
Proximity Ligation Mapping of Microcephaly Associated SMPD4 Shows Association with Components of the Nuclear Pore Membrane



Abstract
:1. Introduction
2. Materials and Methods
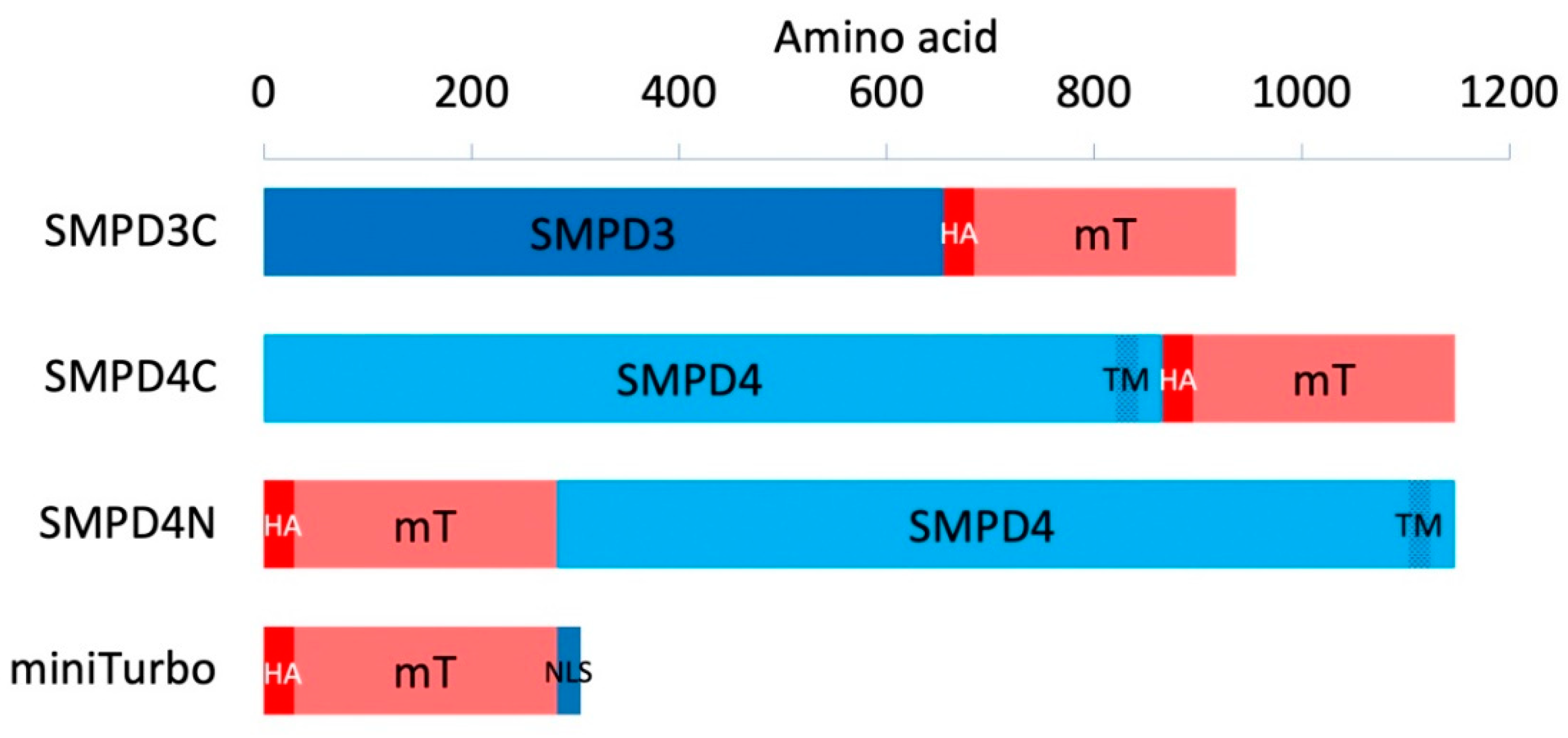
2.1. Cloning, Transformation, Bacterial Culture, and Sequencing
2.2. HEK293T Culture, Transfection, and Biotinylation
2.3. Immunofluorescence
2.4. Western Blotting
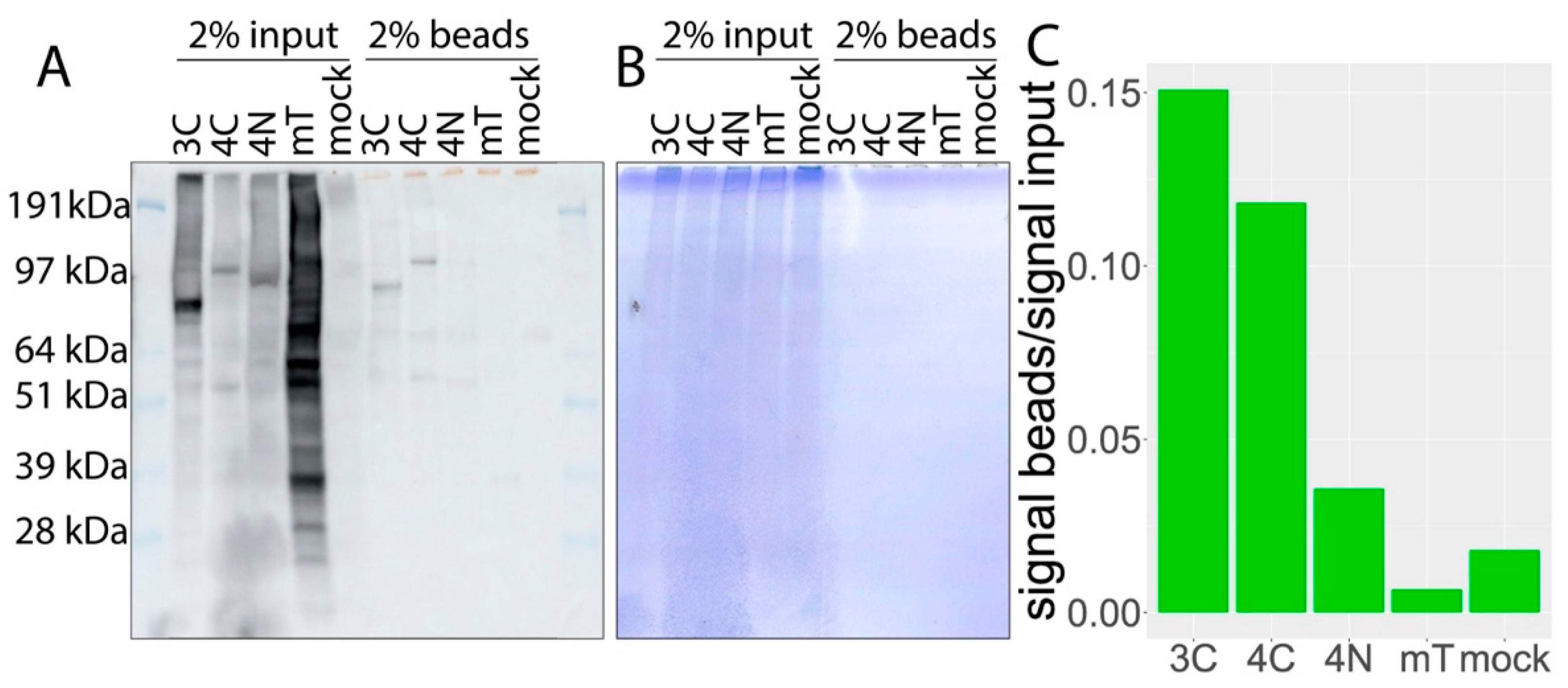
2.5. Biotinylated Protein Pulldown
2.6. Mass Spectrometry
3. Results
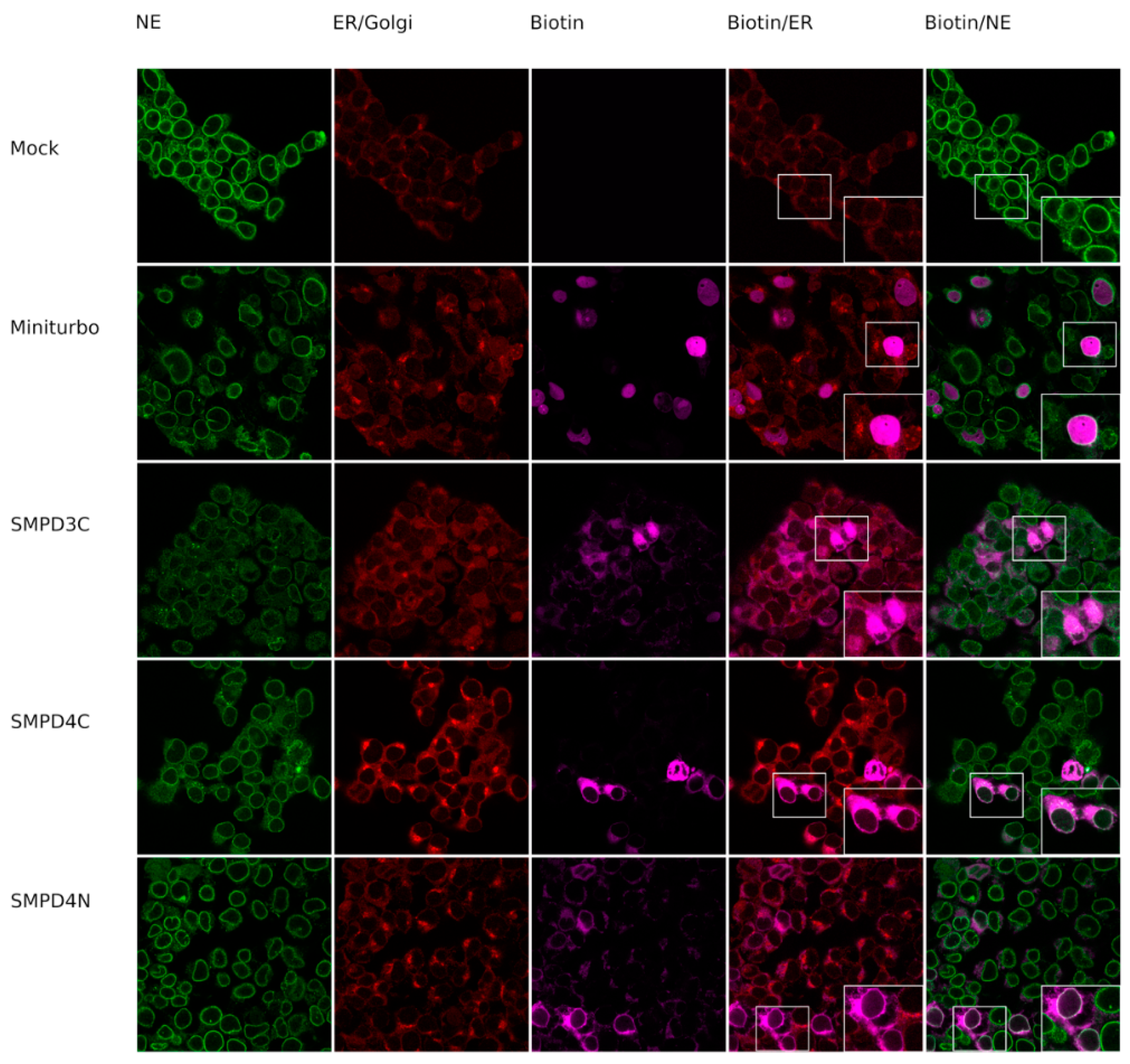
3.1. Biotin-Tagged Proteins Localize at the Nuclear Membrane
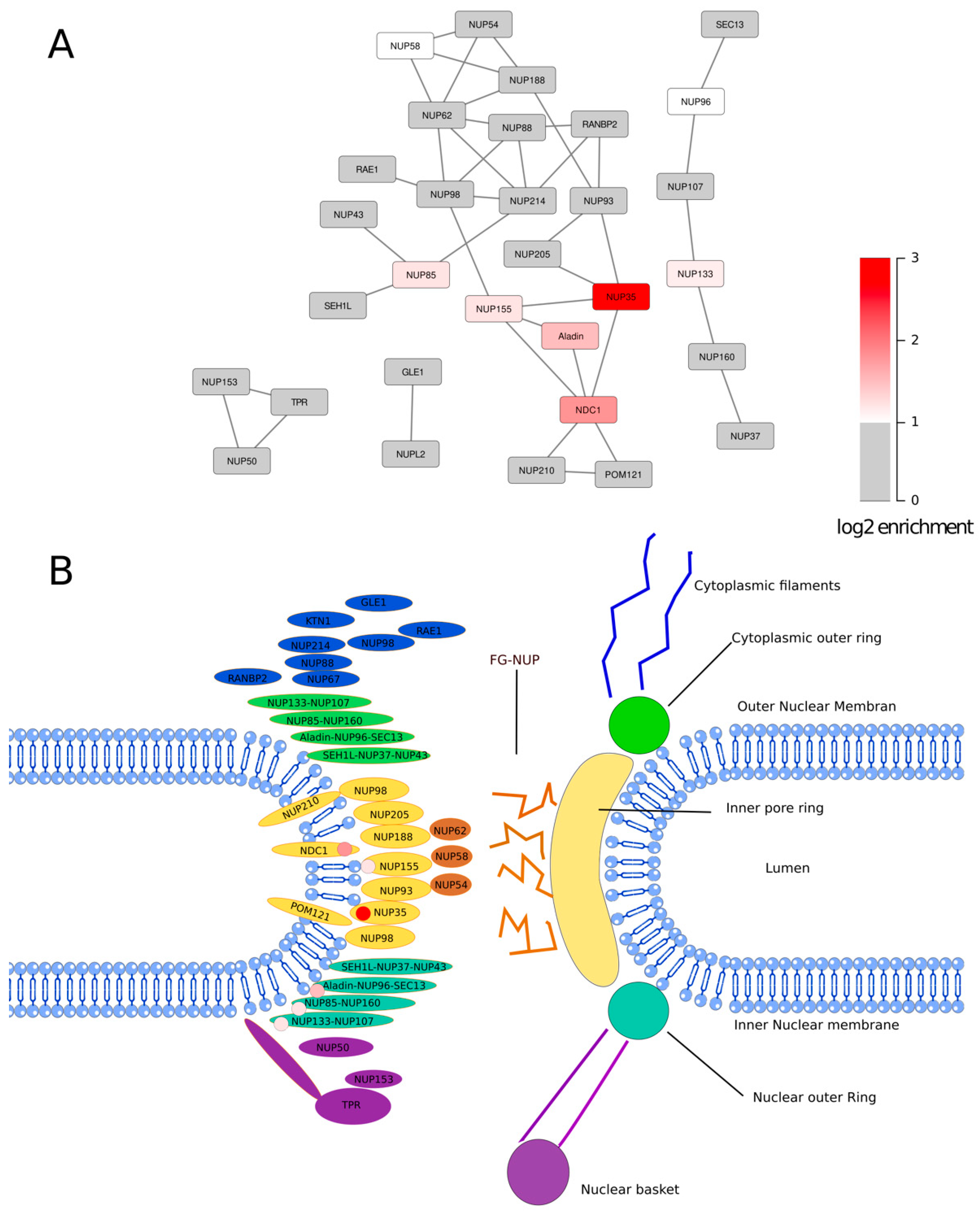
3.2. The SMPD4 Interactome
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody/Reagent | Company | Dilution |
|---|---|---|
| Streptavidin, Alexa Fluor™ 700 conjugate | Invitrogen | 1:280 |
| Streptavidin-HRP Conjugate | Perkin Elmer | 1:5000 |
| anti-BCAP31 (ab237485) | Abcam | 1:600 |
| mAb414 (ab24609) | Abcam | 1:300 |
| Goat anti-Mouse or anti-Rabbit IgG (H + L) Cross-Adsorbed Secondary Antibody, Texas Red or Alexa 488 | ThermoFisher | 1:1000 |
Appendix B
| NotI SMPD3 forward | CTTGCGGCCGCATGGTTTTGTACACGACCCCCTTTCCT |
| NotI SMPD3 reverse | GTGGCGGCCGCTGCCTCCTCCTCCCCCGAAG |
| NotI SMPD4 forward | CTTGCGGCCGCATGACGACTTTCGGCGCCGTG |
| NotI SMPD4 reverse | GTGGCGGCCGCGGGCTGGTGCAGCTTCCCC |
| EcoRI SMPD4 forward | CTTGAATTCATGACGACTTTCGGCGCCGTG |
