
Mechanisms of Action of Extracorporeal Photopheresis in the Control of Bronchiolitis Obliterans Syndrome (BOS): Involvement of Circulating miRNAs

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. ECP Procedures
2.3. miRNAs Selection
2.4. miRNAs Analysis
2.5. miRNAs Target Prediction and Pathway Analysis
2.6. miRNAs and Targets Expression Analyses in PBMCs
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Statistical Analysis
3. Results
3.1. Demographic and Clinical Features of Patients
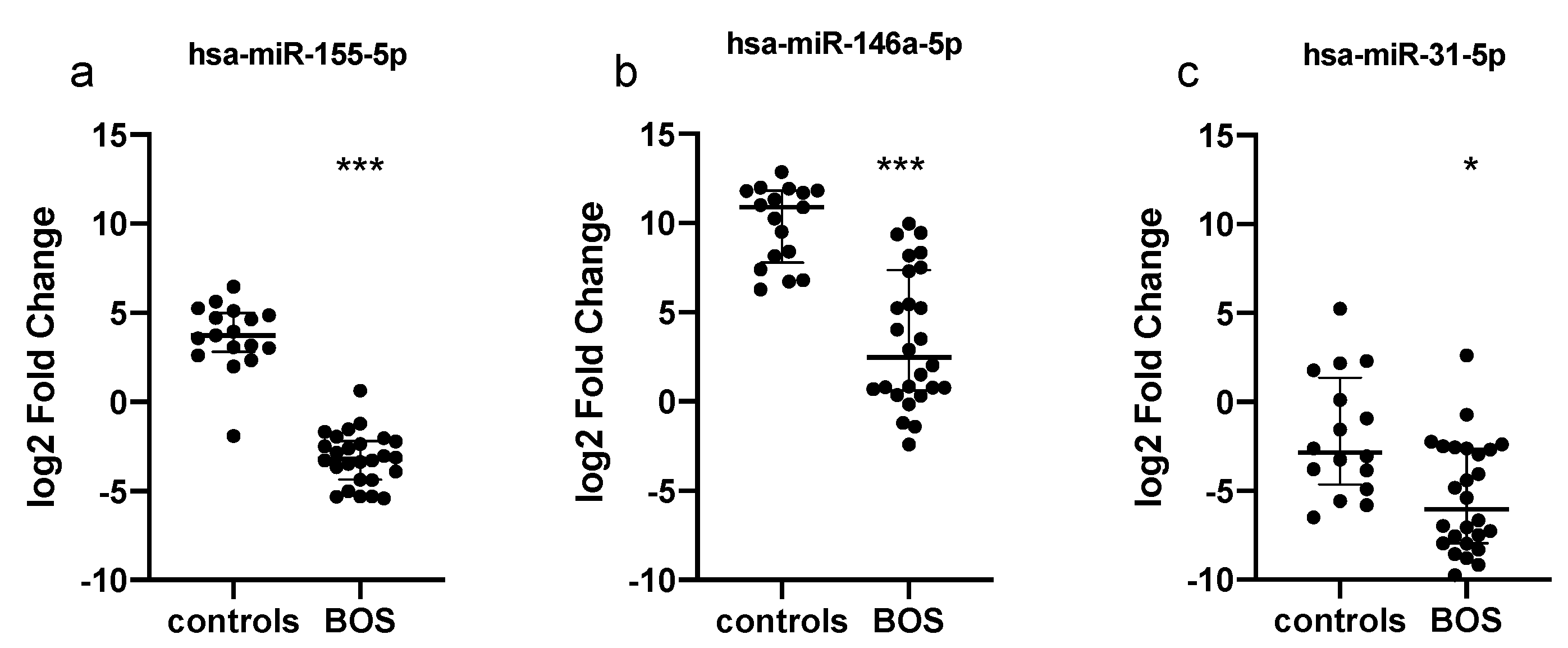
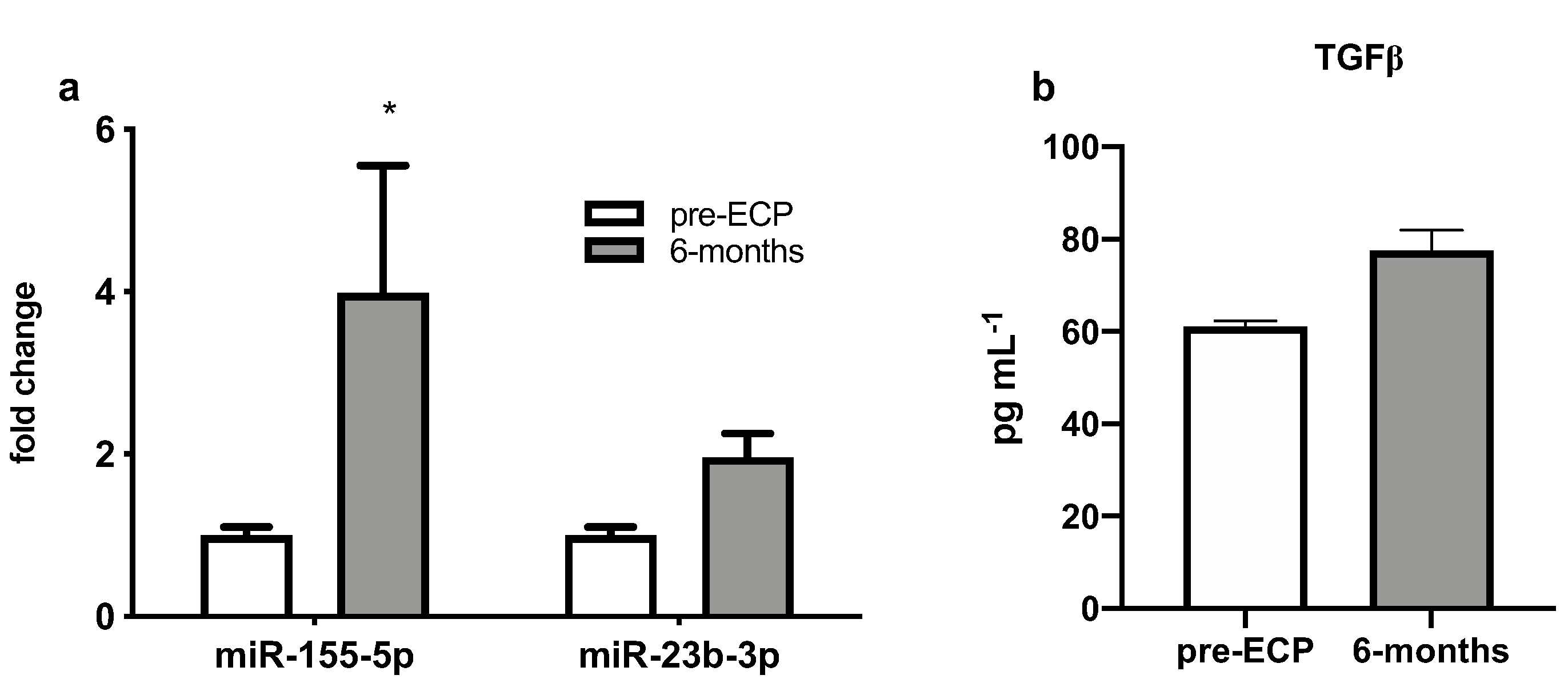
3.2. Serum Expression Levels of Selected miRNAs
3.3. Signaling Pathway Prediction and Targets Analyses
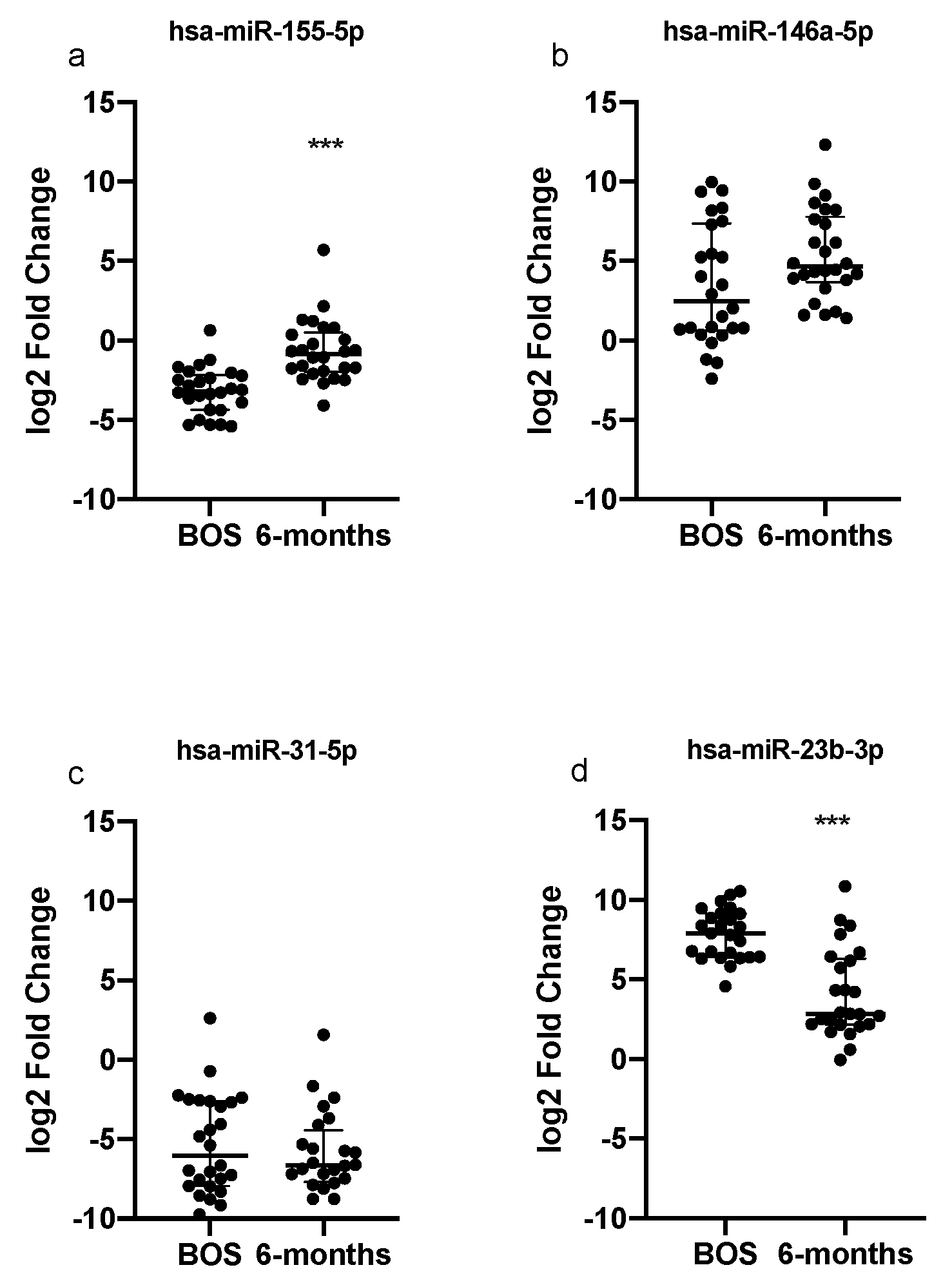
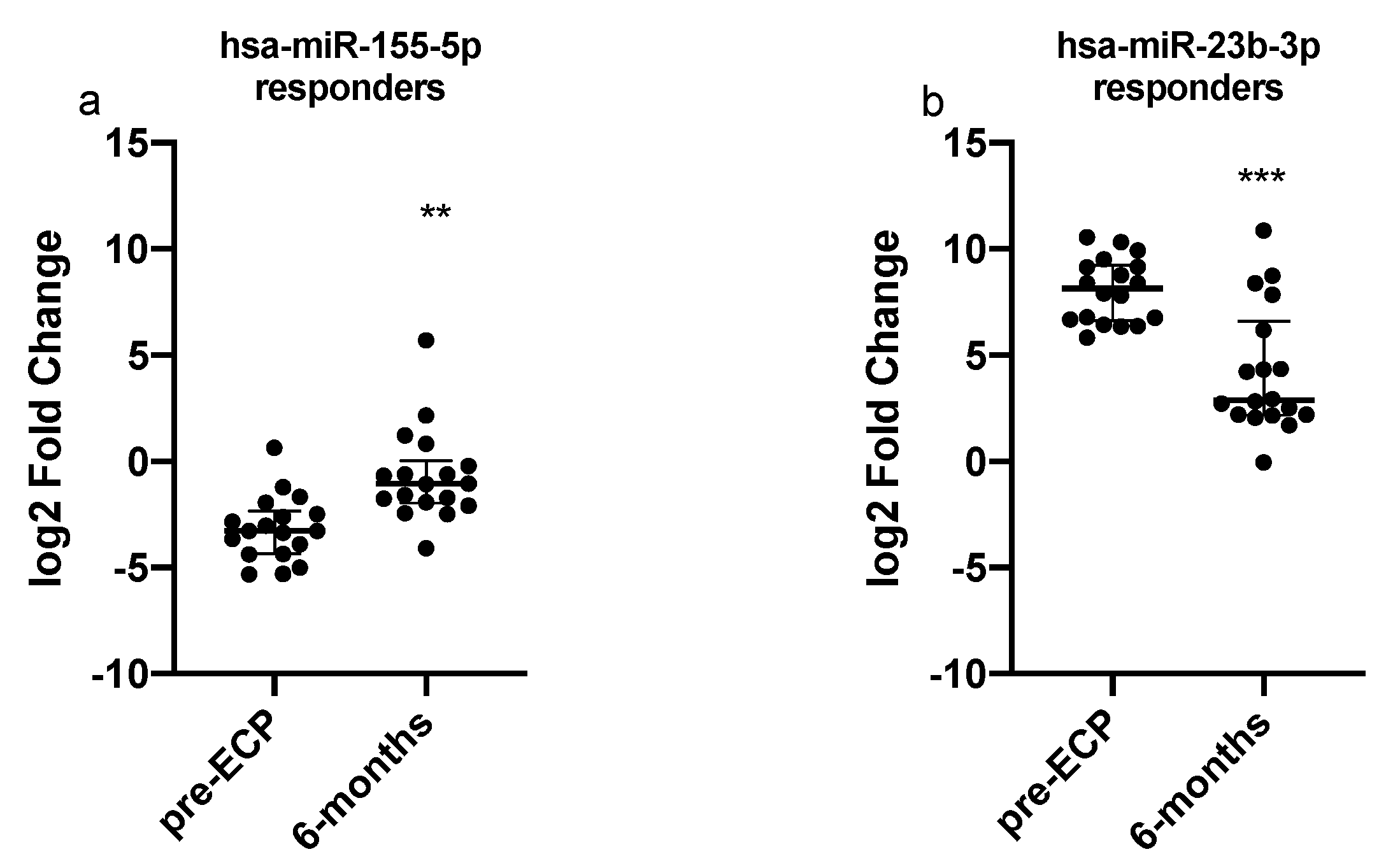
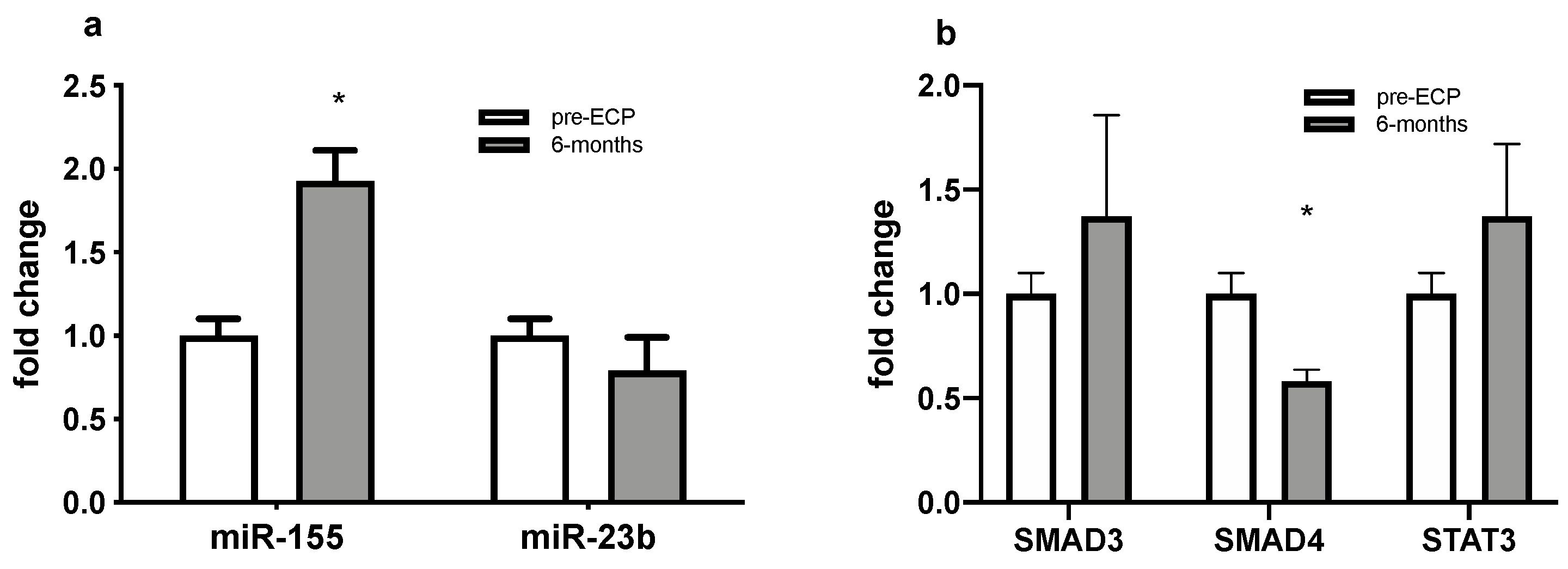
3.4. Dysregulated miRNAs and Selected Targets Expression Analyses in PBMCs
3.5. Expression Levels of Dysregulated miRNAs and TGF-b Levels in BAL Supernatant
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gottlieb, J. Lung Transplantation for Interstitial Lung Diseases and Pulmonary Hypertension. Semin. Respir. Crit. Care Med. 2013, 34, 281–287. [Google Scholar] [CrossRef]
- Yusen, R.D.; Edwards, L.B.; Dipchand, A.I.; Goldfarb, S.B.; Kucheryavaya, A.Y.; Levvey, B.J.; Lund, L.H.; Meiser, B.; Rossano, J.W.; Stehlik, J. International Society for Heart and Lung Transplantation. The Registry of the International Society for Heart and Lung Transplantation: Thirty-third Adult Lung and Heart–Lung Transplant Report—2016; Focus Theme: Primary Diagnostic Indications for Transplant. J. Heart Lung Transplant. 2016, 35, 1170–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barr, M.L. Call It BOS, Call It CLAD—The Need for Prospective Clinical Trials and Elucidating the Mechanism of Extracorporeal Photopheresis. Am. J. Transplant. 2013, 13, 833–834. [Google Scholar] [CrossRef] [PubMed]
- Dieterlen, M.T.; Bittner, H.B.; Pierzchalski, A.; Dhein, S.; Mohr, F.W.; Barten, M.J. Immunological monitoring of extracorporeal photopheresis after heart transplantation. Clin. Exp. Immunol. 2014, 176, 120–128. [Google Scholar] [CrossRef]
