
Mesenchymal Stromal Cell-Derived Extracellular Vesicles for Neonatal Lung Disease: Tiny Particles, Major Promise, Rigorous Requirements for Clinical Translation


Abstract
:1. A Brief Description of Bronchopulmonary Dysplasia
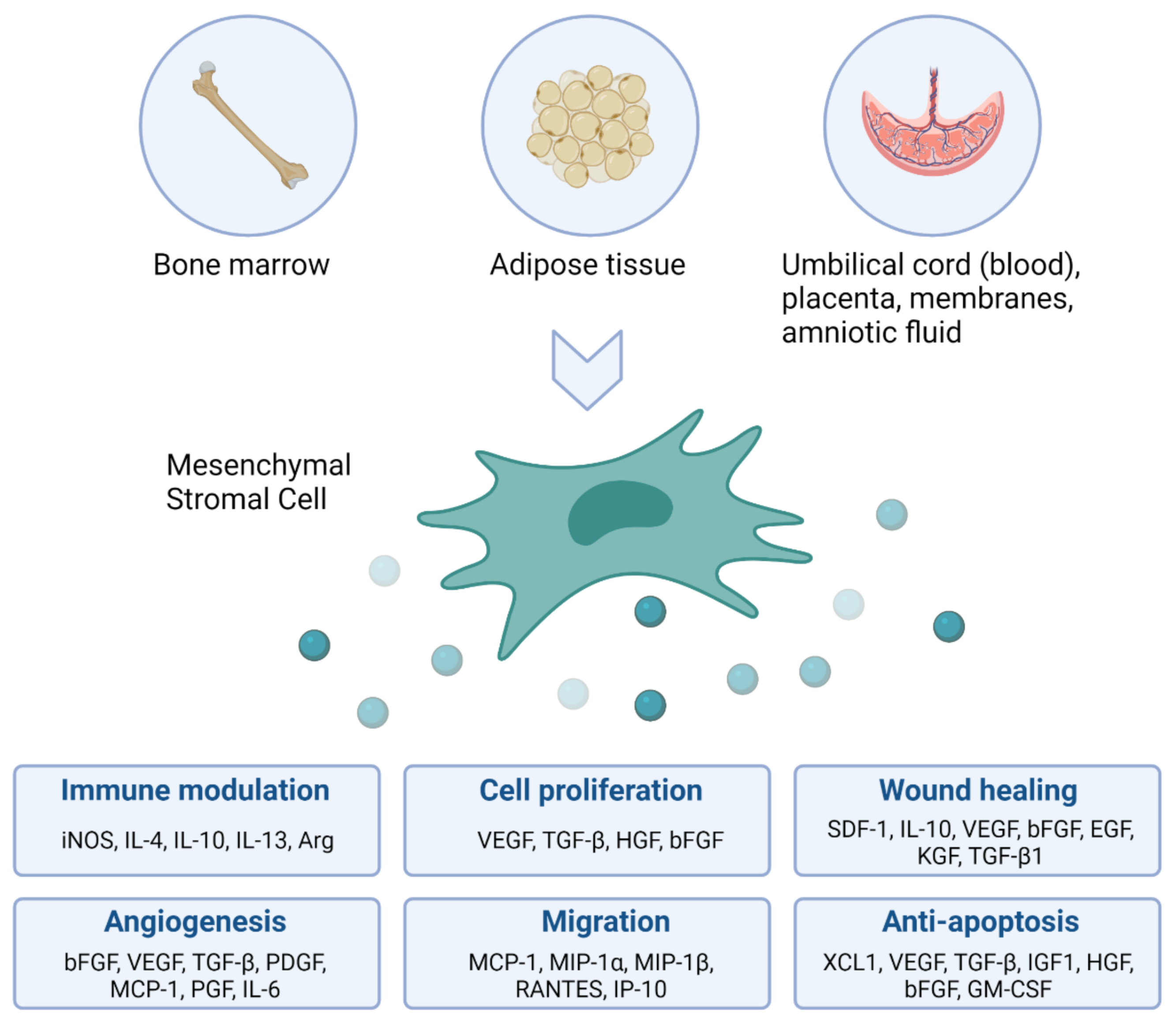
2. The Rationale for Using Mesenchymal Stromal Cells as Immune Modulating Agents in Patients with BPD
3. Cell Therapy without the Cell: The Promise of Extracellular Vesicles
4. Current State-of-the-Art for EV-Based Therapy in BPD Animal Models
5. EV-Based Therapy in BPD Animal Models: Additional Research Required before Clinical Translation
6. Influencing the Bioactive Properties of EVs
7. Standardization of Isolation and Characterization Methods for EV Research
8. Future Directions for the EV Field
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Burri, P.H. Fetal and Postnatal Development of the Lung. Annu. Rev. Physiol. 1984, 46, 617–628. [Google Scholar] [CrossRef] [PubMed]
- McGrath-Morrow, S.A.; Collaco, J.M. Bronchopulmonary Dysplasia: What Are Its Links to COPD? Adv. Respir. Dis. 2019, 13, 1753466619892492. [Google Scholar] [CrossRef] [PubMed]
- Jobe, A.H.; Bancalari, E. Bronchopulmonary Dysplasia. Am. J. Respir. Crit. Care Med. 2001, 163, 1723–1729. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.S.; Sankaran, K.; Aziz, K.; Allen, A.C.; Seshia, M.; Ohlsson, A.; Lee, S.K. Canadian Neonatal Network Outcomes of Preterm Infants <29 Weeks Gestation over 10-Year Period in Canada: A Cause for Concern? J. Perinatol. 2012, 32, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Thébaud, B.; Goss, K.N.; Laughon, M.; Whitsett, J.A.; Abman, S.H.; Steinhorn, R.H.; Aschner, J.L.; Davis, P.G.; McGrath-Morrow, S.A.; Soll, R.F.; et al. Bronchopulmonary Dysplasia. Nat. Rev. Dis. Primers 2019, 5, 78. [Google Scholar] [CrossRef]
- Jobe, A.H. What Is BPD in 2012 and What Will BPD Become? Early Hum. Dev. 2012, 88 (Suppl. 2), S27–S28. [Google Scholar] [CrossRef]
- Wong, P.M.; Lees, A.N.; Louw, J.; Lee, F.Y.; French, N.; Gain, K.; Murray, C.P.; Wilson, A.; Chambers, D.C. Emphysema in Young Adult Survivors of Moderate-to-Severe Bronchopulmonary Dysplasia. Eur. Respir. J. 2008, 32, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Fawke, J.; Lum, S.; Kirkby, J.; Hennessy, E.; Marlow, N.; Rowell, V.; Thomas, S.; Stocks, J. Lung Function and Respiratory Symptoms at 11 Years in Children Born Extremely Preterm: The EPICure Study. Am. J. Respir. Crit. Care Med. 2010, 182, 237–245. [Google Scholar] [CrossRef]
- Tréluyer, L.; Jarreau, P.-H.; Marchand-Martin, L.; Benhammou, V.; Nuytten, A.; Berquin, P.; Marret, S.; Pierrat, V.; Ancel, P.-Y.; Torchin, H. Bronchopulmonary Dysplasia and Risk of Developmental Delay: An EPIPAGE-2 Cohort Study. Neonatology 2021, 119, 124–128. [Google Scholar] [CrossRef]
- Goss, K.N.; Beshish, A.G.; Barton, G.P.; Haraldsdottir, K.; Levin, T.S.; Tetri, L.H.; Battiola, T.J.; Mulchrone, A.M.; Pegelow, D.F.; Palta, M.; et al. Early Pulmonary Vascular Disease in Young Adults Born Preterm. Am. J. Respir. Crit. Care Med. 2018, 198, 1549–1558. [Google Scholar] [CrossRef]
