
Emerging Role of Oxidative Stress on EGFR and OGG1-BER Cross-Regulation: Implications in Thyroid Physiopathology

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
2.2. Cell Viability Assay
2.3. Flow Cytometry and Cell Cycle Assay
2.4. Real-Time Quantitative PCR Analysis (qRT-PCR)
2.5. Gene Dosage Assay of OGG1 and MUTYH
2.6. Western Blotting
2.7. Statistical Analysis and Tools
3. Results
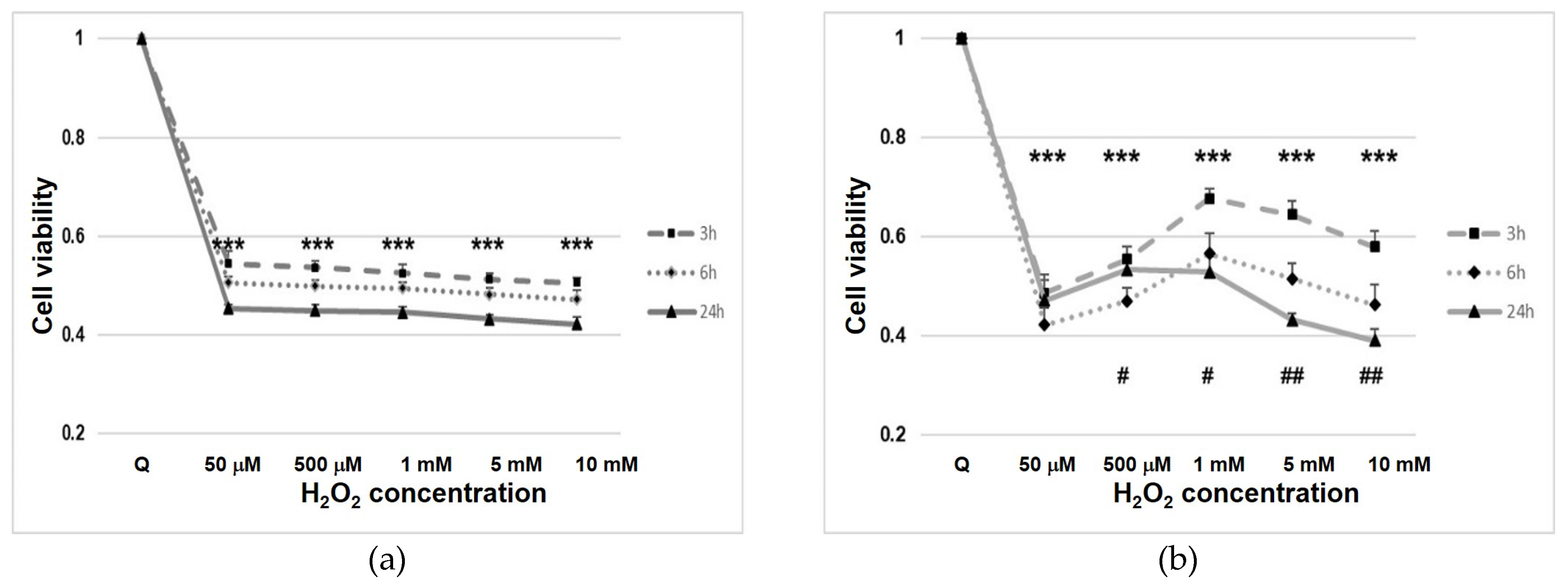
3.1. Cell Line Viability after Hydrogen Peroxide (H2O2) Treatment
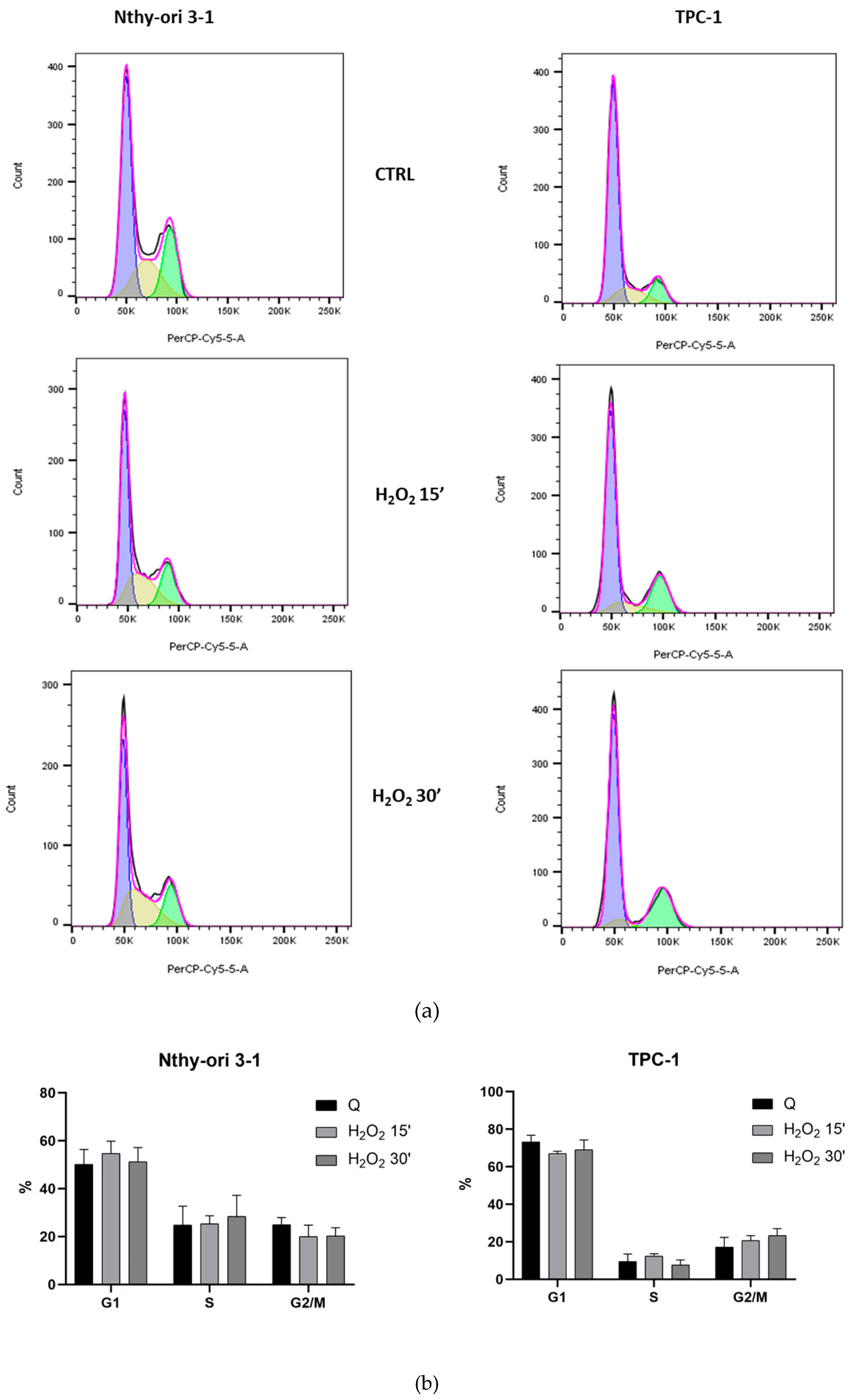
3.2. Cell Cycle Analysis
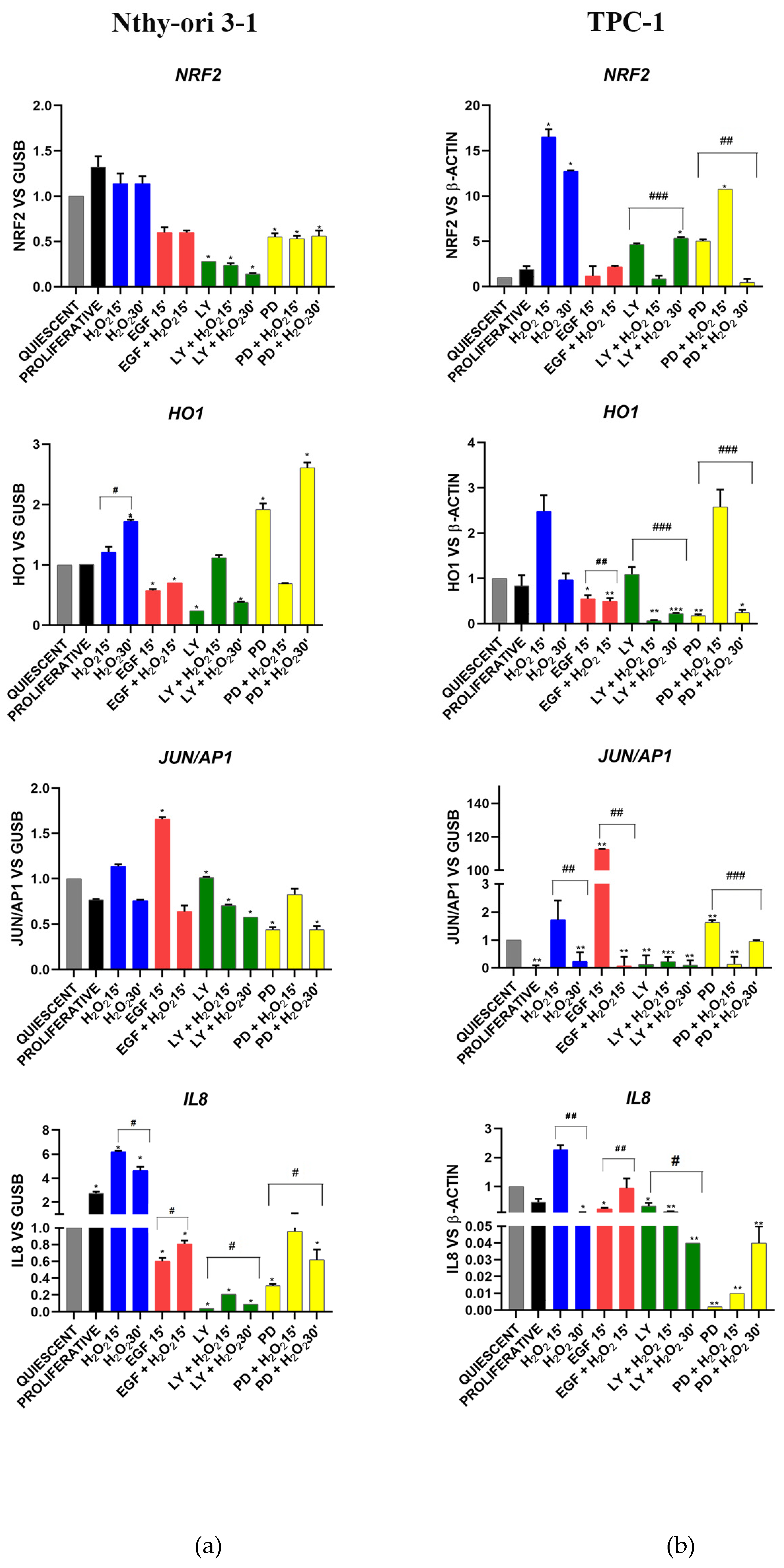
3.3. Gene Expression of BER and EGF Signaling in Nthy-ori 3-1 vs. TPC-1
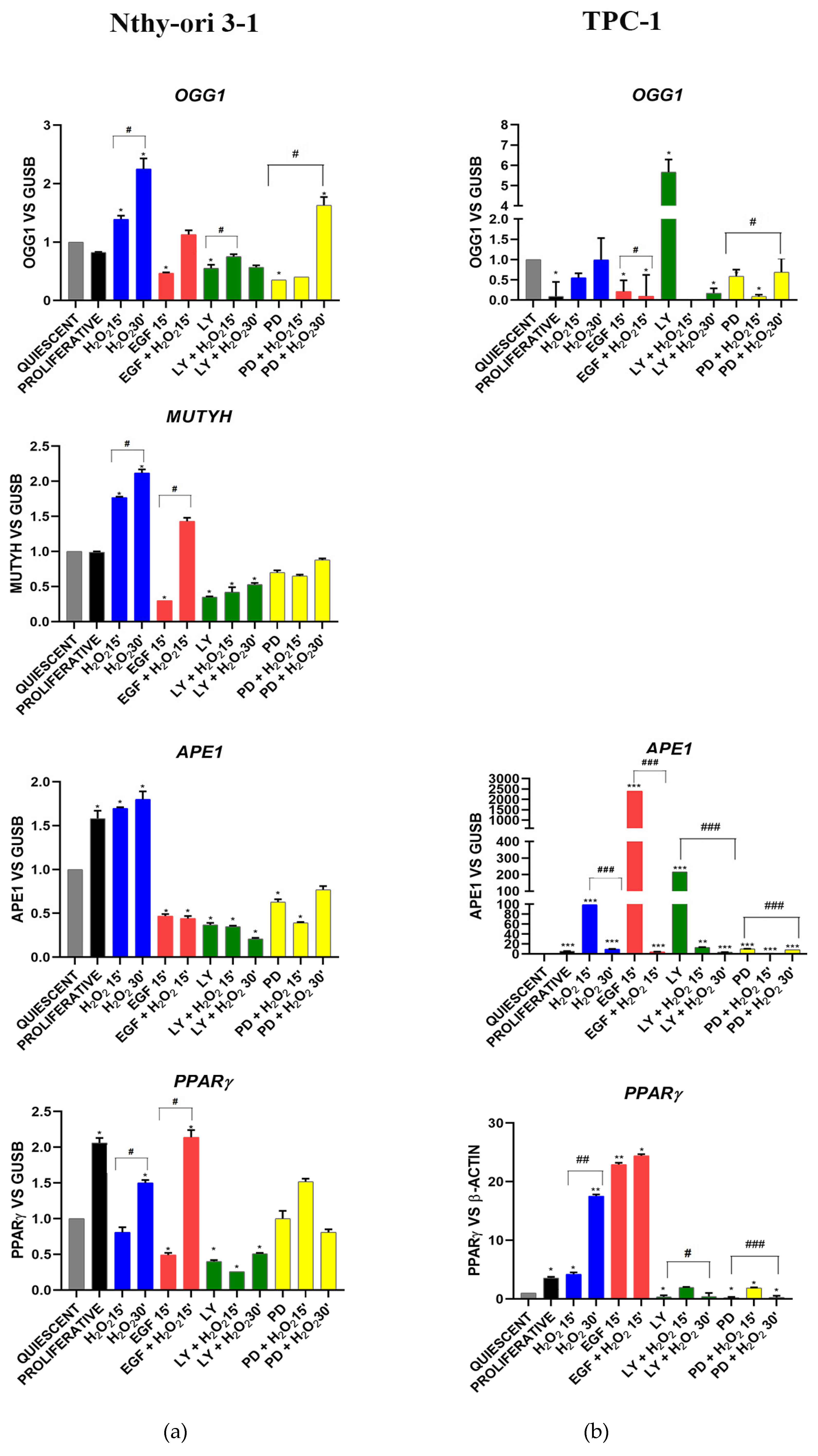
3.3.1. OGG1-BER Signaling
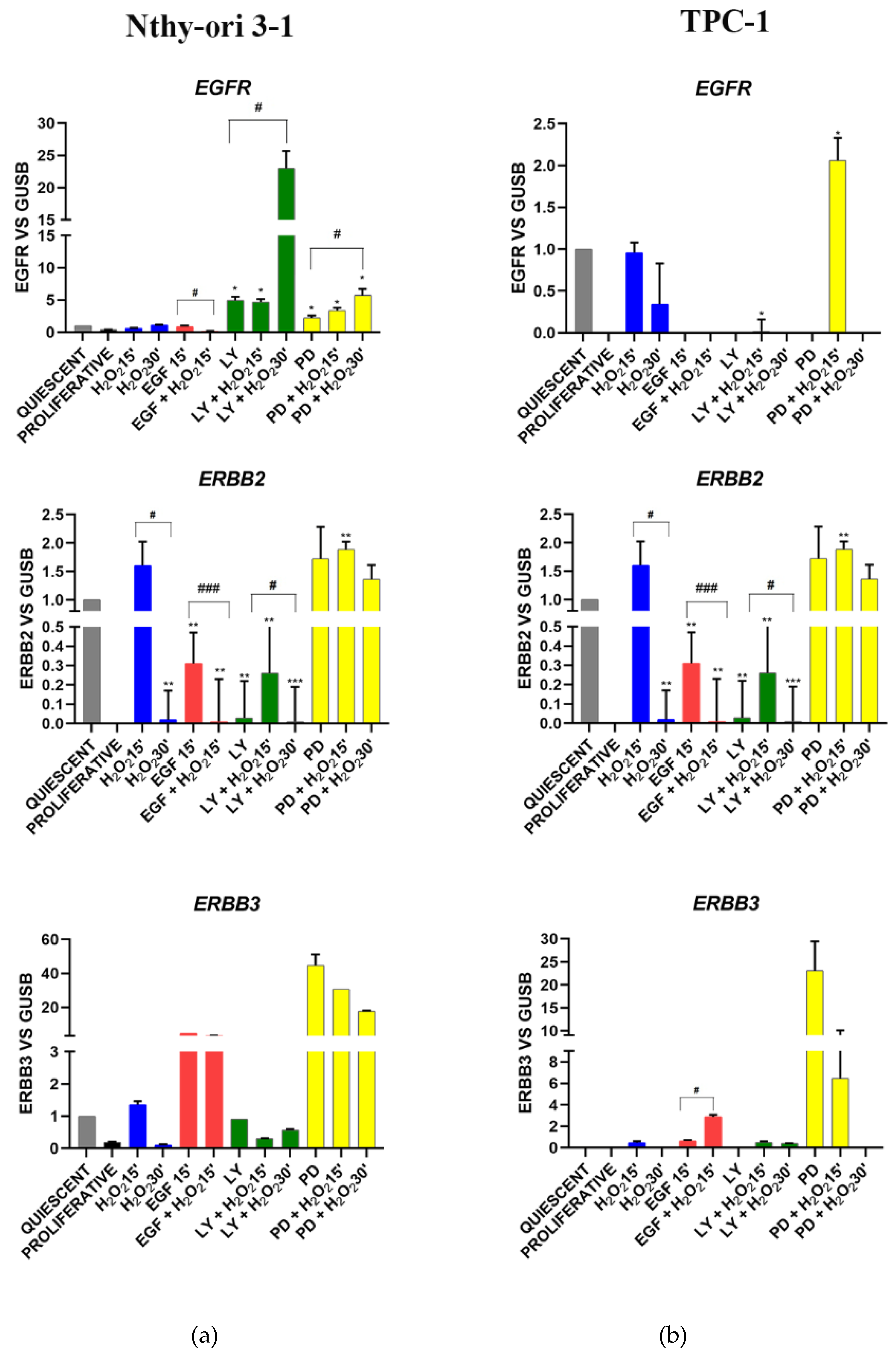
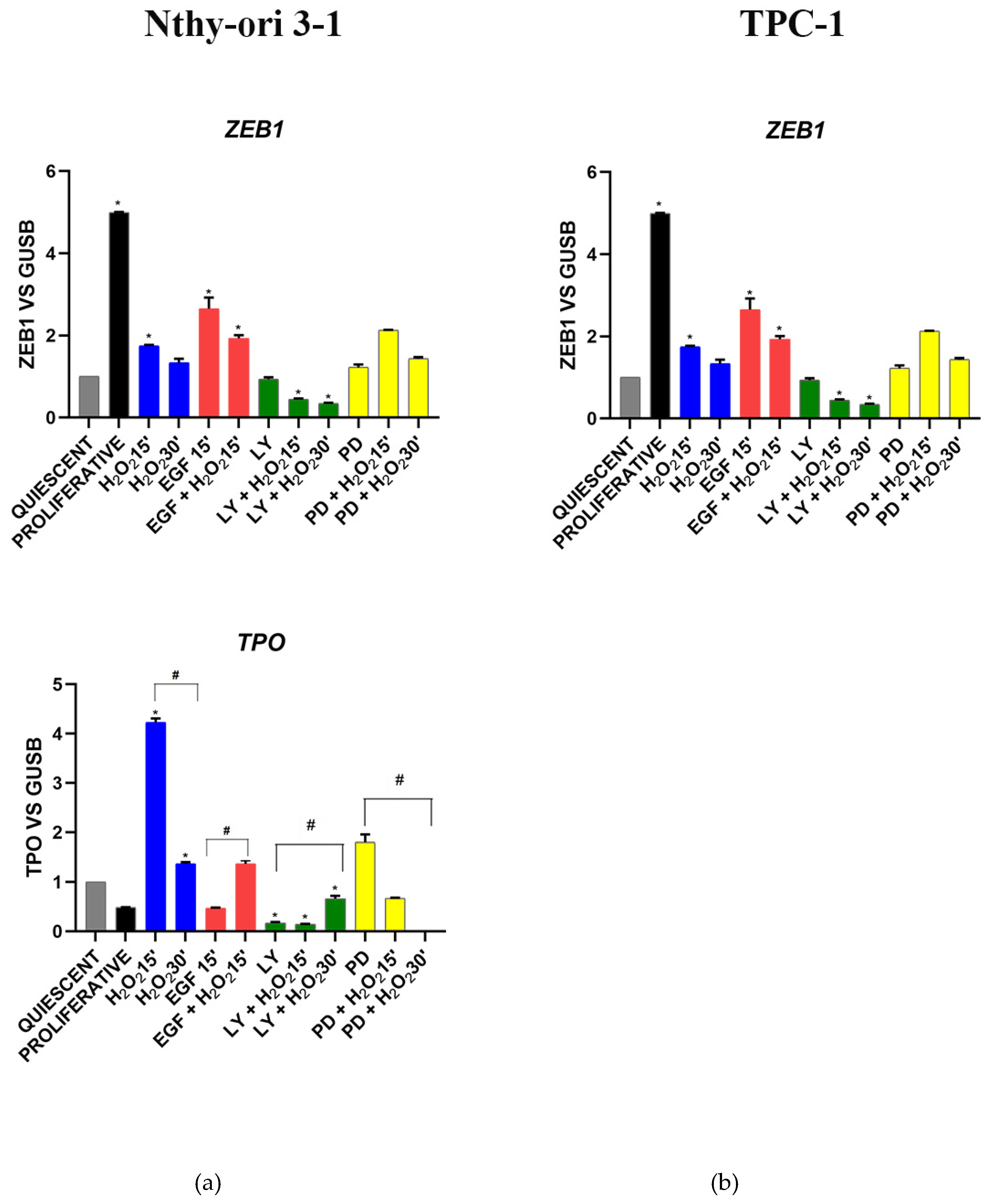
3.3.2. EGF Signaling
3.3.3. Oxidative Stress Signaling
3.3.4. Stemness and Differentiation Markers
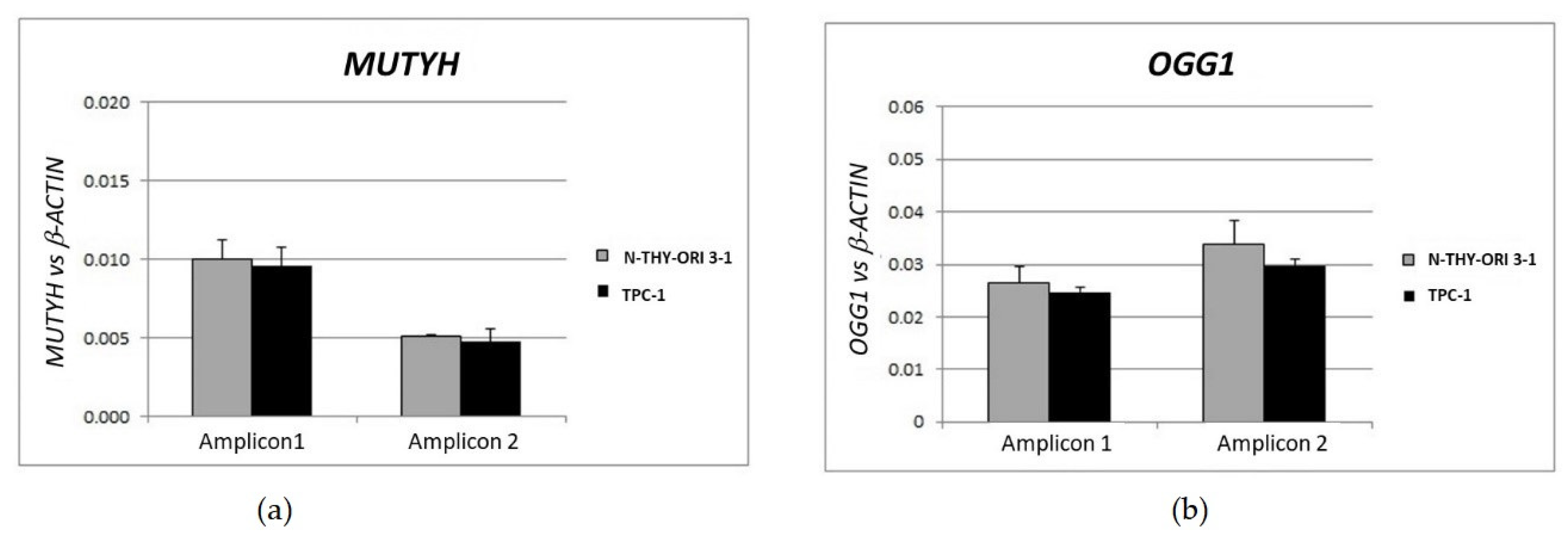
3.4. Gene Dosage Assay of OGG1 and MUTYH in TPC1 Cells
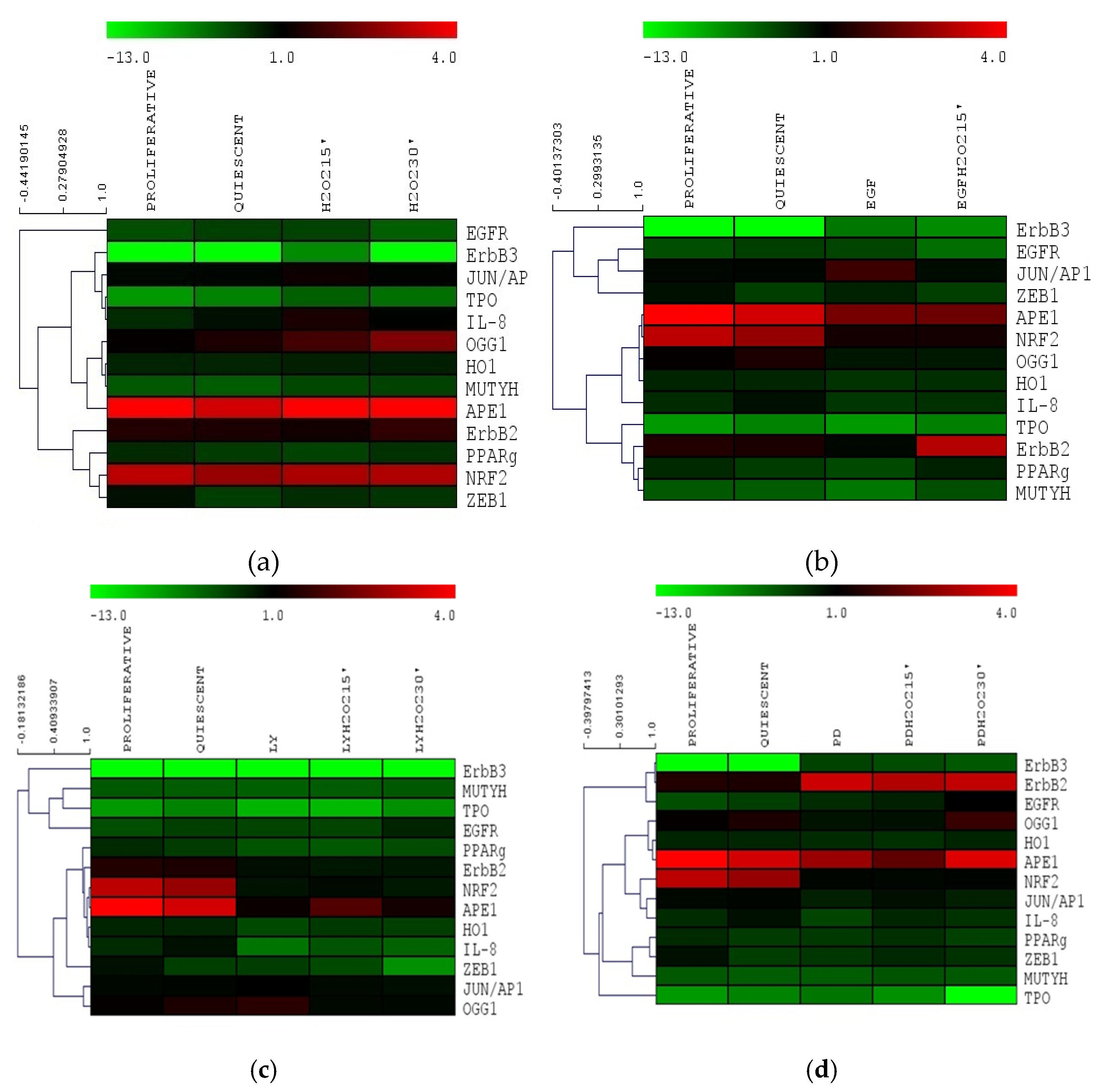
3.5. Gene Expression Cluster Analysis in Nthy-ori 3-1
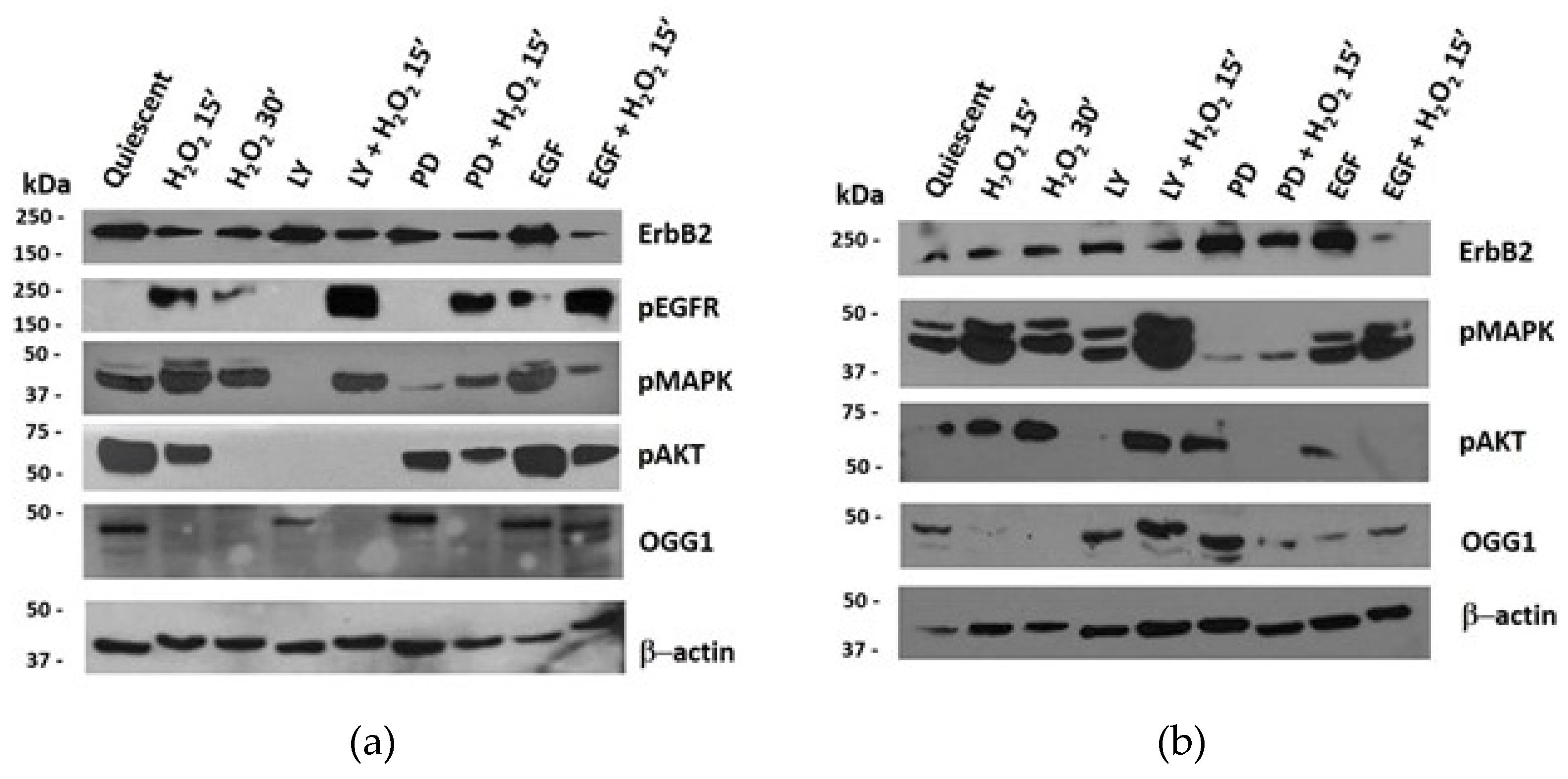
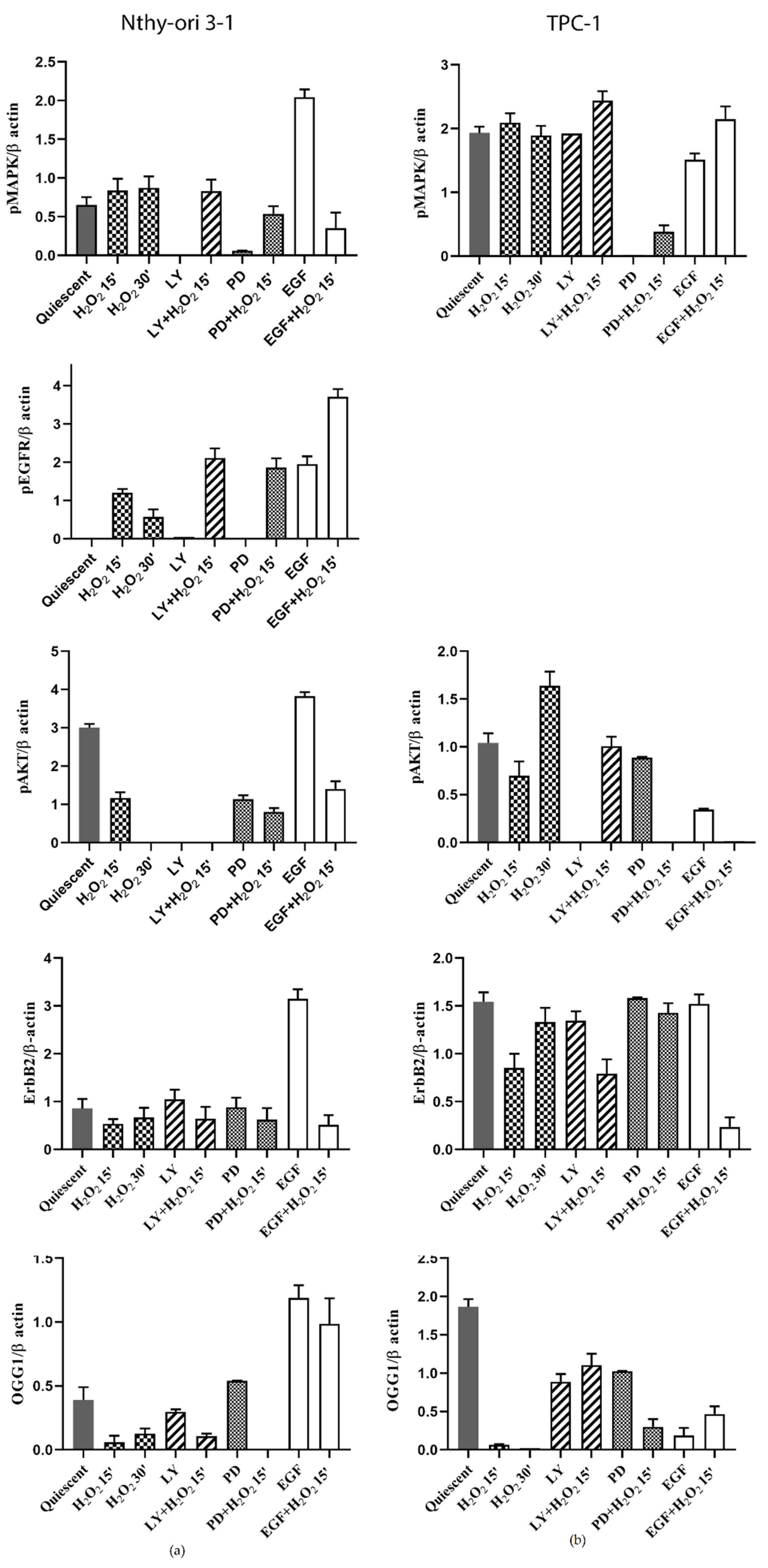
3.6. Protein Expression of ErbB Pathway and OGG1 in NThy-ori 3-1 and TPC-1 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMPK | adenosine-monophosphate-activated protein kinase |
| AKT | protein kinase B |
| BER | base excision repair |
| DUOX2 | dual oxidase 2 |
| APE1/Ref1 | apurinic/apyrimidinic endodeoxyribonuclease-1 |
| EGFR | epidermal growth factor receptor |
| ErbB2 | Erb-B2 receptor tyrosine kinase 2 |
| ErbB3 | Erb-B3 receptor tyrosine kinase 3 |
| ErbB4 | Erb-B4 receptor tyrosine kinase 4 |
| GUSB | glucuronidase Beta |
| HO1 | heme oxygenase 1 |
| IL8 | CXCL8, interleukin 8 |
| Jun/AP1 | Jun proto-oncogene, AP-1 transcription factor subunit |
| LY | LY294002; |
| MAPK | mitogen-activated protein kinase |
| MUTYH | mutY DNA glycosylase |
| NRF2 | realted factor |
| OGG1 | 8-oxoguanine glycosylase-1 |
| PPARg | peroxisome-proliferator-activated receptor gamma |
| PTC | papillary thyroid carcinoma |
| ROS | reactive oxygen species |
| TPO | thyroid peroxidase |
| ZEB-1 | zinc finger E-box-binding homeobox 1 |
| PD | PD98059 |
References
