
Glucose Transporter 9 (GLUT9) Plays an Important Role in the Placental Uric Acid Transport System

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Placental Tissue
2.2. Antibodies
2.3. Immunofluorescence
2.4. Immunohistochemistry
2.5. G9KO Mice
2.6. Statistical Analysis
3. Results
3.1. Localization of GLUT9 Isoforms in Human Placenta
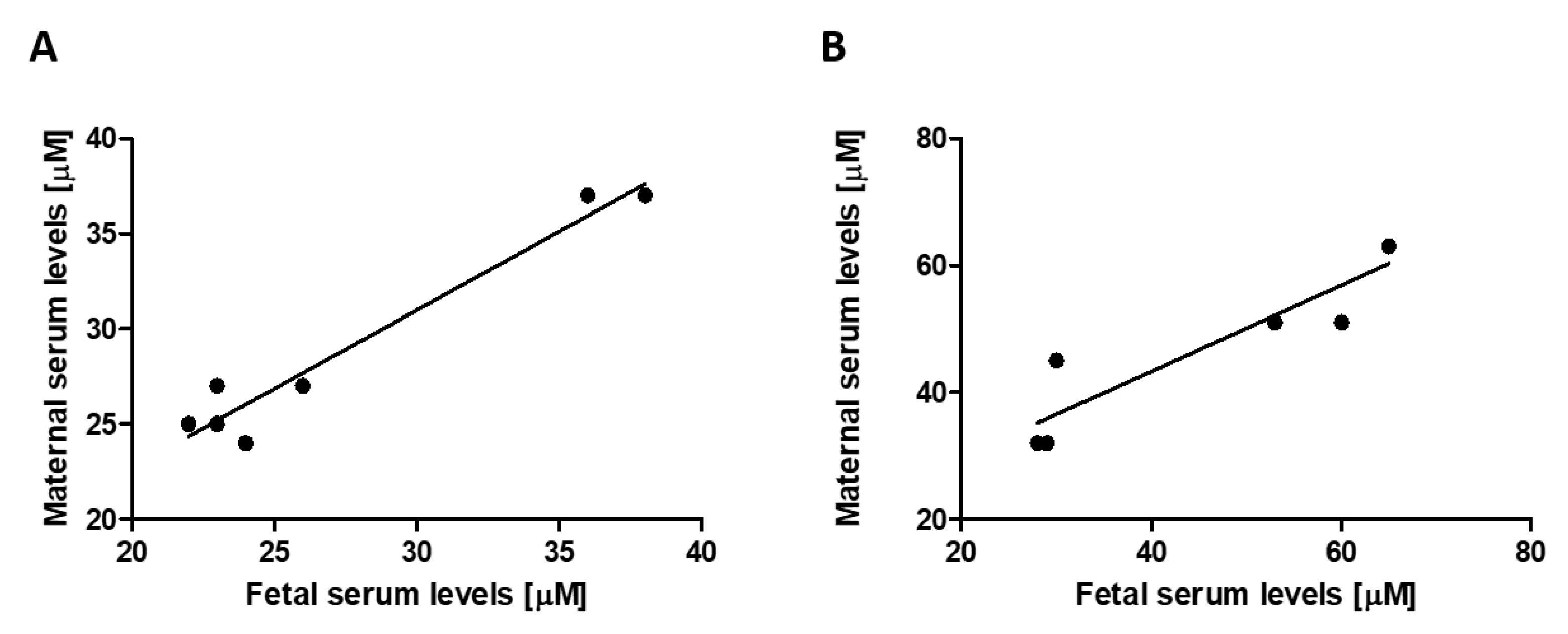
3.2. Transplacental Uric Acid Transport
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bainbridge, S.A.; Roberts, J.M. Uric acid as a pathogenic factor in preeclampsia. Placenta 2008, 29, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.C.; Brown, M.A. Could uric acid have a pathogenic role in pre-eclampsia? Nat. Rev. Nephrol. 2010, 6, 744–748. [Google Scholar] [CrossRef]
- Hediger, M.A.; Johnson, R.J.; Miyazaki, H.; Endou, H. Molecular physiology of urate transport. Physiology 2005, 20, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Bo, S.; Cavallo-Perin, P.; Gentile, L.; Repetti, E.; Pagano, G. Hypouricemia and hyperuricemia in type 2 diabetes: Two different phenotypes. Eur. J. Clin. Investig. 2001, 31, 318–321. [Google Scholar] [CrossRef]
- Feig, D.I.; Kang, D.H.; Nakagawa, T.; Mazzali, M.; Johnson, R.J. Uric acid and hypertension. Curr. Hypertens. Rep. 2006, 8, 111–115. [Google Scholar] [CrossRef]
- Hjortnaes, J.; Algra, A.; Olijhoek, J.; Huisman, M.; Jacobs, J.; van der Graaf, Y.; Visseren, F. Serum uric acid levels and risk for vascular diseases in patients with metabolic syndrome. J. Rheumatol. 2007, 34, 1882–1887. [Google Scholar]
- Onat, A.; Uyarel, H.; Hergenç, G.; Karabulut, A.; Albayrak, S.; Sarı, I.; Yazıcı, M.; Keleş, I. Serum uric acid is a determinant of metabolic syndrome in a population-based study. Am. J. Hypertens. 2006, 19, 1055–1062. [Google Scholar] [CrossRef] [Green Version]
- Panoulas, V.F.; Milionis, H.J.; Douglas, K.M.; Nightingale, P.; Kita, M.D.; Klocke, R.; Elisaf, M.S.; Kitas, G.D. Association of serum uric acid with cardiovascular disease in rheumatoid arthritis. Rheumatology 2007, 46, 1466–1470. [Google Scholar] [CrossRef] [Green Version]
- Rathmann, W.; Haastert, B.; Icks, A.; Giani, G.; Roseman, J.M. Ten-year change in serum uric acid and its relation to changes in other metabolic risk factors in young black and white adults: The CARDIA study. Eur. J. Epidemiol. 2007, 22, 439–445. [Google Scholar] [CrossRef]