- Lorenz, K.; Rommel, K.; Mani, J.; Jin, N.; Hilgendorf, I.; Ho, A.D.; Freund, M.; Schmitt, M.; Schmitt, A. Modulation of lymphocyte subpopulations by extracorporeal photopheresis in pa-tients with acute graft-versus-host disease or graft rejection. Leuk. Lymphoma 2015, 56, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Baskaran, G.; Tiriveedhi, V.; Ramachandran, S.; Aloush, A.; Grossman, B.; Hachem, R.; Mohanakumar, T. Efficacy of extracorporeal photopheresis in clearance of antibodies to donor-specific and lung-specific antigens in lung transplant recipients. J. Heart Lung Transplant. 2014, 33, 950–956. [Google Scholar] [CrossRef] [Green Version]
- Meloni, F.; Cascina, A.; Miserere, S.; Perotti, C.; Vitulo, P.; Fietta, A.M. Peripheral CD4+CD25+ TREG cell counts and the response to extracorporeal photopheresis in lung transplant recipients. Transplant. Proc. 2007, 39, 213–217. [Google Scholar] [CrossRef]
- Hart, J.W.; Shiue, L.H.; Shpall, E.J.; Alousi, A.M. Extracorporeal photopheresis in the treatment of graft-versus-host disease: Evidence and opinion. Ther. Adv. Hematol. 2013, 4, 320–334. [Google Scholar] [CrossRef] [Green Version]
- Edelson, R.L. Mechanistic insights into extracorporeal photochemotherapy: Efficient induction of monocyte-to-dendritic cell maturation. Transfus. Apher. Sci. 2014, 50, 322–329. [Google Scholar] [CrossRef] [Green Version]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of Novel Genes Coding for Small Expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef] [Green Version]
- Zampetaki, A.; Mayr, M. MicroRNAs in vascular and metabolic disease. Circ. Res. 2012, 110, 508–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, R.; Ahmed, S.A. MicroRNA, a new paradigm for understanding immunoregulation, inflammation, and autoimmune diseases. Transl. Res. 2011, 157, 163–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, S. MicroRNAs in fibrosis: Opportunities and challenges. Arthritis Res. Ther. 2016, 18, 11. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-Z.; Schaffert, S.; Fragoso, R.; Loh, C. Regulation of immune responses and tolerance: The microRNA perspective. Immunol. Rev. 2013, 253, 112–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, M.B.; Tuncer, H.H. Photopheresis in solid organ transplant rejection. J. Clin. Apher. 2006, 21, 72–77. [Google Scholar] [CrossRef]
- Perotti, C.; Del Fante, C.; Tinelli, C.; Viarengo, G.; Scudeller, L.; Zecca, M.; Locatelli, F.; Salvaneschi, L. Extracorporeal photochemotherapy in graft-versus-host disease: A longitudinal study on factors influencing the response and survival in pediatric patients. Transfusion 2010, 50, 1359–1369. [Google Scholar] [CrossRef]