- Friedenstein, A.J.; Chailakhjan, R.K.; Lalykina, K.S. The Development of Fibroblast Colonies in Monolayer Cultures of Guinea-Pig Bone Marrow and Spleen Cells. Cell Tissue Kinet 1970, 3, 393–403. [Google Scholar] [CrossRef]
- Friedenstein, A.J.; Gorskaja, J.F.; Kulagina, N.N. Fibroblast Precursors in Normal and Irradiated Mouse Hematopoietic Organs. Exp. Hematol. 1976, 4, 267–274. [Google Scholar]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Luk, F.; Carreras-Planella, L.; Korevaar, S.S.; de Witte, S.F.H.; Borràs, F.E.; Betjes, M.G.H.; Baan, C.C.; Hoogduijn, M.J.; Franquesa, M. Inflammatory Conditions Dictate the Effect of Mesenchymal Stem or Stromal Cells on B Cell Function. Front. Immunol. 2017, 8, 1042. [Google Scholar] [CrossRef] [Green Version]
- Ge, W.; Jiang, J.; Arp, J.; Liu, W.; Garcia, B.; Wang, H. Regulatory T-Cell Generation and Kidney Allograft Tolerance Induced by Mesenchymal Stem Cells Associated with Indoleamine 2,3-Dioxygenase Expression. Transplantation 2010, 90, 1312–1320. [Google Scholar] [CrossRef]
- Spaggiari, G.M.; Capobianco, A.; Abdelrazik, H.; Becchetti, F.; Mingari, M.C.; Moretta, L. Mesenchymal Stem Cells Inhibit Natural Killer-Cell Proliferation, Cytotoxicity, and Cytokine Production: Role of Indoleamine 2,3-Dioxygenase and Prostaglandin E2. Blood 2008, 111, 1327–1333. [Google Scholar] [CrossRef]
- Obermajer, N.; Popp, F.C.; Soeder, Y.; Haarer, J.; Geissler, E.K.; Schlitt, H.J.; Dahlke, M.H. Conversion of Th17 into IL-17A(Neg) Regulatory T Cells: A Novel Mechanism in Prolonged Allograft Survival Promoted by Mesenchymal Stem Cell-Supported Minimized Immunosuppressive Therapy. J. Immunol. 2014, 193, 4988–4999. [Google Scholar] [CrossRef] [Green Version]
- Németh, K.; Leelahavanichkul, A.; Yuen, P.S.T.; Mayer, B.; Parmelee, A.; Doi, K.; Robey, P.G.; Leelahavanichkul, K.; Koller, B.H.; Brown, J.M.; et al. Bone Marrow Stromal Cells Attenuate Sepsis via Prostaglandin E(2)-Dependent Reprogramming of Host Macrophages to Increase Their Interleukin-10 Production. Nat. Med. 2009, 15, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Luz-Crawford, P.; Djouad, F.; Toupet, K.; Bony, C.; Franquesa, M.; Hoogduijn, M.J.; Jorgensen, C.; Noël, D. Mesenchymal Stem Cell-Derived Interleukin 1 Receptor Antagonist Promotes Macrophage Polarization and Inhibits B Cell Differentiation. Stem Cells 2016, 34, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Pierro, M.; Ionescu, L.; Montemurro, T.; Vadivel, A.; Weissmann, G.; Oudit, G.; Emery, D.; Bodiga, S.; Eaton, F.; Péault, B.; et al. Short-Term, Long-Term and Paracrine Effect of Human Umbilical Cord-Derived Stem Cells in Lung Injury Prevention and Repair in Experimental Bronchopulmonary Dysplasia. Thorax 2013, 68, 475–484. [Google Scholar] [CrossRef] [Green Version]
- Lesage, F.; Jimenez, J.; Toelen, J.; Deprest, J. Preclinical Evaluation of Cell-Based Strategies to Prevent or Treat Bronchopulmonary Dysplasia in Animal Models: A Systematic Review. J. Matern. Fetal. Neonatal. Med. 2018, 31, 958–966. [Google Scholar] [CrossRef]
