
Non-Invasive Optical Motion Tracking Allows Monitoring of Respiratory Dynamics in Dystrophin-Deficient Mice

 , , , , ,
, , , , , 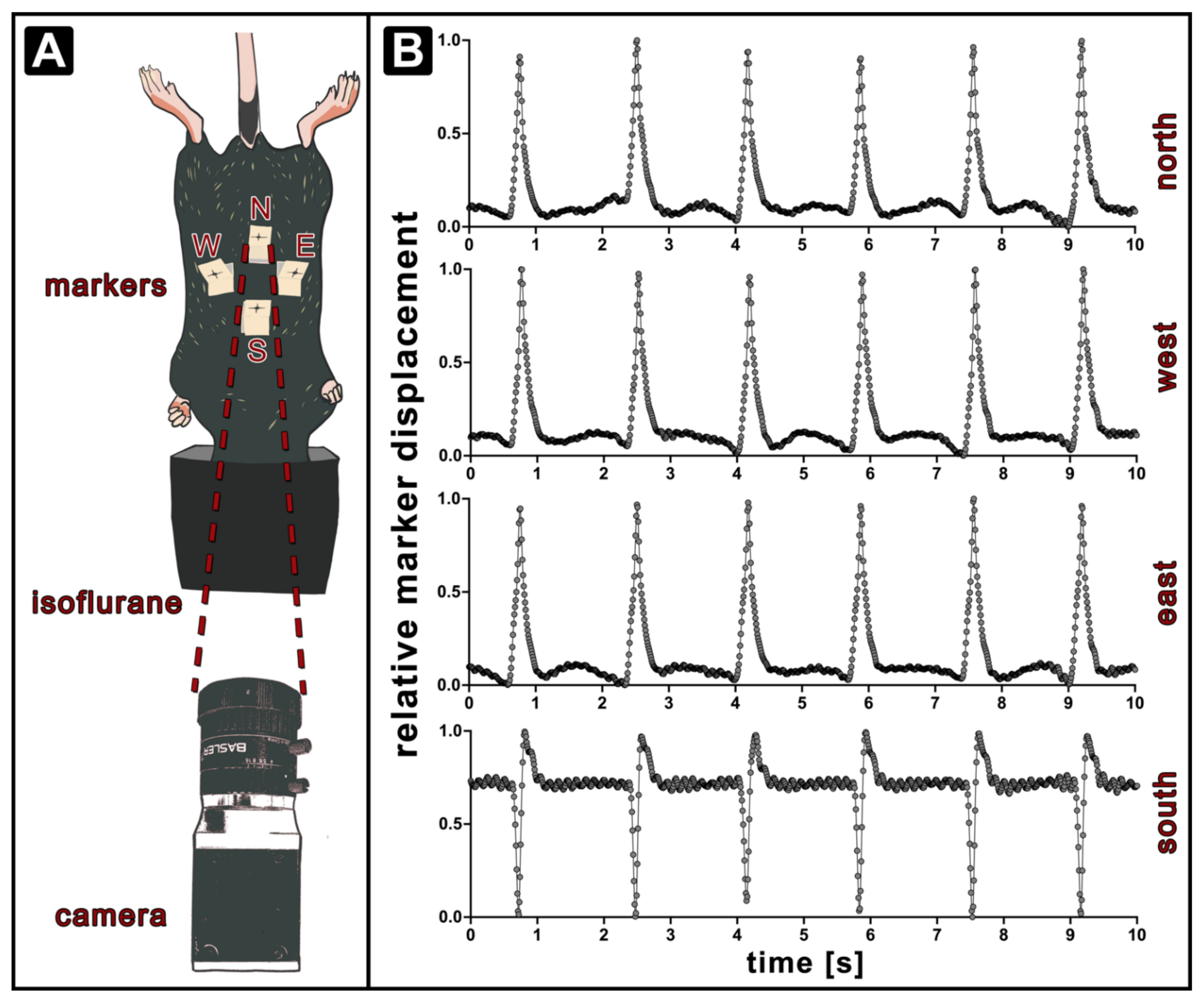{kind=link}
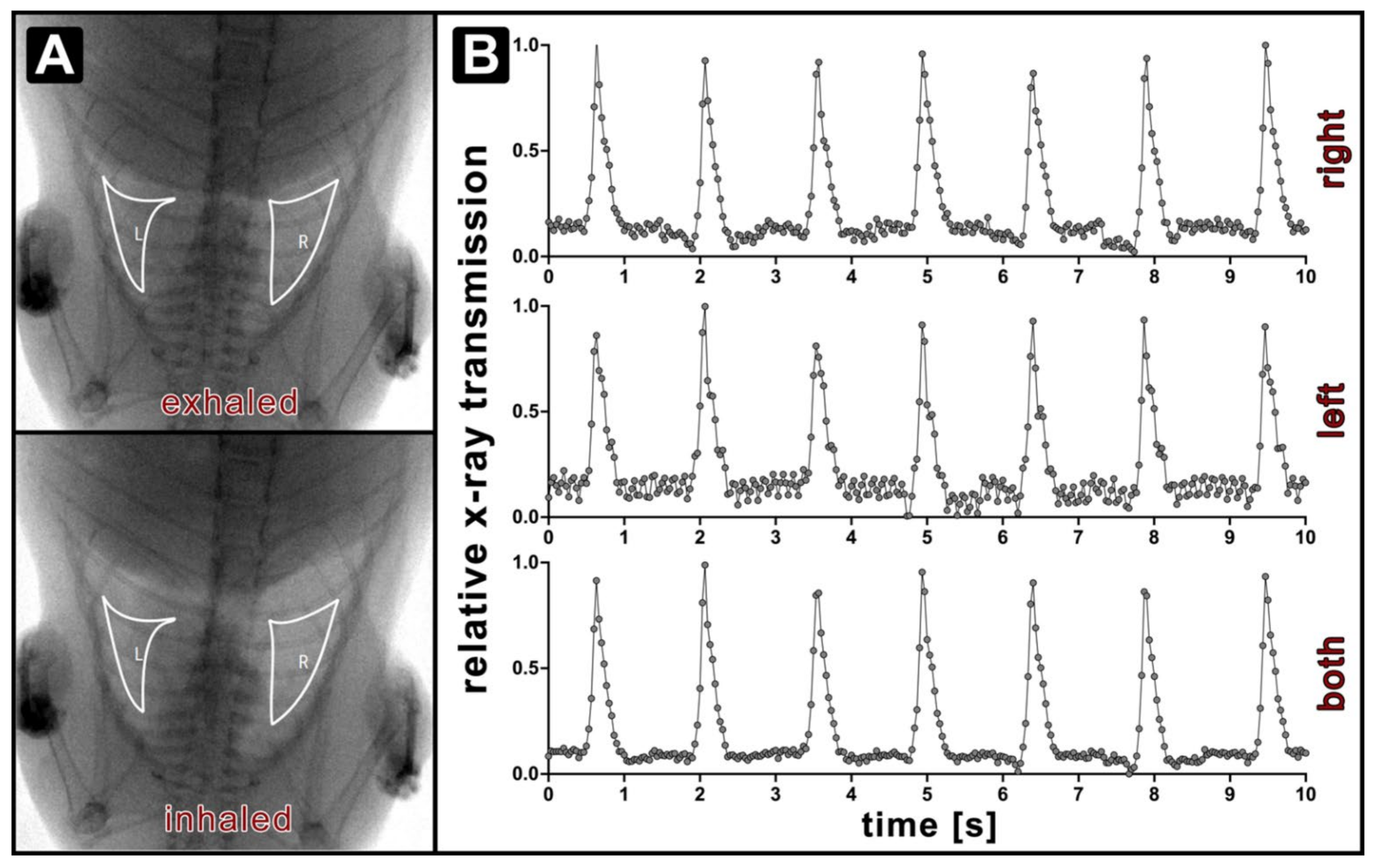{kind=link}
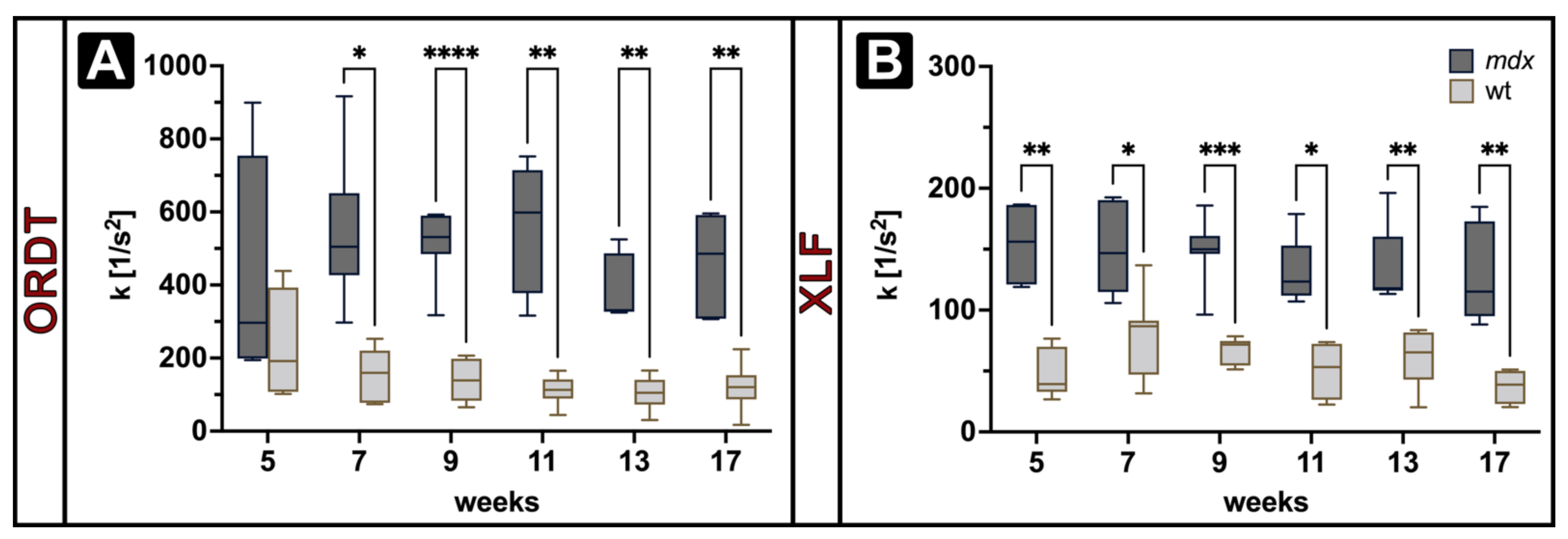{kind=link}
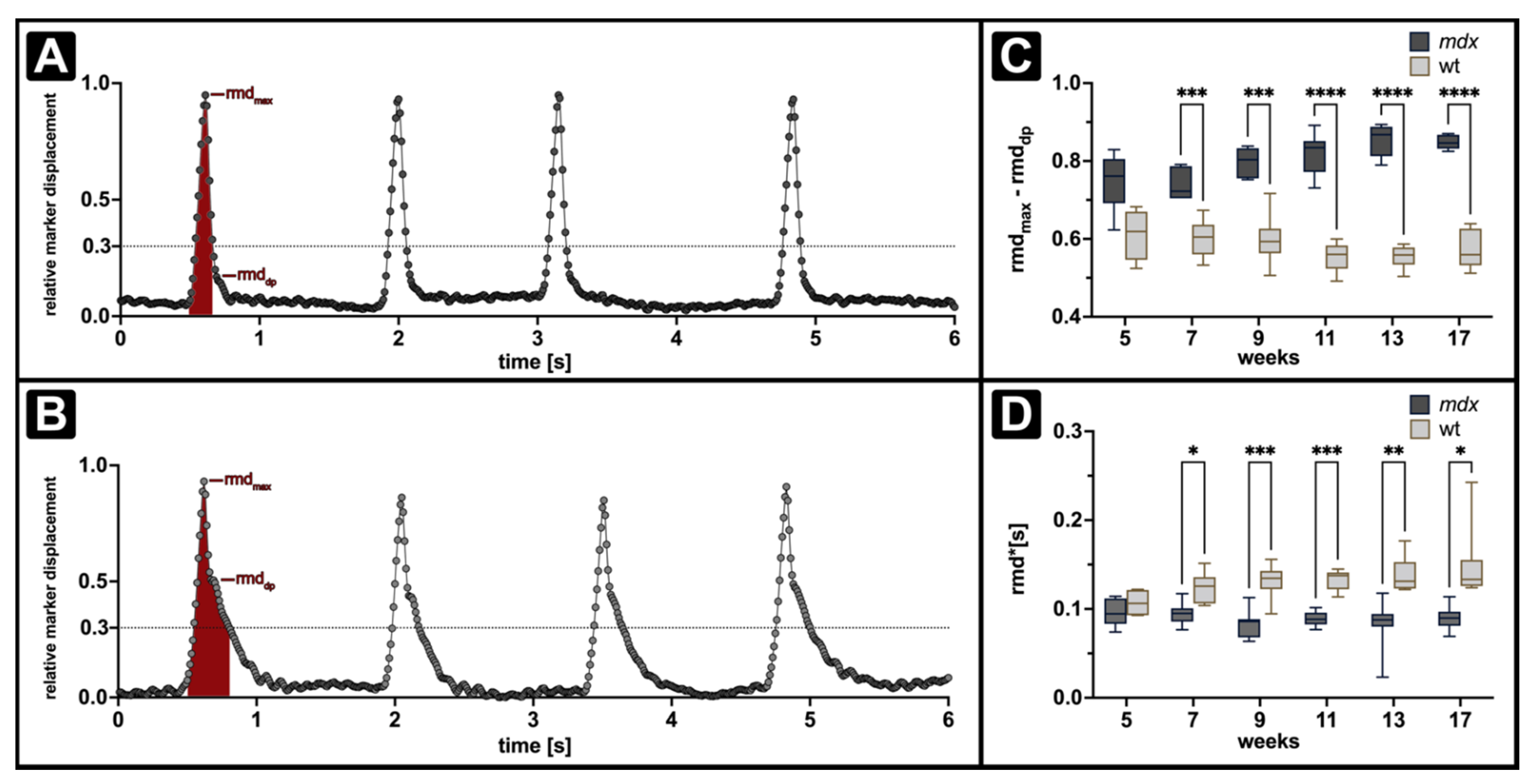{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Model and Experimental Design
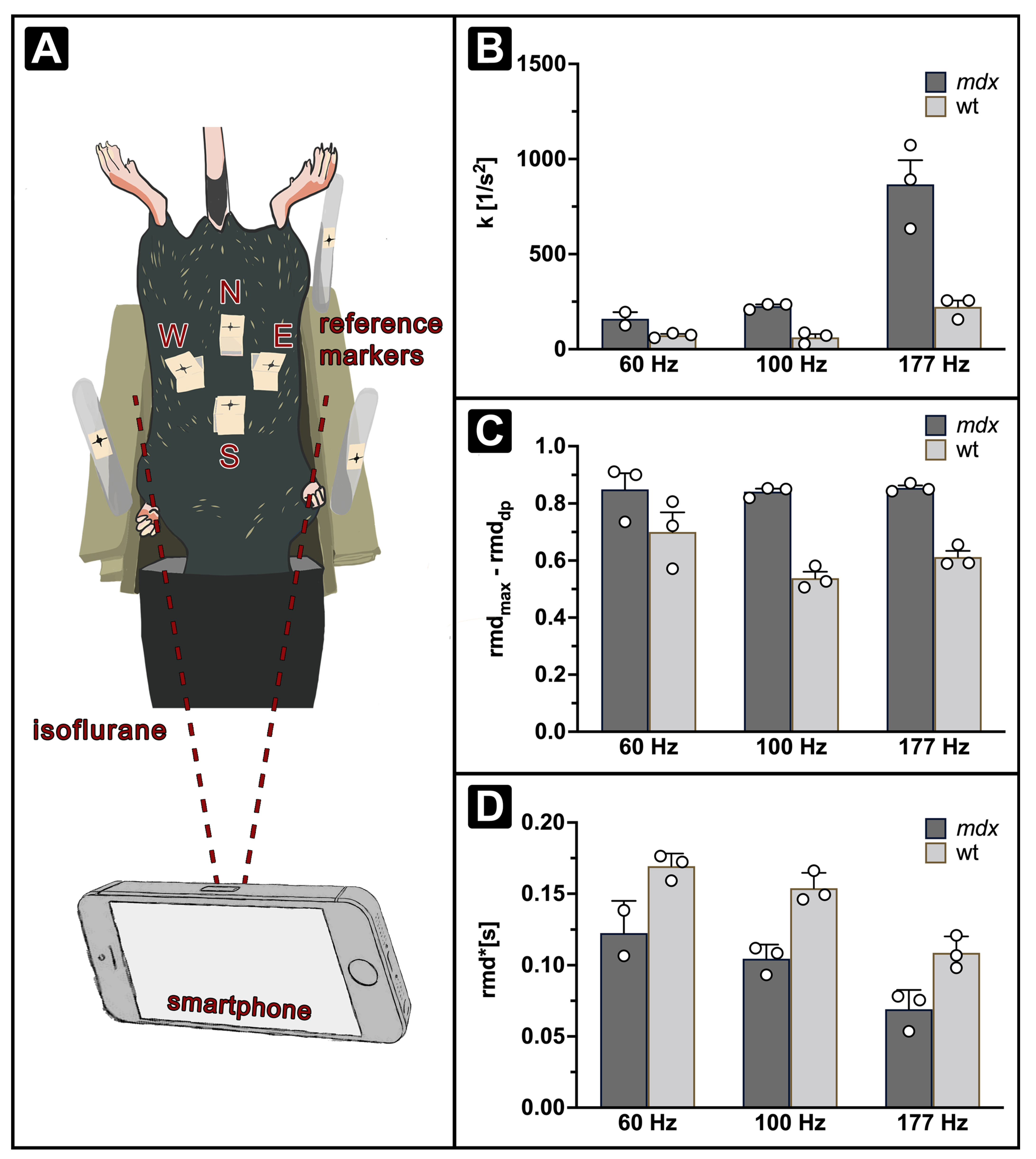
2.2. Optical Respiratory Dynamics Tracking (ORDT)
2.3. X-ray Lung Function Measurement
2.4. Statistical Analysis
3. Results
3.1. Optical Respiratory Dynamics Tracking (ORDT) Workflow and Its Comparison to XLF
3.2. Expiratory Dynamics Measured by ORDT Reproduce Expiratory Constant Differences Found by XLF
3.3. ORDT Identifies Two-Step Expiratory Behaviour which Differs between mdx and wt
3.4. iORDT Reveals the Importance of the Sampling Rate in Diseased Phenotype Differentiation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Emery, A.E. Population Frequencies of Inherited Neuromuscular Diseases—A World Survey. Neuromuscul. Disord. 1991, 1, 19–29. [Google Scholar] [CrossRef]
- Farini, A.; Sitzia, C.; Erratico, S.; Meregalli, M.; Torrente, Y. Influence of Immune Responses in Gene/Stem Cell Therapies for Muscular Dystrophies. BioMed Res. Int. 2014, 2014, 818107. [Google Scholar] [CrossRef] [PubMed]
- Łoboda, A.; Dulak, J. Muscle and Cardiac Therapeutic Strategies for Duchenne Muscular Dystrophy: Past, Present, and Future. Pharmacol. Rep. 2020, 72, 1227–1263. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, E.P.; Brown, R.H.; Kunkel, L.M. Dystrophin: The Protein Product of the Duchenne Muscular Dystrophy Locus. Cell 1987, 51, 919–928. [Google Scholar] [CrossRef]
- Bonilla, E.; Schmidt, B.; Samitt, C.E.; Miranda, A.F.; Hays, A.P.; De Oliveira, A.B.S.; Chang, H.W.; Servidei, S.; Ricci, E.; Younger, D.S.; et al. Normal and Dystrophin-Deficient Muscle Fibers in Carriers of the Gene for Duchenne Muscular Dystrophy. Am. J. Pathol. 1988, 133, 440–445. [Google Scholar]
