
Browning Epicardial Adipose Tissue: Friend or Foe?

 ,
,
Abstract
:1. Introduction
2. The Epicardial Adipose Tissue
2.1. Physiological Features
2.2. EAT as an Endocrine Organ
2.3. Immune Cellular Composition of EAT
2.4. EAT in Heart Diseases
2.4.1. Coronary Artery Disease
2.4.2. Atrial Fibrillation
2.4.3. Cardiac Morphology and Function
2.4.4. EAT and COVID-19
2.5. Effect of Exercise, Weight Loss, Pharmacological Intervention on EAT
3. The Browning of Adipose Tissue
3.1. White, Brown, and Beige Adipocytes Markers
3.1.1. White Adipocyte Markers
3.1.2. Brown Adipocyte Markers
3.1.3. Beige Adipocyte Markers
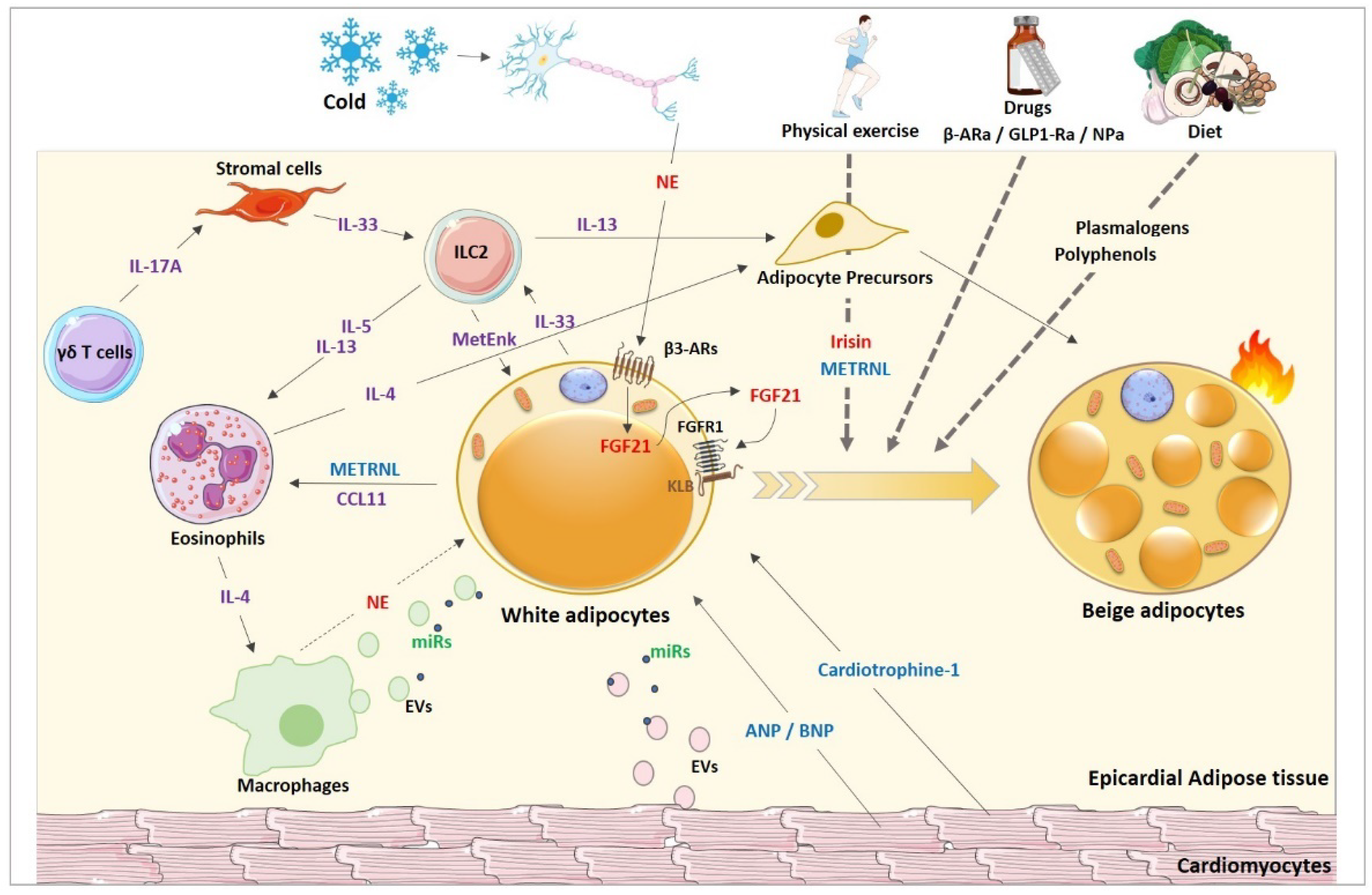
3.2. Browning Factors
3.2.1. Thermogenic Stimuli
3.2.2. Proteins Stimuli
3.2.3. Lipids Stimuli
3.2.4. Natriuretic Peptides
3.2.5. Extracellular Vesicles (EVs) and miRNA
3.2.6. Muscle
3.2.7. Immune Cells
4. Browning EAT: Friend or Foe?
4.1. Evidences of EAT Beige Phenotype
4.2. Proposed Browning Factors within the EAT
4.3. EAT Whitening as Foe
4.4. Perspectives: Non-Invasive Imaging Techniques
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Kannel, W.B.; Abbott, R.D.; Savage, D.D.; McNamara, P.M. Epidemiologic Features of Chronic Atrial Fibrillation: The Framingham Study. N. Engl. J. Med. 1982, 306, 1018–1022. [Google Scholar] [CrossRef] [PubMed]
- Gaborit, B.; Venteclef, N.; Ancel, P.; Pelloux, V.; Gariboldi, V.; Leprince, P.; Amour, J.; Hatem, S.N.; Jouve, E.; Dutour, A.; et al. Human Epicardial Adipose Tissue Has a Specific Transcriptomic Signature Depending on Its Anatomical Peri-Atrial, Peri-Ventricular, or Peri-Coronary Location. Cardiovasc. Res. 2015, 108, 62–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacks, H.S.; Fain, J.N.; Bahouth, S.W.; Ojha, S.; Frontini, A.; Budge, H.; Cinti, S.; Symonds, M.E. Adult Epicardial Fat Exhibits Beige Features. J. Clin. Endocrinol. Metab. 2013, 98, E1448–E1455. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, N.; Walden, T.B.; Shabalina, I.G.; Timmons, J.A.; Cannon, B.; Nedergaard, J. Chronic Peroxisome Proliferator-Activated Receptor Gamma (PPARgamma) Activation of Epididymally Derived White Adipocyte Cultures Reveals a Population of Thermogenically Competent, UCP1-Containing Adipocytes Molecularly Distinct from Classic Brown Adipocytes. J. Biol. Chem. 2010, 285, 7153–7164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, J.; Seale, P. Medicine. Beige Can Be Slimming. Science 2010, 328, 1113–1114. [Google Scholar] [CrossRef]
- Klingenspor, M.; Herzig, S.; Pfeifer, A. Brown Fat Develops a Brite Future. Obes. Facts 2012, 5, 890–896. [Google Scholar] [CrossRef]
- Iacobellis, G.; Corradi, D.; Sharma, A.M. Epicardial Adipose Tissue: Anatomic, Biomolecular and Clinical Relationships with the Heart. Nat. Clin. Pract. Cardiovasc. Med. 2005, 2, 536–543. [Google Scholar] [CrossRef]
- Corradi, D.; Maestri, R.; Callegari, S.; Pastori, P.; Goldoni, M.; Luong, T.V.; Bordi, C. The Ventricular Epicardial Fat Is Related to the Myocardial Mass in Normal, Ischemic and Hypertrophic Hearts. Cardiovasc. Pathol. 2004, 13, 313–316. [Google Scholar] [CrossRef]
- Rabkin, S.W. Epicardial Fat: Properties, Function and Relationship to Obesity. Obes. Rev. 2007, 8, 253–261. [Google Scholar] [CrossRef]
- Marchington, J.M.; Pond, C.M. Site-Specific Properties of Pericardial and Epicardial Adipose Tissue: The Effects of Insulin and High-Fat Feeding on Lipogenesis and the Incorporation of Fatty Acids In Vitro. Int. J. Obes. 1990, 14, 1013–1022. [Google Scholar]
- Marchington, J.M.; Mattacks, C.A.; Pond, C.M. Adipose Tissue in the Mammalian Heart and Pericardium: Structure, Foetal Development and Biochemical Properties. Comp. Biochem. Physiol. Part B Comp. Biochem. 1989, 94, 225–232. [Google Scholar] [CrossRef]
- Iacobellis, G.; Bianco, A.C. Epicardial Adipose Tissue: Emerging Physiological, Pathophysiological and Clinical Features. Trends Endocrinol. Metab. 2011, 22, 450–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caprio, M.; Antelmi, A.; Chetrite, G.; Muscat, A.; Mammi, C.; Marzolla, V.; Fabbri, A.; Zennaro, M.-C.; Fève, B. Antiadipogenic Effects of the Mineralocorticoid Receptor Antagonist Drospirenone: Potential Implications for the Treatment of Metabolic Syndrome. Endocrinology 2011, 152, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Sacks, H.S.; Fain, J.N.; Holman, B.; Cheema, P.; Chary, A.; Parks, F.; Karas, J.; Optican, R.; Bahouth, S.W.; Garrett, E.; et al. Uncoupling Protein-1 and Related Messenger Ribonucleic Acids in Human Epicardial and Other Adipose Tissues: Epicardial Fat Functioning as Brown Fat. J. Clin. Endocrinol. Metab. 2009, 94, 3611–3615. [Google Scholar] [CrossRef] [PubMed]
- Chechi, K.; Vijay, J.; Voisine, P.; Mathieu, P.; Bossé, Y.; Tchernof, A.; Grundberg, E.; Richard, D. UCP1 Expression-Associated Gene Signatures of Human Epicardial Adipose Tissue. JCI Insight 2019, 4, e123618. [Google Scholar] [CrossRef]
- Baker, A.R.; da Silva, N.F.; Quinn, D.W.; Harte, A.L.; Pagano, D.; Bonser, R.S.; Kumar, S.; McTernan, P.G. Human Epicardial Adipose Tissue Expresses a Pathogenic Profile of Adipocytokines in Patients with Cardiovascular Disease. Cardiovasc. Diabetol. 2006, 5, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fain, J.N.; Sacks, H.S.; Bahouth, S.W.; Tichansky, D.S.; Madan, A.K.; Cheema, P.S. Human Epicardial Adipokine Messenger RNAs: Comparisons of Their Expression in Substernal, Subcutaneous, and Omental Fat. Metabolism 2010, 59, 1379–1386. [Google Scholar] [CrossRef]
- Gruzdeva, O.V.; Akbasheva, O.E.; Dyleva, Y.A.; Antonova, L.V.; Matveeva, V.G.; Uchasova, E.G.; Fanaskova, E.V.; Karetnikova, V.N.; Ivanov, S.V.; Barbarash, O.L. Adipokine and Cytokine Profiles of Epicardial and Subcutaneous Adipose Tissue in Patients with Coronary Heart Disease. Bull. Exp. Biol. Med. 2017, 163, 608–611. [Google Scholar] [CrossRef]
- Gruzdeva, O.; Uchasova, E.; Dyleva, Y.; Borodkina, D.; Akbasheva, O.; Antonova, L.; Matveeva, V.; Belik, E.; Ivanov, S.; Sotnikov, A.; et al. Adipocytes Directly Affect Coronary Artery Disease Pathogenesis via Induction of Adipokine and Cytokine Imbalances. Front. Immunol. 2019, 10, 2163. [Google Scholar] [CrossRef]
- Sacks, H.S.; Fain, J.N. Human Epicardial Adipose Tissue: A Review. Am. Heart J. 2007, 153, 907–917. [Google Scholar] [CrossRef]
- Iacobellis, G. Epicardial Adipose Tissue in Endocrine and Metabolic Diseases. Endocrine 2014, 46, 8–15. [Google Scholar] [CrossRef]
- Zhuang, X.; Wu, B.; Li, J.; Shi, H.; Jin, B.; Luo, X. The Emerging Role of Interleukin-37 in Cardiovascular Diseases. Immun. Inflamm. Dis. 2017, 5, 373–379. [Google Scholar] [CrossRef]
- Butcovan, D.; Mocanu, V.; Timofte, D.V.; Costan, V.V.; Danila, R.; Veselin, A.P.; Ciuntu, B.M.; Haliga, R.E.; Sascau, R.A.; Ghiga, G.; et al. Macrophage Accumulation and Angiogenesis in Epicardial Adipose Tissue in Cardiac Patients with or without Chronic Heart Failure. Appl. Sci. 2020, 10, 5871. [Google Scholar] [CrossRef]
- Gaborit, B.; Sengenes, C.; Ancel, P.; Jacquier, A.; Dutour, A. Role of Epicardial Adipose Tissue in Health and Disease: A Matter of Fat? Compr. Physiol. 2017, 7, 1051–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaihov-Teper, O.; Ram, E.; Ballan, N.; Brzezinski, R.Y.; Naftali-Shani, N.; Masoud, R.; Ziv, T.; Lewis, N.; Schary, Y.; Levin-Kotler, L.-P.; et al. Extracellular Vesicles from Epicardial Fat Facilitate Atrial Fibrillation. Circulation 2021, 143, 2475–2493. [Google Scholar] [CrossRef] [PubMed]
- Dutour, A.; Achard, V.; Sell, H.; Naour, N.; Collart, F.; Gaborit, B.; Silaghi, A.; Eckel, J.; Alessi, M.-C.; Henegar, C.; et al. Secretory Type II Phospholipase A2 Is Produced and Secreted by Epicardial Adipose Tissue and Overexpressed in Patients with Coronary Artery Disease. J. Clin. Endocrinol. Metab. 2010, 95, 963–967. [Google Scholar] [CrossRef] [Green Version]
