestine, liver, testis | RT-PCR | LIMK2a and LIMK2b isoforms expressed in various tissues |
| Koshimizu et al. Biochemistry and Biophysical Research Communications, 1997 [39] | LIMK2 mouse | E10 to E18 embryos adult brain, heart, lung, spleen, thymus, kidney, stomach | Northern blot | LIMK2a and LIMK2b isoforms expressed in various tissues |
| Mori et al. Molecular Brain Research, 1997 [36] | LIMK1 LIMK2 rat | E12, E14, E16, E18 embryo | In situ hybridization | LIMK1 and LIMK2 expressed in brain. Differential expression of LIMK1 and LIMK2 in epithelia. High expression in extra-embryonic tissues |
| Takahashi et al. Developmental Dynamics, 1997 [46] | LIMK1 xenopus | Stage 2 to 40 cleavage, gastrula, blastula, neurula | Northern blot, in situ hybridization | Variable expression rates during development. Important role of XLIMK1 in neural development |
| Ikebe et al. Biochemistry and Biophysical Research Communications, 1998 [35] | LIMK2 mouse | adult liver, brain, thymus, lung, heart, stomach, testis | Northern blot, RT-PCR | Identification of LIMK2c, a brain-specific isoform, and LIMK2t, a testis-specific isoform |
| Takahashi et al. Biochemistry and Biophysical Research Communications, 1998 [47] | LIMK1 LIMK2 mouse | adult brain, thymus, lung, spleen, testis, kidney, stomach, heart | Northern blot, in situ hybridization | LIMK2 expressed in all tissues, identification of a testis-specific isoform LIMK2t |
| Nomoto et al. Genes, 1999 [37] | LIMK2 human | fetal and adult brain, stomach, colon, pancreas, liver, lung, kidney, placenta | RT-PCR, RNase protection assay | Identification of LIMK2a and LIMK2b with tissue-specific expression profile. LIMK2a predominantly expressed in fetal and adult tissues compared to LIMK2b |
| Meng et al. Neuron, 2002 [48] | LIMK1 mouse | adult brain | LIMK1 KO mice, immunohistochemistry, primary neurons, brain sections | Dendritic spine morphology and synaptic function alterations |
| Takahashi et al. Developmental Biology, 2002 [41] | LIMK2 mouse | adult testis | LIMK2 KO mice, MEF cells, immunofluorescence, histology | Abnormal spermatogenesis found in LIMK2-KO testis associated with an increased number of apoptotic germ cells |
| Meng et al. Neuropharmacology, 2004 [49] | LIMK1 LIMK2 mouse | adult brain | LIMK1, LIMK2 and LIMK1/2 KO mice, immunohistochemistry, electrophysiology | Normal synaptic plasticity in LIMK2-KO mice, altered synaptic functions in double-LIMK1/2-KO mice |
| Chen et al. Current Biology, 2004 [50] | dLIMK drosophila | from larvae to adult leg morphogenesis | mRNA level | Expression in late larval and pupal stages, suggesting a role in this transition. Defects in leg morphogenesis. Role of the Rho-dLIMK signaling pathway. |
| Foletta et al. Experimental Cell Research, 2004 [51] | LIMK1 rat, mouse, chicken | rat and chick embryos brain and spinal cord mouse adult brain, heart, liver, lung, small intestine, stomach, kidney | Western blot | Expression of LIMK1 in liver, thymus, kidney, heart, lung, small intestine, stomach and brain |
| Ang et al. Developmental Biology, 2006 [52] | dLIMK drosophila | larvae neuromuscular junctions (abdominal muscle fibers), antennal lobe glomeruli | dLIMK active/inactive, drosophila strains, immunohistochemistry, electrophysiology | Role of LIMK in synapse development and in glomeruli of antennal lobe. LIMK is a downstream effector of PAK |
| Acevedo et al. Journal of Histochemistry and Cytochemistry, 2006 [38] | LIMK2 mouse | E14 olfactory epithelium, heart, liver, intestine, urogenital sinus, thymus, spinal cord adult brain, heart, spleen, stomach, intestine, lung, skin, kidney, ovary, eyes, testes, uterus | Embryo sections, immunohistochemistry, western blot | Variable LIMK2 expression levels in embryonic and adult tissues, similar expression pattern than LIMK1 except in testis |
| Menzel et al. Mechanism of Development, 2007 [53] | dLIMK drosophila | adult eyes | Genetic screen, mutant, drosophila strains, immunohistochemistry of photoreceptor cell | PAK-LIMK-cofilin pathway are involved in photoreceptor cell morphogenesis by regulating adherent junctions and actin dynamics |
| Ott et al. Gene Expression Patterns, 2007 [54] | LIMK1 LIMK2 zebrafish | all embryonic stages | In situ hybridization | Temporal and spatial expression of LIMK1 and LIMK2 during embryogenesis |
| Lindström et al. Gene Expression Patterns, 2011 [33] | LIMK1 mouse | E10.5 to E18.5 EMT- and MET-tissues, limb, eye, heart, lung, skin, kidney, intestine, testes | Embryo sections, immunohistochemistry | LIMK1 highly expressed in many neuronal and epithelial tissues undergoing EMT and MET |
| Rice et al. PLoS ONE, 2012 [40] | LIMK2 mouse | E14.5 E15.5 E18.5 and newborns P1.5 ocular tissue adult brain, testis, eyes, rate, lung | LIMK2-KO mice, RT-PCR, in situ hybridization, western blot, immunohistochemistry | Phenotype of EOB “eyes open at birth” of LIMK2-KO mice, abnormal migration of keratinocytes during eyelid development |