- Ibraghimov-Beskrovnaya, O.; Ervasti, J.M.; Leveille, C.J.; Slaughter, C.A.; Sernett, S.W.; Campbell, K.P. Primary Structure of Dystrophin-Associated Glycoproteins Linking Dystrophin to the Extracellular Matrix. Nature 1992, 355, 696–702. [Google Scholar] [CrossRef]
- Ervasti, J.M. Dystrophin, Its Interactions with Other Proteins, and Implications for Muscular Dystrophy. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2007, 1772, 108–117. [Google Scholar] [CrossRef]
- Deconinck, N.; Dan, B. Pathophysiology of Duchenne Muscular Dystrophy: Current Hypotheses. Pediatr. Neurol. 2007, 36, 1–7. [Google Scholar] [CrossRef]
- Villalta, S.A.; Deng, B.; Rinaldi, C.; Wehling-Henricks, M.; Tidball, J.G. IFN-γ Promotes Muscle Damage in the Mdx Mouse Model of Duchenne Muscular Dystrophy by Suppressing M2 Macrophage Activation and Inhibiting Muscle Cell Proliferation. J. Immunol. 2011, 187, 5419–5428. [Google Scholar] [CrossRef]
- Nigro, G.; Comi, L.I.; Politano, L.; Bain, R.J.I. The Incidence and Evolution of Cardiomyopathy in Duchenne Muscular Dystrophy. Int. J. Cardiol. 1990, 26, 271–277. [Google Scholar] [CrossRef]
- Siegel, I.M. Pulmonary Problems in Duchenne Muscular Dystrophy. Diagnosis, Prophylaxis, and Treatment. Phys. Ther. 1975, 55, 160–162. [Google Scholar] [CrossRef]
- Bushby, K.; Finkel, R.; Birnkrant, D.J.; Case, L.E.; Clemens, P.R.; Cripe, L.; Kaul, A.; Kinnett, K.; McDonald, C.; Pandya, S.; et al. Diagnosis and Management of Duchenne Muscular Dystrophy, Part 1: Diagnosis, and Pharmacological and Psychosocial Management. Lancet Neurol. 2010, 9, 77–93. [Google Scholar] [CrossRef]
- Ryder, S.; Leadley, R.M.; Armstrong, N.; Westwood, M.; de Kock, S.; Butt, T.; Jain, M.; Kleijnen, J. The Burden, Epidemiology, Costs and Treatment for Duchenne Muscular Dystrophy: An Evidence Review. Orphanet J. Rare Dis. 2017, 12, 79. [Google Scholar] [CrossRef]
- Pradhan, S.; Mittal, B. Infraspinatus Muscle Hypertrophy and Wasting of Axillary Folds as the Important Signs in Duchenne Muscular Dystrophy. Clin. Neurol. Neurosurg. 1995, 97, 134–138. [Google Scholar] [CrossRef]
