
G-Protein Coupled Receptor Dysregulation May Play Roles in Severe Preeclampsia—A Weighted Gene Correlation Network Analysis of Placental Gene Expression Profile


Abstract
:1. Introduction
2. Materials and Methods
2.1. Population and Evaluation of Gene Expression
2.2. Identification of Possible Molecular Targets through WGCNA
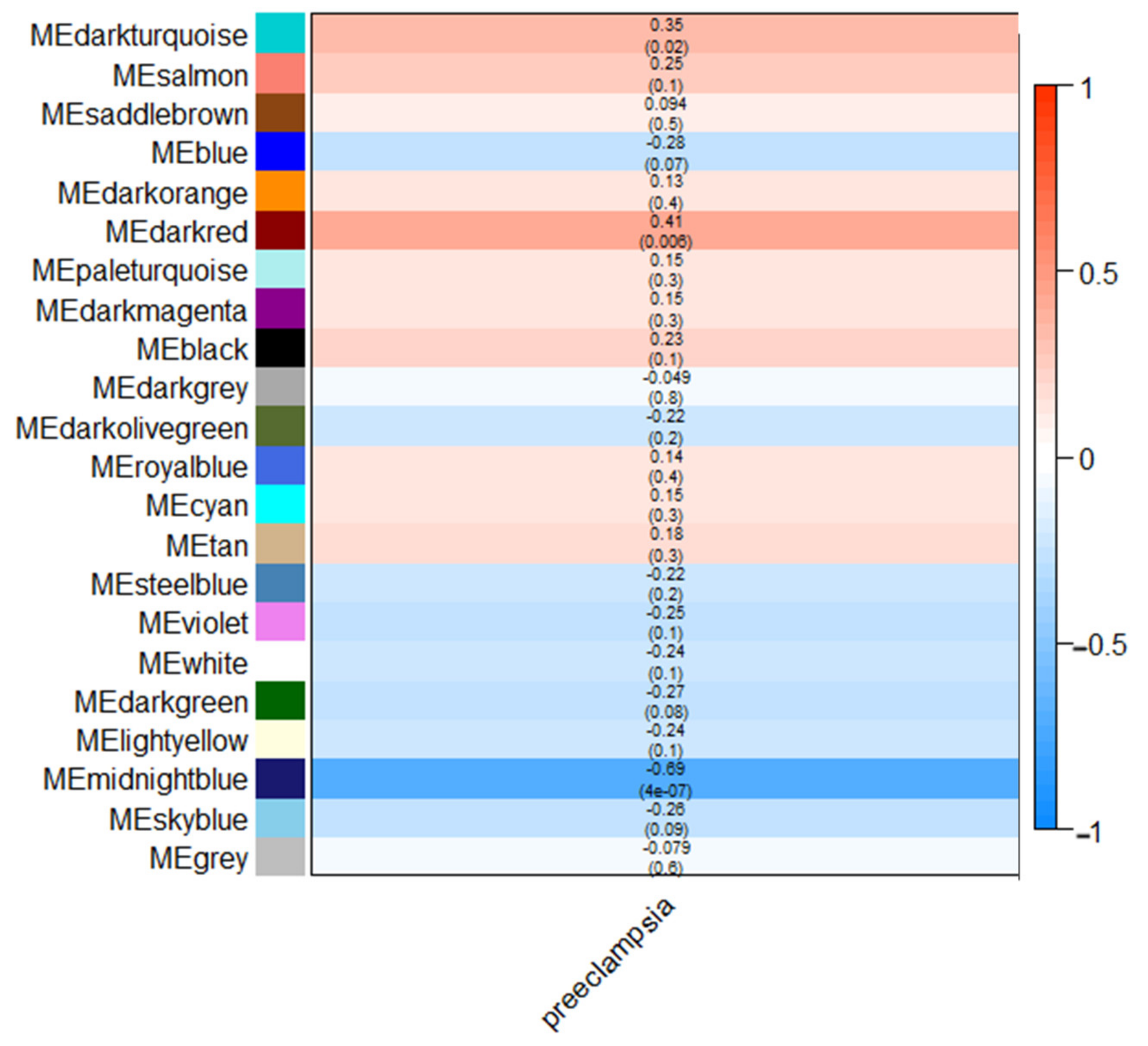
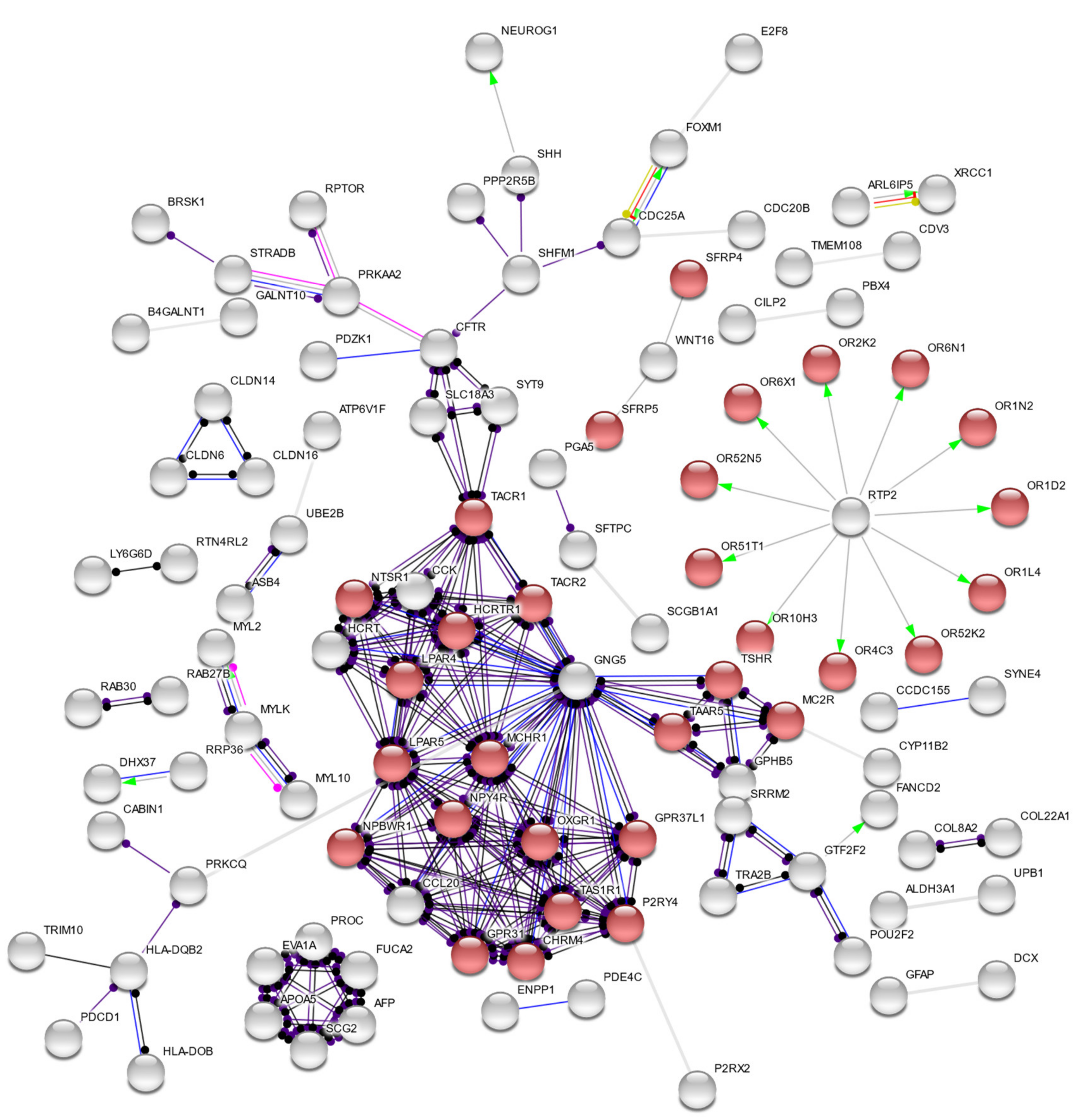
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Philippine Obstetrics and Gynecologic Society. Clinical Practice Guidelines on Hypertension in Pregnancy; Philippine Obstetrics and Gynecologic Society: Quezon City, Philippines, 2015. [Google Scholar]
- Department of Health—National Epidemiology Center. The 2013 Philippine Health Statistics; Epidemiology Bureau: Manila, Philippines, 2014. Available online: http://www.dohgovph/sites/default/files/publications/2013PHScompressed_0.pdf (accessed on 10 March 2020).