- Silaghi, A.; Achard, V.; Paulmyer-Lacroix, O.; Scridon, T.; Tassistro, V.; Duncea, I.; Clément, K.; Dutour, A.; Grino, M. Expression of Adrenomedullin in Human Epicardial Adipose Tissue: Role of Coronary Status. Am. J. Physiol.-Endocrinol. Metab. 2007, 293, E1443–E1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacks, H.S.; Fain, J.N.; Cheema, P.; Bahouth, S.W.; Garrett, E.; Wolf, R.Y.; Wolford, D.; Samaha, J. Inflammatory Genes in Epicardial Fat Contiguous with Coronary Atherosclerosis in the Metabolic Syndrome and Type 2 Diabetes: Changes Associated with Pioglitazone. Diabetes Care 2011, 34, 730–733. [Google Scholar] [CrossRef] [Green Version]
- Cherian, S.; Lopaschuk, G.D.; Carvalho, E. Cellular Cross-Talk between Epicardial Adipose Tissue and Myocardium in Relation to the Pathogenesis of Cardiovascular Disease. Am. J. Physiol.-Endocrinol. Metab. 2012, 303, E937–E949. [Google Scholar] [CrossRef] [Green Version]
- Gaborit, B.; Abdesselam, I.; Dutour, A. Epicardial Fat: More than Just an “Epi” Phenomenon? Horm. Metab. Res. 2013, 45, 991–1001. [Google Scholar] [CrossRef]
- Cheng, K.-H.; Chu, C.-S.; Lee, K.-T.; Lin, T.-H.; Hsieh, C.-C.; Chiu, C.-C.; Voon, W.-C.; Sheu, S.-H.; Lai, W.-T. Adipocytokines and Proinflammatory Mediators from Abdominal and Epicardial Adipose Tissue in Patients with Coronary Artery Disease. Int. J. Obes. 2008, 32, 268–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpato, S.; Guralnik, J.M.; Ferrucci, L.; Balfour, J.; Chaves, P.; Fried, L.P.; Harris, T.B. Cardiovascular Disease, Interleukin-6, and Risk of Mortality in Older Women: The Women’s Health and Aging Study. Circulation 2001, 103, 947–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, R. The Role of TNF in Cardiovascular Disease. Pharmacol. Res. 1999, 40, 97–105. [Google Scholar] [CrossRef]
- Akoumianakis, I.; Antoniades, C. The Interplay between Adipose Tissue and the Cardiovascular System: Is Fat Always Bad? Cardiovasc. Res. 2017, 113, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulos, A.S.; Margaritis, M.; Verheule, S.; Recalde, A.; Sanna, F.; Herdman, L.; Psarros, C.; Nasrallah, H.; Coutinho, P.; Akoumianakis, I.; et al. Mutual Regulation of Epicardial Adipose Tissue and Myocardial Redox State by PPAR-γ/Adiponectin Signalling. Circ. Res. 2016, 118, 842–855. [Google Scholar] [CrossRef]
- Camarena, V.; Sant, D.; Mohseni, M.; Salerno, T.; Zaleski, M.L.; Wang, G.; Iacobellis, G. Novel Atherogenic Pathways from the Differential Transcriptome Analysis of Diabetic Epicardial Adipose Tissue. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 739–750. [Google Scholar] [CrossRef]
- Clément, K.; Basdevant, A.; Dutour, A. Weight of Pericardial Fat on Coronaropathy. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 615–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karastergiou, K.; Evans, I.; Ogston, N.; Miheisi, N.; Nair, D.; Kaski, J.-C.; Jahangiri, M.; Mohamed-Ali, V. Epicardial Adipokines in Obesity and Coronary Artery Disease Induce Atherogenic Changes in Monocytes and Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1340–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenney, M.L.; Schultz, K.A.; Boyd, J.H.; Byrd, J.P.; Alloosh, M.; Teague, S.D.; Arce-Esquivel, A.A.; Fain, J.N.; Laughlin, M.H.; Sacks, H.S.; et al. Epicardial Adipose Excision Slows the Progression of Porcine Coronary Atherosclerosis. J. Cardiothorac. Surg. 2014, 9, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venteclef, N.; Guglielmi, V.; Balse, E.; Gaborit, B.; Cotillard, A.; Atassi, F.; Amour, J.; Leprince, P.; Dutour, A.; Clément, K.; et al. Human Epicardial Adipose Tissue Induces Fibrosis of the Atrial Myocardium through the Secretion of Adipo-Fibrokines. Eur. Heart J. 2015, 36, 795–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazurek, T.; Zhang, L.; Zalewski, A.; Mannion, J.D.; Diehl, J.T.; Arafat, H.; Sarov-Blat, L.; O’Brien, S.; Keiper, E.A.; Johnson, A.G.; et al. Human Epicardial Adipose Tissue Is a Source of Inflammatory Mediators. Circulation 2003, 108, 2460–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, S.; Merlin, J.; Lee, M.K.S.; Murphy, A.J.; Guinamard, R.R. Biology and Function of Adipose Tissue Macrophages, Dendritic Cells and B Cells. Atherosclerosis 2018, 271, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the Full Spectrum of Macrophage Activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Tordjman, J.; Poitou, C.; Darakhshan, F.; Hugol, D.; Basdevant, A.; Aissat, A.; Guerre-Millo, M.; Clément, K. Human Adipose Tissue Macrophages: M1 and M2 Cell Surface Markers in Subcutaneous and Omental Depots and after Weight Loss. J. Clin. Endocrinol. Metab. 2009, 94, 4619–4623. [Google Scholar] [CrossRef] [Green Version]
- Hirata, Y.; Tabata, M.; Kurobe, H.; Motoki, T.; Akaike, M.; Nishio, C.; Higashida, M.; Mikasa, H.; Nakaya, Y.; Takanashi, S.; et al. Coronary Atherosclerosis Is Associated with Macrophage Polarization in Epicardial Adipose Tissue. J. Am. Coll. Cardiol. 2011, 58, 248–255. [Google Scholar] [CrossRef] [Green Version]
- Vianello, E.; Dozio, E.; Arnaboldi, F.; Marazzi, M.G.; Martinelli, C.; Lamont, J.; Tacchini, L.; Sigrüner, A.; Schmitz, G.; Corsi Romanelli, M.M. Epicardial Adipocyte Hypertrophy: Association with M1-Polarization and Toll-like Receptor Pathways in Coronary Artery Disease Patients. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Gurses, K.M.; Ozmen, F.; Kocyigit, D.; Yersal, N.; Bilgic, E.; Kaya, E.; Kopru, C.Z.; Soyal, T.; Doganci, S.; Tokgozoglu, L.; et al. Netrin-1 Is Associated with Macrophage Infiltration and Polarization in Human Epicardial Adipose Tissue in Coronary Artery Disease. J. Cardiol. 2017, 69, 851–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, F.; Niiro, H.; Iino, T.; Yoshida, S.; Saito, N.; Onohara, S.; Miyamoto, T.; Minagawa, H.; Fujii, S.-I.; Shultz, L.D.; et al. The Developmental Program of Human Dendritic Cells Is Operated Independently of Conventional Myeloid and Lymphoid Pathways. Blood 2007, 110, 3591–3660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanovic-Racic, M.; Yang, X.; Turner, M.S.; Mantell, B.S.; Stolz, D.B.; Sumpter, T.L.; Sipula, I.J.; Dedousis, N.; Scott, D.K.; Morel, P.A.; et al. Dendritic Cells Promote Macrophage Infiltration and Comprise a Substantial Proportion of Obesity-Associated Increases in CD11c+ Cells in Adipose Tissue and Liver. Diabetes 2012, 61, 2330–2339. [Google Scholar] [CrossRef] [Green Version]
- Mráz, M.; Cinkajzlová, A.; Kloučková, J.; Lacinová, Z.; Kratochvílová, H.; Lipš, M.; Pořízka, M.; Kopecký, P.; Lindner, J.; Kotulák, T.; et al. Dendritic Cells in Subcutaneous and Epicardial Adipose Tissue of Subjects with Type 2 Diabetes, Obesity, and Coronary Artery Disease. Mediat. Inflamm. 2019, 2019, 5481725. [Google Scholar] [CrossRef]
- Divoux, A.; Moutel, S.; Poitou, C.; Lacasa, D.; Veyrie, N.; Aissat, A.; Arock, M.; Guerre-Millo, M.; Clément, K. Mast Cells in Human Adipose Tissue: Link with Morbid Obesity, Inflammatory Status, and Diabetes. J. Clin. Endocrinol. Metab. 2012, 97, E1677–E1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandstadter, J.D.; Yang, Y. Natural Killer Cell Responses to Viral Infection. J. Innate Immun. 2011, 3, 274–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonamichi, B.D.S.F.; Lee, J. Unusual Suspects in the Development of Obesity-Induced Inflammation and Insulin Resistance: NK Cells, INKT Cells, and ILCs. Diabetes Metab. J. 2017, 41, 229–250. [Google Scholar] [CrossRef] [Green Version]