- Lim, H.; Devesa, S.S.; Sosa, J.A.; Check, D.; Kitahara, C.M. Trends in Thyroid Cancer Incidence and Mortality in the United States, 1974-2013. JAMA 2017, 317, 1338–1348. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.A.; Tublin, M.E. Postoperative surveillance of differentiated thyroid carcinoma: Rationale, techniques, and controversies. Radiology 2008, 249, 429–444. [Google Scholar] [CrossRef]
- Arora, N.; Scognamiglio, T.; Zhu, B.; Fahey, T.J. Do benign thyroid nodules have malignant potential? An evidence-based review. World J. Surg. 2008, 32, 1237–1246. [Google Scholar] [CrossRef]
- Ameziane El Hassani, R.; Buffet, C.; Leboulleux, S.; Dupuy, C. Oxidative stress in thyroid carcinomas: Biological and clinical significance. Endocr. Relat. Cancer. 2019, 26, R131–R143. [Google Scholar] [CrossRef] [Green Version]
- Moloney, J.N.; Cotter, T.G. ROS signaling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Ekholm, R. Iodination of thyroglobulin: An intracellular or extracellular process? Mol. Cell. Endocrinol. 1981, 24, 141–163. [Google Scholar] [CrossRef]
- Howie, A.F.; Arthur, J.R.; Nicol, F.; Walker, S.W.; Beech, S.G.; Beckett, G.J. Identification of a 57-kilodalton selenoprotein in human thyrocytes as thioredoxin reductase and evidence that its expression is regulated through the calcium-phosphoinositol signaling pathway. J. Clin. Endocrinol. Metab. 1998, 83, 2052–2058. [Google Scholar]
- Carvalho, D.P.; Dupuy, C. Thyroid hormone biosynthesis and release. Mol. Cell. Endocrinol. 2017, 458, 6–15. [Google Scholar] [CrossRef]
- Sigurdson, A.J.; Hauptmann, M.; Alexander, B.H.; Doody, M.M.; Thomas, C.B.; Struewing, J.P.; Jones, I.M. DNA damage among thyroid cancer and multiple cancer cases, controls, and long-lived individuals. Mutat. Res. 2005, 586, 173–188. [Google Scholar] [CrossRef] [Green Version]
- Sies, H. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef]
- Kumari, S.; Badana, A.K.; Mohan, G.M.; Shailender, G.; Malla, R. Reactive Oxygen Species: A Key Constituent in Cancer Survival. Biomark. Insights 2018, 13, 1177271918755391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, M.S.; Chang, J.H.; Hung, W.Y.; Yang, Y.C.; Chien, M.H. The interplay of reactive oxygen species and the epidermal growth factor receptor in tumor progression and drug resistance. J. Exp. Clin. Cancer Res. 2018, 37, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metere, A.M.; Frezzotti, F.; Graves, C.E.; Vergine, M.; De Luca, A.; Pietraforte, D.; Giacomelli, L.A. Possible role for selenoprotein glutathione peroxidase (GPx1) and thioredoxin reductases (TrxR1) in thyroid cancer: Our experience in thyroid surgery. Cancer Cell Int. 2018, 18, 7. [Google Scholar] [CrossRef] [PubMed]