- Romero, R.; Chaiworapongsa, T. Preeclampsia: A link between trophoblast dysregulation and an antiangiogenic state. J. Clin. Investig. 2013, 123, 2775–2777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staff, A.C. The two-stage placental model of preeclampsia: An update. J. Reprod. Immunol. 2019, 134–135, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Herting, A. Vascular pathology in hypertensive albuminuric toxemias of pregnancy. Clinics 1945, 4, 602. [Google Scholar]
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia: Pathophysiology, challenges, and perspectives. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef]
- Sitras, V.; Paulssen, R.; Grønaas, H.; Leirvik, J.A.; Hanssen, T.; Vårtun, Å.; Acharya, G. Differential Placental Gene Expression in Severe Preeclampsia. Placenta 2009, 30, 424–433. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- McGuane, J.T.; Conrad, K.P. GPCRs as potential therapeutic targets in preeclampsia. Drug Discov. Today Dis. Model. 2012, 9, e119–e127. [Google Scholar] [CrossRef] [Green Version]
- Iwasawa, Y.; Fujii, T.; Nagamatsu, T.; Kawana, K.; Okudaira, S.; Miura, S.; Matsumoto, J.; Tomio, A.; Hyodo, H.; Yamashita, T.; et al. Original Article: Expression of Autotaxin, an Ectoenzyme that Produces Lysophosphatidic Acid, in Human Placenta: Autotaxin in Human Placenta. Am. J. Reprod. Immunol. 2009, 62, 90–95. [Google Scholar] [CrossRef]
- Tokumura, A.; Kanaya, Y.; Miyake, M.; Yamano, S.; Irahara, M.; Fukuzawa, K. Increased production of bioactive lysophosphatidic acid by serum lysophospholipase D in human pregnancy. Biol. Reprod. 2002, 67, 1386–1392. [Google Scholar] [CrossRef] [Green Version]
- Fujii, T.; Nagamatsu, T.; Schust, D.J.; Ichikawa, M.; Kumasawa, K.; Yabe, S.; Iriyama, T.; Hirota, Y.; Osuga, Y.; Aoki, J.; et al. Placental expression of lysophosphatidic acid receptors in normal pregnancy and preeclampsia. Am. J. Reprod. Immunol. 2019, 82, e13176. [Google Scholar] [CrossRef]
- Ichikawa, M.; Nagamatsu, T.; Nishimori, Y.; Fujii, T.; Kumasawa, K.; Iriyama, T.; Osuga, Y.; Fujii, T. Role of placental ATX-LPA system and its pathologic relevance to Hypertensive disorders of pregnancy. Placenta 2019, 87, 64–65. [Google Scholar] [CrossRef]
- Yang, L.; Kraemer, M.; Fang, X.F.; Angel, P.M.; Drake, R.R.; Morris, A.J.; Smyth, S.S. LPA receptor 4 deficiency attenuates experimental atherosclerosis. J. Lipid Res. 2019, 60, 972–980. [Google Scholar] [CrossRef]
- Kozian, D.H.; Evers, A.; Florian, P.; Wonerow, P.; Joho, S.; Nazare, M. Selective non-lipid modulator of LPA5 activity in human platelets. Bioorg. Med. Chem. Lett. 2012, 22, 5239–5243. [Google Scholar] [CrossRef]
- Tamaruya, Y.; Suzuki, M.; Kamura, G.; Kanai, M.; Hama, K.; Shimizu, K.; Aoki, J.; Arai, H.; Shibasaki, M. Identifying Specific Conformations by Using a Carbohydrate Scaffold: Discovery of Subtype-Selective LPA-Receptor Agonists and an Antagonist. Angew. Chem. Int. Ed. 2004, 43, 2834–2837. [Google Scholar] [CrossRef]