- Augustine, S.; Avey, M.T.; Harrison, B.; Locke, T.; Ghannad, M.; Moher, D.; Thébaud, B. Mesenchymal Stromal Cell Therapy in Bronchopulmonary Dysplasia: Systematic Review and Meta-Analysis of Preclinical Studies. Stem Cells Transl. Med. 2017, 6, 2079–2093. [Google Scholar] [CrossRef]
- Chang, Y.S.; Ahn, S.Y.; Yoo, H.S.; Sung, S.I.; Choi, S.J.; Oh, W.I.; Park, W.S. Mesenchymal Stem Cells for Bronchopulmonary Dysplasia: Phase 1 Dose-Escalation Clinical Trial. J. Pediatr. 2014, 164, 966–972.e6. [Google Scholar] [CrossRef]
- Lim, R.; Malhotra, A.; Tan, J.; Chan, S.T.; Lau, S.; Zhu, D.; Mockler, J.C.; Wallace, E.M. First-In-Human Administration of Allogeneic Amnion Cells in Premature Infants with Bronchopulmonary Dysplasia: A Safety Study. Stem Cells Transl. Med. 2018, 7, 628–635. [Google Scholar] [CrossRef] [Green Version]
- Powell, S.B.; Silvestri, J.M. Safety of Intratracheal Administration of Human Umbilical Cord Blood Derived Mesenchymal Stromal Cells in Extremely Low Birth Weight Preterm Infants. J. Pediatr. 2019, 210, 209–213.e2. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Chang, Y.S.; Lee, M.H.; Sung, S.I.; Lee, B.S.; Kim, K.S.; Kim, A.-R.; Park, W.S. Stem Cells for Bronchopulmonary Dysplasia in Preterm Infants: A Randomized Controlled Phase II Trial. Stem Cells Transl. Med. 2021, 10, 1129–1137. [Google Scholar] [CrossRef]
- Lo Cicero, A.; Stahl, P.D.; Raposo, G. Extracellular Vesicles Shuffling Intercellular Messages: For Good or for Bad. Curr. Opin. Cell Biol. 2015, 35, 69–77. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Ban, J.-J.; Lee, M.; Im, W.; Kim, M. Low PH Increases the Yield of Exosome Isolation. Biochem. Biophys. Res. Commun. 2015, 461, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Yang, F.; Miao, S.; Liu, W.; Wang, C.; Shu, Y.; Shen, H. Role of Hypoxia-Induced Exosomes in Tumor Biology. Mol. Cancer 2018, 17, 120. [Google Scholar] [CrossRef]
- Willis, G.R.; Fernandez-Gonzalez, A.; Anastas, J.; Vitali, S.H.; Liu, X.; Ericsson, M.; Kwong, A.; Mitsialis, S.A.; Kourembanas, S. Mesenchymal Stromal Cell Exosomes Ameliorate Experimental Bronchopulmonary Dysplasia and Restore Lung Function through Macrophage Immunomodulation. Am. J. Respir. Crit. Care Med. 2018, 197, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Willis, G.R.; Fernandez-Gonzalez, A.; Reis, M.; Yeung, V.; Liu, X.; Ericsson, M.; Andrews, N.A.; Mitsialis, S.A.; Kourembanas, S. Mesenchymal Stromal Cell-Derived Small Extracellular Vesicles Restore Lung Architecture and Improve Exercise Capacity in a Model of Neonatal Hyperoxia-Induced Lung Injury. J. Extracell Vesicles 2020, 9, 1790874. [Google Scholar] [CrossRef] [PubMed]
- Reis, M.; Willis, G.R.; Fernandez-Gonzalez, A.; Yeung, V.; Taglauer, E.; Magaletta, M.; Parsons, T.; Derr, A.; Liu, X.; Maehr, R.; et al. Mesenchymal Stromal Cell-Derived Extracellular Vesicles Restore Thymic Architecture and T Cell Function Disrupted by Neonatal Hyperoxia. Front. Immunol. 2021, 12, 640595. [Google Scholar] [CrossRef]