- Stumpfova, Z.; Hezova, R.; Meli, A.C.; Slaby, O.; Michalek, J. MicroRNA Profiling of Activated and Tolerogenic Human Dendritic Cells. Mediat. Inflamm. 2014, 2014, 259689. [Google Scholar] [CrossRef]
- Vigorito, E.; Perks, K.L.; Abreu-Goodger, C.; Bunting, S.; Xiang, Z.; Kohlhaas, S.; Das, P.P.; Miska, E.; Rodriguez, A.; Bradley, A.; et al. microRNA-155 Regulates the Generation of Immunoglobulin Class-Switched Plasma Cells. Immunity 2007, 27, 847–859. [Google Scholar] [CrossRef] [Green Version]
- Lai, M.; Xiao, C. Functional interactions among members of the miR-17–92 cluster in lymphocyte development, differentiation and malignant transformation. Int. Immunopharmacol. 2015, 28, 854–858. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Li, N.; Liu, G. The Role of MicroRNAs in Regulatory T Cells. J. Immunol. Res. 2020, 2020, 3232061. [Google Scholar] [CrossRef]
- Jebbawi, F.; Fayyad-Kazan, H.; Merimi, M.; Lewalle, P.; Verougstraete, J.C.J.; Leo, O.; Romero, P.; Burny, A.; Badran, B.; Martiat, P.; et al. A microRNA profile of human CD8+ regulatory T cells and characterization of the effects of microRNAs on Treg cell-associated genes. J. Transl. Med. 2014, 12, 218. [Google Scholar] [CrossRef] [PubMed]
- Hippen, K.L.; Loschi, M.; Nicholls, J.; MacDonald, K.P.A.; Blazar, B.R. Effects of MicroRNA on Regulatory T Cells and Implications for Adoptive Cellular Therapy to Ameliorate Graft-versus-Host Disease. Front. Immunol. 2018, 9, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlachos, I.S.; Kostoulas, N.; Vergoulis, T.; Georgakilas, G.; Reczko, M.; Maragkakis, M.; Paraskevopoulou, M.D.; Prionidis, K.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA miRPath v.2.0: Investigating the combinatorial effect of microRNAs in pathways. Nucleic Acids Res. 2012, 40, W498–W504. [Google Scholar] [CrossRef] [PubMed]
- Hossain, D.M.S.; Panda, A.K.; Chakrabarty, S.; Bhattacharjee, P.; Kajal, K.; Mohanty, S.; Sarkar, I.; Sarkar, D.K.; Kar, S.K.; Sa, G. MEK inhibition prevents tumour-shed transforming growth factor-β-induced T-regulatory cell augmentation in tumour milieu. Immunology 2015, 144, 561–573. [Google Scholar] [CrossRef] [Green Version]
- Iizuka-Koga, M.; Nakatsukasa, H.; Ito, M.; Akanuma, T.; Lu, Q.; Yoshimura, A. Induction and maintenance of regulatory T cells by transcription factors and epigenetic modifications. J. Autoimmun. 2017, 83, 113–121. [Google Scholar] [CrossRef]