- Roda, J.M.; Parihar, R.; Magro, C.; Nuovo, G.J.; Tridandapani, S.; Carson, W.E. Natural Killer Cells Produce T Cell-Recruiting Chemokines in Response to Antibody-Coated Tumor Cells. Cancer Res. 2006, 66, 517–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bendelac, A.; Savage, P.B.; Teyton, L. The Biology of NKT Cells. Annu. Rev. Immunol. 2007, 25, 297–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, L.; Michelet, X.; Zhang, S.; Brennan, P.J.; Moseman, A.; Lester, C.; Besra, G.; Vomhof-Dekrey, E.E.; Tighe, M.; Koay, H.-F.; et al. Regulatory INKT Cells Lack Expression of the Transcription Factor PLZF and Control the Homeostasis of T(Reg) Cells and Macrophages in Adipose Tissue. Nat. Immunol. 2015, 16, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Brestoff, J.R.; Kim, B.S.; Saenz, S.A.; Stine, R.R.; Monticelli, L.A.; Sonnenberg, G.F.; Thome, J.J.; Farber, D.L.; Lutfy, K.; Seale, P.; et al. Group 2 Innate Lymphoid Cells Promote Beiging of Adipose and Limit Obesity. Nature 2015, 519, 242–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molofsky, A.B.; Nussbaum, J.C.; Liang, H.-E.; Van Dyken, S.J.; Cheng, L.E.; Mohapatra, A.; Chawla, A.; Locksley, R.M. Innate Lymphoid Type 2 Cells Sustain Visceral Adipose Tissue Eosinophils and Alternatively Activated Macrophages. J. Exp. Med. 2013, 210, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Nussbaum, J.C.; Van Dyken, S.J.; von Moltke, J.; Cheng, L.E.; Mohapatra, A.; Molofsky, A.B.; Thornton, E.E.; Krummel, M.F.; Chawla, A.; Liang, H.-E.; et al. Type 2 Innate Lymphoid Cells Control Eosinophil Homeostasis. Nature 2013, 502, 245–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohlgruber, A.C.; Gal-Oz, S.T.; LaMarche, N.M.; Shimazaki, M.; Duquette, D.; Koay, H.-F.; Nguyen, H.N.; Mina, A.I.; Paras, T.; Tavakkoli, A.; et al. Γδ T Cells Producing Interleukin-17A Regulate Adipose Regulatory T Cell Homeostasis and Thermogenesis. Nat. Immunol. 2018, 19, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Eto, K.; Yamashita, H.; Ohsugi, M.; Otsu, M.; Hara, K.; Ueki, K.; Sugiura, S.; et al. CD8+ Effector T Cells Contribute to Macrophage Recruitment and Adipose Tissue Inflammation in Obesity. Nat. Med. 2009, 15, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Ying, W.; Wollam, J.; Ofrecio, J.M.; Bandyopadhyay, G.; El Ouarrat, D.; Lee, Y.S.; Oh, D.Y.; Li, P.; Osborn, O.; Olefsky, J.M. Adipose Tissue B2 Cells Promote Insulin Resistance through Leukotriene LTB4/LTB4R1 Signaling. J. Clin. Invest. 2017, 127, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Mráz, M.; Cinkajzlová, A.; Kloučková, J.; Lacinová, Z.; Kratochvílová, H.; Lipš, M.; Pořízka, M.; Kopecký, P.; Pierzynová, A.; Kučera, T.; et al. Coronary Artery Disease Is Associated with an Increased Amount of T Lymphocytes in Human Epicardial Adipose Tissue. Mediat. Inflamm. 2019, 2019, 4075086. [Google Scholar] [CrossRef]
- Silaghi, A.; Piercecchi-Marti, M.-D.; Grino, M.; Leonetti, G.; Alessi, M.C.; Clement, K.; Dadoun, F.; Dutour, A. Epicardial Adipose Tissue Extent: Relationship with Age, Body Fat Distribution, and Coronaropathy. Obesity 2008, 16, 2424–2430. [Google Scholar] [CrossRef]
- Greif, M.; Becker, A.; von Ziegler, F.; Lebherz, C.; Lehrke, M.; Broedl, U.C.; Tittus, J.; Parhofer, K.; Becker, C.; Reiser, M.; et al. Pericardial Adipose Tissue Determined by Dual Source CT Is a Risk Factor for Coronary Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 781–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Hsu, F.-C.; Harris, T.B.; Liu, Y.; Kritchevsky, S.B.; Szklo, M.; Ouyang, P.; Espeland, M.A.; Lohman, K.K.; Criqui, M.H.; et al. The Association of Pericardial Fat with Incident Coronary Heart Disease: The Multi-Ethnic Study of Atherosclerosis (MESA). Am. J. Clin. Nutr. 2009, 90, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Hell, M.M.; Ding, X.; Rubeaux, M.; Slomka, P.; Gransar, H.; Terzopoulos, D.; Hayes, S.; Marwan, M.; Achenbach, S.; Berman, D.S.; et al. Epicardial Adipose Tissue Volume but Not Density Is an Independent Predictor for Myocardial Ischemia. J. Cardiovasc. Comput. Tomogr. 2016, 10, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Shan, D.; Wang, X.; Dou, G.; Zhang, W.; Jing, J.; He, B.; Li, Y.; Chen, Y.; Yang, J. Vascular-Specific Epicardial Adipose Tissue in Predicting Functional Myocardial Ischemia for Patients with Stable Chest Pain. J. Thromb. Thrombolysis 2021, 51, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Mahabadi, A.A.; Berg, M.H.; Lehmann, N.; Kälsch, H.; Bauer, M.; Kara, K.; Dragano, N.; Moebus, S.; Jöckel, K.-H.; Erbel, R.; et al. Association of Epicardial Fat with Cardiovascular Risk Factors and Incident Myocardial Infarction in the General Population: The Heinz Nixdorf Recall Study. J. Am. Coll. Cardiol. 2013, 61, 1388–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaborit, B.; Kober, F.; Jacquier, A.; Moro, P.J.; Flavian, A.; Quilici, J.; Cuisset, T.; Simeoni, U.; Cozzone, P.; Alessi, M.-C.; et al. Epicardial Fat Volume Is Associated with Coronary Microvascular Response in Healthy Subjects: A Pilot Study. Obesity 2012, 20, 1200–1205. [Google Scholar] [CrossRef] [PubMed]
- Bachar, G.N.; Dicker, D.; Kornowski, R.; Atar, E. Epicardial Adipose Tissue as a Predictor of Coronary Artery Disease in Asymptomatic Subjects. Am. J. Cardiol. 2012, 110, 534–538. [Google Scholar] [CrossRef] [PubMed]
- Yerramasu, A.; Dey, D.; Venuraju, S.; Anand, D.V.; Atwal, S.; Corder, R.; Berman, D.S.; Lahiri, A. Increased Volume of Epicardial Fat Is an Independent Risk Factor for Accelerated Progression of Sub-Clinical Coronary Atherosclerosis. Atherosclerosis 2012, 220, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Hirata, Y.; Kurobe, H.; Akaike, M.; Chikugo, F.; Hori, T.; Bando, Y.; Nishio, C.; Higashida, M.; Nakaya, Y.; Kitagawa, T.; et al. Enhanced Inflammation in Epicardial Fat in Patients with Coronary Artery Disease. Int. Heart J. 2011, 52, 139–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dozio, E.; Vianello, E.; Briganti, S.; Fink, B.; Malavazos, A.E.; Scognamiglio, E.T.; Dogliotti, G.; Sigrüener, A.; Schmitz, G.; Romanelli, M.M.C. Increased Reactive Oxygen Species Production in Epicardial Adipose Tissues from Coronary Artery Disease Patients Is Associated with Brown-to-White Adipocyte Trans-Differentiation. Int. J. Cardiol. 2014, 174, 413–414. [Google Scholar] [CrossRef]
- Fioranelli, M.; Bottaccioli, A.G.; Bottaccioli, F.; Bianchi, M.; Rovesti, M.; Roccia, M.G. Stress and Inflammation in Coronary Artery Disease: A Review Psychoneuroendocrineimmunology-Based. Front. Immunol. 2018, 9, 2031. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Somoza, A.; Teijeira-Fernández, E.; Fernández, A.L.; González-Juanatey, J.R.; Eiras, S. Proteomic Analysis of Epicardial and Subcutaneous Adipose Tissue Reveals Differences in Proteins Involved in Oxidative Stress. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H202–H209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prati, F.; Arbustini, E.; Labellarte, A.; Sommariva, L.; Pawlowski, T.; Manzoli, A.; Pagano, A.; Motolese, M.; Boccanelli, A. Eccentric Atherosclerotic Plaques with Positive Remodelling Have a Pericardial Distribution: A Permissive Role of Epicardial Fat? A Three-Dimensional Intravascular Ultrasound Study of Left Anterior Descending Artery Lesions. Eur. Heart J. 2003, 24, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Oikonomou, E.K.; Williams, M.C.; Kotanidis, C.P.; Desai, M.Y.; Marwan, M.; Antonopoulos, A.S.; Thomas, K.E.; Thomas, S.; Akoumianakis, I.; Fan, L.M.; et al. A Novel Machine Learning-Derived Radiotranscriptomic Signature of Perivascular Fat Improves Cardiac Risk Prediction Using Coronary CT Angiography. Eur. Heart J. 2019, 40, 3529–3543. [Google Scholar] [CrossRef]
- Oikonomou, E.K.; Antonopoulos, A.S.; Schottlander, D.; Marwan, M.; Mathers, C.; Tomlins, P.; Siddique, M.; Klüner, L.V.; Shirodaria, C.; Mavrogiannis, M.C.; et al. Standardized Measurement of Coronary Inflammation Using Cardiovascular Computed Tomography: Integration in Clinical Care as a Prognostic Medical Device. Cardiovasc. Res. 2021, 117, 2677–2690. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.X.; Abed, H.S.; Molaee, P.; Nelson, A.J.; Brooks, A.G.; Sharma, G.; Leong, D.P.; Lau, D.H.; Middeldorp, M.E.; Roberts-Thomson, K.C.; et al. Pericardial Fat Is Associated with Atrial Fibrillation Severity and Ablation Outcome. J. Am. Coll. Cardiol. 2011, 57, 1745–1751. [Google Scholar] [CrossRef]