- Willis, G.R.; Reis, M.; Gheinani, A.H.; Fernandez-Gonzalez, A.; Taglauer, E.S.; Yeung, V.; Liu, X.; Ericsson, M.; Haas, E.; Mitsialis, S.A.; et al. Extracellular Vesicles Protect the Neonatal Lung from Hyperoxic Injury through the Epigenetic and Transcriptomic Reprogramming of Myeloid Cells. Am. J. Respir. Crit. Care Med. 2021, 204, 1418–1432. [Google Scholar] [CrossRef]
- Porzionato, A.; Zaramella, P.; Dedja, A.; Guidolin, D.; Van Wemmel, K.; Macchi, V.; Jurga, M.; Perilongo, G.; De Caro, R.; Baraldi, E.; et al. Intratracheal Administration of Clinical-Grade Mesenchymal Stem Cell-Derived Extracellular Vesicles Reduces Lung Injury in a Rat Model of Bronchopulmonary Dysplasia. Am. J. Physiol. Lung Cell Mol. Physiol. 2019, 316, L6–L19. [Google Scholar] [CrossRef]
- Braun, R.K.; Chetty, C.; Balasubramaniam, V.; Centanni, R.; Haraldsdottir, K.; Hematti, P.; Eldridge, M.W. Intraperitoneal Injection of MSC-Derived Exosomes Prevent Experimental Bronchopulmonary Dysplasia. Biochem. Biophys. Res. Commun. 2018, 503, 2653–2658. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Park, W.S.; Kim, Y.E.; Sung, D.K.; Sung, S.I.; Ahn, J.Y.; Chang, Y.S. Vascular Endothelial Growth Factor Mediates the Therapeutic Efficacy of Mesenchymal Stem Cell-Derived Extracellular Vesicles against Neonatal Hyperoxic Lung Injury. Exp. Mol. Med. 2018, 50, 26. [Google Scholar] [CrossRef] [Green Version]
- Chaubey, S.; Thueson, S.; Ponnalagu, D.; Alam, M.A.; Gheorghe, C.P.; Aghai, Z.; Singh, H.; Bhandari, V. Early Gestational Mesenchymal Stem Cell Secretome Attenuates Experimental Bronchopulmonary Dysplasia in Part via Exosome-Associated Factor TSG-6. Stem Cell Res. 2018, 9, 173. [Google Scholar] [CrossRef]
- Li, Z.; Gong, X.; Li, D.; Yang, X.; Shi, Q.; Ju, X. Intratracheal Transplantation of Amnion-Derived Mesenchymal Stem Cells Ameliorates Hyperoxia-Induced Neonatal Hyperoxic Lung Injury via Aminoacyl-Peptide Hydrolase. Int. J. Stem Cells 2020, 13, 221–236. [Google Scholar] [CrossRef] [Green Version]
- Abele, A.N.; Taglauer, E.S.; Almeda, M.; Wilson, N.; Abikoye, A.; Seedorf, G.J.; Mitsialis, S.A.; Kourembanas, S.; Abman, S. Antenatal Mesenchymal Stromal Cell Extracellular Vesicle Treatment Preserves Lung Development in a Model of Bronchopulmonary Dysplasia Due to Chorioamnionitis. Am. J. Physiol. Lung Cell Mol. Physiol. 2021. [Google Scholar] [CrossRef]
- Porzionato, A.; Zaramella, P.; Dedja, A.; Guidolin, D.; Bonadies, L.; Macchi, V.; Pozzobon, M.; Jurga, M.; Perilongo, G.; De Caro, R.; et al. Intratracheal Administration of Mesenchymal Stem Cell-Derived Extracellular Vesicles Reduces Lung Injuries in a Chronic Rat Model of Bronchopulmonary Dysplasia. Am. J. Physiol. Lung Cell Mol. Physiol. 2021, 320, L688–L704. [Google Scholar] [CrossRef]
- You, J.; Zhou, O.; Liu, J.; Zou, W.; Zhang, L.; Tian, D.; Dai, J.; Luo, Z.; Liu, E.; Fu, Z.; et al. Human Umbilical Cord Mesenchymal Stem Cell-Derived Small Extracellular Vesicles Alleviate Lung Injury in Rat Model of Bronchopulmonary Dysplasia by Affecting Cell Survival and Angiogenesis. Stem Cells Dev. 2020, 29, 1520–1532. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, Z.; Li, J.; Zhong, H.; Yuan, R.; Deng, Z.; Wu, X. Mechanism of Adipose-Derived Mesenchymal Stem Cell-Derived Extracellular Vesicles Carrying MiR-21-5p in Hyperoxia-Induced Lung Injury. Stem Cell Rev. Rep. 2021, 18, 1007–1024. [Google Scholar] [CrossRef]