- Kano, K.; Matsumoto, H.; Inoue, A.; Yukiura, H.; Kanai, M.; Chun, J.; Ishii, S.; Shimizu, T.; Aoki, J. Molecular mechanism of lysophosphatidic acid-induced hypertensive response. Sci. Rep. 2019, 9, 2662. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.-L.; Vegiraju, S.; Chauhan, M.; Gangula, P.R.R.; Hankins, G.D.V.; Goodrum, L.; Yallampalli, C. Involvement of calcitonin gene-related peptide in control of human fetoplacental vascular tone. Am. J. Physiol. Circ. Physiol. 2004, 286, H230–H239. [Google Scholar] [CrossRef] [Green Version]
- Ariza, A.C.; Bobadilla, N.; Díaz, L.; Avila, E.; Larrea, F.; Halhali, A. Placental gene expression of calcitonin gene-related peptide and nitric oxide synthases in preeclampsia: Effects of magnesium sulfate. Magnes. Res. 2009, 22, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, M.; Balakrishnan, M.; Yallampalli, C. 334: Regulation of CRLR and RAMPs by nitric oxide in endothelial cells. Am. J. Obstet. Gynecol. 2016, 214, S188. [Google Scholar] [CrossRef]
- Dong, Y.-L.; Green, K.E.; Vegiragu, S.; Hankins, G.D.V.; Martin, E.; Chauhan, M.; Thota, C.; Yallampalli, C. Evidence for Decreased Calcitonin Gene-Related Peptide (CGRP) Receptors and Compromised Responsiveness to CGRP of Fetoplacental Vessels in Preeclamptic Pregnancies. J. Clin. Endocrinol. Metab. 2005, 90, 2336–2343. [Google Scholar] [CrossRef] [Green Version]
- Khatun, S.; Kanayama, N.; Belayet, H.M.; Bhuiyan, A.B.; Jahan, S.; Begum, A.; Kobayashi, T.; Terao, T. Increased concentrations of plasma neuropeptide Y in patients with eclampsia and preeclampsia. Am. J. Obstet. Gynecol. 2000, 182, 896–900. [Google Scholar] [CrossRef]
- Paiva, S.P.C.; Veloso, C.A.; Campos, F.F.C.; Carneiro, M.M.; Tilan, J.U.; Wang, H.; Umans, J.G.; Zukowska, Z.; Kitlinska, J. Elevated levels of neuropeptide Y in preeclampsia: A pilot study implicating a role for stress in pathogenesis of the disease. Neuropeptides 2016, 55, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Klinjampa, R.; Sitticharoon, C.; Souvannavong-Vilivong, X.; Sripong, C.; Keadkraichaiwat, I.; Churintaraphan, M.; Chatree, S.; Lertbunnaphong, T. Placental Neuropeptide Y (NPY) and NPY receptors expressions and serum NPY levels in preeclampsia. Exp. Biol. Med. 2019, 244, 380–388. [Google Scholar] [CrossRef] [Green Version]
- Hua, X.; Lundberg, J.M.; Theodorsson-Norheim, E.; Brodin, E. Comparison of cardiovascular and bronchoconstrictor effects of substance P, substance K and other tachykinins. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakol. 1984, 328, 196–201. [Google Scholar] [CrossRef]
- Walsh, D.A.; McWilliams, D.F. Tachykinins and the Cardiovascular System. Curr. Drug Targets 2006, 7, 1031–1042. [Google Scholar] [CrossRef]
- Bovolenta, P.; Esteve, P.; Ruiz, J.M.; Cisneros, E.; Lopez-Rios, J. Beyond Wnt inhibition: New functions of secreted Frizzled-related proteins in development and disease. J. Cell Sci. 2008, 121, 737–746. [Google Scholar] [CrossRef] [Green Version]