- Nagashima, K.; Okumura, Y.; Watanabe, I.; Nakai, T.; Ohkubo, K.; Kofune, T.; Kofune, M.; Mano, H.; Sonoda, K.; Hirayama, A. Association between Epicardial Adipose Tissue Volumes on 3-Dimensional Reconstructed CT Images and Recurrence of Atrial Fibrillation after Catheter Ablation. Circ. J. 2011, 75, 2559–2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, K.; Fukuda, S.; Tanaka, A.; Otsuka, K.; Sakamoto, M.; Taguchi, H.; Yoshikawa, J.; Shimada, K.; Yoshiyama, M. Peri-Atrial Epicardial Adipose Tissue Is Associated with New-Onset Nonvalvular Atrial Fibrillation. Circ. J. 2012, 76, 2748–2754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.-I.; Kim, B.-J.; Cha, T.-J.; Heo, J.-H.; Kim, H.-S.; Lee, J.-W. Impact of Duration and Dosage of Statin Treatment and Epicardial Fat Thickness on the Recurrence of Atrial Fibrillation after Electrical Cardioversion. Heart Vessel. 2015, 30, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Thanassoulis, G.; Massaro, J.M.; O’Donnell, C.J.; Hoffmann, U.; Levy, D.; Ellinor, P.T.; Wang, T.J.; Schnabel, R.B.; Vasan, R.S.; Fox, C.S.; et al. Pericardial Fat Is Associated with Prevalent Atrial Fibrillation: The Framingham Heart Study. Circ. Arrhythm. Electrophysiol. 2010, 3, 345–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Chekakie, M.O.; Welles, C.C.; Metoyer, R.; Ibrahim, A.; Shapira, A.R.; Cytron, J.; Santucci, P.; Wilber, D.J.; Akar, J.G. Pericardial Fat Is Independently Associated with Human Atrial Fibrillation. J. Am. Coll. Cardiol. 2010, 56, 784–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frustaci, A.; Chimenti, C.; Bellocci, F.; Morgante, E.; Russo, M.A.; Maseri, A. Histological Substrate of Atrial Biopsies in Patients with Lone Atrial Fibrillation. Circulation 1997, 96, 1180–1184. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Sekiguchi, A.; Iwasaki, Y.; Date, T.; Sagara, K.; Tanabe, H.; Suma, H.; Sawada, H.; Aizawa, T. Recruitment of Immune Cells across Atrial Endocardium in Human Atrial Fibrillation. Circ. J. 2010, 74, 262–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatem, S.N.; Sanders, P. Epicardial Adipose Tissue and Atrial Fibrillation. Cardiovasc. Res. 2014, 102, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Poggi, A.; Gaborit, B.; Schindler, T.; Liberale, L.; Montecucco, F.; Carbone, F. Epicardial Fat and Atrial Fibrillation: The Perils of Atrial Failure. Europace, 2022; Accepted for Publication. [Google Scholar]
- Iacobellis, G.; Leonetti, F.; Singh, N.; Sharma, A.M. Relationship of Epicardial Adipose Tissue with Atrial Dimensions and Diastolic Function in Morbidly Obese Subjects. Int. J. Cardiol. 2007, 115, 272–273. [Google Scholar] [CrossRef] [PubMed]
- Iacobellis, G.; Ribaudo, M.C.; Zappaterreno, A.; Iannucci, C.V.; Leonetti, F. Relation between Epicardial Adipose Tissue and Left Ventricular Mass. Am. J. Cardiol. 2004, 94, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Crendal, E.; Dutheil, F.; Naughton, G.; McDonald, T.; Obert, P. Increased Myocardial Dysfunction, Dyssynchrony, and Epicardial Fat across the Lifespan in Healthy Males. BMC Cardiovasc. Disord. 2014, 14, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, L.; Binkley, C.M.; Suever, J.D.; Umasankar, N.; Haggerty, C.M.; Rich, J.; Wehner, G.J.; Hamlet, S.M.; Powell, D.K.; Radulescu, A.; et al. Cardiac Remodeling and Dysfunction in Childhood Obesity: A Cardiovascular Magnetic Resonance Study. J. Cardiovasc. Magn. Reson. 2016, 18, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasbleiz, A.; Gaborit, B.; Soghomonian, A.; Bartoli, A.; Ancel, P.; Jacquier, A.; Dutour, A. COVID-19 and Obesity: Role of Ectopic Visceral and Epicardial Adipose Tissues in Myocardial Injury. Front. Endocrinol. 2021, 12, 726967. [Google Scholar] [CrossRef] [PubMed]
- Bihan, H.; Heidar, R.; Beloeuvre, A.; Allard, L.; Ouedraogo, E.; Tatulashvili, S.; Tandjaoui, Y.; Gaudry, S.; Brillet, P.-Y.; Cosson, E. Epicardial Adipose Tissue and Severe Coronavirus Disease 19. Cardiovasc. Diabetol. 2021, 20, 147. [Google Scholar] [CrossRef] [PubMed]
- Iacobellis, G.; Secchi, F.; Capitanio, G.; Basilico, S.; Schiaffino, S.; Boveri, S.; Sardanelli, F.; Corsi Romanelli, M.M.; Malavazos, A.E. Epicardial Fat Inflammation in Severe COVID-19. Obesity 2020, 28, 2260–2262. [Google Scholar] [CrossRef] [PubMed]
- Iacobellis, G.; Malavazos, A.E.; Basilico, S.; Tresoldi, S.; Rinaldo, R.F.; Dubini, C.; Capitanio, G.; Serpi, F.; Schiaffino, S.; Oliva, O.A.; et al. Epicardial Fat Inflammation Response to COVID-19 Therapies. Obesity 2021, 29, 1427–1433. [Google Scholar] [CrossRef] [PubMed]
- Malavazos, A.E.; Goldberger, J.J.; Iacobellis, G. Does Epicardial Fat Contribute to COVID-19 Myocardial Inflammation? Eur. Heart J. 2020, 41, 2333. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Qin, M.; Cai, Y.; Liu, T.; Shen, B.; Yang, F.; Cao, S.; Liu, X.; Xiang, Y.; Zhao, Q.; et al. Characteristics and Clinical Significance of Myocardial Injury in Patients with Severe Coronavirus Disease 2019. Eur. Heart J. 2020, 41, 2070–2079. [Google Scholar] [CrossRef] [PubMed]
- Gaborit, B.; Jacquier, A.; Kober, F.; Abdesselam, I.; Cuisset, T.; Boullu-Ciocca, S.; Emungania, O.; Alessi, M.-C.; Clément, K.; Bernard, M.; et al. Effects of Bariatric Surgery on Cardiac Ectopic Fat: Lesser Decrease in Epicardial Fat Compared to Visceral Fat Loss and No Change in Myocardial Triglyceride Content. J. Am. Coll. Cardiol. 2012, 60, 1381–1389. [Google Scholar] [CrossRef] [PubMed]
- Dutour, A.; Abdesselam, I.; Ancel, P.; Kober, F.; Mrad, G.; Darmon, P.; Ronsin, O.; Pradel, V.; Lesavre, N.; Martin, J.C.; et al. Exenatide Decreases Liver Fat Content and Epicardial Adipose Tissue in Patients with Obesity and Type 2 Diabetes: A Prospective Randomized Clinical Trial Using Magnetic Resonance Imaging and Spectroscopy. Diabetes Obes. Metab. 2016, 18, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Saco-Ledo, G.; Valenzuela, P.L.; Castillo-García, A.; Arenas, J.; León-Sanz, M.; Ruilope, L.M.; Lucia, A. Physical Exercise and Epicardial Adipose Tissue: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Obes. Rev. 2021, 22, e13103. [Google Scholar] [CrossRef] [PubMed]
- Launbo, N.; Zobel, E.H.; von Scholten, B.J.; Faerch, K.; Jørgensen, P.G.; Christensen, R.H. Targeting Epicardial Adipose Tissue with Exercise, Diet, Bariatric Surgery or Pharmaceutical Interventions: A Systematic Review and Meta-Analysis. Obes. Rev. 2021, 22, e13136. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-H.; Park, Y.S.; Kim, Y.J.; Lee, I.S.; Kim, J.H.; Lee, J.-H.; Choi, S.W.; Jeong, J.-O.; Seong, I.-W. Effects of Statins on the Epicardial Fat Thickness in Patients with Coronary Artery Stenosis Underwent Percutaneous Coronary Intervention: Comparison of Atorvastatin with Simvastatin/Ezetimibe. J. Cardiovasc. Ultrasound 2010, 18, 121–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Aizawa, Y.; Yuasa, S.; Kishi, S.; Fuse, K.; Fujita, S.; Ikeda, Y.; Kitazawa, H.; Takahashi, M.; Sato, M.; et al. The Effect of Dapagliflozin Treatment on Epicardial Adipose Tissue Volume. Cardiovasc. Diabetol. 2018, 17, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Aizawa, Y.; Yuasa, S.; Fujita, S.; Ikeda, Y.; Okabe, M. The Effect of Dapagliflozin Treatment on Epicardial Adipose Tissue Volume and P-Wave Indices: An Ad-Hoc Analysis of The Previous Randomized Clinical Trial. J. Atheroscler. Thromb. 2020, 27, 1348–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaborit, B.; Ancel, P.; Abdullah, A.E.; Maurice, F.; Abdesselam, I.; Calen, A.; Soghomonian, A.; Houssays, M.; Varlet, I.; Eisinger, M.; et al. Effect of Empagliflozin on Ectopic Fat Stores and Myocardial Energetics in Type 2 Diabetes: The EMPACEF Study. Cardiovasc. Diabetol. 2021, 20, 57. [Google Scholar] [CrossRef] [PubMed]
- Jonker, J.T.; de Mol, P.; de Vries, S.T.; Widya, R.L.; Hammer, S.; van Schinkel, L.D.; van der Meer, R.W.; Gans, R.O.B.; Webb, A.G.; Kan, H.E.; et al. Exercise and Type 2 Diabetes Mellitus: Changes in Tissue-Specific Fat Distribution and Cardiac Function. Radiology 2013, 269, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Parisi, V.; Petraglia, L.; D’Esposito, V.; Cabaro, S.; Rengo, G.; Caruso, A.; Grimaldi, M.G.; Baldascino, F.; De Bellis, A.; Vitale, D.; et al. Statin Therapy Modulates Thickness and Inflammatory Profile of Human Epicardial Adipose Tissue. Int. J. Cardiol. 2019, 274, 326–330. [Google Scholar] [CrossRef] [Green Version]
- Kotanidis, C.P.; Antoniades, C. Perivascular Fat Imaging by Computed Tomography (CT): A Virtual Guide. Br. J. Pharmacol. 2021, 178, 4270–4290. [Google Scholar] [CrossRef]