- Lithopoulos, M.A.; Strueby, L.; O’Reilly, M.; Zhong, S.; Möbius, M.A.; Eaton, F.; Fung, M.; Hurskainen, M.; Cyr-Depauw, C.; Suen, C.; et al. Pulmonary and Neurologic Effects of Mesenchymal Stromal Cell Extracellular Vesicles in a Multifactorial Lung Injury Model. Am. J. Respir. Crit Care Med. 2022. [Google Scholar] [CrossRef]
- Ai, D.; Shen, J.; Sun, J.; Zhu, Z.; Gao, R.; Du, Y.; Yuan, L.; Chen, C.; Zhou, J. Mesenchymal Stem Cell-Derived Extracellular Vesicles Suppress Hyperoxia-Induced Transdifferentiation of Rat Alveolar Type 2 Epithelial Cells. Stem Cells Dev. 2022, 31, 53–66. [Google Scholar] [CrossRef]
- Qin, S.; Chen, M.; Ji, H.; Liu, G.-Y.; Mei, H.; Li, K.; Chen, T. MiR-21-5p Regulates Type II Alveolar Epithelial Cell Apoptosis in Hyperoxic Acute Lung Injury. Mol. Med. Rep. 2018, 17, 5796–5804. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Saegusa, C.; Schehr, A.; Grant, S.; Whitsett, J.A.; Ikegami, M. C/EBP{alpha} Is Required for Pulmonary Cytoprotection during Hyperoxia. Am. J. Physiol. Lung Cell Mol. Physiol. 2009, 297, L286–L298. [Google Scholar] [CrossRef] [Green Version]
- Abdelwahab, E.M.M.; Rapp, J.; Feller, D.; Csongei, V.; Pal, S.; Bartis, D.; Thickett, D.R.; Pongracz, J.E. Wnt Signaling Regulates Trans-Differentiation of Stem Cell like Type 2 Alveolar Epithelial Cells to Type 1 Epithelial Cells. Respir. Res. 2019, 20, 204. [Google Scholar] [CrossRef]
- Chang, Y.S.; Ahn, S.Y.; Jeon, H.B.; Sung, D.K.; Kim, E.S.; Sung, S.I.; Yoo, H.S.; Choi, S.J.; Oh, W.I.; Park, W.S. Critical Role of Vascular Endothelial Growth Factor Secreted by Mesenchymal Stem Cells in Hyperoxic Lung Injury. Am. J. Respir. Cell Mol. Biol. 2014, 51, 391–399. [Google Scholar] [CrossRef]
- Franco, M.; Roswall, P.; Cortez, E.; Hanahan, D.; Pietras, K. Pericytes Promote Endothelial Cell Survival through Induction of Autocrine VEGF-A Signaling and Bcl-w Expression. Blood 2011, 118, 2906–2917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Lin, X.; Dai, Y.; Hu, X.; Zhu, H.; Jiang, Y.; Zhang, S. Endometrial Stem Cells Repair Injured Endometrium and Induce Angiogenesis via AKT and ERK Pathways. Reproduction 2016, 152, 389–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tieu, A.; Hu, K.; Gnyra, C.; Montroy, J.; Fergusson, D.A.; Allan, D.S.; Stewart, D.J.; Thébaud, B.; Lalu, M.M. Mesenchymal Stromal Cell Extracellular Vesicles as Therapy for Acute and Chronic Respiratory Diseases: A Meta-Analysis. J. Extracell Vesicles 2021, 10, e12141. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.Y.; Chang, Y.S.; Sung, D.K.; Yoo, H.S.; Sung, S.I.; Choi, S.J.; Park, W.S. Cell Type-Dependent Variation in Paracrine Potency Determines Therapeutic Efficacy against Neonatal Hyperoxic Lung Injury. Cytotherapy 2015, 17, 1025–1035. [Google Scholar] [CrossRef] [Green Version]