- Birnkrant, D.J.; Bushby, K.; Bann, C.M.; Apkon, S.D.; Blackwell, A.; Colvin, M.K.; Cripe, L.; Herron, A.R.; Kennedy, A.; Kinnett, K.; et al. Diagnosis and Management of Duchenne Muscular Dystrophy, Part 3: Primary Care, Emergency Management, Psychosocial Care, and Transitions of Care across the Lifespan. Lancet Neurol. 2018, 17, 445–455. [Google Scholar] [CrossRef]
- Eagle, M.; Bourke, J.; Bullock, R.; Gibson, M.; Mehta, J.; Giddings, D.; Straub, V.; Bushby, K. Managing Duchenne Muscular Dystrophy—The Additive Effect of Spinal Surgery and Home Nocturnal Ventilation in Improving Survival. Neuromuscul. Disord. 2007, 17, 470–475. [Google Scholar] [CrossRef]
- Bushby, K.; Finkel, R.; Wong, B.; Barohn, R.; Campbell, C.; Comi, G.P.; Connolly, A.M.; Day, J.W.; Flanigan, K.M.; Goemans, N.; et al. Ataluren Treatment of Patients with Nonsense Mutation Dystrophinopathy. Muscle Nerve 2014, 50, 477–487. [Google Scholar] [CrossRef]
- Mendell, J.R.; Rodino-Klapac, L.R.; Sahenk, Z.; Roush, K.; Bird, L.; Lowes, L.P.; Alfano, L.; Gomez, A.M.; Lewis, S.; Kota, J.; et al. Eteplirsen for the Treatment of Duchenne Muscular Dystrophy. Ann. Neurol. 2013, 74, 637–647. [Google Scholar] [CrossRef]
- Manini, A.; Abati, E.; Nuredini, A.; Corti, S.; Comi, G.P. Adeno-Associated Virus (AAV)-Mediated Gene Therapy for Duchenne Muscular Dystrophy: The Issue of Transgene Persistence. Front. Neurol. 2022, 12, 814174. [Google Scholar] [CrossRef]
- Olson, E.N. Toward the Correction of Muscular Dystrophy by Gene Editing. Proc. Natl. Acad. Sci. USA 2021, 118, e2004840117. [Google Scholar] [CrossRef]
- Sun, C.; Serra, C.; Lee, G.; Wagner, K.R. Stem Cell-Based Therapies for Duchenne Muscular Dystrophy. Exp. Neurol. 2020, 323, 113086. [Google Scholar] [CrossRef] [PubMed]