- Elnabawi, Y.A.; Oikonomou, E.K.; Dey, A.K.; Mancio, J.; Rodante, J.A.; Aksentijevich, M.; Choi, H.; Keel, A.; Erb-Alvarez, J.; Teague, H.L.; et al. Association of Biologic Therapy with Coronary Inflammation in Patients with Psoriasis as Assessed by Perivascular Fat Attenuation Index. JAMA Cardiol. 2019, 4, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Zechner, R.; Zimmermann, R.; Eichmann, T.O.; Kohlwein, S.D.; Haemmerle, G.; Lass, A.; Madeo, F. FAT SIGNALS—Lipases and Lipolysis in Lipid Metabolism and Signaling. Cell. Metab. 2012, 15, 279–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballinger, M.A.; Andrews, M.T. Nature’s Fat-Burning Machine: Brown Adipose Tissue in a Hibernating Mammal. J. Exp. Biol. 2018, 221, jeb162586. [Google Scholar] [CrossRef] [Green Version]
- Burlington, R.F.; Therriault, D.G.; Hubbard, R.W. Lipid Changes in Isolated Brown Fat Cells from Hibernating and Aroused Thirteen-Lined Ground Squirrels (Citellus Tridecemlineatus). Comp. Biochem. Physiol. 1969, 29, 431–437. [Google Scholar] [CrossRef]
- Barger, J.L.; Barnes, B.M.; Boyer, B.B. Regulation of UCP1 and UCP3 in Arctic Ground Squirrels and Relation with Mitochondrial Proton Leak. J. Appl. Physiol. 2006, 101, 339–347. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Burman, A.; Nichols, C.; Alila, L.; Showe, L.C.; Showe, M.K.; Boyer, B.B.; Barnes, B.M.; Marr, T.G. Detection of Differential Gene Expression in Brown Adipose Tissue of Hibernating Arctic Ground Squirrels with Mouse Microarrays. Physiol. Genom. 2006, 25, 346–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malatesta, M.; Battistelli, S.; Rocchi, M.B.; Zancanaro, C.; Fakan, S.; Gazzanelli, G. Fine Structural Modifications of Liver, Pancreas and Brown Adipose Tissue Mitochondria from Hibernating, Arousing and Euthermic Dormice. Cell Biol. Int. 2001, 25, 131–138. [Google Scholar] [CrossRef]
- Bartelt, A.; Heeren, J. Adipose Tissue Browning and Metabolic Health. Nat. Rev. Endocrinol. 2014, 10, 24–36. [Google Scholar] [CrossRef]
- Cinti, S. The Adipose Organ at a Glance. Dis. Model Mech. 2012, 5, 588–594. [Google Scholar] [CrossRef] [Green Version]
- Cannon, B.; Nedergaard, J. Brown Adipose Tissue: Function and Physiological Significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- Lowell, B.B.; Spiegelman, B.M. Towards a Molecular Understanding of Adaptive Thermogenesis. Nature 2000, 404, 652–660. [Google Scholar] [CrossRef]
- Nicholls, D.G.; Locke, R.M. Thermogenic Mechanisms in Brown Fat. Physiol. Rev. 1984, 64, 1–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nedergaard, J.; Golozoubova, V.; Matthias, A.; Asadi, A.; Jacobsson, A.; Cannon, B. UCP1: The Only Protein Able to Mediate Adaptive Non-Shivering Thermogenesis and Metabolic Inefficiency. Biochim. Biophys. Acta 2001, 1504, 82–106. [Google Scholar] [CrossRef] [Green Version]
- Nedergaard, J.; Bengtsson, T.; Cannon, B. Unexpected Evidence for Active Brown Adipose Tissue in Adult Humans. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E444–E452. [Google Scholar] [CrossRef] [PubMed]
- Cypess, A.M.; Lehman, S.; Williams, G.; Tal, I.; Rodman, D.; Goldfine, A.B.; Kuo, F.C.; Palmer, E.L.; Tseng, Y.-H.; Doria, A.; et al. Identification and Importance of Brown Adipose Tissue in Adult Humans. N. Engl. J. Med. 2009, 360, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- Young, P.; Arch, J.R.; Ashwell, M. Brown Adipose Tissue in the Parametrial Fat Pad of the Mouse. FEBS Lett. 1984, 167, 10–14. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.-H.; Khandekar, M.; Nuutila, P.; Schaart, G.; Huang, K.; et al. Beige Adipocytes Are a Distinct Type of Thermogenic Fat Cell in Mouse and Human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Barbatelli, G.; Murano, I.; Madsen, L.; Hao, Q.; Jimenez, M.; Kristiansen, K.; Giacobino, J.P.; De Matteis, R.; Cinti, S. The Emergence of Cold-Induced Brown Adipocytes in Mouse White Fat Depots Is Determined Predominantly by White to Brown Adipocyte Transdifferentiation. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E1244–E1253. [Google Scholar] [CrossRef] [Green Version]
- Sacco, E.; Bientinesi, R.; Tienforti, D.; Racioppi, M.; Gulino, G.; D’Agostino, D.; Vittori, M.; Bassi, P. Discovery History and Clinical Development of Mirabegron for the Treatment of Overactive Bladder and Urinary Incontinence. Expert Opin. Drug Discov. 2014, 9, 433–448. [Google Scholar] [CrossRef]
- Himms-Hagen, J.; Melnyk, A.; Zingaretti, M.C.; Ceresi, E.; Barbatelli, G.; Cinti, S. Multilocular Fat Cells in WAT of CL-316243-Treated Rats Derive Directly from White Adipocytes. Am. J. Physiol. Cell Physiol. 2000, 279, C670–C681. [Google Scholar] [CrossRef]
- Weyer, C.; Tataranni, P.A.; Snitker, S.; Danforth, E.; Ravussin, E. Increase in Insulin Action and Fat Oxidation after Treatment with CL 316,243, a Highly Selective Beta3-Adrenoceptor Agonist in Humans. Diabetes 1998, 47, 1555–1561. [Google Scholar] [CrossRef]
- van Baak, M.A.; Hul, G.B.J.; Toubro, S.; Astrup, A.; Gottesdiener, K.M.; DeSmet, M.; Saris, W.H.M. Acute Effect of L-796568, a Novel Beta 3-Adrenergic Receptor Agonist, on Energy Expenditure in Obese Men. Clin. Pharmacol. Ther. 2002, 71, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Larsen, T.M.; Toubro, S.; van Baak, M.A.; Gottesdiener, K.M.; Larson, P.; Saris, W.H.M.; Astrup, A. Effect of a 28-d Treatment with L-796568, a Novel Beta(3)-Adrenergic Receptor Agonist, on Energy Expenditure and Body Composition in Obese Men. Am. J. Clin. Nutr. 2002, 76, 780–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buemann, B.; Toubro, S.; Astrup, A. Effects of the Two β3-Agonists, ZD7114 and ZD2079 on 24 Hour Energy Expenditure and Respiratory Quotient in Obese Subjects. Int. J. Obes. 2000, 24, 1553–1560. [Google Scholar] [CrossRef] [Green Version]
- Lockie, S.H.; Heppner, K.M.; Chaudhary, N.; Chabenne, J.R.; Morgan, D.A.; Veyrat-Durebex, C.; Ananthakrishnan, G.; Rohner-Jeanrenaud, F.; Drucker, D.J.; DiMarchi, R.; et al. Direct Control of Brown Adipose Tissue Thermogenesis by Central Nervous System Glucagon-Like Peptide-1 Receptor Signaling. Diabetes 2012, 61, 2753–2762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kooijman, S.; Wang, Y.; Parlevliet, E.T.; Boon, M.R.; Edelschaap, D.; Snaterse, G.; Pijl, H.; Romijn, J.A.; Rensen, P.C.N. Central GLP-1 Receptor Signalling Accelerates Plasma Clearance of Triacylglycerol and Glucose by Activating Brown Adipose Tissue in Mice. Diabetologia 2015, 58, 2637–2646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beiroa, D.; Imbernon, M.; Gallego, R.; Senra, A.; Herranz, D.; Villarroya, F.; Serrano, M.; Fernø, J.; Salvador, J.; Escalada, J.; et al. GLP-1 Agonism Stimulates Brown Adipose Tissue Thermogenesis and Browning through Hypothalamic AMPK. Diabetes 2014, 63, 3346–3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Lin, B.; Zheng, X.; Chen, Z.; Cao, H.; Xu, H.; Liang, H.; Weng, J. GLP-1 Receptor Agonist Promotes Brown Remodelling in Mouse White Adipose Tissue through SIRT1. Diabetologia 2016, 59, 1059–1069. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, X.; Yin, H.; Zhang, L.; Feng, A.; Zhang, Q.-X.; Lin, Y.; Bao, B.; Hernandez, L.L.; Shi, G.-P.; et al. Functional Inactivation of Mast Cells Enhances Subcutaneous Adipose Tissue Browning in Mice. Cell Rep. 2019, 28, 792–803.e4. [Google Scholar] [CrossRef]
- Frontini, A.; Vitali, A.; Perugini, J.; Murano, I.; Romiti, C.; Ricquier, D.; Guerrieri, M.; Cinti, S. White-to-Brown Transdifferentiation of Omental Adipocytes in Patients Affected by Pheochromocytoma. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2013, 1831, 950–959. [Google Scholar] [CrossRef] [Green Version]
- Gesta, S.; Tseng, Y.-H.; Kahn, C.R. Developmental Origin of Fat: Tracking Obesity to Its Source. Cell 2007, 131, 242–256. [Google Scholar] [CrossRef] [Green Version]