| EcoRI SMPD4 reverse | GTGGAATTCTCAGGGCTGGTGCAGCTTCCC |
References
- Beck, M.; Hurt, E. The Nuclear Pore Complex: Understanding Its Function through Structural Insight. Nat. Rev. Mol. Cell Biol. 2017, 18, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.H.; Hoelz, A. The Structure of the Nuclear Pore Complex (An Update). Annu. Rev. Biochem. 2019, 88, 725–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platani, M.; Santarella-Mellwig, R.; Posch, M.; Walczak, R.; Swedlow, J.R.; Mattaj, I.W. The Nup107-160 Nucleoporin Complex Promotes Mitotic Events via Control of the Localization State of the Chromosome Passenger Complex. Mol. Biol. Cell 2009, 20, 5260–5275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, P.; Wang, Y.; Wei, J.-H.; van Deursen, J.; Yu, H.; Malureanu, L.; Dasso, M.; Forbes, D.J.; Levy, D.E.; Seemann, J.; et al. Nucleoporin Levels Regulate Cell Cycle Progression and Phase-Specific Gene Expression. Dev. Cell 2008, 15, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Jiménez, N.; Pradillo, M. The Role of the Nuclear Envelope in the Regulation of Chromatin Dynamics during Cell Division. J. Exp. Bot. 2020, 71, 5148–5159. [Google Scholar] [CrossRef]
- Sakuma, S.; Raices, M.; Borlido, J.; Guglielmi, V.; Zhu, E.Y.S.; D’Angelo, M.A. Inhibition of Nuclear Pore Complex Formation Selectively Induces Cancer Cell Death. Cancer Discov. 2021, 11, 176–193. [Google Scholar] [CrossRef]