- Dong, M.; Wang, X.; Li, T.; Wang, J.; Yang, Y.; Liu, Y.; Jing, Y.; Zhao, H.; Chen, J. Mir-27a-3p attenuates bronchiolitis obliterans in vivo via the regulation of dendritic cells’ maturation and the suppression of myofibroblasts’ differentiation. Clin. Transl. Med. 2020, 10, e140. [Google Scholar] [CrossRef]
- Pan, H.; Ding, E.; Hu, M.; Lagoo, A.S.; Datto, M.B.; Lagoo-Deenadayalan, S.A. SMAD4 Is Required for Development of Maximal Endotoxin Tolerance. J. Immunol. 2010, 184, 5502–5509. [Google Scholar] [CrossRef] [Green Version]
- Verleden, G.M.; Glanville, A.R.; Lease, E.D.; Fisher, A.J.; Calabrese, F.; Corris, P.A.; Ensor, C.R.; Gottlieb, J.; Hachem, R.R.; Lama, V.; et al. Chronic lung allograft dysfunction: Definition, diagnostic criteria, and approaches to treatment―A consensus report from the Pulmonary Council of the ISHLT. J. Hear. Lung Transplant. 2019, 38, 493–503. [Google Scholar] [CrossRef] [Green Version]
- Pecoraro, Y.; Carillo, C.; Diso, D.; Mantovani, S.; Cimino, G.; De Giacomo, T.; Troiani, P.; Shafii, M.; Gherzi, L.; Amore, D.; et al. Efficacy of Extracorporeal Photopheresis in Patients with Bronchiolitis Obliterans Syndrome after Lung Transplantation. Transplant. Proc. 2017, 49, 695–698. [Google Scholar] [CrossRef]
- The EPI Study Group; Hage, C.A.; Klesney-Tait, J.; Wille, K.; Arcasoy, S.; Yung, G.; Hertz, M.; Chan, K.M.; Morrell, M.; Goldberg, H.; et al. Extracorporeal photopheresis to attenuate decline in lung function due to refractory obstructive allograft dysfunction. Transfus. Med. 2021, 31, 292–302. [Google Scholar] [CrossRef]
- Morrell, M.R.; Despotis, G.J.; Lublin, D.M.; Patterson, G.A.; Trulock, E.P.; Hachem, R.R. The efficacy of photopheresis for bronchiolitis obliterans syndrome after lung transplantation. J. Hear. Lung Transplant. 2010, 29, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Piccirillo, N.; Putzulu, R.; Massini, G.; Di Giovanni, A.; Giammarco, S.; Metafuni, E.; Sica, S.; Zini, G.; Chiusolo, P. Inline and offline extracorporeal photopheresis: Device performance, cell yields and clinical response. J. Clin. Apher. 