- Rodeheffer, M.S.; Birsoy, K.; Friedman, J.M. Identification of White Adipocyte Progenitor Cells In Vivo. Cell 2008, 135, 240–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ussar, S.; Lee, K.Y.; Dankel, S.N.; Boucher, J.; Haering, M.-F.; Kleinridders, A.; Thomou, T.; Xue, R.; Macotela, Y.; Cypess, A.M.; et al. ASC-1, PAT2, and P2RX5 Are Cell Surface Markers for White, Beige, and Brown Adipocytes. Sci. Transl. Med. 2014, 6, 247ra103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, L.Z.; Shinoda, K.; Ohno, H.; Scheel, D.W.; Tomoda, E.; Ruiz, L.; Hu, H.; Wang, L.; Pavlova, Z.; Gilsanz, V.; et al. Human BAT Possesses Molecular Signatures That Resemble Beige/Brite Cells. PLoS ONE 2012, 7, e49452. [Google Scholar] [CrossRef]
- Garcia, R.A.; Roemmich, J.N.; Claycombe, K.J. Evaluation of Markers of Beige Adipocytes in White Adipose Tissue of the Mouse. Nutr. Metab. 2016, 13, 24. [Google Scholar] [CrossRef] [Green Version]
- Waldén, T.B.; Hansen, I.R.; Timmons, J.A.; Cannon, B.; Nedergaard, J. Recruited vs. Nonrecruited Molecular Signatures of Brown, “Brite,” and White Adipose Tissues. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E19–E31. [Google Scholar] [CrossRef] [Green Version]
- Cypess, A.M.; White, A.P.; Vernochet, C.; Schulz, T.J.; Xue, R.; Sass, C.A.; Huang, T.L.; Roberts-Toler, C.; Weiner, L.S.; Sze, C.; et al. Anatomical Localization, Gene Expression Profiling and Functional Characterization of Adult Human Neck Brown Fat. Nat. Med. 2013, 19, 635–639. [Google Scholar] [CrossRef] [PubMed]
- Comas, F.; Martínez, C.; Sabater, M.; Ortega, F.; Latorre, J.; Díaz-Sáez, F.; Aragonés, J.; Camps, M.; Gumà, A.; Ricart, W.; et al. Neuregulin 4 Is a Novel Marker of Beige Adipocyte Precursor Cells in Human Adipose Tissue. Front. Physiol. 2019, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-H.; Petkova, A.P.; Mottillo, E.P.; Granneman, J.G. In Vivo Identification of Bipotential Adipocyte Progenitors Recruited by Β3-Adrenoceptor Activation and High-Fat Feeding. Cell Metab. 2012, 15, 480–491. [Google Scholar] [CrossRef] [Green Version]
- Pilkington, A.-C.; Paz, H.A.; Wankhade, U.D. Beige Adipose Tissue Identification and Marker Specificity-Overview. Front. Endocrinol. 2021, 12, 599134. [Google Scholar] [CrossRef]
- de Jong, J.M.A.; Larsson, O.; Cannon, B.; Nedergaard, J. A Stringent Validation of Mouse Adipose Tissue Identity Markers. Am. J. Physiol.-Endocrinol. Metab. 2015, 308, E1085–E1105. [Google Scholar] [CrossRef] [Green Version]
- Kajimura, S.; Spiegelman, B.M.; Seale, P. Brown and Beige Fat: Physiological Roles beyond Heat Generation. Cell Metab. 2015, 22, 546–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wankhade, U.D.; Lee, J.-H.; Dagur, P.K.; Yadav, H.; Shen, M.; Chen, W.; Kulkarni, A.B.; McCoy, J.P.; Finkel, T.; Cypess, A.M.; et al. TGF-β Receptor 1 Regulates Progenitors That Promote Browning of White Fat. Mol. Metab. 2018, 16, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Kajimura, S.; Yang, W.; Chin, S.; Rohas, L.; Uldry, M.; Tavernier, G.; Langin, D.; Spiegelman, B.M. Transcriptional Control of Brown Fat Determination by PRDM16. Cell Metab. 2007, 6, 38–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puigserver, P.; Wu, Z.; Park, C.W.; Graves, R.; Wright, M.; Spiegelman, B.M. A Cold-Inducible Coactivator of Nuclear Receptors Linked to Adaptive Thermogenesis. Cell 1998, 92, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Uldry, M.; Yang, W.; St-Pierre, J.; Lin, J.; Seale, P.; Spiegelman, B.M. Complementary Action of the PGC-1 Coactivators in Mitochondrial Biogenesis and Brown Fat Differentiation. Cell Metab. 2006, 3, 333–341. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, T.; Sakai, J.; Kajimura, S. Transcriptional and Epigenetic Control of Brown and Beige Adipose Cell Fate and Function. Nat. Rev. Mol. Cell Biol. 2016, 17, 480–495. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Zhong, L.; Lee, J.T.H.; Zhang, J.; Wu, D.; Geng, L.; Wang, Y.; Wong, C.-M.; Xu, A. The FGF21-CCL11 Axis Mediates Beiging of White Adipose Tissues by Coupling Sympathetic Nervous System to Type 2 Immunity. Cell Metab. 2017, 26, 493–508.e4. [Google Scholar] [CrossRef] [Green Version]
- Harms, M.; Seale, P. Brown and Beige Fat: Development, Function and Therapeutic Potential. Nat. Med. 2013, 19, 1252–1263. [Google Scholar] [CrossRef] [Green Version]
- van Marken Lichtenbelt, W.D.; Vanhommerig, J.W.; Smulders, N.M.; Drossaerts, J.M.A.F.L.; Kemerink, G.J.; Bouvy, N.D.; Schrauwen, P.; Teule, G.J.J. Cold-Activated Brown Adipose Tissue in Healthy Men. N. Engl. J. Med. 2009, 360, 1500–1508. [Google Scholar] [CrossRef] [Green Version]
- Huttunen, P.; Hirvonen, J.; Kinnula, V. The Occurrence of Brown Adipose Tissue in Outdoor Workers. Eur. J. Appl. Physiol. Occup. Physiol. 1981, 46, 339–345. [Google Scholar] [CrossRef]
- Saito, M.; Okamatsu-Ogura, Y.; Matsushita, M.; Watanabe, K.; Yoneshiro, T.; Nio-Kobayashi, J.; Iwanaga, T.; Miyagawa, M.; Kameya, T.; Nakada, K.; et al. High Incidence of Metabolically Active Brown Adipose Tissue in Healthy Adult Humans: Effects of Cold Exposure and Adiposity. Diabetes 2009, 58, 1526–1531. [Google Scholar] [CrossRef] [Green Version]
- Sidossis, L.; Kajimura, S. Brown and Beige Fat in Humans: Thermogenic Adipocytes That Control Energy and Glucose Homeostasis. J. Clin. Invest. 2015, 125, 478–486. [Google Scholar] [CrossRef]
- Ghorbani, M.; Claus, T.H.; Himms-Hagen, J. Hypertrophy of Brown Adipocytes in Brown and White Adipose Tissues and Reversal of Diet-Induced Obesity in Rats Treated with a Beta3-Adrenoceptor Agonist. Biochem. Pharmacol. 1997, 54, 121–131. [Google Scholar] [CrossRef]
- Ghorbani, M.; Himms-Hagen, J. Appearance of Brown Adipocytes in White Adipose Tissue during CL 316,243-Induced Reversal of Obesity and Diabetes in Zucker Fa/Fa Rats. Int. J. Obes. Relat. Metab. Disord. 1997, 21, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Cousin, B.; Cinti, S.; Morroni, M.; Raimbault, S.; Ricquier, D.; Pénicaud, L.; Casteilla, L. Occurrence of Brown Adipocytes in Rat White Adipose Tissue: Molecular and Morphological Characterization. J. Cell Sci. 1992, 103 Pt 4, 931–942. [Google Scholar] [CrossRef]
- Jimenez, M.; Barbatelli, G.; Allevi, R.; Cinti, S.; Seydoux, J.; Giacobino, J.-P.; Muzzin, P.; Preitner, F. Beta 3-Adrenoceptor Knockout in C57BL/6J Mice Depresses the Occurrence of Brown Adipocytes in White Fat. Eur. J. Biochem. 2003, 270, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Roh, H.C.; Tsai, L.T.Y.; Shao, M.; Tenen, D.; Shen, Y.; Kumari, M.; Lyubetskaya, A.; Jacobs, C.; Dawes, B.; Gupta, R.K.; et al. Warming Induces Significant Reprogramming of Beige, but Not Brown, Adipocyte Cellular Identity. Cell Metab. 2018, 27, 1121–1137.e5. [Google Scholar] [CrossRef]
- Hardie, D.G.; Carling, D.; Carlson, M. The AMP-Activated/SNF1 Protein Kinase Subfamily: Metabolic Sensors of the Eukaryotic Cell? Annu. Rev. Biochem. 1998, 67, 821–855. [Google Scholar] [CrossRef] [PubMed]
- Rutter, G.A.; Da Silva Xavier, G.; Leclerc, I. Roles of 5’-AMP-Activated Protein Kinase (AMPK) in Mammalian Glucose Homoeostasis. Biochem. J. 2003, 375, 1–16. [Google Scholar] [CrossRef]
- López, M.; Nogueiras, R.; Tena-Sempere, M.; Diéguez, C. Hypothalamic AMPK: A Canonical Regulator of Whole-Body Energy Balance. Nat. Rev. Endocrinol. 2016, 12, 421–432. [Google Scholar] [CrossRef] [PubMed]
- López, M.; Varela, L.; Vázquez, M.J.; Rodríguez-Cuenca, S.; González, C.R.; Velagapudi, V.R.; Morgan, D.A.; Schoenmakers, E.; Agassandian, K.; Lage, R.; et al. Hypothalamic AMPK and Fatty Acid Metabolism Mediate Thyroid Regulation of Energy Balance. Nat. Med. 2010, 16, 1001–1008. [Google Scholar] [CrossRef]
- Martínez-Sánchez, N.; Moreno-Navarrete, J.M.; Contreras, C.; Rial-Pensado, E.; Fernø, J.; Nogueiras, R.; Diéguez, C.; Fernández-Real, J.-M.; López, M. Thyroid Hormones Induce Browning of White Fat. J. Endocrinol. 2016, 232, 351–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.; He, A.; Lodhi, I.J. Lipid Regulators of Thermogenic Fat Activation. Trends Endocrinol. Metab. 2019, 30, 710–723. [Google Scholar] [CrossRef] [PubMed]
- Guilherme, A.; Pedersen, D.J.; Henchey, E.; Henriques, F.S.; Danai, L.V.; Shen, Y.; Yenilmez, B.; Jung, D.; Kim, J.K.; Lodhi, I.J.; et al. Adipocyte Lipid Synthesis Coupled to Neuronal Control of Thermogenic Programming. Mol. Metab. 2017, 6, 781–796. [Google Scholar] [CrossRef] [PubMed]