- Szanto, I.; Pusztaszeri, M.; Mavromati, M. H2O2 Metabolism in Normal Thyroid Cells and in Thyroid Tumorigenesis: Focus on NADPH Oxidases. Antioxidants 2019, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Yi, J.W.; Park, J.Y.; Sung, J.Y.; Kwak, S.H.; Yu, J.; Chang, J.H.; Kim, J.H.; Ha, S.Y.; Paik, E.K.; Lee, W.S.; et al. Genomic evidence of reactive oxygen species elevation in papillary thyroid carcinoma with Hashimoto thyroiditis. Endocr. J. 2015, 62, 857–877. [Google Scholar] [CrossRef] [Green Version]
- Soultanakis, R.P.; Melamede, R.J.; Bespalov, I.A.; Wallace, S.S.; Beckman, K.B.; Ames, B.N.; Taatjes, D.J.; Janssen-Heininger, Y.M. Fluorescence detection of 8-oxoguanine in nuclear and mitochondrial DNA of cultured cells using a recombinant Fab and confocal scanning laser microscopy. Free Radic. Biol. Med. 2000, 28, 987–998. [Google Scholar] [CrossRef]
- Mo, J.Y.; Maki, H.; Sekiguchi, M. Hydrolytic elimination of a mutagenic nucleotide, 8-oxodGTP by human 18-kilodalton protein: Sanitization of nucleotide pool. Proc. Natl. Acad. Sci. USA 1992, 89, 11021–11025. [Google Scholar] [CrossRef] [Green Version]
- Oka, S.; Leon, J.; Tsuchimoto, D.; Sakumi, K.; Nakabeppu, Y. MUTYH, an adenine DNA glycosylase, mediates p53 tumor suppression via PARP-dependent cell death. Oncogenesis. 2015, 4, e142. [Google Scholar] [CrossRef] [Green Version]
- Banda, D.M.; Nuñez, N.N.; Burnside, M.A.; Bradshaw, K.M.; David, S.S. Repair of 8-oxoG:A mismatches by the MUTYH glycosylase:Mechanism, metals and medicine. Free Radic. Biol. Med. 2017, 107, 202–215. [Google Scholar] [CrossRef]
- Paschke, R. Molecular pathogenesis of nodular goiter. Langenbecks Arch. Surg. 2011, 396, 1127–1397. [Google Scholar] [CrossRef]
- Svilar, D.; Goellner, E.M.; Almeida, K.H.; Sobol, R.W. Base excision repair and lesion-dependent subpathways for repair of oxidative DNA damage. Antioxid. Redox Signal 2011, 14, 2491–2507. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Li, C.; Qiao, P.; Xue, Y.; Zheng., X.; Chen, H.; Zeng, X.; Liu, W.; Boldogh, I.; Ba, X. OGG1-initiated base excision repair exacerbates oxidative stress-induced parthanatos. Cell Death Dis. 2018, 9, 628. [Google Scholar] [CrossRef] [PubMed]