- Otsuka, S.; Ellenberg, J. Mechanisms of Nuclear Pore Complex Assembly—Two Different Ways of Building One Molecular Machine. FEBS Lett. 2018, 592, 475–488. [Google Scholar] [CrossRef]
- Corcoran, C.A.; He, Q.; Ponnusamy, S.; Ogretmen, B.; Huang, Y.; Sheikh, M.S. Neutral Sphingomyelinase-3 Is a DNA Damage and Nongenotoxic Stress-Regulated Gene That Is Deregulated in Human Malignancies. Mol. Cancer Res. 2008, 6, 795–807. [Google Scholar] [CrossRef] [Green Version]
- Moylan, J.S.; Smith, J.D.; Wolf Horrell, E.M.; McLean, J.B.; Deevska, G.M.; Bonnell, M.R.; Nikolova-Karakashian, M.N.; Reid, M.B. Neutral Sphingomyelinase-3 Mediates TNF-Stimulated Oxidant Activity in Skeletal Muscle. Redox Biol. 2014, 2, 910–920. [Google Scholar] [CrossRef] [Green Version]
- Krut, O.; Wiegmann, K.; Kashkar, H.; Yazdanpanah, B.; Krönke, M. Novel Tumor Necrosis Factor-Responsive Mammalian Neutral Sphingomyelinase-3 Is a C-Tail-Anchored Protein. J. Biol. Chem. 2006, 281, 13784–13793. [Google Scholar] [CrossRef] [Green Version]
- Atilla-Gokcumen, G.E.; Muro, E.; Relat-Goberna, J.; Sasse, S.; Bedigian, A.; Coughlin, M.L.; Garcia-Manyes, S.; Eggert, U.S. Dividing Cells Regulate Their Lipid Composition and Localization. Cell 2014, 156, 428–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magini, P.; Smits, D.J.; Vandervore, L.; Schot, R.; Columbaro, M.; Kasteleijn, E.; van der Ent, M.; Palombo, F.; Lequin, M.H.; Dremmen, M.; et al. Loss of SMPD4 Causes a Developmental Disorder Characterized by Microcephaly and Congenital Arthrogryposis. Am. J. Hum. Genet. 