- Lodhi, I.J.; Yin, L.; Jensen-Urstad, A.P.L.; Funai, K.; Coleman, T.; Baird, J.H.; El Ramahi, M.K.; Razani, B.; Song, H.; Fu-Hsu, F.; et al. Inhibiting Adipose Tissue Lipogenesis Reprograms Thermogenesis and PPARγ Activation to Decrease Diet-Induced Obesity. Cell Metab. 2012, 16, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Sadurskis, A.; Dicker, A.; Cannon, B.; Nedergaard, J. Polyunsaturated Fatty Acids Recruit Brown Adipose Tissue: Increased UCP Content and NST Capacity. Am. J. Physiol. 1995, 269, E351–E360. [Google Scholar] [CrossRef]
- Kim, M.; Goto, T.; Yu, R.; Uchida, K.; Tominaga, M.; Kano, Y.; Takahashi, N.; Kawada, T. Fish Oil Intake Induces UCP1 Upregulation in Brown and White Adipose Tissue via the Sympathetic Nervous System. Sci. Rep. 2015, 5, 18013. [Google Scholar] [CrossRef] [Green Version]
- Ghandour, R.A.; Colson, C.; Giroud, M.; Maurer, S.; Rekima, S.; Ailhaud, G.; Klingenspor, M.; Amri, E.-Z.; Pisani, D.F. Impact of Dietary Ω3 Polyunsaturated Fatty Acid Supplementation on Brown and Brite Adipocyte Function. J. Lipid Res. 2018, 59, 452–461. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; He, A.; Tan, M.; Johnson, J.M.; Dean, J.M.; Pietka, T.A.; Chen, Y.; Zhang, X.; Hsu, F.-F.; Razani, B.; et al. Peroxisome-Derived Lipids Regulate Adipose Thermogenesis by Mediating Cold-Induced Mitochondrial Fission. J. Clin. Invest. 2019, 129, 694–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafontan, M.; Moro, C.; Berlan, M.; Crampes, F.; Sengenes, C.; Galitzky, J. Control of Lipolysis by Natriuretic Peptides and Cyclic GMP. Trends Endocrinol. Metab. 2008, 19, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Zois, N.E.; Bartels, E.D.; Hunter, I.; Kousholt, B.S.; Olsen, L.H.; Goetze, J.P. Natriuretic Peptides in Cardiometabolic Regulation and Disease. Nat. Rev. Cardiol. 2014, 11, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Collins, S. A Heart-Adipose Tissue Connection in the Regulation of Energy Metabolism. Nat. Rev. Endocrinol. 2014, 10, 157–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpe, M.; Rubattu, S.; Burnett, J. Natriuretic Peptides in Cardiovascular Diseases: Current Use and Perspectives. Eur. Heart J. 2014, 35, 419–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Ceddia, R.P.; Collins, S. Cardiac Natriuretic Peptides Promote Adipose “browning” through MTOR Complex-1. Mol. Metab. 2018, 9, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Tran, C.M.; Mukherjee, S.; Ye, L.; Frederick, D.W.; Kissig, M.; Davis, J.G.; Lamming, D.W.; Seale, P.; Baur, J.A. Rapamycin Blocks Induction of the Thermogenic Program in White Adipose Tissue. Diabetes 2016, 65, 927–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Bordicchia, M.; Zhang, C.; Fang, H.; Wei, W.; Li, J.-L.; Guilherme, A.; Guntur, K.; Czech, M.P.; Collins, S. Activation of MTORC1 Is Essential for β-Adrenergic Stimulation of Adipose Browning. J. Clin. Invest. 2016, 126, 1704–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, H.; Nagoshi, T.; Oi, Y.; Yoshii, A.; Tanaka, Y.; Takahashi, H.; Kashiwagi, Y.; Tanaka, T.D.; Yoshimura, M. Treatment with Atrial Natriuretic Peptide Induces Adipose Tissue Browning and Exerts Thermogenic Actions In Vivo. Sci. Rep. 2021, 11, 17466. [Google Scholar] [CrossRef] [PubMed]
- Bordicchia, M.; Liu, D.; Amri, E.-Z.; Ailhaud, G.; Dessì-Fulgheri, P.; Zhang, C.; Takahashi, N.; Sarzani, R.; Collins, S. Cardiac Natriuretic Peptides Act via P38 MAPK to Induce the Brown Fat Thermogenic Program in Mouse and Human Adipocytes. J. Clin. Invest. 2012, 122, 1022–1036. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Xu, A. Adipose Extracellular Vesicles in Intercellular and Inter-Organ Crosstalk in Metabolic Health and Diseases. Front. Immunol. 2021, 12, 608680. [Google Scholar] [CrossRef] [PubMed]
- van der Pol, E.; Böing, A.N.; Harrison, P.; Sturk, A.; Nieuwland, R. Classification, Functions, and Clinical Relevance of Extracellular Vesicles. Pharmacol. Rev. 2012, 64, 676–705. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Shang, Q.; Pan, Z.; Bai, Y.; Li, Z.; Zhang, H.; Zhang, Q.; Guo, C.; Zhang, L.; Wang, Q. Exosomes from Adipose-Derived Stem Cells Attenuate Adipose Inflammation and Obesity Through Polarizing M2 Macrophages and Beiging in White Adipose Tissue. Diabetes 2018, 67, 235–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.J.; Kim, H.K.; Cho, Y.; Choi, J.S.; Woo, C.H.; Lee, K.S.; Sul, J.H.; Lee, C.M.; Han, J.; Park, J.H.; et al. Cell Reprogramming Using Extracellular Vesicles from Differentiating Stem Cells into White/Beige Adipocytes. Sci. Adv. 2020, 6, eaay6721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-Derived Circulating MiRNAs Regulate Gene Expression in Other Tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Nakagami, H.; Rodriguez-Araujo, G.; Nimura, K.; Kaneda, Y. Essential Role for MiR-196a in Brown Adipogenesis of White Fat Progenitor Cells. PLoS Biol. 2012, 10, e1001314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Siegel, F.; Kipschull, S.; Haas, B.; Fröhlich, H.; Meister, G.; Pfeifer, A. MiR-155 Regulates Differentiation of Brown and Beige Adipocytes via a Bistable Circuit. Nat. Commun. 2013, 4, 1769. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.-L.; Li, Z.-Y.; Song, J.; Liu, J.-M.; Miao, C.-Y. Metrnl: A Secreted Protein with New Emerging Functions. Acta Pharmacol. Sin. 2016, 37, 571–579. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.R.; Long, J.Z.; White, J.P.; Svensson, K.J.; Lou, J.; Lokurkar, I.; Jedrychowski, M.P.; Ruas, J.L.; Wrann, C.D.; Lo, J.C.; et al. Meteorin-like Is a Hormone That Regulates Immune-Adipose Interactions to Increase Beige Fat Thermogenesis. Cell 2014, 157, 1279–1291. [Google Scholar] [CrossRef] [Green Version]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-Dependent Myokine That Drives Brown-Fat-like Development of White Fat and Thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Raschke, S.; Elsen, M.; Gassenhuber, H.; Sommerfeld, M.; Schwahn, U.; Brockmann, B.; Jung, R.; Wisløff, U.; Tjønna, A.E.; Raastad, T.; et al. Evidence against a Beneficial Effect of Irisin in Humans. PLoS ONE 2013, 8, e73680. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-W.; Odegaard, J.I.; Mukundan, L.; Qiu, Y.; Molofsky, A.B.; Nussbaum, J.C.; Yun, K.; Locksley, R.M.; Chawla, A. Activated Type 2 Innate Lymphoid Cells Regulate Beige Fat Biogenesis. Cell 2015, 160, 74–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Molofsky, A.B.; Liang, H.-E.; Ricardo-Gonzalez, R.R.; Jouihan, H.A.; Bando, J.K.; Chawla, A.; Locksley, R.M. Eosinophils Sustain Adipose Alternatively Activated Macrophages Associated with Glucose Homeostasis. Science 2011, 332, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Nguyen, K.D.; Odegaard, J.I.; Cui, X.; Tian, X.; Locksley, R.M.; Palmiter, R.D.; Chawla, A. Eosinophils and Type 2 Cytokine Signaling in Macrophages Orchestrate Development of Functional Beige Fat. Cell 2014, 157, 1292–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, K.D.; Qiu, Y.; Cui, X.; Goh, Y.P.S.; Mwangi, J.; David, T.; Mukundan, L.; Brombacher, F.; Locksley, R.M.; Chawla, A. Alternatively Activated Macrophages Produce Catecholamines to Sustain Adaptive Thermogenesis. Nature 2011, 480, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Herz, C.T.; Kiefer, F.W. Adipose Tissue Browning in Mice and Humans. J. Endocrinol. 2019, 241, R97–R109. [Google Scholar] [CrossRef]
- Chechi, K.; Blanchard, P.-G.; Mathieu, P.; Deshaies, Y.; Richard, D. Brown Fat like Gene Expression in the Epicardial Fat Depot Correlates with Circulating HDL-Cholesterol and Triglycerides in Patients with Coronary Artery Disease. Int. J. Cardiol. 2013, 167, 2264–2270. [Google Scholar] [CrossRef]
- Barchuk, M.; Dutour, A.; Ancel, P.; Svilar, L.; Miksztowicz, V.; Lopez, G.; Rubio, M.; Schreier, L.; Nogueira, J.P.; Valéro, R.; et al. Untargeted Lipidomics Reveals a Specific Enrichment in Plasmalogens in Epicardial Adipose Tissue and a Specific Signature in Coronary Artery Disease. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 986–1000. [Google Scholar] [CrossRef]