- Bafico, A.; Gazit, A.; Pramila, T.; Finch, P.W.; Yaniv, A.; Aaronson, S.A. Interaction of Frizzled Related Protein (FRP) with Wnt Ligands and the Frizzled Receptor Suggests Alternative Mechanisms for FRP Inhibition of Wnt Signaling. J. Biol. Chem. 1999, 274, 16180–16187. [Google Scholar] [CrossRef] [Green Version]
- Lacher, M.D.; Siegenthaler, A.; Jäger, R.; Yan, X.; Hett, S.; Xuan, L.; Saurer, S.; Lareu, R.R.; Dharmarajan, A.M.; Friis, R. Role of DDC-4/sFRP-4, a secreted Frizzled-related protein, at the onset of apoptosis in mammary involution. Cell Death Differ. 2003, 10, 528–538. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, L.; Zhang, L.; Jia, L.; Wang, P.; Gao, Y. Association of Wnt2 and sFRP4 Expression in the Third Trimester Placenta in Women With Severe Preeclampsia. Reprod. Sci. 2013, 20, 981–989. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Zhang, S.; Nakano, H.; Simmons, D.G.; Wang, S.; Kong, S.; Wang, Q.; Shen, L.; Tu, Z.; Wang, W.; et al. A Positive Feedback Loop Involving Gcm1 and Fzd5 Directs Chorionic Branching Morphogenesis in the Placenta. PLoS Biol. 2013, 11, e1001536. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ran, Y.; Ma, Y.; Huang, H.; Chen, Y.; Qi, H. Elevated Serum SFRP5 Levels During Preeclampsia and Its Potential Association with Trophoblast Dysfunction via Wnt/β-Catenin Suppression. Reprod. Sci. 2022, 29, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Bar, H.Y.; Yan, J.; Jones, S.; Brannon, P.M.; West, A.A.; Perry, C.A.; Ganti, A.; Pressman, E.; Devapatla, S.; et al. A higher maternal choline intake among third-trimester pregnant women lowers placental and circulating concentrations of the antiangiogenic factor fms-like tyrosine kinase-1 (sFLT1). FASEB J. 2013, 27, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Gomez, T.; Ona, K.; Kapidzic, M.; Gormley, M.; Simón, C.; Genbacev, O.; Fisher, S.J. Severe preeclampsia is associated with alterations in cytotrophoblasts of the smooth chorion. Development 2017, 144, 767–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, E.L. Pregnancy and olfaction: A review. Front. Psychol. 2014, 5, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konečná, B.; Lauková, L.; Vlková, B. Immune Activation by Nucleic Acids: A Role in Pregnancy Complications. Scand. J. Immunol. 2018, 87, e12651. [Google Scholar] [CrossRef] [PubMed]
- Shu, C.; Liu, Z.; Cui, L.; Wei, C.; Wang, S.; Tang, J.J.; Cui, M.; Lian, G.; Li, W.; Liu, X.; et al. Protein Profiling of Preeclampsia Placental Tissues. PLoS ONE 2014, 9, e112890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Song, G.; Zhao, G.; Meng, T. Gene polymorphism associated with TGF-β1 and susceptibility to preeclampsia: A meta-analysis and trial sequential analysis. J. Obstet. Gynaecol. Res. 2021, 47, 2031–2041. [Google Scholar] [CrossRef]
- Liu, Y.; Li, P. Meta-Analysis of Angiotensin-Converting Enzyme Insersion/Delection Polymorphism and Pre-Eclampsia Susceptibility. J. Obstet. Gynaecol. Res. 2020, 46, 1744–1757. [Google Scholar] [CrossRef]