| Andrews et al. Biology Open, 2013 [55] | LIMK2 mouse | E13.5, E15.5 brain | In situ hybridization, siRNA transfections, in utero electroporation | Role of LIMK2 in growth cone collapse in response to Sema3A by regulating PlexinA1 expression level |
| Kawano et al. Bone, 2013 [56] | LIMK1 mouse | newborns PD3-PD5 tibiae, femur | LIMK1-KO mice, bone histomorphometry, microCT, primary osteoblasts, osteoclasts and bone marrow cells | Bone mass reduction in LIMK1-KO mice, abnormal osteoblast differentiation and defective osteoblastic and osteoclastic functions |
| Abe et al. Development, 2014 [57] | dLIMK drosophila | newborns P2-P3 brain | Drosophila strains, immunohistochemistry | Involvement of Rac-Sickie-SSH and Rac-PAK-LIMK pathways in axonal growth |
| Piccioli et al. Journal of Neuroscience, 2014 [58] | dLIMK drosophila | larvae neuromuscular junctions | Drosophila strains, live imaging of synaptic growth and bouton budding | Role of BMPRII-LIMK-cofilin-actin signaling in potentialization of neuromuscular junctions |
| Yang et al. Bone, 2015 [59] | LIMK2 mouse | newborns PD2-PD3 primary osteoblasts | siRNA transfections, immunofluorescence, fluid shear stress | Contribution of LIMK2 in the mechanosensitivity of osteoblasts |
| Xie et al. Histochemistry and Cell Biology, 2017 [60] | LIMK1 mouse | E15.5, E18.5, newborns P1 brain | In utero electroporation, brain sections, immunofluorescence | Altered neuronal migration and number of neurites due to aberrant expression of LIMK1 |
| Duan et al. Cell Cycle, 2018 [32] | LIMK1 LIMK2 mouse | 2, 4, 8 -cells morula, blastocyst | In vitro fertilization, embryo culture, immunofluorescence, inhibition of LIMKs activity | LIMK1 and LIMK2 are involved in early stages of embryo development and regulate actin assembly |
| Saxena et al. Development, 2018 [61] | LIMK mouse | E11.5, E13.5, E15.5 cortex and cortical neurons postnatal P0, P6, P21 neurons | Immunohistochemistry, cell proliferation, in utero-electroporation, P-SMAD labeling | Regulation of dendritic branching by LIMK-mediated non-canonical BMP signaling and involvement of both canonical and non-canonical BMP signaling in neuronal migration |
| Mao et al. Molecular Brain, 2019 [62] | LIMK1 LIMK2 mouse | E14.5 brain | LIMK1-KO, LIMK2-KO and double LIMK1/2-KO mice, immunohistochemistry | Contribution of LIMK1 and LIMK2 in progenitor cell proliferation and migration. Role of LIMK2 in embryonic cell apoptosis |
| Fang et al. Scientific Reports, 2019 [63] | LIMK1 LIMK2 mouse | E3 and from P3 to P30 cochlea | LIMK1/2-KO mice, immunohistochemistry, auditory measurement | No alteration of cochlear development and auditory function in LIMK1/2-KO mice |
| Kwon et al. Asia-Australasian Journal of Animal Sciences, 2020 [31] | LIMK1 LIMK2 porcine | 1, 2, 4-cells morula, blastocyst | RT-QPCR, LIMK1/2 activity inhibition, LIMK1/2 dsRNA injection, embryo culture, immunofluorescence | Role of LIMK1 and LIMK2 in embryo cleavage and compaction through actin regulation and the maintenance of cell–cell junctions |
| He et al. In Vitro Cellular and Developmental Biology, 2021 [64] | LIMK2 human | Embryonic Stem Cells | endodermal differentiation, siRNA transfection, RT-QPCR, immuno-fluorescence | Control of actin assembly, EMT-related genes expression and cell migration by LIMK2 in endodermal lineage |
3. Signaling Pathways Involving LIMKs during Development
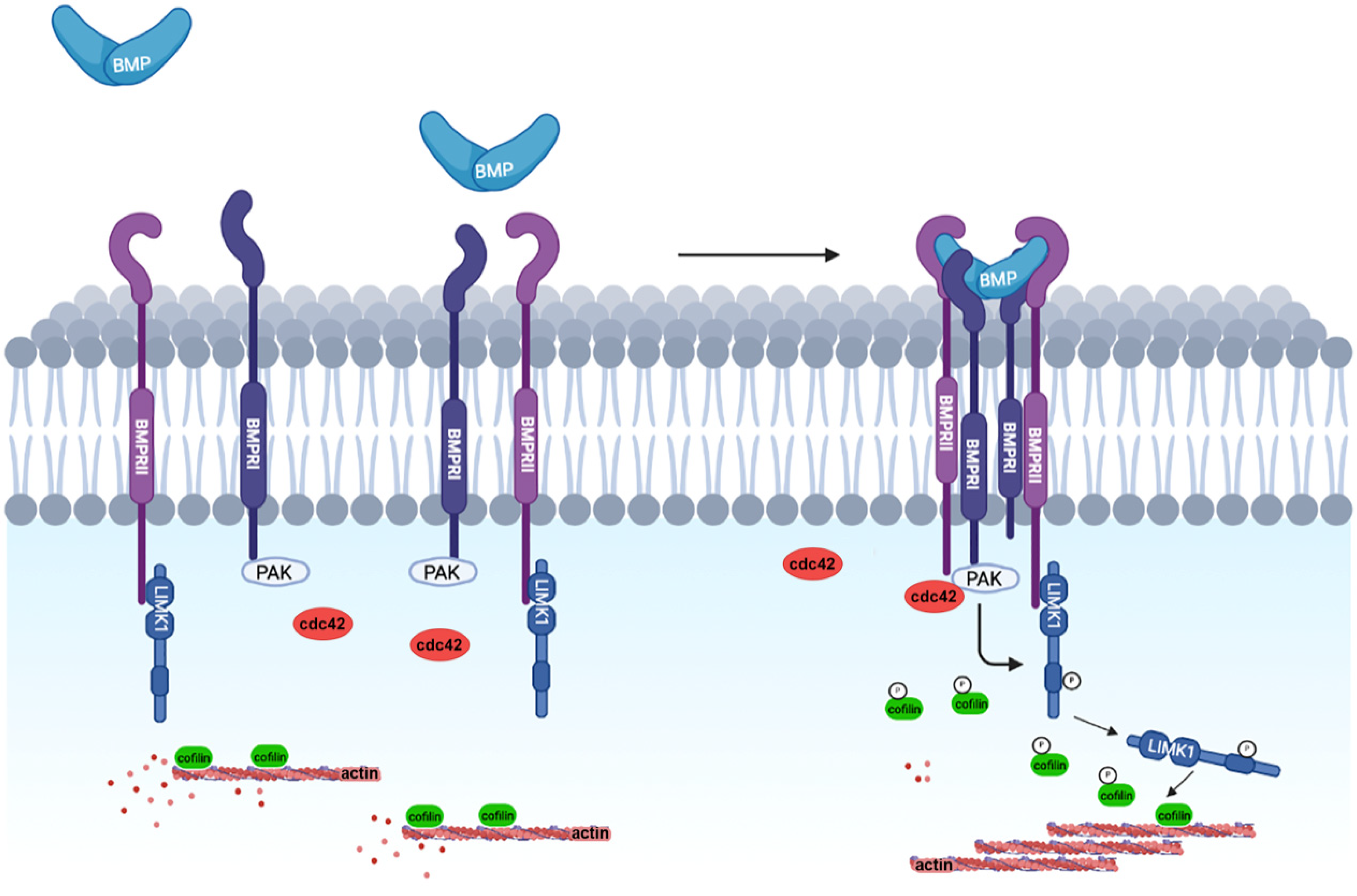
3.1. The Non-Canonical BMP Pathway
3.2. Nerve Growth Factor
3.3. Semaphorins
4. The Role of LIMKs during Embryonic Cell Migration
4.1. Role of LIM Kinases in Neural Progenitor Migration
4.2. Role of LIM Kinases in Keratinocyte Migration
5. Role of LIMKs in Epithelial-Mesenchymal Transitions
6. Impact of LIMKs in Cell Differentiation
6.1. Neuron Differentiation
6.2. Bone Cell Differentiation
6.3. Gonadal Cell Differentiation
6.4. Gland Morphogenesis
6.5. Blood Cells
7. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mizuno, K.; Okano, I.; Ohashi, K.; Nunoue, K.; Kuma, K.; Miyata, T.; Nakamura, T. Identification of a human cDNA encoding a novel protein kinase with two repeats of the LIM/double zinc finger motif. Oncogene 1994, 9, 1605–1612. [Google Scholar]
- Bernard, O.; Ganiatsas, S.; Kannourakis, G.; Dringen, R. Kiz-1, a protein with LIM zinc finger and kinase domains, is expressed mainly in neurons. Cell Growth Differ. 1994, 5, 1159–1171. [Google Scholar]
- Nunoue, K.; Ohashi, K.; Okano, I.; Mizuno, K. LIMK-1 and LIMK-2, two members of a LIM motif-containing protein kinase family. Oncogene 1995, 11, 701–710. [Google Scholar]
- Kadrmas, J.L.; Beckerle, M.C. The LIM domain: From the cytoskeleton to the nucleus. Nat. Rev. Mol. Cell Biol. 2004, 5, 920–931. [Google Scholar] [CrossRef]
- Te Velthuis, A.J.W.; Bagowski, C.P. PDZ and LIM domain-encoding genes: Molecular interactions and their role in development. Sci. World J. 2007, 7, 1470–1492. [Google Scholar] [CrossRef] [Green Version]