- Bladen, C.L.; Salgado, D.; Monges, S.; Foncuberta, M.E.; Kekou, K.; Kosma, K.; Dawkins, H.; Lamont, L.; Roy, A.J.; Chamova, T.; et al. The TREAT-NMD DMD Global Database: Analysis of More than 7000 Duchenne Muscular Dystrophy Mutations. Hum. Mutat. 2015, 36, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Grounds, M.D.; Radley, H.G.; Lynch, G.S.; Nagaraju, K.; De Luca, A. Towards Developing Standard Operating Procedures for Pre-Clinical Testing in the Mdx Mouse Model of Duchenne Muscular Dystrophy. Neurobiol. Dis. 2008, 31, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Stedman, H.H.; Sweeney, H.L.; Shrager, J.B.; Maguire, H.C.; Panettieri, R.A.; Petrof, B.; Narusawa, M.; Leferovich, J.M.; Sladky, J.T.; Kelly, A.M. The Mdx Mouse Diaphragm Reproduces the Degenerative changes of Duchenne muscular dystrophy. Nature 1991, 352, 536–539. [Google Scholar] [CrossRef]
- Ryder-Cook, A.S.; Sicinski, P.; Thomas, K.; Davies, K.E.; Worton, R.G.; Barnard, E.A.; Darlison, M.G.; Barnard, P.J. Localization of the Mdx Mutation within the Mouse Dystrophin Gene. EMBO J. 1988, 7, 3017–3021. [Google Scholar] [CrossRef]
- Sicinski, P.; Geng, Y.; Ryder-Cook, A.S.; Barnard, E.A.; Darlison, M.G.; Barnard, P.J. The Molecular Basis of Muscular Dystrophy in the Mdx Mouse: A Point Mutation. Science 1989, 244, 1578–1580. [Google Scholar] [CrossRef]
- Bulfield, G.; Siller, W.G.; Wight, P.A.; Moore, K.J. X Chromosome-Linked Muscular Dystrophy (Mdx) in the Mouse. Proc. Natl. Acad. Sci. USA 1984, 81, 1189–1192. [Google Scholar] [CrossRef]
- Lefaucheur, J.P.; Pastoret, C.; Sebille, A. Phenotype of Dystrophinopathy in Old Mdx Mice. Anat. Rec. 1995, 242, 70–76. [Google Scholar] [CrossRef]
- Khurana, T.J. Experimental Protocols for DMD Animal Models/Respiratory System Evaluation. Available online: https://treat-nmd.org/resources-support/research-overview/preclinical-research/experimental-protocols-for-dmd-animal-models/ (accessed on 29 December 2021).
- Gilchrist, J.M. Overview of Neuromuscular Disorders Affecting Respiratory Function. Semin. Respir. Crit. Care Med. 2002, 23, 191–200. [Google Scholar] [CrossRef]
- Bourke, S.C.; Gibson, G.J. Sleep and Breathing in Neuromuscular Disease. Eur. Respir. J. 2002, 19, 1194–1201. [Google Scholar] [CrossRef]
- Sahni, A.S.; Wolfe, L. Respiratory Care in Neuromuscular Diseases. Respir. Care 2018, 63, 601–608. [Google Scholar] [CrossRef]