2019, 105, 689–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hein, M.Y.; Hubner, N.C.; Poser, I.; Cox, J.; Nagaraj, N.; Toyoda, Y.; Gak, I.A.; Weisswange, I.; Mansfeld, J.; Buchholz, F.; et al. A Human Interactome in Three Quantitative Dimensions Organized by Stoichiometries and Abundances. Cell 2015, 163, 712–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, G.D.; Coyaud, É.; Gonçalves, J.; Mojarad, B.A.; Liu, Y.; Wu, Q.; Gheiratmand, L.; Comartin, D.; Tkach, J.M.; Cheung, S.W.T.; et al. A Dynamic Protein Interaction Landscape of the Human Centrosome-Cilium Interface. Cell 2015, 163, 1484–1499. [Google Scholar] [CrossRef] [Green Version]
- Airola, M.V.; Shanbhogue, P.; Shamseddine, A.A.; Guja, K.E.; Senkal, C.E.; Maini, R.; Bartke, N.; Wu, B.X.; Obeid, L.M.; Garcia-Diaz, M.; et al. Structure of Human NSMase2 Reveals an Interdomain Allosteric Activation Mechanism for Ceramide Generation. Proc. Natl. Acad. Sci. USA 2017, 114, E5549–E5558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branon, T.C.; Bosch, J.A.; Sanchez, A.D.; Udeshi, N.D.; Svinkina, T.; Carr, S.A.; Feldman, J.L.; Perrimon, N.; Ting, A.Y. Efficient Proximity Labeling in Living Cells and Organisms with TurboID. Nat. Biotechnol. 2018, 36, 880–887. [Google Scholar] [CrossRef]