- Powers, R.W.; Bodnar, L.M.; Ness, R.B.; Cooper, K.M.; Gallaher, M.J.; Frank, M.P.; Daftary, A.R.; Roberts, J.M. Uric acid concentrations in early pregnancy among preeclamptic women with gestational hyperuricemia at delivery. Am. J. Obstet. Gynecol. 2006, 194, 160.e1–160.e8. [Google Scholar] [CrossRef]
- Chen, Y.; Ou, W.; Lin, D.; Lin, M.; Huang, X.; Ni, S.; Chen, S.; Yong, J.; O’Gara, M.C.; Tan, X.; et al. Increased Uric Acid, Gamma-Glutamyl Transpeptidase and Alkaline Phosphatase in Early-Pregnancy Associated With the Development of Gestational Hypertension and Preeclampsia. Front. Cardiovasc. Med. 2021, 8, 756140. [Google Scholar] [CrossRef]
- Voto, L.S.; Illia, R.; Darbon-Grosso, H.A.; Imaz, F.U.; Margulies, M. Uric acid levels: A useful index of the severity of preeclampsia and perinatal prognosis. J. Perinat. Med. 1988, 16, 123–126. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.M.; Bodnar, L.M.; Lain, K.Y.; Hubel, C.A.; Markovic, N.; Ness, R.B.; Powers, R.W. Uric acid is as important as proteinuria in identifying fetal risk in women with gestational hypertension. Hypertension 2005, 46, 1263–1269. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Xiong, X.; Fraser, W.D.; Luo, Z.C. Association of uric acid with progression to preeclampsia and development of adverse conditions in gestational hypertensive pregnancies. Am. J. Hypertens. 2012, 25, 711–717. [Google Scholar] [CrossRef]
- Livingston, J.R.; Payne, B.; Brown, M.; Roberts, J.M.; Côté, A.M.; Magee, L.A.; von Dadelszen, P.; PIERS Study Group. Uric Acid as a predictor of adverse maternal and perinatal outcomes in women hospitalized with preeclampsia. J. Obstet. Gynaecol. Can. 2014, 36, 870–877. [Google Scholar] [CrossRef]
- Bibert, S.; Hess, S.K.; Firsov, D.; Thorens, B.; Geering, K.; Horisberger, J.D.; Bonny, O. Mouse GLUT9: Evidences for a urate uniporter. Am. J. Physiol.-Ren. Physiol. 2009, 297, F612–F619. [Google Scholar] [CrossRef] [Green Version]
- Augustin, R.; Carayannopoulos, M.O.; Dowd, L.O.; Phay, J.E.; Moley, J.F.; Moley, K.H. Identification and characterization of human glucose transporter-like protein-9 (GLUT9): Alternative splicing alters trafficking. J. Biol. Chem. 2004, 279, 16229–16236. [Google Scholar] [CrossRef] [Green Version]
- Phay, J.E.; Hussain, H.B.; Moley, J.F. Cloning and expression analysis of a novel member of the facilitative glucose transporter family, SLC2A9 (GLUT9). Genomics 2000, 66, 217–220. [Google Scholar] [CrossRef]