2021, 36, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Helmberg, W.; Sipurzynski, S.; Groselje-Strehle, A.; Greinix, H.; Schlenke, P. Does Offline Beat Inline Treatment: Investigation into Extracorporeal Photopheresis. Transfus. Med. Hemotherapy 2020, 47, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Bueno, J.; Alonso, R.; Gonzalez-Santillana, C.; Naya, D.; Romera, I.; Alarcón, A.; Aguilar, M.; Bautista, G.; Duarte, R.; Ussetti, P.; et al. A paired trial comparing mononuclear cell collection in two machines for further inactivation through an inline or offline extracorporeal photopheresis procedure. Transfusion 2019, 59, 340–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, W.; Kontekakis, A.; Maas, C.; Kuchenbecker, U.; Behlke, S.; Schennach, H. Comparison of procedure times and collection efficiencies using integrated and multistep nonintegrated procedures for extracorporeal photopheresis. J. Clin. Apher. 2022. [Google Scholar] [CrossRef]
- Maeda, A.; Schwarz, A.; Kernebeck, K.; Gross, N.; Aragane, Y.; Peritt, D.; Schwarz, T. Intravenous Infusion of Syngeneic Apoptotic Cells by Photopheresis Induces Antigen-Specific Regulatory T Cells. J. Immunol. 2005, 174, 5968–5976. [Google Scholar] [CrossRef] [Green Version]
- Hequet, O.; Nosbaum, A.; Guironnet-Paquet, A.; Blasco, E.; Nicolas-Virelizier, E.; Griffith, T.S.; Rigal, D.; Cognasse, F.; Nicolas, J.; Vocanson, M. CD8+ T cells mediate ultraviolet A-induced immunomodulation in a model of extracorporeal photochemotherapy. Eur. J. Immunol. 2020, 50, 725–735. [Google Scholar] [CrossRef]
- Dieterlen, M.-T.; Garbade, J.; Misfeld, M.; Lehmann, S.; Klaeske, K.; Borger, M.A.; Barten, M.J. Indication-specific immunomodulatory effects of extracorporeal photopheresis: A pilot study in heart transplanted patients. J. Clin. Apher. 2018, 33, 591–599. [Google Scholar] [CrossRef]
- Hachem, R.; Corris, P. Extracorporeal Photopheresis for Bronchiolitis Obliterans Syndrome after Lung Transplantation. Transplantation 2018, 102, 1059–1065. [Google Scholar] [CrossRef]
- Robinson, C.A.; Inci, I.; Naegeli, M.; Murer, C.; Schuurmans, M.M.; Urosevic-Maiwald, M.; Schüpbach, R.; Weder, W.; Benden, C. Extracorporeal photopheresis as second-line treatment therapy in life-threatening primary graft dysfunction following lung transplantation. Pediatr. Transplant. 2018, 22, e13145. [Google Scholar] [CrossRef]