- Pfeffer, G.; Povitz, M. Respiratory Management of Patients with Neuromuscular Disease: Current Perspectives. Degener. Neurol. Neuromuscul. Dis. 2016, 6, 111–118. [Google Scholar] [CrossRef]
- Ishizaki, M.; Suga, T.; Kimura, E.; Shiota, T.; Kawano, R.; Uchida, Y.; Uchino, K.; Yamashita, S.; Maeda, Y.; Uchino, M. Mdx Respiratory Impairment Following Fibrosis of the Diaphragm. Neuromuscul. Disord. 2008, 18, 342–348. [Google Scholar] [CrossRef]
- Burns, D.P.; Edge, D.; O’Malley, D.; O’Halloran, K.D. Respiratory Control in the Mdx Mouse Model of Duchenne Muscular Dystrophy. Adv. Exp. Med. Biol. 2015, 860, 239–244. [Google Scholar] [CrossRef]
- Burns, D.P.; Canavan, L.; Rowland, J.; O’Flaherty, R.; Brannock, M.; Drummond, S.E.; O’Malley, D.; Edge, D.; O’Halloran, K.D. Recovery of Respiratory Function in Mdx Mice Co-Treated with Neutralizing Interleukin-6 Receptor Antibodies and Urocortin-2. J. Physiol. 2018, 596, 5175–5197. [Google Scholar] [CrossRef]
- Burns, D.P.; Roy, A.; Lucking, E.F.; McDonald, F.B.; Gray, S.; Wilson, R.J.; Edge, D.; O’Halloran, K.D. Sensorimotor Control of Breathing in the Mdx Mouse Model of Duchenne Muscular Dystrophy. J. Physiol. 2017, 595, 6653–6672. [Google Scholar] [CrossRef]
- Quindry, J.C.; Ballmann, C.G.; Epstein, E.E.; Selsby, J.T. Plethysmography Measurements of Respiratory Function in Conscious Unrestrained Mice. J. Physiol. Sci. 2016, 66, 157–164. [Google Scholar] [CrossRef]
- Huang, P.; Cheng, G.; Lu, H.; Aronica, M.; Ransohoff, R.M.; Zhou, L. Impaired respiratory function in mdx and mdx/utrn+/− mice. Muscle Nerve 2011, 43, 263–267. [Google Scholar] [CrossRef]
- Khan, A.; Markus, A.; Rittmann, T.; Albers, J.; Alves, F.; Hülsmann, S.; Dullin, C. Simple Low Dose Radiography Allows Precise Lung Volume Assessment in Mice. Sci. Rep. 2021, 11, 4163. [Google Scholar] [CrossRef]
- Dullin, C.; Markus, M.A.; Larsson, E.; Tromba, G.; Huelsmann, S.; Alves, F. X-ray Based Lung Function Measurement—A Sensitive Technique to Quantify Lung Function in Allergic Airway Inflammation Mouse Models. Sci. Rep. 2016, 6, 36297. [Google Scholar] [CrossRef]