- Krcmery, J.; Camarata, T.; Kulisz, A.; Simon, H.-G. Nucleocytoplasmic functions of the PDZ-LIM protein family: New insights into organ development. BioEssays 2010, 32, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Arber, S.; Barbayannis, F.A.; Hanser, H.; Schneider, C.; Stanyon, C.A.; Bernard, O.; Caroni, P. Regulation of actin dynamics through phosphorylation of cofilin by LIM-kinase. Nature 1998, 393, 805–809. [Google Scholar] [CrossRef]
- Yang, N.; Higuchi, O.; Ohashi, K.; Nagata, K.; Wada, A.; Kangawa, K.; Nishida, E.; Mizuno, K. Cofilin phosphorylation by LIM-kinase 1 and its role in Rac-mediated actin reorganization. Nature 1998, 393, 809–812. [Google Scholar] [CrossRef]
- Sumi, T.; Matsumoto, K.; Takai, Y.; Nakamura, T. Cofilin phosphorylation and actin cytoskeletal dynamics regulated by rho- and Cdc42-activated LIM-kinase 2. J. Cell Biol. 1999, 147, 1519–1532. [Google Scholar] [CrossRef] [Green Version]
- Nikhil, K.; Chang, L.; Viccaro, K.; Jacobsen, M.; McGuire, C.; Satapathy, S.R.; Tandiary, M.; Broman, M.M.; Cresswell, G.; He, Y.J.; et al. Identification of LIMK2 as a therapeutic target in castration resistant prostate cancer. Cancer Lett. 2019, 448, 182–196. [Google Scholar] [CrossRef] [Green Version]
- Ou, S.; Tan, M.-H.; Weng, T.; Li, H.; Koh, C.-G. LIM kinase1 regulates mitotic centrosome integrity via its activity on dynein light intermediate chains. Open Biol. 2018, 8, 170202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prunier, C.; Prudent, R.; Kapur, R.; Sadoul, K.; Lafanechère, L. LIM kinases: Cofilin and beyond. Oncotarget 2017, 8, 41749–41763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCullough, B.R.; Grintsevich, E.E.; Chen, C.K.; Kang, H.; Hutchison, A.L.; Henn, A.; Cao, W.; Suarez, C.; Martiel, J.L.; Blanchoin, L.; et al. Cofilin-linked changes in actin filament flexibility promote severing. Biophys. J. 2011, 101, 151–159. [Google Scholar] [CrossRef] [Green Version]
- McCullough, B.R.; Blanchoin, L.; Martiel, J.L.; De la Cruz, E.M. Cofilin increases the bending flexibility of actin filaments: Implications for severing and cell mechanics. J. Mol. Biol. 2008, 381, 550–558. [Google Scholar] [CrossRef] [Green Version]
- Toshima, J.; Toshima, J.Y.; Takeuchi, K.; Mori, R.; Mizuno, K. Cofilin Phosphorylation and Actin Reorganization Activities of Testicular Protein Kinase 2 and Its Predominant Expression in Testicular Sertoli Cells. J. Biol. Chem. 2001, 276, 31449–31458. [Google Scholar] [CrossRef] [Green Version]
- Toshima, J.; Toshima, J.Y.; Amano, T.; Yang, N.; Narumiya, S.; Mizuno, K. Cofilin phosphorylation by protein kinase testicular protein kinase 1 and its role in integrin-mediated actin reorganization and focal adhesion formation. Mol. Biol. Cell 2001, 12, 1131–1145. [Google Scholar] [CrossRef] [Green Version]
- Nakano, K.; Kanai-Azuma, M.; Kanai, Y.; Moriyama, K.; Yazaki, K.; Hayashi, Y.; Kitamura, N. Cofilin phosphorylation and actin polymerization by NRK/NESK, a member of the germinal center kinase family. Exp. Cell Res. 2003, 287, 219–227. [Google Scholar] [CrossRef]
- Ritchey, L.; Chakrabarti, R. Aurora A kinase modulates actin cytoskeleton through phosphorylation of Cofilin: Implication in the mitotic process. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 2719–2729. [Google Scholar] [CrossRef] [Green Version]
- Niwa, R.; Nagata-Ohashi, K.; Takeichi, M.; Mizuno, K.; Uemura, T. Control of actin reorganization by Slingshot, a family of phosphatases that dephosphorylate ADF/cofilin. Cell 2002, 108, 233–246. [Google Scholar] [CrossRef] [Green Version]
- Gohla, A.; Birkenfeld, J.; Bokoch, G.M. Chronophin, a novel HAD-type serine protein phosphatase, regulates cofilin-dependent actin dynamics. Nat. Cell Biol. 2005, 7, 21–29. [Google Scholar] [CrossRef]
- Scott, R.W.; Olson, M.F. LIM kinases: Function, regulation and association with human disease. J. Mol. Med. 2007, 85, 555–568. [Google Scholar] [CrossRef]
- Ohashi, K. Roles of cofilin in development and its mechanisms of regulation. Dev. Growth Differ. 2015, 57, 275–290. [Google Scholar] [CrossRef]
- Pober, B.R. Medical progress: Williams-Beuren syndrome. N. Engl. J. Med. 2010, 362, 239–252. [Google Scholar] [CrossRef]
- Li, H.H.; Roy, M.; Kuscuoglu, U.; Spencer, C.M.; Halm, B.; Harrison, K.C.; Bayle, J.H.; Splendore, A.; Ding, F.; Meltzer, L.A.; et al. Induced chromosome deletions cause hypersociability and other features of Williams-Beuren syndrome in mice. EMBO Mol. Med. 2009, 1, 50–65. [Google Scholar] [CrossRef]
- Ben Zablah, Y.; Zhang, H.; Gugustea, R.; Jia, Z. LIM-Kinases in synaptic plasticity, memory, and brain diseases. Cells 2021, 10, 2079. [Google Scholar] [CrossRef] [PubMed]
- Frangiskakis, J.M.; Ewart, A.K.; Morris, C.A.; Mervis, C.B.; Bertrand, J.; Robinson, B.F.; Klein, B.P.; Ensing, G.J.; Everett, L.A.; Green, E.D.; et al. LIM-kinase1 hemizygosity implicated in impaired visuospatial constructive cognition. Cell 1996, 86, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Kundu, J.K.; Chae, J.I.; Shim, J.H. Targeting ROCK/LIMK/cofilin signaling pathway in cancer. Arch. Pharm. Res. 2019, 42, 481–491. [Google Scholar] [CrossRef]
- Manetti, F. Recent findings confirm LIM domain kinases as emerging target candidates for cancer therapy. Curr. Cancer Drug Targets 2012, 12, 543–560. [Google Scholar] [CrossRef]