- Lamioni, A.; Parisi, F.; Isacchi, G.; Giorda, E.; Di Cesare, S.; Landolfo, A.; Cenci, F.; Bottazzo, G.F.; Carsetti, R. The Immunological Effects of Extracorporeal Photopheresis Unraveled: Induction of Tolerogenic Dendritic Cells In Vitro and Regulatory T Cells In Vivo. Transplantation 2005, 79, 846–850. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Passerini, L.; Piening, B.D.; Uyeda, M.J.; Goodwin, M.; Gregori, S.; Snyder, M.P.; Bertaina, A.; Roncarolo, M.; Bacchetta, R. Human-engineered Treg-like cells suppress FOXP3-deficient T cells but preserve adaptive immune responses in vivo. Clin. Transl. Immunol. 2020, 9, e1214. [Google Scholar] [CrossRef] [PubMed]
- Bergantini, L.; D’Alessandro, M.; De Vita, E.; Perillo, F.; Fossi, A.; Luzzi, L.; Paladini, P.; Perrone, A.; Rottoli, P.; Sestini, P.; et al. Regulatory and Effector Cell Disequilibrium in Patients with Acute Cellular Rejection and Chronic Lung Allograft Dysfunction after Lung Transplantation: Comparison of Peripheral and Alveolar Distribution. Cells 2021, 10, 780. [Google Scholar] [CrossRef] [PubMed]
- Mas, V.R.; Dumur, C.I.; Scian, M.J.; Gehrau, R.C.; Maluf, D.G. MicroRNAs as Biomarkers in Solid Organ Transplantation. Am. J. Transplant. 2013, 13, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, A.; Krams, S.M.; Martinez, O.M. MicroRNAs as Immune Regulators: Implications for Transplantation. Am. J. Transplant. 2010, 10, 713–719. [Google Scholar] [CrossRef] [Green Version]
- Montoya, R.T.; López-Godino, O.; Garcia-Barbera, N.; Cifuentes-Riquelme, R.; Sola, M.; Heras, I.; Perotti, C.; Garcia, V.V.; González-Conejero, R.; Del Fante, C.; et al. Identification of Circulating microRNA Signatures As Potential Noninvasive Biomarkers for Prediction to Response to Extracorporeal Photoapheresis in Patients with Graft Versus Host Disease. Blood 2019, 134, 4466. [Google Scholar] [CrossRef]
- Cabral, A.; Candido, D.D.S.; Monteiro, S.M.; Lemos, F.; Saitovitch, D.; Noronha, I.L.; Alves, L.; Geraldo, M.V.; Kalil, J.; Cunha-Neto, E.; et al. Differential microRNA Profile in Operational Tolerance: A Potential Role in Favoring Cell Survival. Front. Immunol. 2019, 10, 740. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.; Vigorito, E.; Clare, S.; Warren, M.V.; Couttet, P.; Soond, D.R.; van Dongen, S.; Grocock, R.J.; Das, P.P.; Miska, E.A.; et al. Requirement of bic/microRNA-155 for Normal Immune Function. Science 2007, 316, 608–611. [Google Scholar] [CrossRef] [Green Version]
- Kohlhaas, S.; Garden, O.A.; Scudamore, C.; Turner, M.; Okkenhaug, K.; Vigorito, E. Cutting Edge: The Foxp3 Target miR-155 Contributes to the Development of Regulatory T Cells. J. Immunol. 2009, 182, 2578–2582. [Google Scholar] [CrossRef]