- Markus, M.A.; Borowik, S.; Reichardt, M.; Tromba, G.; Alves, F.; Dullin, C. X-ray-Based Lung Function Measurement Reveals Persistent Loss of Lung Tissue Elasticity in Mice Recovered from Allergic Airway Inflammation. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2017, 313, L763–L771. [Google Scholar] [CrossRef]
- Ershov, D.; Phan, M.-S.; Pylvänäinen, J.W.; Rigaud, S.U.; Blanc, L.L.; Charles-Orszag, A.; Conway, J.R.W.; Laine, R.F.; Roy, N.H.; Bonazzi, D.; et al. Bringing TrackMate into the Era of Machine-Learning and Deep-Learning. bioRxiv 2021. [Google Scholar] [CrossRef]
- Marques, L.; de Freitas Fregonezi, G.A.; Marcelino, A.A.; Teixeira Dourado, E.M., Jr.; Sarmento, A.; Pennati, F.; Aliverti, A.; Resqueti, V.R. Thoracoabdominal Asynchrony and Paradoxical Motion in Duchenne Muscular Dystrophy and Healthy Subjects during Cough: A Case Control Study. Pediatr. Pulmonol. 2021, 56, 2136–2145. [Google Scholar] [CrossRef]
- LoMauro, A.; D’Angelo, M.G.; Aliverti, A. Assessment and Management of Respiratory Function in Patients with Duchenne Muscular Dystrophy: Current and Emerging Options. Ther. Clin. Risk Manag. 2015, 11, 1475–1488. [Google Scholar] [CrossRef]
- Lo Mauro, A.; Aliverti, A. Physiology of Respiratory Disturbances in Muscular Dystrophies. Breathe 2016, 12, 318–327. [Google Scholar] [CrossRef]
- Gosselin, L.E.; Williams, J.E. Pentoxifylline Fails to Attenuate Fibrosis in Dystrophic (Mdx) Diaphragm Muscle. Muscle Nerve Off. J. Am. Assoc. Electrodiagn. Med. 2006, 33, 820–823. [Google Scholar] [CrossRef]
- Krupnick, A.S.; Zhu, J.; Nguyen, T.; Kreisel, D.; Balsara, K.R.; Lankford, E.B.; Clark, C.C.; Levine, S.; Stedman, H.H.; Shrager, J.B. Inspiratory Loading Does Not Accelerate Dystrophy in Mdx Mouse Diaphragm: Implications for Regenerative Therapy. J. Appl. Physiol. 2003, 94, 411–419. [Google Scholar] [CrossRef]
- Niebroj-Dobosz, I.; Fidzianska, A.; Glinka, Z. Comparative Studies of Hind Limb and Diaphragm Muscles of Mdx Mice. Basic Appl. Myol. 1997, 7, 381–386. [Google Scholar]