- Manetti, F. LIM kinases are attractive targets with many macromolecular partners and only a few small molecule regulators. Med. Res. Rev. 2012, 32, 968–998. [Google Scholar] [CrossRef]
- Chatterjee, D.; Preuss, F.; Dederer, V.; Knapp, S.; Mathea, S. Structural Aspects of LIMK Regulation and Pharmacology. Cells 2022, 11, 142. [Google Scholar] [CrossRef]
- Kwon, J.; Seong, M.J.; Piao, X.; Jo, Y.J.; Kim, N.H. Limk1/2 are required for actin filament and cell junction assembly in porcine embryos developing in vitro. Asian-Australas. J. Anim. Sci. 2020, 33, 1579–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, X.; Zhang, H.L.; Wu, L.L.; Liu, M.Y.; Pan, M.H.; Ou, X.H.; Sun, S.C. Involvement of LIMK1/2 in actin assembly during mouse embryo development. Cell Cycle 2018, 17, 1381–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindström, N.O.; Neves, C.; McIntosh, R.; Miedzybrodzka, Z.; Vargesson, N.; Martin Collinson, J. Tissue specific characterisation of Lim-kinase 1 expression during mouse embryogenesis. Gene Expr. Patterns 2011, 11, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okano, I.; Hiraoka, J.; Otera, H.; Nunoue, K.; Ohashi, K.; Iwashita, S.; Hirai, M.; Mizuno, K. Identification and characterization of a novel family of serine/threonine kinases containing two N-terminal LIM motifs. J. Biol. Chem. 1995, 270, 31321–31330. [Google Scholar] [CrossRef] [Green Version]
- Ikebe, C.; Ohashi, K.; Mizuno, K. Identification of testis-specific (Limk2t) and brain-specific (Limk2c) isoforms of mouse LIM-Kinase 2 gene transcripts. Biochem. Biophys. Res. Commun. 1998, 246, 307–312. [Google Scholar] [CrossRef]
- Mori, T.; Okano, I.; Mizuno, K.; Tohyama, M.; Wanaka, A. Comparison of tissue distribution of two novel serine/threonine kinase genes containing the LIM motif (LIMK-1 and LIMK-2) in the developing rat. Mol. Brain Res. 1997, 45, 247–254. [Google Scholar] [CrossRef]
- Nomoto, S.; Tatematsu, Y.; Takahashi, T.; Osada, H. Cloning and characterization of the alternative promoter regions of the human LIMK2 gene responsible for alternative transcripts with tissue-specific expression. Gene 1999, 236, 259–271. [Google Scholar] [CrossRef]
- Acevedo, K.; Moussi, N.; Li, R.; Soo, P.; Bernard, O. LIM kinase 2 is widely expressed in all tissues. J. Histochem. Cytochem. 2006, 54, 487–501. [Google Scholar] [CrossRef] [Green Version]
- Koshimizu, U.; Takahashi, H.; Yoshida, M.C.; Nakamura, T. cDNA cloning, genomic organization, and chromosomal localization of the mouse LIM motif-containing kinase gene, Limk2. Biochem. Biophys. Res. Commun. 1997, 241, 243–250. [Google Scholar] [CrossRef]
- Rice, D.S.; Hansen, G.M.; Liu, F.; Crist, M.J.; Newhouse, M.M.; Potter, D.; Xu, N.; Abuin, A.; Vogel, P.J.; Zambrowicz, B.P. Keratinocyte migration in the developing eyelid requires LIMK2. PLoS ONE 2012, 7, e47168. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Koshimizu, U.; Miyazaki, J.; Nakamura, T. Impaired spermatogenic ability of testicular germ cells in mice deficient in the LIM-kinase 2 gene. Dev. Biol. 2002, 241, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, K.; Toshima, J.; Tajinda, K.; Nakamura, T.; Mizuno, K. Molecular cloning of a chicken lung cDNA encoding a novel protein kinase with N-terminal two lim/double zinc finger motifs. J. Biochem. 1994, 116, 636–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, A.K.; Robertson, E.J. The murine LIM-kinase gene (limk) encodes a novel serine threonine kinase expressed predominantly in trophoblast giant cells and the developing nervous system. Mech. Dev. 1995, 52, 187–197. [Google Scholar] [CrossRef]
- Pröschel, C.; Blouin, M.J.; Gutowski, N.J.; Ludwig, R.; Noble, M. Limk1 is predominantly expressed in neural tissues and phosphorylates serine, threonine and tyrosine residues in vitro. Oncogene 1995, 11, 1271–1281. [Google Scholar] [PubMed]
- Ikebe, C.; Ohashi, K.; Fujimori, T.; Bernard, O.; Noda, T.; Robertson, E.J.; Mizuno, K. Mouse LIM-kinase 2 gene: cDNA cloning, genomic organization, and tissue-specific expression of two alternatively initiated transcripts. Genomics 1997, 46, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Aoki, S.; Nakamura, T.; Koshimizu, U.; Matsumoto, K.; Nakamura, T. Xenopus LIM motif-containing protein kinase, Xlimk1, is expressed in the developing head structure of the embryo. Dev. Dyn. 1997, 209, 196–205. [Google Scholar] [CrossRef]
- Takahashi, H.; Koshimizu, U.; Nakamura, T. Novel transcript encoding truncated LIM kinase 2 is specifically expressed in male germ cells undergoing meiosis. Biochem. Biophys. Res. Commun. 1998, 249, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Zhang, Y.; Tregoubov, V.; Janus, C.; Cruz, L.; Jackson, M.; Lu, W.Y.; MacDonald, J.F.; Wang, J.Y.; Falls, D.L.; et al. Abnormal spine morphology and enhanced LTP in LIMK-1 knockout mice. Neuron 2002, 35, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Takahashi, H.; Meng, J.; Zhang, Y.; Lu, G.; Asrar, S.; Nakamura, T.; Jia, Z. Regulation of ADF/cofilin phosphorylation and synaptic function by LIM-kinase. Neuropharmacology 2004, 47, 746–754. [Google Scholar] [CrossRef]