- Wiklander, O.P.B.; Brennan, M.Á.; Lötvall, J.; Breakefield, X.O.; El Andaloussi, S. Advances in therapeutic applications of extracellular vesicles. Sci. Transl. Med. 2019, 11, eaav8521. [Google Scholar] [CrossRef]
- Guess, A.J.; Daneault, B.; Wang, R.; Bradbury, H.; La Perle, K.M.D.; Fitch, J.; Hedrick, S.L.; Hamelberg, E.; Astbury, C.; White, P.; et al. Safety Profile of Good Manufacturing Practice Manufactured Interferon γ-Primed Mesenchymal Stem/Stromal Cells for Clinical Trials. Stem Cells Transl. Med. 2017, 6, 1868–1879. [Google Scholar] [CrossRef]
- Markoutsa, E.; Mayilsamy, K.; Gulick, D.; Mohapatra, S.S.; Mohapatra, S. Extracellular Vesicles Derived from Inflammatory-Educated Stem Cells Reverse Brain Inflammation-Implication of MiRNAs. Mol. Ther. 2022, 30, 816–830. [Google Scholar] [CrossRef]
- Li, P.; Kaslan, M.; Lee, S.H.; Yao, J.; Gao, Z. Progress in Exosome Isolation Techniques. Theranostics 2017, 7, 789–804. [Google Scholar] [CrossRef]
- Gardiner, C.; Di Vizio, D.; Sahoo, S.; Théry, C.; Witwer, K.W.; Wauben, M.; Hill, A.F. Techniques Used for the Isolation and Characterization of Extracellular Vesicles: Results of a Worldwide Survey. J. Extracell Vesicles 2016, 5, 32945. [Google Scholar] [CrossRef]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3–22. [Google Scholar] [CrossRef]
- Raposo, G.; Nijman, H.W.; Stoorvogel, W.; Liejendekker, R.; Harding, C.V.; Melief, C.J.; Geuze, H.J. B Lymphocytes Secrete Antigen-Presenting Vesicles. J. Exp. Med. 1996, 183, 1161–1172. [Google Scholar] [CrossRef]
- EV-TRACK Consortium; Van Deun, J.; Mestdagh, P.; Agostinis, P.; Akay, Ö.; Anand, S.; Anckaert, J.; Martinez, Z.A.; Baetens, T.; Beghein, E.; et al. EV-TRACK: Transparent Reporting and Centralizing Knowledge in Extracellular Vesicle Research. Nat. Methods 2017, 14, 228–232. [Google Scholar] [CrossRef]
- Van Deun, J.; Hendrix, A. EV-TRACK consortium Is Your Article EV-TRACKed? J. Extracell Vesicles 2017, 6, 1379835. [Google Scholar] [CrossRef] [Green Version]
- Witwer, K.W.; Goberdhan, D.C.; O’Driscoll, L.; Théry, C.; Welsh, J.A.; Blenkiron, C.; Buzás, E.I.; Di Vizio, D.; Erdbrügger, U.; Falcón-Pérez, J.M.; et al. Updating MISEV: Evolving the Minimal Requirements for Studies of Extracellular Vesicles. J. Extracell Vesicles 2021, 10, e12182. [Google Scholar] [CrossRef]
- Kim, D.-K.; Kang, B.; Kim, O.Y.; Choi, D.-S.; Lee, J.; Kim, S.R.; Go, G.; Yoon, Y.J.; Kim, J.H.; Jang, S.C.; et al. EVpedia: An Integrated Database of High-Throughput Data for Systemic Analyses of Extracellular Vesicles. J. Extracell Vesicles 2013, 2, 20384. [Google Scholar] [CrossRef]
- Kalra, H.; Simpson, R.J.; Ji, H.; Aikawa, E.; Altevogt, P.; Askenase, P.; Bond, V.C.; Borràs, F.E.; Breakefield, X.; Budnik, V.; et al. Vesiclepedia: A Compendium for Extracellular Vesicles with Continuous Community Annotation. PLoS Biol. 2012, 10, e1001450. [Google Scholar] [CrossRef] [Green Version]
- Simpson, R.J.; Kalra, H.; Mathivanan, S. ExoCarta as a Resource for Exosomal Research. J. Extracell Vesicles 2012, 1, 18374. [Google Scholar] [CrossRef]