- Laws, N.; Hoey, A. Progression of Kyphosis in Mdx Mice. J. Appl. Physiol. 2004, 97, 1970–1977. [Google Scholar] [CrossRef]
- Murphy, K.H. Assessment of Respiratory Motor Units in the Mdx Mouse Model of Duchenne Muscular Dystrophy. Master’s Thesis, University College Cork, Cork, Ireland, 2021. [Google Scholar]
- O’Halloran, K. Contribution of Extra-Diaphragmatic Inspiratory Muscles to Peak Inspiratory Pressure in Wild-Type and Dystrophic (Mdx) Mice. FASEB J. 2021, 35, 831–848. [Google Scholar] [CrossRef]
- Pellegrini, M.; Hedenstierna, G.; Roneus, A.; Segelsjö, M.; Larsson, A.; Perchiazzi, G. The Diaphragm Acts as a Brake during Expiration to Prevent Lung Collapse. Am. J. Respir. Crit. Care Med. 2017, 195, 1608–1616. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, M.; Gudmundsson, M.; Bencze, R.; Segelsjö, M.; Freden, F.; Rylander, C.; Hedenstierna, G.; Larsson, A.S.; Perchiazzi, G. Expiratory Resistances Prevent Expiratory Diaphragm Contraction, Flow Limitation, and Lung Collapse. Am. J. Respir. Crit. Care Med. 2020, 201, 1218–1229. [Google Scholar] [CrossRef] [PubMed]
- Muller, N.; Volgyesi, G.; Becker, L.; Bryan, M.H.; Bryan, A.C. Diaphragmatic Muscle Tone. J. Appl. Physiol. 1979, 47, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Easton, P.A.; Katagiri, M.; Kieser, T.M.; Platt, R.S. Postinspiratory Activity of Costal and Crural Diaphragm. J. Appl. Physiol. 1999, 87, 582–589. [Google Scholar] [CrossRef]
- Roberts, N.W.; Holley-Cuthrell, J.; Gonzalez-Vega, M.; Mull, A.J.; Heydemann, A. Biochemical and Functional Comparisons of Mdx and Sgcg−/− Muscular Dystrophy Mouse Models. BioMed Res. Int. 2015, 2015, e131436. [Google Scholar] [CrossRef]
- Giordano, K.; Rodriguez, E.; Green, N.; Armani, M.; Richards, J.; Shaffer, T.H.; Attia, M.W. Pulmonary Function Tests in Emergency Department Pediatric Patients with Acute Wheezing/Asthma Exacerbation. Pulm. Med. 2012, 2012, e724139. [Google Scholar] [CrossRef]
- Rahman, T.; Page, R.; Page, C.; Bonnefoy, J.-R.; Cox, T.; Shaffer, T.H. PneuRIPTM: A Novel Respiratory Inductance Plethysmography Monitor. J. Med. Devices 2017, 11, 110101–110106. [Google Scholar] [CrossRef]