- Chen, G.-C.; Gajowniczek, P.; Settleman, J. Rho-LIM Kinase Signaling Regulates Ecdysone-Induced Gene Expression and Morphogenesis during Drosophila Metamorphosis. Curr. Biol. 2004, 14, 309–313. [Google Scholar] [CrossRef]
- Foletta, V.C.; Moussi, N.; Sarmiere, P.D.; Bamburg, J.R.; Bernard, O. LIM kinase 1, a key regulator of actin dynamics, is widely expressed in embryonic and adult tissues. Exp. Cell Res. 2004, 294, 392–405. [Google Scholar] [CrossRef] [PubMed]
- Ang, L.H.; Chen, W.; Yao, Y.; Ozawa, R.; Tao, E.; Yonekura, J.; Uemura, T.; Keshishian, H.; Hing, H. Lim kinase regulates the development of olfactory and neuromuscular synapses. Dev. Biol. 2006, 293, 178–190. [Google Scholar] [CrossRef] [Green Version]
- Menzel, N.; Schneeberger, D.; Raabe, T. The Drosophila p21 activated kinase Mbt regulates the actin cytoskeleton and adherens junctions to control photoreceptor cell morphogenesis. Mech. Dev. 2007, 124, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Ott, E.B.; te Velthuis, A.J.W.; Bagowski, C.P. Comparative analysis of splice form-specific expression of LIM Kinases during zebrafish development. Gene Expr. Patterns 2007, 7, 620–629. [Google Scholar] [CrossRef] [PubMed]
- Andrews, W.D.; Zito, A.; Memi, F.; Jones, G.; Tamamaki, N.; Parnavelas, J.G. Limk2 mediates semaphorin signalling in cortical interneurons migrating through the subpallium. Biol. Open 2013, 2, 277–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawano, T.; Zhu, M.; Troiano, N.; Horowitz, M.; Bian, J.; Gundberg, C.; Kolodziejczak, K.; Insogna, K. LIM kinase 1 deficient mice have reduced bone mass. Bone 2013, 52, 70–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, T.; Yamazaki, D.; Murakami, S.; Hiroi, M.; Nitta, Y.; Maeyama, Y.; Tabata, T. The NAV2 homolog sickie regulates F-actin-mediated axonal growth in Drosophila mushroom body neurons via the noncanonical Rac-Cofilin pathway. Development 2014, 141, 4716–4728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccioli, Z.D.; Littleton, J.T. Retrograde BMP signaling modulates rapid activity-dependent synaptic growth via presynaptic lim kinase regulation of Cofilin. J. Neurosci. 2014, 34, 4371–4381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Tan, S.; Shen, Y.; Chen, R.; Wu, C.; Xu, Y.; Song, Z.; Fu, Q. Inhibition of FSS-induced actin cytoskeleton reorganization by silencing LIMK2 gene increases the mechanosensitivity of primary osteoblasts. Bone 2015, 74, 182–190. [Google Scholar] [CrossRef]
- Xie, J.; Li, X.; Zhang, W.; Chai, X.; Huang, Y.; Li, K.; Cheng, X.; Zhao, S. Aberrant expression of LIMK1 impairs neuronal migration during neocortex development. Histochem. Cell Biol. 2017, 147, 471–479. [Google Scholar] [CrossRef]
- Saxena, M.; Agnihotri, N.; Sen, J. Perturbation of canonical and non-canonical BMP signaling affects migration, polarity and dendritogenesis of mouse cortical neurons. Development 2018, 145, dev147157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, R.; Deng, R.; Wei, Y.; Han, L.; Meng, Y.; Xie, W.; Jia, Z. LIMK1 and LIMK2 regulate cortical development through affecting neural progenitor cell proliferation and migration. Mol. Brain 2019, 12, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Q.; Zhang, Y.; Da, P.; Shao, B.; Pan, H.; He, Z.; Cheng, C.; Li, D.; Guo, J.; Wu, X.; et al. Deletion of Limk1 and Limk2 in mice does not alter cochlear development or auditory function. Sci. Rep. 2019, 9, 3357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Zhang, L.; He, Y.; Yu, H.; Li, S.; Li, Q. Involvement of LIMK2 in actin cytoskeleton remodeling during the definitive endoderm differentiation. Vitr. Cell. Dev. Biol. Anim. 2021, 57, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Foletta, V.C.; Lim, M.A.; Soosairajah, J.; Kelly, A.P.; Stanley, E.G.; Shannon, M.; He, W.; Das, S.; Massague, J.; Bernard, O.; et al. Direct signaling by the BMP type II receptor via the cytoskeletal regulator LIMK1. J. Cell Biol. 2003, 162, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Lee-Hoeflich, S.T.; Causing, C.G.; Podkowa, M.; Zhao, X.; Wrana, J.L.; Attisano, L. Activation of LIMK1 by binding to the BMP receptor, BMPRII, regulates BMP-dependent dendritogenesis. EMBO J. 2004, 23, 4792–4801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podkowa, M.; Christova, T.; Zhao, X.; Jian, Y.; Attisano, L. P21-Activated kinase (PAK) is required for Bone Morphogenetic Protein (BMP)-induced dendritogenesis in cortical neurons. Mol. Cell. Neurosci. 2013, 57, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K.; Varadarajan, S.G.; Li, J.E.; Butler, S.J. Type Ib BMP receptors mediate the rate of commissural axon extension through inhibition of cofilin activity. Development 2013, 140, 333–342. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.; Han, L.; Bamburg, J.R.; Shim, S.; Ming, G.L.; Zheng, J.Q. BMP gradients steer nerve growth cones by a balancing act of LIM kinase and Slingshot phosphatase on ADF/cofilin. J. Cell Biol. 2007, 178, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Phan, K.D.; Hazen, V.M.; Frendo, M.; Jia, Z.; Butler, S.J. The bone morphogenetic protein roof plate chemorepellent regulates the rate of commissural axonal growth. J. Neurosci. 2010, 30, 15430–15440. [Google Scholar] [CrossRef]