- Ahmadi, N.; Nabavi, V.; Hajsadeghi, F.; Zeb, I.; Flores, F.; Ebrahimi, R.; Budoff, M. Aged Garlic Extract with Supplement Is Associated with Increase in Brown Adipose, Decrease in White Adipose Tissue and Predict Lack of Progression in Coronary Atherosclerosis. Int. J. Cardiol. 2013, 168, 2310–2314. [Google Scholar] [CrossRef] [PubMed]
- Dozio, E.; Vianello, E.; Malavazos, A.E.; Tacchini, L.; Schmitz, G.; Iacobellis, G.; Corsi Romanelli, M.M. Epicardial Adipose Tissue GLP-1 Receptor Is Associated with Genes Involved in Fatty Acid Oxidation and White-to-Brown Fat Differentiation: A Target to Modulate Cardiovascular Risk? Int. J. Cardiol. 2019, 292, 218–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldassano, S.; Amato, A.; Rappa, F.; Cappello, F.; Mulè, F. Influence of Endogenous Glucagon-like Peptide-2 on Lipid Disorders in Mice Fed a High-Fat Diet. Endocr. Res. 2016, 41, 317–324. [Google Scholar] [CrossRef]
- Shafiei-Jahani, P.; Hurrell, B.P.; Galle-Treger, L.; Helou, D.G.; Howard, E.; Painter, J.; Lo, R.; Lewis, G.; Soroosh, P.; Akbari, O. DR3 Stimulation of Adipose Resident ILC2s Ameliorates Type 2 Diabetes Mellitus. Nat. Commun. 2020, 11, 4718. [Google Scholar] [CrossRef]
- Distel, E.; Penot, G.; Cadoudal, T.; Balguy, I.; Durant, S.; Benelli, C. Early Induction of a Brown-like Phenotype by Rosiglitazone in the Epicardial Adipose Tissue of Fatty Zucker Rats. Biochimie 2012, 94, 1660–1667. [Google Scholar] [CrossRef] [PubMed]
- Crewe, C.; Funcke, J.-B.; Li, S.; Joffin, N.; Gliniak, C.M.; Ghaben, A.L.; An, Y.A.; Sadek, H.A.; Gordillo, R.; Akgul, Y.; et al. Extracellular Vesicle-Based Interorgan Transport of Mitochondria from Energetically Stressed Adipocytes. Cell Metab. 2021, 33, 1853–1868.e11. [Google Scholar] [CrossRef] [PubMed]
- Loyer, X.; Boulanger, C.M.; Le Lay, S. Adipocyte Extracellular Vesicles: Rescuers of Cardiac Mitochondrial Stress. Trends Endocrinol. Metab. 2022, 33, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Ku, H.-C.; Cheng, C.-F. Browning the Epicardial Adipose Tissues in Cardiovascular Regulation. Biomed. J. Sci. Tech. Res. 2019, 12, 9332–9334. [Google Scholar] [CrossRef] [Green Version]
- Iacobellis, G. Aging Effects on Epicardial Adipose Tissue. Front. Aging 2021, 2, 12. [Google Scholar] [CrossRef]
- Singh, S.P.; McClung, J.A.; Thompson, E.; Glick, Y.; Greenberg, M.; Acosta-Baez, G.; Edris, B.; Shapiro, J.I.; Abraham, N.G. Cardioprotective Heme Oxygenase-1-PGC1α Signaling in Epicardial Fat Attenuates Cardiovascular Risk in Humans as in Obese Mice. Obesity 2019, 27, 1634–1643. [Google Scholar] [CrossRef]
- Oikonomou, E.K.; Siddique, M.; Antoniades, C. Artificial Intelligence in Medical Imaging: A Radiomic Guide to Precision Phenotyping of Cardiovascular Disease. Cardiovasc. Res. 2020, 116, 2040–2054. [Google Scholar] [CrossRef] [Green Version]
- Commandeur, F.; Goeller, M.; Betancur, J.; Cadet, S.; Doris, M.; Chen, X.; Berman, D.S.; Slomka, P.J.; Tamarappoo, B.K.; Dey, D. Deep Learning for Quantification of Epicardial and Thoracic Adipose Tissue from Non-Contrast CT. IEEE Trans Med. Imaging 2018, 37, 1835–1846. [Google Scholar] [CrossRef]
- Antonopoulos, A.S.; Sanna, F.; Sabharwal, N.; Thomas, S.; Oikonomou, E.K.; Herdman, L.; Margaritis, M.; Shirodaria, C.; Kampoli, A.-M.; Akoumianakis, I.; et al. Detecting Human Coronary Inflammation by Imaging Perivascular Fat. Sci. Transl. Med. 2017, 9, eaal2658. [Google Scholar] [CrossRef] [Green Version]
- Oikonomou, E.K.; Marwan, M.; Desai, M.Y.; Mancio, J.; Alashi, A.; Hutt Centeno, E.; Thomas, S.; Herdman, L.; Kotanidis, C.P.; Thomas, K.E.; et al. Non-Invasive Detection of Coronary Inflammation Using Computed Tomography and Prediction of Residual Cardiovascular Risk (the CRISP CT Study): A Post-Hoc Analysis of Prospective Outcome Data. Lancet 2018, 392, 929–939. [Google Scholar] [CrossRef] [Green Version]
- Oikonomou, E.K.; Desai, M.Y.; Marwan, M.; Kotanidis, C.P.; Antonopoulos, A.S.; Schottlander, D.; Channon, K.M.; Neubauer, S.; Achenbach, S.; Antoniades, C. Perivascular Fat Attenuation Index Stratifies Cardiac Risk Associated with High-Risk Plaques in the CRISP-CT Study. J. Am. Coll. Cardiol. 2020, 76, 755–757. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ding, J.; Zhao, Y.; Ju, S.; Mao, H.; Peng, X.-G. Irisin Induces White Adipose Tissue Browning in Mice as Assessed by Magnetic Resonance Imaging. Exp. Biol. Med. 2021, 246, 1597–1606. [Google Scholar] [CrossRef]
- Hartimath, S.V.; Khanapur, S.; Boominathan, R.; Jiang, L.; Cheng, P.; Yong, F.F.; Tan, P.W.; Robins, E.G.; Goggi, J.L. Imaging Adipose Tissue Browning Using the TSPO-18kDa Tracer [18F]FEPPA. Mol. Metab. 2019, 25, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.Y.; Cypess, A.M.; Laughlin, M.R.; Haft, C.R.; Hu, H.H.; Bredella, M.A.; Enerbäck, S.; Kinahan, P.E.; van Marken Lichtenbelt, W.; Lin, F.I.; et al. Brown Adipose Reporting Criteria in Imaging STudies (BARCIST 1.0): Recommendations for Standardized FDG-PET/CT Experiments in Humans. Cell Metab. 2016, 24, 210–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Zhang, H.; Parhat, K.; Xu, H.; Li, M.; Wang, X.; Ran, C. Molecular Imaging of Brown Adipose Tissue Mass. Int. J. Mol. Sci. 2021, 22, 9436. [Google Scholar] [CrossRef]
- Chondronikola, M.; Beeman, S.C.; Wahl, R.L. Non-Invasive Methods for the Assessment of Brown Adipose Tissue in Humans: BAT Assessment Methods. J. Physiol. 2018, 596, 363–378. [Google Scholar] [CrossRef]
- van der Lans, A.A.J.J.; Wierts, R.; Vosselman, M.J.; Schrauwen, P.; Brans, B.; van Marken Lichtenbelt, W.D. Cold-Activated Brown Adipose Tissue in Human Adults: Methodological Issues. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2014, 307, R103–R113. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Immune Cells | EAT | VAT | ||||
|---|---|---|---|---|---|---|
| EAT | References | VAT | References | |||
| Innate immune cells | Macrophages | M1 | ↑ | Hirata et al., 2011; Vianello et al., 2016; Gurses et al., 2017 | ↑ | Wisnewsky et al., 2009; Aron-Wisnewsky et al., 2009; Morris et al., 2011 |
| M2 | ↓ | Hirata et al., 2011; Vianello et al., 2016; Gurses et al., 2017 | ↓ | Wisnewsky et al., 2009; Aron-Wisnewsky et al., 2009; Morris et al., 2011 | ||
| Eosinophils | N/A | - | ↑ | Wu D et al., 2011; Molofsky et al., 2013 | ||
| Mast cells | Presence | Laine et al. 1999; Mazurek et al., 2003 | ↑ | Divoux et al., 2012; Shi and Shi 2012 | ||
| Neutrophils | N/A | - | ↑ | Carmon et al., 2008; Talukdar et al., 2012 | ||
| Natural killers | ↓ | Mráz et al., 2019 | ↑↑ | O’Rourke et al., 2013; Trim et al., 2018; Lee et al., 2016; Wensveen et al., 2015 | ||
| Dendritic cells | ↑ | Mráz et al., 2019; Horcksman et al., 2017 | ↑↑ | Berthola et al., 2012; Bapat et al., 2015 | ||
| Innate lymphoid Cells (ILCs) | ILC1 | N/A | - | ↑ | Everaere et al., 2017 | |
| ILC2 | N/A | - | ↓ | Everaere et al., 2017 | ||
| ILC3 | N/A | - | N/A | - | ||
| Adaptative immune cells | T Lymphocytes | LTreg | N/A | - | ↑ | Feuerer et al., 2009; Bapat et al., 2015 |
| CD4+ | ND | Hirata et al., 2011 | ↑/↓ | Nishimura et al., 2009; Lee et al., 2016 | ||
| CD8+ | ↑ | Hirata et al., 2011 | ↑↑ | Duffaut et al., 2009; Bapat et al., 2015 | ||
| B Lymphocytes | ↑ | Mráz et al., 2019 | ↑ | Bapat et al., 2015 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doukbi, E.; Soghomonian, A.; Sengenès, C.; Ahmed, S.; Ancel, P.; Dutour, A.; Gaborit, B. Browning Epicardial Adipose Tissue: Friend or Foe? Cells 2022, 11, 991. https://doi.org/10.3390/cells11060991
Doukbi E, Soghomonian A, Sengenès C, Ahmed S, Ancel P, Dutour A, Gaborit B. Browning Epicardial Adipose Tissue: Friend or Foe? Cells. 2022; 11(6):991. https://doi.org/10.3390/cells11060991
Chicago/Turabian StyleDoukbi, Elisa, Astrid Soghomonian, Coralie Sengenès, Shaista Ahmed, Patricia Ancel, Anne Dutour, and Bénédicte Gaborit. 2022. "Browning Epicardial Adipose Tissue: Friend or Foe?" Cells 11, no. 6: 991. https://doi.org/10.3390/cells11060991
APA StyleDoukbi, E., Soghomonian, A., Sengenès, C., Ahmed, S., Ancel, P., Dutour, A., & Gaborit, B. (2022). Browning Epicardial Adipose Tissue: Friend or Foe? Cells, 11(6), 991. https://doi.org/10.3390/cells11060991