- Mincione, G.; Di Marcantonio, M.C.; Tarantelli, C.; D’Inzeo, S.; Nicolussi, A.; Nardi, F.; Donini, C.F.; Coppa, A. EGF and TGF-β1 Effects on Thyroid Function. J. Thyroid Res. 2011, 2011, 431718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ely, A.; Bischoff, L.A.; Weiss, V.L. Wnt signaling in thyroid homeostasis and carcinogenesis. Genes 2018, 9, 204. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.J.; Nirodi, C.S. The epidermal growth factor receptor: A role in repair of radiation- induced DNA damage. Clin. Cancer Res. 2007, 13, 6555–6560. [Google Scholar] [CrossRef] [Green Version]
- Bensimon, A.; Aebersold, R.; Shiloh, Y. Beyond ATM: The protein kinase landscape of the DNA damage response. FEBS Lett. 2011, 585, 1625–1639. [Google Scholar] [CrossRef] [Green Version]
- Oberthür, R.; Seemann, H.; Gehrig, J.; Rave-Fränk, M.; Bremmer, F.; Halpape, R.; Conradi, L.C.; Scharf, J.G.; Burfeind, P.; Kaulfuß, S. Simultaneous inhibition of IGF1R and EGFR enhances the efficacy of standard treatment for colorectal cancer by the impairment of DNA repair the induction of cell death. Cancer. Lett. 2017, 407, 93–105. [Google Scholar] [CrossRef]
- Saiselet, M.; Floor, S.; Tarabichi, M.; Dom, G.; Hébrant, A.; van Staveren, W.C.; Maenhaut, C. Thyroid cancer cell lines: An overview. Front. Endocrinol. 2012, 3, 133. [Google Scholar] [CrossRef] [Green Version]
- Nicolussi, A.; D’Inzeo, S.; Mincione, G.; Buffone, A.; Di Marcantonio, M.C.; Cotellese, R.; Cichella, A.; Capalbo, C.; Di Gioia, C.; Nardi, F.; et al. PRDX1 and PRDX6 are repressed in papillary thyroid carcinomas via BRAF V600E-dependent and -independent mechanisms. Int. J. Oncol. 2014, 44, 548–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirkmajer, S.; Chibalin, A.V. Serum starvation: Caveat emptor. Am. J. Physiol. Cell. Physiol. 2011, 301, C272–C279. [Google Scholar] [CrossRef] [Green Version]
- Massart, C.; Hoste, C.; Virion, A.; Ruf, J.; Dumont, J.E.; Van Sande, J. Cell biology of H2O2 generation in the thyroid: Investigation of the control of dual oxidases (DUOX) activity in intact ex vivo thyroid tissue and cell lines. Mol. Cell Endocrinol. 2011, 343, 32–44. [Google Scholar] [CrossRef]