- Schwertman, P.; Lagarou, A.; Dekkers, D.H.W.; Raams, A.; van der Hoek, A.C.; Laffeber, C.; Hoeijmakers, J.H.J.; Demmers, J.A.A.; Fousteri, M.; Vermeulen, W.; et al. UV-Sensitive Syndrome Protein UVSSA Recruits USP7 to Regulate Transcription-Coupled Repair. Nat. Genet. 2012, 44, 598–602. [Google Scholar] [CrossRef]
- Cox, J.; Matic, I.; Hilger, M.; Nagaraj, N.; Selbach, M.; Olsen, J.V.; Mann, M. A Practical Guide to the MaxQuant Computational Platform for SILAC-Based Quantitative Proteomics. Nat. Protoc. 2009, 4, 698–705. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Marchesini, N.; Osta, W.; Bielawski, J.; Luberto, C.; Obeid, L.M.; Hannun, Y.A. Role for Mammalian Neutral Sphingomyelinase 2 in Confluence-Induced Growth Arrest of MCF7 Cells. J. Biol. Chem. 2004, 279, 25101–25111. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, K.; Tomiuk, S.; Wolff, G.; Stoffel, W. Cloning and Characterization of the Mammalian Brain-Specific, Mg2+-Dependent Neutral Sphingomyelinase. Proc. Natl. Acad. Sci. USA 2000, 97, 5895–5900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishihama, Y.; Oda, Y.; Tabata, T.; Sato, T.; Nagasu, T.; Rappsilber, J.; Mann, M. Exponentially Modified Protein Abundance Index (EmPAI) for Estimation of Absolute Protein Amount in Proteomics by the Number of Sequenced Peptides per Protein. Mol. Cell. Proteom. 