- Stieger, B.; Hagenbuch, B.; Landmann, L.; Höchli, M.; Schroeder, A.; Meier, P.J. In situ localization of the hepatocytic Na+/Taurocholate cotransporting polypeptide in rat liver. Gastroenterology 1994, 107, 1781–1787. [Google Scholar] [CrossRef]
- Preitner, F.; Bonny, O.; Laverrière, A.; Rotman, S.; Firsov, D.; da Costa, A.; Metref, S.; Thorens, B. Glut9 is a major regulator of urate homeostasis and its genetic inactivation induces hyperuricosuria and urate nephropathy. Proc. Natl. Acad. Sci. USA 2009, 106, 15501–15506. [Google Scholar] [CrossRef] [Green Version]
- Lüscher, B.; Surbek, D.; Clemençon, B.; Huang, X.; Albrecht, C.; Marini, C.; Hediger, M.; Baumann, M.U. Different Pharmacological Properties of GLUT9a and GLUT9b: Potential Implications in Preeclampsia. Cell. Physiol. Biochem. 2019, 53, 508–517. [Google Scholar] [PubMed]
- Kimura, T.; Takahashi, M.; Yan, K.; Sakurai, H. Expression of SLC2A9 isoforms in the kidney and their localization in polarized epithelial cells. PLoS ONE 2014, 9, e84996. [Google Scholar] [CrossRef] [Green Version]
- Hagos, Y.; Stein, D.; Ugele, B.; Burckhardt, G.; Bahn, A. Human renal organic anion transporter 4 operates as an asymmetric urate transporter. J. Am. Soc. Nephrol. 2007, 18, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Staff, A.C.; Johnsen, G.M.; Dechend, R.; Redman, C.W.G. Preeclampsia and uteroplacental acute atherosis: Immune and inflammatory factors. J. Reprod. Immunol. 2014, 101–102, 120–126. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lüscher, B.P.; Albrecht, C.; Stieger, B.; Surbek, D.V.; Baumann, M.U. Glucose Transporter 9 (GLUT9) Plays an Important Role in the Placental Uric Acid Transport System. Cells 2022, 11, 633. https://doi.org/10.3390/cells11040633
Lüscher BP, Albrecht C, Stieger B, Surbek DV, Baumann MU. Glucose Transporter 9 (GLUT9) Plays an Important Role in the Placental Uric Acid Transport System. Cells. 2022; 11(4):633. https://doi.org/10.3390/cells11040633
Chicago/Turabian StyleLüscher, Benjamin P., Christiane Albrecht, Bruno Stieger, Daniel V. Surbek, and Marc U. Baumann. 2022. "Glucose Transporter 9 (GLUT9) Plays an Important Role in the Placental Uric Acid Transport System" Cells 11, no. 4: 633. https://doi.org/10.3390/cells11040633
APA StyleLüscher, B. P., Albrecht, C., Stieger, B., Surbek, D. V., & Baumann, M. U. (2022). Glucose Transporter 9 (GLUT9) Plays an Important Role in the Placental Uric Acid Transport System. Cells, 11(4), 633. https://doi.org/10.3390/cells11040633