- Lanuti, P.; Fuhrmann, S.; Lachmann, R.; Marchisio, M.; Miscia, S.; Kern, F. Simultaneous characterization of phospho-proteins and cell cycle in activated T cell subsets. Int. J. Immunopathol. Pharmacol. 2009, 22, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Bologna, G.; Lanuti, P.; D’Ambrosio, P.; Tonucci, L.; Pierdomenico, L.; D’Emilio, C.; Celli, N.; Marchisio, M.; d’Alessandro, N.; Santavenere, E.; et al. Water-soluble platinum phthalocyanines as potential antitumor agents. Biometals 2014, 27, 575–589. [Google Scholar] [CrossRef]
- Moscatello, C.; Di Nicola, M.; Veschi, S.; Di Gregorio, P.; Cianchetti, E.; Stuppia, L.; Battista, P.; Cama, A.; Curia, M.C.; Aceto, G.M. Relationship between MUTYH, OGG1 and BRCA1 mutations and mRNA expression in breast and ovarian cancer predisposition. Mol. Clin. Oncol. 2021, 14, 15. [Google Scholar] [CrossRef]
- Aceto, G.M.; Fantini, F.; De Iure, S.; Di Nicola, M.; Palka, G.; Valanzano, R.; Di Gregorio, P.; Stigliano, V.; Genuardi, M.; Battista, P.; et al. Correlation between mutations and mRNA expression of APC and MUTYH genes: New insight into hereditary colorectal polyposis predisposition. J. Exp. Clin. Cancer Res. 2015, 34, 131. [Google Scholar] [CrossRef] [Green Version]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furihata, C. An active alternative splicing isoform of human mitochondrial 8-oxoguanine DNA glycosylase (OGG1). Genes Environ. 2015, 37, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, R.; Ramdzan, Z.M.; Kaur, S.; Duquette, P.M.; Marcotte, R.; Leduy, L.; Davoudi, S.; Lamarche-Vane, N.; Iulianella., A.; Nepveu., A. CUX2 protein functions as an accessory factor in the repair of oxidative DNA damage. J. Biol. Chem. 2015, 290, 22520–22531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böttcher, Y.; Eszlinger, M.; Tönjes, A.; Paschke, R. The genetics of euthyroid familial goiter. Trends Endocrinol. Metab. 2005, 16, 314–319. [Google Scholar] [CrossRef]
- Yarden, Y.; Sliwkowski, M.X. Untangling the ErbB signaling network. Nat. Rev. Mol. Cell Biol. 2001, 2, 127–137. [Google Scholar] [CrossRef]
- Jena, N.R.; Mishra, P.C. Mechanisms of formation of 8-oxoguanine due to reactions of one and two OH* radicals and the H2O2 molecule with guanine: A quantum computational study. J. Phys. Chem. B 2005, 109, 14205–14218. [Google Scholar] [CrossRef] [PubMed]