2005, 4, 1265–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The UniProt Consortium. UniProt: The Universal Protein Knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively Expanding the Structural Coverage of Protein-Sequence Space with High-Accuracy Models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Hawryluk-Gara, L.A.; Platani, M.; Santarella, R.; Wozniak, R.W.; Mattaj, I.W. Nup53 Is Required for Nuclear Envelope and Nuclear Pore Complex Assembly. Mol. Biol. Cell. 2008, 19, 1753–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huttlin, E.L.; Bruckner, R.J.; Navarrete-Perea, J.; Cannon, J.R.; Baltier, K.; Gebreab, F.; Gygi, M.P.; Thornock, A.; Zarraga, G.; Tam, S.; et al. Dual Proteome-Scale Networks Reveal Cell-Specific Remodeling of the Human Interactome. Cell 2021, 184, 3022–3040.e28. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, B.; Schooley, A.; Sachdev, R.; Eisenhardt, N.; Schneider, A.M.; Sieverding, C.; Madlung, J.; Gerken, U.; Macek, B.; Antonin, W. Dimerization and Direct Membrane Interaction of Nup53 Contribute to Nuclear Pore Complex Assembly. EMBO J. 2012, 31, 4072–4084. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.-C.; Baboo, S.; Lindsay, C.; Brusman, L.; Martinez-Bartolomé, S.; Tapia, O.; Zhang, X.; Yates, J.R.; Gerace, L. Identification of New Transmembrane Proteins Concentrated at the Nuclear Envelope Using Organellar Proteomics of Mesenchymal Cells. Nucleus 2019, 10, 126–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peeters, B.W.A.; Piët, A.C.A.; Fornerod, M. Generating Membrane Curvature at the Nuclear Pore: A Lipid Point of View. Cells 2022, 11, 469. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piët, A.C.A.; Post, M.; Dekkers, D.; Demmers, J.A.A.; Fornerod, M. Proximity Ligation Mapping of Microcephaly Associated SMPD4 Shows Association with Components of the Nuclear Pore Membrane. Cells 2022, 11, 674. https://doi.org/10.3390/cells11040674
Piët ACA, Post M, Dekkers D, Demmers JAA, Fornerod M. Proximity Ligation Mapping of Microcephaly Associated SMPD4 Shows Association with Components of the Nuclear Pore Membrane. Cells. 2022; 11(4):674. https://doi.org/10.3390/cells11040674
Chicago/Turabian StylePiët, Alexandra C. A., Marco Post, Dick Dekkers, Jeroen A. A. Demmers, and Maarten Fornerod. 2022. "Proximity Ligation Mapping of Microcephaly Associated SMPD4 Shows Association with Components of the Nuclear Pore Membrane" Cells 11, no. 4: 674. https://doi.org/10.3390/cells11040674
APA StylePiët, A. C. A., Post, M., Dekkers, D., Demmers, J. A. A., & Fornerod, M. (2022). Proximity Ligation Mapping of Microcephaly Associated SMPD4 Shows Association with Components of the Nuclear Pore Membrane. Cells, 11(4), 674. https://doi.org/10.3390/cells11040674