- Frendo, M.E.; da Silva, A.; Phan, K.D.; Riche, S.; Butler, S.J. The cofilin/limk1 pathway controls the growth rate of both developing and regenerating motor axons. J. Neurosci. 2019, 39, 9316–9327. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ren, T.; Jiao, G.; Huang, Y.; Bao, X.; Zhang, F.; Liu, K.; Zheng, B.; Sun, K.; Guo, W. BMPR2 promotes invasion and metastasis via the RhoA-ROCKLIMK2 pathway in human osteosarcoma cells. Oncotarget 2017, 8, 58625–58641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Tang, P.; Guo, F.; Zhang, M.; Chen, Y.; Yan, Y.; Tian, Z.; Xu, P.; Zhang, L.; Zhang, L.; et al. RhoA regulates Activin B-induced stress fiber formation and migration of bone marrow-derived mesenchymal stromal cell through distinct signaling. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 3011–3018. [Google Scholar] [CrossRef] [PubMed]
- Vardouli, L.; Moustakas, A.; Stournaras, C. LIM-kinase 2 and cofilin phosphorylation mediate actin cytoskeleton reorganization induced by transforming growth factor-β. J. Biol. Chem. 2005, 280, 11448–11457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, M.; Ohashi, K.; Mizuno, K. LIM kinase and slingshot are critical for neurite extension. J. Biol. Chem. 2007, 282, 13692–13702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aizawa, H.; Wakatsuki, S.; Ishii, A.; Moriyama, K.; Sasaki, Y.; Ohashi, K.; Sekine-Aizawa, Y.; Sehara-Fujisawa, A.; Mizuno, K.; Goshima, Y.; et al. Phosphorylation of cofilin by LIM-kinase is necessary for semaphorin 3A-induced growth cone collapse. Nat. Neurosci. 2001, 4, 367–373. [Google Scholar] [CrossRef]
- Duncan, B.W.; Mohan, V.; Wade, S.D.; Truong, Y.; Kampov-Polevoi, A.; Temple, B.R.; Maness, P.F. Semaphorin3F Drives Dendritic Spine Pruning Through Rho-GTPase Signaling. Mol. Neurobiol. 2021, 58, 3817–3834. [Google Scholar] [CrossRef]
- Das, R.M.; Storey, K.G. Apical abscission alters cell polarity and dismantles the primary cilium during neurogenesis. Science. 2014, 343, 200–204. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, A. Neuronal Delamination and Outer Radial Glia Generation in Neocortical Development. Front. Cell Dev. Biol. 2021, 8, 623573. [Google Scholar] [CrossRef]
- Thumkeo, D.; Shimizu, Y.; Sakamoto, S.; Yamada, S.; Narumiya, S. ROCK-I and ROCK-II cooperatively regulate closure of eyelid and ventral body wall in mouse embryo. Genes Cells 2005, 10, 825–834. [Google Scholar] [CrossRef]
- Shimizu, Y.; Thumkeo, D.; Keel, J.; Ishizaki, T.; Oshima, H.; Oshima, M.; Noda, Y.; Matsumura, F.; Taketo, M.M.; Narumiya, S. ROCK-I regulates closure of the eyelids and ventral body wall by inducing assembly of actomyosin bundles. J. Cell Biol. 2005, 168, 941–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.; Zhang, Y.; Tregoubov, V.; Falls, D.L.; Jia, Z. Regulation of spine morphology and synaptic function by LIMK and the actin cytoskeleton. Rev. Neurosci. 2003, 14, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.W.; Hooper, S.; Crighton, D.; Li, A.; Konig, I.; Munro, J.; Trivier, E.; Wickman, G.; Morin, P.; Croft, D.R.; et al. LIM kinases are required for invasive path generation by tumor and tumor-associated stromal cells. J. Cell Biol. 2010, 191, 169–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- De Craene, B.; Berx, G. Regulatory networks defining EMT during cancer initiation and progression. Nat. Rev. Cancer 2013, 13, 97–110. [Google Scholar] [CrossRef]
- Sakurai, K.; Talukdar, I.; Patil, V.S.; Dang, J.; Li, Z.; Chang, K.Y.; Lu, C.C.; Delorme-Walker, V.; Dermardirossian, C.; Anderson, K.; et al. Kinome-wide functional analysis highlights the role of cytoskeletal remodeling in somatic cell reprogramming. Cell Stem Cell 2014, 14, 523–534. [Google Scholar] [CrossRef] [Green Version]
- Dupin, E.; Calloni, G.W.; Coelho-Aguiar, J.M.; Le Douarin, N.M. The issue of the multipotency of the neural crest cells. Dev. Biol. 2018, 444 (Suppl. 1), S47–S59. [Google Scholar] [CrossRef]
- Radu, A.G.; Torch, S.; Fauvelle, F.; Pernet-Gallay, K.; Lucas, A.; Blervaque, R.; Delmas, V.; Schlattner, U.; Lafanechère, L.; Hainaut, P.; et al. LKB1 specifies neural crest cell fates through pyruvate-alanine cycling. Sci. Adv. 2019, 5, eaau5106. [Google Scholar] [CrossRef] [Green Version]
- Park, K.S.; Gumbiner, B.M. Cadherin-6B stimulates an epithelial mesenchymal transition and the delamination of cells from the neural ectoderm via LIMK/cofilin mediated non-canonical BMP receptor signaling. Dev. Biol. 2012, 366, 232–243. [Google Scholar] [CrossRef] [Green Version]
- Andrieu, C.; Montigny, A.; Bibonne, A.; Despin-Guitard, E.; Alfandari, D.; Théveneau, E. MMP14 is required for delamination of chick neural crest cells independently of its catalytic activity. Development 2020, 147, dev183954. [Google Scholar] [CrossRef]