- DeVanna, J.C.; Kornegay, J.N.; Bogan, D.J.; Bogan, J.R.; Dow, J.L.; Hawkins, E.C. Respiratory Dysfunction in Unsedated Dogs with Golden Retriever Muscular Dystrophy. Neuromuscul. Disord. 2014, 24, 63–73. [Google Scholar] [CrossRef]
- Hawkins, E.C.; Bettis, A.K.; Kornegay, J.N. Expiratory Dysfunction in Young Dogs with Golden Retriever Muscular Dystrophy. Neuromuscul. Disord. NMD 2020, 30, 930–937. [Google Scholar] [CrossRef]
- Howard, Z.M.; Lowe, J.; Blatnik, A.J.; Roberts, D.; Burghes, A.H.M.; Bansal, S.S.; Rafael-Fortney, J.A. Early Inflammation in Muscular Dystrophy Differs between Limb and Respiratory Muscles and Increases with Dystrophic Severity. Am. J. Pathol. 2021, 191, 730–747. [Google Scholar] [CrossRef]
- Leith, D.E. Comparative Mammalian Respiratory Mechanics. Physiologist 1976, 19, 485–510. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Svetlove, A.; Albers, J.; Hülsmann, S.; Markus, M.A.; Zschüntzsch, J.; Alves, F.; Dullin, C. Non-Invasive Optical Motion Tracking Allows Monitoring of Respiratory Dynamics in Dystrophin-Deficient Mice. Cells 2022, 11, 918. https://doi.org/10.3390/cells11050918
Svetlove A, Albers J, Hülsmann S, Markus MA, Zschüntzsch J, Alves F, Dullin C. Non-Invasive Optical Motion Tracking Allows Monitoring of Respiratory Dynamics in Dystrophin-Deficient Mice. Cells. 2022; 11(5):918. https://doi.org/10.3390/cells11050918
Chicago/Turabian StyleSvetlove, Angelika, Jonas Albers, Swen Hülsmann, Marietta Andrea Markus, Jana Zschüntzsch, Frauke Alves, and Christian Dullin. 2022. "Non-Invasive Optical Motion Tracking Allows Monitoring of Respiratory Dynamics in Dystrophin-Deficient Mice" Cells 11, no. 5: 918. https://doi.org/10.3390/cells11050918
APA StyleSvetlove, A., Albers, J., Hülsmann, S., Markus, M. A., Zschüntzsch, J., Alves, F., & Dullin, C. (2022). Non-Invasive Optical Motion Tracking Allows Monitoring of Respiratory Dynamics in Dystrophin-Deficient Mice. Cells, 11(5), 918. https://doi.org/10.3390/cells11050918