- Lu, L.-F.; Boldin, M.; Chaudhry, A.; Lin, L.-L.; Taganov, K.D.; Hanada, T.; Yoshimura, A.; Baltimore, D.; Rudensky, A.Y. Function of miR-146a in Controlling Treg Cell-Mediated Regulation of Th1 Responses. Cell 2010, 142, 914–929. [Google Scholar] [CrossRef] [Green Version]
- Nuin, B.C.R.; De Bejar, Á.; Martinez-Alarcon, L.; Herrero, J.I.; Martínez-Cáceres, C.M.; Ramírez, P.; Baroja-Mazo, A.; Pons-Miñano, J.A. Differential profile of activated regulatory T cell subsets and microRNAs in tolerant liver transplant recipients. Liver Transplant. 2017, 23, 933–945. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Xie, N.; Cui, H.; Banerjee, S.; Abraham, E.; Thannickal, V.J.; Liu, G. miR-31 is a negative regulator of fibrogenesis and pulmonary fibrosis. FASEB J. 2012, 26, 3790–3799. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Jiang, H.-Y.; Li, J.; Tang, H.-C.; Zhang, X.-M.; Wang, X.-R.; Du, J.-T.; Li, H.-B.; Xu, G. MicroRNA-23b promotes tolerogenic properties of dendritic cells in vitro through inhibiting Notch1/NF-κB signalling pathways. Allergy 2012, 67, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; O’Connell, R.M. MiR-23b is a safeguard against autoimmunity. Nat. Med. 2012, 18, 1009–1010. [Google Scholar] [CrossRef]
- Liu, X.; Ni, S.; Li, C.; Xu, N.; Chen, W.; Wu, M.; van Wijnen, A.J.; Wang, Y. Circulating microRNA-23b as a new biomarker for rheumatoid arthritis. Gene 2019, 712, 143911. [Google Scholar] [CrossRef]
- Yang, Z.; Xiao, Z.; Guo, H.; Fang, X.; Liang, J.; Zhu, J.; Yang, J.; Li, H.; Pan, R.; Yuan, S.; et al. Novel role of the clustered miR-23b-3p and miR-27b-3p in enhanced expression of fibrosis-associated genes by targeting TGFBR3 in atrial fibroblasts. J. Cell. Mol. Med. 2019, 23, 3246–3256. [Google Scholar] [CrossRef]
- Song, S.-S.; Yuan, P.-F.; Chen, J.-Y.; Fu, J.-J.; Wu, H.-X.; Lu, J.T.; Wei, W. TGF-β favors bone marrow-derived dendritic cells to acquire tolerogenic properties. Immunol. Investig. 2014, 43, 360–369. [Google Scholar] [CrossRef]
- Meng, X.-M.; Huang, X.R.; Xiao, J.; Chung, A.C.; Qin, W.; Chen, H.-Y.; Lan, H.Y. Disruption of Smad4 impairs TGF-β/Smad3 and Smad7 transcriptional regulation during renal inflammation and fibrosis in vivo and in vitro. Kidney Int. 2012, 81, 266–279. [Google Scholar] [CrossRef] [Green Version]