- Wang, X.; Sun, T.; Chen, G.; Gao, H. Association between Vascular Endothelial Growth Factor Gene Polymorphisms and Pre-Eclampsia Susceptibility: An Updated Meta-Analysis. Immunol. Investig. 2020, 49, 120–133. [Google Scholar] [CrossRef]
- Miller, A.K.; Azhibekov, T.; O’Toole, J.F.; Sedor, J.R.; Williams, S.M.; Redline, R.W.; Bruggeman, L.A. Association of preeclampsia with infant APOL1 genotype in African Americans. BMC Med. Genet. 2020, 21, 110. [Google Scholar] [CrossRef]
- Kang, Q.; Li, W.; Xiao, J.; Yu, N.; Fan, L.; Sha, M.; Ma, S.; Wu, J.; Chen, S. Integrated Analysis of Multiple Microarray Studies to Identify Novel Gene Signatures in Preeclampsia. Placenta 2021, 105, 104–118. [Google Scholar] [CrossRef]
- Meng, Y.; Li, C.; Liu, C.-X. Immune Cell Infiltration Landscape and Immune Marker Molecular Typing in Preeclampsia. Bioengineered 2021, 12, 540–554. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Module Color | GS Value | Number of Genes | Top Five Module Genes |
|---|---|---|---|
| Dark red | 0.41 (p = 0.006) | 3162 | MRPL19, FLJ12903, C2orf33, SLC25A5, ZNF627 |
| Dark turquoise | 0.35 (p = 0.02) | 402 | GPR92, GPR7, PADI3, MATN4, BAZ2B |
| Midnight blue | −0.69 (p = 4 × 10−7) | 1340 | HKE2, BANF1, CAPNS1, CREBL1, C6orf109 |
| #Term ID | Description | Observed Gene Count | Background Gene Count | False Discovery Rate |
|---|---|---|---|---|
| GO:0004930 | G-protein-coupled receptor activity | 38 | 824 | 0.0007 |
| GO:0008188 | neuropeptide receptor activity | 8 | 47 | 0.003 |
| GO:0004888 | transmembrane signaling receptor activity | 45 | 1226 | 0.0044 |
| GO:0008528 | G-protein-coupled peptide receptor activity | 12 | 132 | 0.0044 |
| GO:0060089 | molecular transducer activity | 50 | 1483 | 0.0105 |
| GO:0038023 | signaling receptor activity | 48 | 1429 | 0.0137 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vidal, M.S., Jr.; Deguit, C.D.T.; Yu, G.F.B.; Amosco, M.D. G-Protein Coupled Receptor Dysregulation May Play Roles in Severe Preeclampsia—A Weighted Gene Correlation Network Analysis of Placental Gene Expression Profile. Cells 2022, 11, 763. https://doi.org/10.3390/cells11050763
Vidal MS Jr., Deguit CDT, Yu GFB, Amosco MD. G-Protein Coupled Receptor Dysregulation May Play Roles in Severe Preeclampsia—A Weighted Gene Correlation Network Analysis of Placental Gene Expression Profile. Cells. 2022; 11(5):763. https://doi.org/10.3390/cells11050763
Chicago/Turabian StyleVidal, Manuel S., Jr., Christian Deo T. Deguit, Gracia Fe B. Yu, and Melissa D. Amosco. 2022. "G-Protein Coupled Receptor Dysregulation May Play Roles in Severe Preeclampsia—A Weighted Gene Correlation Network Analysis of Placental Gene Expression Profile" Cells 11, no. 5: 763. https://doi.org/10.3390/cells11050763
APA StyleVidal, M. S., Jr., Deguit, C. D. T., Yu, G. F. B., & Amosco, M. D. (2022). G-Protein Coupled Receptor Dysregulation May Play Roles in Severe Preeclampsia—A Weighted Gene Correlation Network Analysis of Placental Gene Expression Profile. Cells, 11(5), 763. https://doi.org/10.3390/cells11050763