- Steffen, A.; Le Dez, G.; Poincloux, R.; Recchi, C.; Nassoy, P.; Rottner, K.; Galli, T.; Chavrier, P. MT1-MMP-Dependent Invasion Is Regulated by TI-VAMP/VAMP7. Curr. Biol. 2008, 18, 926–931. [Google Scholar] [CrossRef] [PubMed]
- Lagoutte, E.; Villeneuve, C.; Lafanechère, L.; Wells, C.M.C.M.; Jones, G.E.G.E.; Chavrier, P.; Rossé, C. LIMK Regulates Tumor-Cell Invasion and Matrix Degradation Through Tyrosine Phosphorylation of MT1-MMP. Sci. Rep. 2016, 6, 24925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Estrada, O.M.; Lettice, L.A.; Essafi, A.; Guadix, J.A.; Slight, J.; Velecela, V.; Hall, E.; Reichmann, J.; Devenney, P.S.; Hohenstein, P.; et al. Wt1 is required for cardiovascular progenitor cell formation through transcriptional control of Snail and E-cadherin. Nat. Genet. 2010, 42, 89–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamprecht, R. Actin cytoskeleton role in the maintenance of neuronal morphology and long-term memory. Cells 2021, 10, 1795. [Google Scholar] [CrossRef] [PubMed]
- George, J.; Soares, C.; Montersino, A.; Beique, J.C.; Thomas, G.M. Palmitoylation of LIM Kinase-1 ensures Spine-Specific actin polymerization and morphological plasticity. elife 2015, 2015, e06327. [Google Scholar] [CrossRef]
- Chen, P.; Jing, H.; Xiong, M.; Zhang, Q.; Lin, D.; Ren, D.; Wang, S.; Yin, D.; Chen, Y.; Zhou, T.; et al. Spine impairment in mice high-expressing neuregulin 1 due to LIMK1 activation. Cell Death Dis. 2021, 12, 403. [Google Scholar] [CrossRef]
- Gory-Fauré, S.; Powell, R.; Jonckheere, J.; Lanté, F.; Denarier, E.; Peris, L.; Nguyen, C.H.; Buisson, A.; Lafanechère, L.; Andrieux, A. Pyr1-Mediated Pharmacological Inhibition of LIM Kinase Restores Synaptic Plasticity and Normal Behavior in a Mouse Model of Schizophrenia. Front. Pharmacol. 2021, 12, 323. [Google Scholar] [CrossRef]
- Chen, L.; Shi, K.; Frary, C.E.; Ditzel, N.; Hu, H.; Qiu, W.; Kassem, M. Inhibiting actin depolymerization enhances osteoblast differentiation and bone formation in human stromal stem cells. Stem Cell Res. 2015, 15, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Huang, Y.; Li, H.; Hu, J.; Liu, X.; Jiang, T.; Sun, G.; Tang, A.; Sun, X.; Qian, W.; et al. Excess of rare variants in genes that are key epigenetic regulators of spermatogenesis in the patients with non-obstructive azoospermia. Sci. Rep. 2015, 5, 8785. [Google Scholar] [CrossRef]
- Romarowski, A.; Battistone, M.A.; La Spina, F.A.; Puga Molina, L.d.C.; Luque, G.M.; Vitale, A.M.; Cuasnicu, P.S.; Visconti, P.E.; Krapf, D.; Buffone, M.G. PKA-dependent phosphorylation of LIMK1 and Cofilin is essential for mouse sperm acrosomal exocytosis. Dev. Biol. 2015, 405, 237–249. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Koshimizu, U.; Abe, H.; Obinata, T.; Nakamura, T. Functional involvement of Xenopus LIM kinases in progression of oocyte maturation. Dev. Biol. 2001, 229, 554–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhu, Y.; Cao, Y.; Wang, Q.; Du, J.; Tian, J.; Liang, Y.; Ma, W. LIM kinase activity is required for microtubule organising centre positioning in mouse oocyte meiosis. Reprod. Fertil. Dev. 2017, 29, 791–804. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.X.; Duan, X.; Song, S.J.; Sun, S.C. LIMK1/2 inhibitor LIMKi 3 suppresses porcine oocyte maturation. PeerJ 2016, 2016, e2553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, X.; Zhang, Y.; Chen, K.L.; Zhang, H.L.; Wu, L.L.; Liu, H.L.; Wang, Z.B.; Sun, S.C. The small GTPase RhoA regulates the LIMK1/2-cofilin pathway to modulate cytoskeletal dynamics in oocyte meiosis. J. Cell. Physiol. 2018, 233, 6088–6097. [Google Scholar] [CrossRef] [PubMed]
- Bourdais, A.; Dehapiot, B.; Halet, G. Cofilin regulates actin network homeostasis and microvilli length in mouse oocytes. J. Cell Sci. 2021, 134, jcs259237. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Fanti, J.A.; Macedo, D.P.; Larsen, M. LIM kinase regulation of cytoskeletal dynamics is required for salivary gland branching morphogenesis. Mol. Biol. Cell 2014, 25, 2393–2407. [Google Scholar] [CrossRef] [PubMed]
- Dimri, M.; Bilogan, C.; Pierce, L.X.; Naegele, G.; Vasanji, A.; Gibson, I.; McClendon, A.; Tae, K.; Sakaguchi, T.F. Three-dimensional structural analysis reveals a Cdk5-mediated kinase cascade regulating hepatic biliary network branching in zebrafish. Development 2017, 144, 2595–2605. [Google Scholar] [CrossRef] [Green Version]
- Machlus, K.R.; Thon, J.N.; Italiano, J.E. Interpreting the developmental dance of the megakaryocyte: A review of the cellular and molecular processes mediating platelet formation. Br. J. Haematol. 2014, 165, 227–236. [Google Scholar] [CrossRef]
- Heib, T.; Hermanns, H.M.; Manukjan, G.; Englert, M.; Kusch, C.; Becker, I.C.; Gerber, A.; Wackerbarth, L.M.; Burkard, P.; Dandekar, T.; et al. RhoA/Cdc42 signaling drives cytoplasmic maturation but not endomitosis in megakaryocytes. Cell Rep. 2021, 35, 109102. [Google Scholar] [CrossRef]