- Morishita, Y.; Yoshizawa, H.; Watanabe, M.; Ishibashi, K.; Muto, S.; Kusano, E.; Nagata, D. siRNAs targeted to Smad4 prevent renal fibrosis in vivo. Sci. Rep. 2014, 4, 6424. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| On-Line (n = 16) | Off-Line (n = 10) | |
|---|---|---|
| Males (n, %) | 11 (69%) | 7 (70%) |
| Age at transplant (mean, DS) | 49 ± 11 | 43 ± 7 |
| Age at ECP (mean, DS) | 54 ± 11 | 52 ± 9 |
| Underlying diagnosis: Chronic obstructive pulmonary disease (COPD) Idiopathic pulmonary fibrosis (IPF) Cystic fibrosis (CF) Other | 7 (44%) 2 (12.5%) 1 (6%) 6 (37.5%) | 0 4 (40%) 3 (30%) 3 (30%) |
| Bilateral lung transplantation (n, %) | 11 (69%) | 7 (70%) |
| ECP outcome at last follow-up: Responder Non responder | 11 (69%) 5 (31%) | 7 (70%) 3 (30%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bozzini, S.; Del Fante, C.; Morosini, M.; Berezhinskiy, H.O.; Auner, S.; Cattaneo, E.; Della Zoppa, M.; Pandolfi, L.; Cacciatore, R.; Perotti, C.; et al. Mechanisms of Action of Extracorporeal Photopheresis in the Control of Bronchiolitis Obliterans Syndrome (BOS): Involvement of Circulating miRNAs. Cells 2022, 11, 1117. https://doi.org/10.3390/cells11071117
Bozzini S, Del Fante C, Morosini M, Berezhinskiy HO, Auner S, Cattaneo E, Della Zoppa M, Pandolfi L, Cacciatore R, Perotti C, et al. Mechanisms of Action of Extracorporeal Photopheresis in the Control of Bronchiolitis Obliterans Syndrome (BOS): Involvement of Circulating miRNAs. Cells. 2022; 11(7):1117. https://doi.org/10.3390/cells11071117
Chicago/Turabian StyleBozzini, Sara, Claudia Del Fante, Monica Morosini, Hatice Oya Berezhinskiy, Sophia Auner, Elena Cattaneo, Matteo Della Zoppa, Laura Pandolfi, Rosalia Cacciatore, Cesare Perotti, and et al. 2022. "Mechanisms of Action of Extracorporeal Photopheresis in the Control of Bronchiolitis Obliterans Syndrome (BOS): Involvement of Circulating miRNAs" Cells 11, no. 7: 1117. https://doi.org/10.3390/cells11071117
APA StyleBozzini, S., Del Fante, C., Morosini, M., Berezhinskiy, H. O., Auner, S., Cattaneo, E., Della Zoppa, M., Pandolfi, L., Cacciatore, R., Perotti, C., Hoetzenecker, K., Jaksch, P., Benazzo, A., & Meloni, F. (2022). Mechanisms of Action of Extracorporeal Photopheresis in the Control of Bronchiolitis Obliterans Syndrome (BOS): Involvement of Circulating miRNAs. Cells, 11(7), 1117. https://doi.org/10.3390/cells11071117