- Ohno, M.; Oka, S.; Nakabeppu, Y. Quantitative analysis of oxidized guanine, 8-oxoguanine, in mitochondrial DNA by immunofluorescence method. Methods Mol. Biol. 2009, 554, 199–212. [Google Scholar] [PubMed]
- Barbouti, A.; Amorgianiotis, C.; Kolettas, E.; Kanavaros, P.; Galaris, D. Hydrogen peroxide inhibits caspase-dependent apoptosis by inactivating procaspase-9 in an iron-dependent manner. Free Radic. Biol. Med. 2007, 43, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Cell culture, oxidative stress, and antioxidants: Avoiding pitfalls. Biomed J. 2014, 37, 99–105. [Google Scholar] [CrossRef]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug. Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef]
- Wu, W.S. The signaling mechanism of ROS in tumor progression. Cancer Metastasis Rev. 2006, 25, 695–705. [Google Scholar] [CrossRef]
- Knobel, M.; Medeiros-Neto, G. An outline of inherited disorders of the thyroid hormone generating system. Thyroid 2003, 13, 771–801. [Google Scholar] [CrossRef]
- Kim, K.C.; Lee, I.K.; Kang, K.A.; Cha, J.W.; Cho, S.J.; Na, S.Y.; Chae, S.; Kim, H.S.; Kim, S.; Hyun, J.W. 7,8-Dihydroxyflavone suppresses oxidative stress-induced base modification in DNA via induction of the repair enzyme 8-oxoguanine DNA glycosylase-1. BioMed Res. Int. 2013, 2013, 863720. [Google Scholar] [CrossRef] [Green Version]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell Mol. Life Sci. 2016. 73, 3221–3247. [CrossRef] [Green Version]
- Wang, L.; Jiang, H.; Yin, Z.; Aschner, M.; Cai, J. Methylmercury toxicity and Nrf2-dependent detoxification in astrocytes. Toxicol. Sci. 2009, 107, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, R.; Gupta, S.; Fisher, B.J.; Ghosh, S.; Fowler, A.A., 3rd. Nitric oxide suppresses IL-8 transcription by inhibiting c-Jun N-terminal kinase-induced AP-1 activation. Exp. Cell. Res. 2001, 266, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Paszti-Gere, E.; Szeker, K.; Csibrik-Nemeth, E.; Csizinszky, R.; Marosi, A.; Palocz, O.; Farkas, O.; Galfi, P. Metabolites of Lactobacillus plantarum 2142 prevent oxidative stress-induced overexpression of proinflammatory cytokines in IPEC-J2 cell line. Inflammation 2012, 35, 1487–1499. [Google Scholar] [CrossRef] [PubMed]
- Waugh, D.J.; Wilson, C. The interleukin-8 pathway in cancer. Clin. Cancer Res. 2008, 14, 6735–6741. [Google Scholar] [CrossRef] [Green Version]