- Li, S.; Li, Y.; Chen, B.; Zhao, J.; Yu, S.; Tang, Y.; Zheng, Q.; Li, Y.; Wang, P.; He, X.; et al. ExoRBase: A Database of CircRNA, LncRNA and MRNA in Human Blood Exosomes. Nucleic Acids Res. 2018, 46, D106–D112. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, Q.; Zhang, J.; Li, C.; Miao, Y.-R.; Lei, Q.; Li, Q.; Guo, A.-Y. EVmiRNA: A Database of MiRNA Profiling in Extracellular Vesicles. Nucleic Acids Res. 2019, 47, D89–D93. [Google Scholar] [CrossRef] [Green Version]
- Groot, M.; Lee, H. Sorting Mechanisms for MicroRNAs into Extracellular Vesicles and Their Associated Diseases. Cells 2020, 9, 1044. [Google Scholar] [CrossRef]
- Haraszti, R.A.; Didiot, M.-C.; Sapp, E.; Leszyk, J.; Shaffer, S.A.; Rockwell, H.E.; Gao, F.; Narain, N.R.; DiFiglia, M.; Kiebish, M.A.; et al. High-Resolution Proteomic and Lipidomic Analysis of Exosomes and Microvesicles from Different Cell Sources. J. Extracell Vesicles 2016, 5, 32570. [Google Scholar] [CrossRef]
- Lener, T.; Gimona, M.; Aigner, L.; Börger, V.; Buzas, E.; Camussi, G.; Chaput, N.; Chatterjee, D.; Court, F.A.; Del Portillo, H.A.; et al. Applying Extracellular Vesicles Based Therapeutics in Clinical Trials-an ISEV Position Paper. J. Extracell Vesicles 2015, 4, 30087. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Publication | Model | Species | EV Source | Dose Analysis | Dose | Route | Timing of EV Administration: Prevention | Timing of Data Collection |
|---|---|---|---|---|---|---|---|---|
| Willis 2018 [32] | Hyperoxia (75% O2, PN1-7) | Mouse | h UmC-MSC, hBM-MSC | Cell equivalent | 0.5 × 106 | IV | PN4 | PN7, PN14, PN42 |
| Porzionato 2019 [36] | Hyperoxia (60% O2, PN1-14) | Rat | h UmC-MSC | Particle count | 8 × 108 at PN3; 4.5 × 108 at PN7; 3 × 108 at PN10 | IT | PN3, PN7, PN10 | PN14 |
| Braun 2018 [37] | Hyperoxia (85% O2, PN1-14) | Rat | r BM-MSC | Protein concentration/particle count | 15 µg/3.4 × 109 | IP | PN1—14, daily | PN14, PN21, PN56 |
| Ahn 2018 [38] | Hyperoxia (90% O2, PN1-14) | Rat | h UCB-MSC | Protein concentration | 20 µg | IT | PN5 | PN14 |
| Chaubey 2018 [39] | Hyperoxia (95% O2, PN1-4) | Mouse | h UmC-MSC | Cell equivalent | 0.7 × 106 | IP | PN2, PN4 | PN14 |
| Li 2020 [40] | Hyperoxia (80% O2, PN1-14) | Rat | h AT-MSC | Protein concentration | 300 ng | IT | PN7 | PN14 |
| Willis 2020 [33] | Hyperoxia (75% O2, PN1-14) | Mouse | h UmC-MSC | Cell equivalent | Early: 0.5 × 106; Bolus late 1 × 106; Serial late: 1 × 106 | IV | Early: PN4; Bolus late: PN18, Serial late: PN18-25-32-39 | Early: PN60; Bolus late: PN28; Serial late: PN60 |
| Abele 2021 [41] | Chorioamnionitis (Endotoxin, E20) | Rat | h BM-MSC | Cell equivalent | 0.25 × 106 | IA | E20 | PN14 |
| Porzionato 2021 [42] | Hyperoxia (60% O2, PN0-14) | Rat | h UmC-MSC | Particle count | 8 × 108 at PN3; 4.5 × 108 at PN7; 3 × 108 at PN10 | IT | PN3, PN7, PN10 and PN21 | PN42 |
| Reis 2021 [34] | Hyperoxia (75% O2, PN1-7) | Mouse | h UmC-MSC | Cell equivalent | 0.5 × 106 | IV | PN4 | PN14 |
| Willis 2021 [35] | Hyperoxia (75% O2, PN1-14) | Mouse | h UmC-MSC | Cell equivalent | 0.5 × 106 | IV | PN4 | PN28 |