- Kauskot, A.; Poirault-Chassac, S.; Adam, F.; Muczynski, V.; Aymé, G.; Casari, C.; Bordet, J.-C.; Soukaseum, C.; Rothschild, C.; Proulle, V.; et al. LIM kinase/cofilin dysregulation promotes macrothrombocytopenia in severe von Willebrand disease-type 2B. JCI Insight 2016, 1, e88643. [Google Scholar] [CrossRef]
- Becker, I.C.; Scheller, I.; Wackerbarth, L.M.; Beck, S.; Heib, T.; Aurbach, K.; Manukjan, G.; Gross, C.; Spindler, M.; Nagy, Z.; et al. Actin/microtubule crosstalk during platelet biogenesis in mice is critically regulated by Twinfilin1 and Cofilin1. Blood Adv. 2020, 4, 2124–2134. [Google Scholar] [CrossRef] [PubMed]
- Kosoff, R.E.; Aslan, J.E.; Kostyak, J.C.; Dulaimi, E.; Chow, H.Y.; Prudnikova, T.Y.; Radu, M.; Kunapuli, S.P.; McCarty, O.J.T.; Chernoff, J. Pak2 restrains endomitosis during megakaryopoiesis and alters cytoskeleton organization. Blood 2015, 125, 2995–3005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geue, S.; Aurbach, K.; Manke, M.C.; Manukjan, G.; Münzer, P.; Stegner, D.; Brähler, C.; Walker-Allgaier, B.; Märklin, M.; Borst, C.E.; et al. Pivotal role of PDK1 in megakaryocyte cytoskeletal dynamics and polarization during platelet biogenesis. Blood 2019, 134, 1847–1858. [Google Scholar] [CrossRef] [PubMed]
- Kapur, R.; Shi, J.; Ghosh, J.; Munugalavadla, V.; Sims, E.; Martin, H.; Wei, L.; Mali, R.S. ROCK1 via LIM kinase regulates growth, maturation and actin based functions in mast cells. Oncotarget 2016, 7, 16936–16947. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Ghosh, P.; Prasad, R.; Ghosh, A.; Das, K.; Roy, A.; Mallik, S.; Sinha, D.K.; Sen, P. MAP Kinase driven actomyosin rearrangement is a crucial regulator of monocyte to macrophage differentiation. Cell. Signal. 2020, 73, 109691. [Google Scholar] [CrossRef]
- Diagouraga, B.; Grichine, A.; Fertin, A.; Wang, J.; Khochbin, S.; Sadoul, K. Motor-driven marginal band coiling promotes cell shape change during platelet activation. J. Cell Biol. 2014, 204, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Antonipillai, J.; Rigby, S.; Bassler, N.; Peter, K.; Bernard, O. Pharmacological inhibition of LIM kinase pathway impairs platelets functionality and facilitates thrombolysis. Exp. Cell Res. 2019, 382, 111458. [Google Scholar] [CrossRef] [PubMed]
- Antonipillai, J.; Mittelstaedt, K.; Rigby, S.; Bassler, N.; Bernard, O. LIM kinase 2 (LIMK2) may play an essential role in platelet function. Exp. Cell Res. 2020, 388, 111822. [Google Scholar] [CrossRef]
- Estevez, B.; Stojanovic-Terpo, A.; Delaney, M.K.; O’Brien, K.A.; Berndt, M.C.; Ruan, C.; Du, X.; O’Brien, K.A.; Berndt, M.C.; Ruan, C.; et al. LIM Kinase-1 selectively promotes glycoprotein Ib-IX-mediated TXA2 synthesis, platelet activation, and thrombosis. Blood 2013, 121, 4586–45094. [Google Scholar] [CrossRef] [Green Version]
- Bettegazzi, B.; Bellani, S.; Cattaneo, S.; Codazzi, F.; Grohovaz, F.; Zacchetti, D. Gα13 Contributes to LPS-Induced Morphological Alterations and Affects Migration of Microglia. Mol. Neurobiol. 2021, 58, 6397–6414. [Google Scholar] [CrossRef]
- Duvall, M.G.; Fuhlbrigge, M.E.; Reilly, R.B.; Walker, K.H.; Kılıç, A.; Levy, B.D. Human NK Cell Cytoskeletal Dynamics and Cytotoxicity Are Regulated by LIM Kinase. J. Immunol. 2020, 205, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Salah, E.; Chatterjee, D.; Beltrami, A.; Tumber, A.; Preuss, F.; Canning, P.; Chaikuad, A.; Knaus, P.; Knapp, S.; Bullock, A.N.; et al. Lessons from LIMK1 enzymology and their impact on inhibitor design. Biochem. J. 2019, 476, 3197–3209. [Google Scholar] [CrossRef] [PubMed]
- Cuberos, H.; Vallée, B.; Vourc’h, P.; Tastet, J.; Andres, C.R.; Bénédetti, H. Roles of LIM kinases in central nervous system function and dysfunction. FEBS Lett. 2015, 589, 3795–3806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Eddy, R.; Condeelis, J. The cofilin pathway in breast cancer invasion and metastasis. Nat. Rev. Cancer 2007, 7, 429–440. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Kim, S.W.; Cho, M.C. The Role of LIM Kinase in the Male Urogenital System. Cells 2022, 11, 78. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribba, A.-S.; Fraboulet, S.; Sadoul, K.; Lafanechère, L. The Role of LIM Kinases during Development: A Lens to Get a Glimpse of Their Implication in Pathologies. Cells 2022, 11, 403. https://doi.org/10.3390/cells11030403
Ribba A-S, Fraboulet S, Sadoul K, Lafanechère L. The Role of LIM Kinases during Development: A Lens to Get a Glimpse of Their Implication in Pathologies. Cells. 2022; 11(3):403. https://doi.org/10.3390/cells11030403
Chicago/Turabian StyleRibba, Anne-Sophie, Sandrine Fraboulet, Karin Sadoul, and Laurence Lafanechère. 2022. "The Role of LIM Kinases during Development: A Lens to Get a Glimpse of Their Implication in Pathologies" Cells 11, no. 3: 403. https://doi.org/10.3390/cells11030403
APA StyleRibba, A. -S., Fraboulet, S., Sadoul, K., & Lafanechère, L. (2022). The Role of LIM Kinases during Development: A Lens to Get a Glimpse of Their Implication in Pathologies. Cells, 11(3), 403. https://doi.org/10.3390/cells11030403