- Weetman, A.P.; Bennett, G.L.; Wong, W.L. Thyroid follicular cells produce interleukin-8. J. Clin. Endocrinol. Metab. 1992, 75, 328–330. [Google Scholar] [PubMed]
- Menendez, J.A. Fine-tuning the lipogenic/lipolytic balance to optimize the metabolic requirements of cancer cell growth: Molecular mechanisms and therapeutic perspectives. Biochim. Biophys. Acta. 2010, 1801, 381–391. [Google Scholar] [CrossRef]
- Carter, R.J.; Parsons, J.L. Base Excision Repair a Pathway Regulated by Posttranslational Modifications. Mol. Cell. Biol. 2016, 36, 1426–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, S.L. Mechanism of activation of AMPK and upregulation of OGG1 by rapamycin in cancer cells. Oncotarget 2011, 2, 958–959. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.R.; Parsons, J.L. The E3 Ubiquitin Ligase NEDD4L Targets OGG1 for Ubiquitylation and Modulates the Cellular DNA Damage Response. Front Cell Dev. Biol. 2020, 12, 607060. [Google Scholar] [CrossRef] [PubMed]
- Curia, M.C.; Catalano, T.; Aceto, G.M. MUTYH: Not just polyposis. World J. Clin. Oncol. 2020, 11, 428–449. [Google Scholar] [CrossRef] [PubMed]
- Frasca, F.; Vella, V.; Nicolosi, M.L.; Messina, R.L.; Gianì, F.; Lotta, S.; Vigneri, P.; Regalbuto, C.; Vigneri, R. Thyroid cancer cell resistance to gefitinib depends on the constitutive oncogenic activation of the ERK pathway. J. Clin. Endocrinol. Metab. 2013, 98, 2502–2512. [Google Scholar] [CrossRef] [Green Version]
- Borrello, M.G.; Alberti, L.; Fischer, A.; Degl’innocenti, D.; Ferrario, C.; Gariboldi, M.; Marchesi, F.; Allavena, P.; Greco, A.; Collini, P.; et al. Induction of a proinflammatory program in normal human thyrocytes by the RET/PTC1 oncogene. Proc. Natl. Acad. Sci. USA 2005, 102, 14825–14830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Yuan, K.; Yan, C.; Fox, J., 3rd; Gaid, M.; Breitwieser, W.; Bansal, A.K.; Zeng, H.; Gao, H.; Wu, M. 8-Oxoguanine-DNA glycosylase 1 deficiency modifies allergic airway inflammation by regulating STAT6 and IL-4 in cells and in mice. Free Radic Biol. Med. 2012, 52, 392–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ba, X.; Aguilera-Aguirre, L.; Rashid, Q.T.; Bacsi, A.; Radak, Z.; Sur, S.; Hosoki, K.; Hegde, M.L.; Boldogh, I. The role of 8-oxoguanine DNA glycosylase-1 in inflammation. Int. J. Mol. Sci. 