| You 2021 [43] | Hyperoxia (85% O2, PN0-14) | Rat | h UmC-MSC | Protein concentration | 20 µg | IT | PN7 | PN14 |
| Wu 2021 [44] | Hyperoxia (95% O2, PN1-3) | Mouse | m AT-MSC | Protein concentration | 30 or 300 ng | IT | PN1 | PN3 |
| Lithopoulos 2022 [45] | Endotoxin (PN7/8) + Ventilation (PN9/10, 40% O2, 8 h) | Mouse | h UmC-MSC | Protein concentration/Particle count | 0.005 μg/g; approximately 1 × 106 particles/g | IT | PN9/10 | 8 h after EV delivery |
| Ai 2022 [46] | Hyperoxia (75%, O2, PN1-14) | Rat | h UmC-MSC | Protein concentration | 10 or 15 µg | IP | PN4 | PN14, PN21, PN42 |
| REQ1: EV Quantification | REQ2: Protein Characterization | REQ3: Single EVs | ||||||
|---|---|---|---|---|---|---|---|---|
| Publication | Isolation Method | BCA | NTA | Presence of Trans-Membrane Proteins | Presence of Cytosolic Proteins | Absence of non-EV Proteins | TEM | NTA |
| Willis 2018 [32] | TFF + UC | ✓ | CD63, CD9, CD81 | HSP70 | ✓ | ✓ | ||
| Porzionato 2019 [36] | TFF | ✓ | CD63, CD9, CD81 | ANNEXIN V | ALBUMIN | ✓ | ||
| Braun 2018 [37] | UC | ✓ | ✓ | CD63, CD9, CD81 | ✓ | ✓ | ||
| Ahn 2018 [38] | UC | ✓ | ✓ | CD63, CD9 | GM130, FIBRILLARIN | ✓ | ✓ | |
| Chaubey 2018 [39] | UC | ✓ | ✓ | CD63, CD81 | ALIX1 | TGN48 | ✓ | ✓ |
| Li 2020 [40] | UC | ✓ | CD63, CD9, CD81 | HSP70 | ✓ | |||
| Willis 2020 [33] | TFF + UC | ✓ | CD63, CD9, CD81 | FLOT1, ALIX, TSG101 | GM130 | ✓ | ✓ | |
| Abele 2021 [41] | TFF + UC | ✓ | ✓ | ✓ | ||||
| Porzionato 2021 [42] | TFF | ✓ | CD63, CD9, CD81 | ANNEXIN V | ALBUMIN | ✓ | ||
| Reis 2021 [34] | TFF + UC | ✓ | CD63, CD81 | TSG101, SDCBP | CALNEXIN | ✓ | ✓ | |
| Willis 2021 [35] | TFF + UC | ✓ | CD63, CD9 | FLOT1, ALIX, TSG101 | GM130, CALNEXIN | ✓ | ✓ | |
| You 2020 [43] | UC | ✓ | ✓ | CD63 | ALIX | ✓ | ✓ | |
| Wu 2021 [44] | UC | ✓ | ✓ | CD63, CD9 | CALNEXIN | ✓ | ✓ | |
| Lithopoulos 2022 [45] | UC | ✓ | ✓ | CD63 | FLOT1 | CALNEXIN | ✓ | ✓ |
| Ai 2021 [46] | UC | ✓ | ✓ | CD63, CD9 | FLOT1 | ✓ | ✓ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lesage, F.; Thébaud, B. Mesenchymal Stromal Cell-Derived Extracellular Vesicles for Neonatal Lung Disease: Tiny Particles, Major Promise, Rigorous Requirements for Clinical Translation. Cells 2022, 11, 1176. https://doi.org/10.3390/cells11071176
Lesage F, Thébaud B. Mesenchymal Stromal Cell-Derived Extracellular Vesicles for Neonatal Lung Disease: Tiny Particles, Major Promise, Rigorous Requirements for Clinical Translation. Cells. 2022; 11(7):1176. https://doi.org/10.3390/cells11071176
Chicago/Turabian StyleLesage, Flore, and Bernard Thébaud. 2022. "Mesenchymal Stromal Cell-Derived Extracellular Vesicles for Neonatal Lung Disease: Tiny Particles, Major Promise, Rigorous Requirements for Clinical Translation" Cells 11, no. 7: 1176. https://doi.org/10.3390/cells11071176
APA StyleLesage, F., & Thébaud, B. (2022). Mesenchymal Stromal Cell-Derived Extracellular Vesicles for Neonatal Lung Disease: Tiny Particles, Major Promise, Rigorous Requirements for Clinical Translation. Cells, 11(7), 1176. https://doi.org/10.3390/cells11071176