2014, 15, 16975–16997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.W.; Hwang, J.H.; Suh, J.M.; Kim, H.; Song, J.H.; Hwang, E.S.; Hwang, I.Y.; Park, K.C.; Chung, H.K.; Kim, J.M.; et al. RET/PTC (rearranged in transformation/papillary thyroid carcinomas) tyrosine kinase phosphorylates and activates phosphoinositide-dependent kinase1 (PDK1): An alternative phosphatidylinositol 3-kinase-independent pathway to activate PDK1. Mol. Endocrinol. 2003, 17, 1382–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oka, S.; Nakabeppu, Y. DNA glycosylase encoded by MUTYH functions as a molecular switch for programmed cell death under oxidative stress to suppress tumorigenesis. Cancer Sci. 2011, 102, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.A.; Akhter, M.S.; Siejka, A.; Catravas, J.D.; Barabutis, N. P53 supports endothelial barrier function via APE1/Ref1 suppression. Immunobiology 2019, 224, 532–538. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moscatello, C.; Di Marcantonio, M.C.; Savino, L.; D’Amico, E.; Spacco, G.; Simeone, P.; Lanuti, P.; Muraro, R.; Mincione, G.; Cotellese, R.; et al. Emerging Role of Oxidative Stress on EGFR and OGG1-BER Cross-Regulation: Implications in Thyroid Physiopathology. Cells 2022, 11, 822. https://doi.org/10.3390/cells11050822
Moscatello C, Di Marcantonio MC, Savino L, D’Amico E, Spacco G, Simeone P, Lanuti P, Muraro R, Mincione G, Cotellese R, et al. Emerging Role of Oxidative Stress on EGFR and OGG1-BER Cross-Regulation: Implications in Thyroid Physiopathology. Cells. 2022; 11(5):822. https://doi.org/10.3390/cells11050822
Chicago/Turabian StyleMoscatello, Carmelo, Maria Carmela Di Marcantonio, Luca Savino, Emira D’Amico, Giordano Spacco, Pasquale Simeone, Paola Lanuti, Raffaella Muraro, Gabriella Mincione, Roberto Cotellese, and et al. 2022. "Emerging Role of Oxidative Stress on EGFR and OGG1-BER Cross-Regulation: Implications in Thyroid Physiopathology" Cells 11, no. 5: 822. https://doi.org/10.3390/cells11050822
APA StyleMoscatello, C., Di Marcantonio, M. C., Savino, L., D’Amico, E., Spacco, G., Simeone, P., Lanuti, P., Muraro, R., Mincione, G., Cotellese, R., & Aceto, G. M. (2022). Emerging Role of Oxidative Stress on EGFR and OGG1-BER Cross-Regulation: Implications in Thyroid Physiopathology. Cells, 11(5), 822. https://doi.org/10.3390/cells11050822