
Interspecies Diversity of Osmotic Gradient Deformability of Red Blood Cells in Human and Seven Vertebrate Animal Species

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Volunteer Participants, Experimental Animals, and Blood Sampling
2.2. Hematological Measurements
2.3. Red Blood Cell Deformability (Conventional and Osmotic Gradient Ektacytometry)
2.4. Statistical Analysis
3. Results
3.1. Hematological Parameters
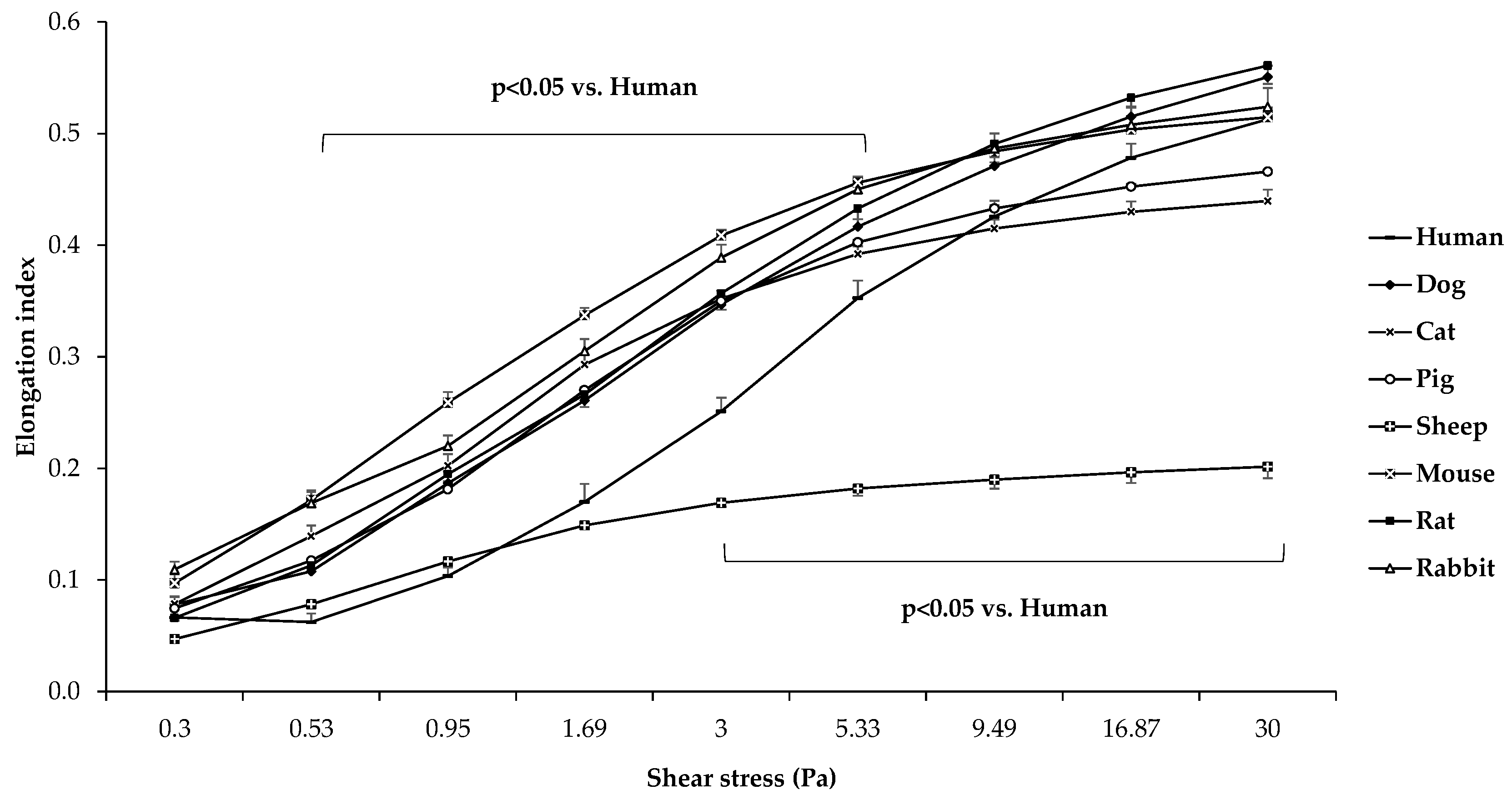
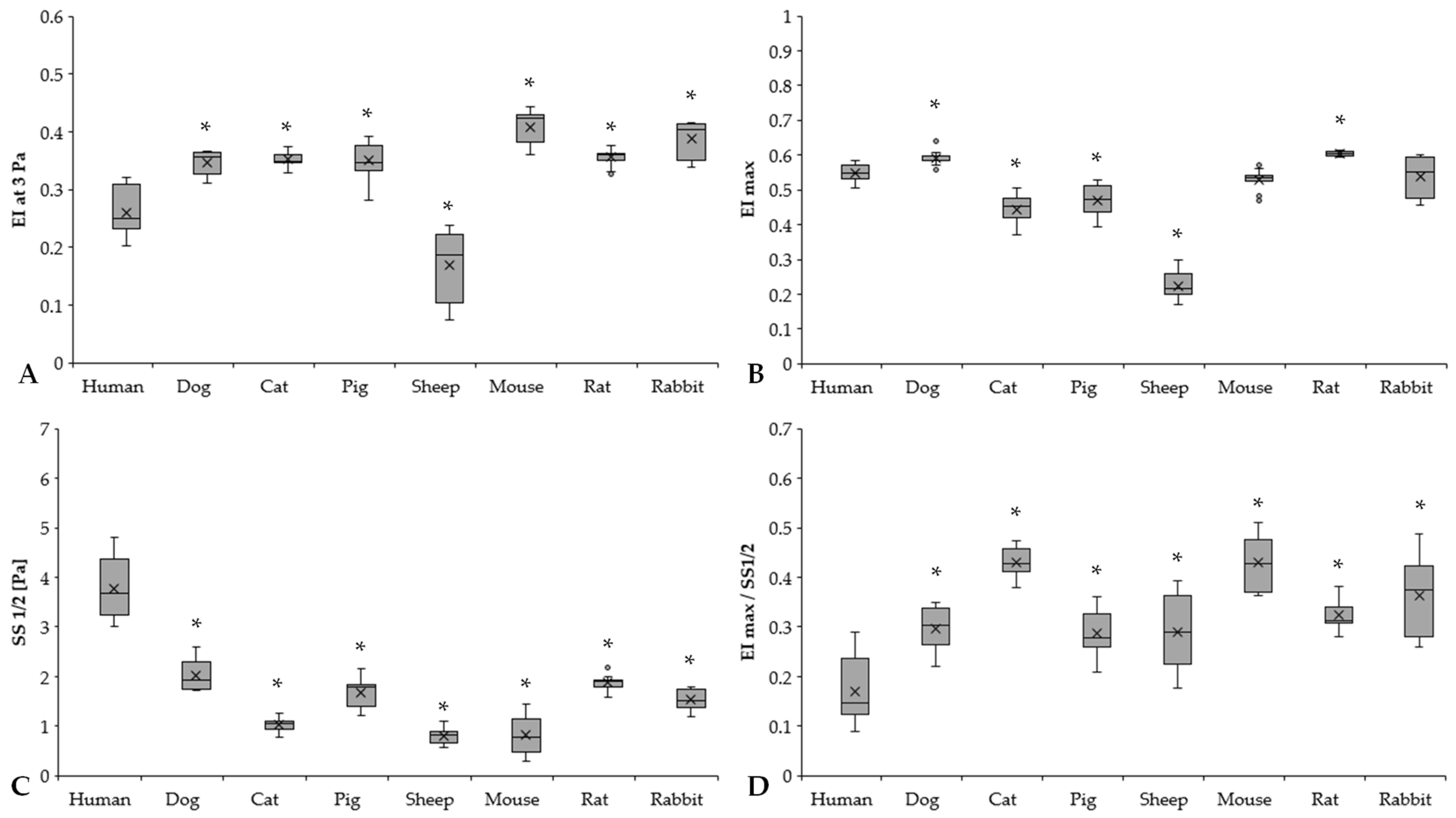
3.2. Red Blood Cell Deformability
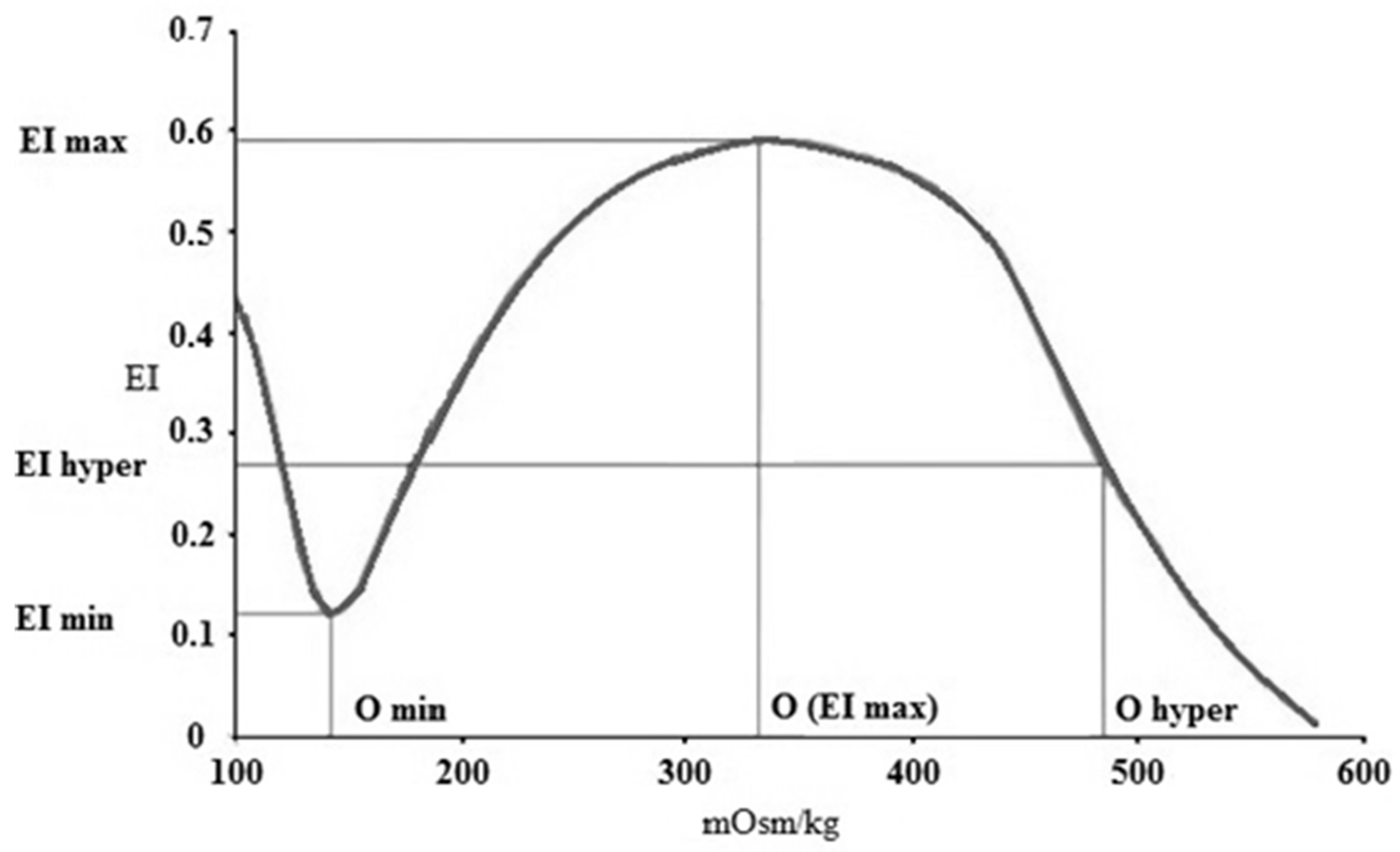
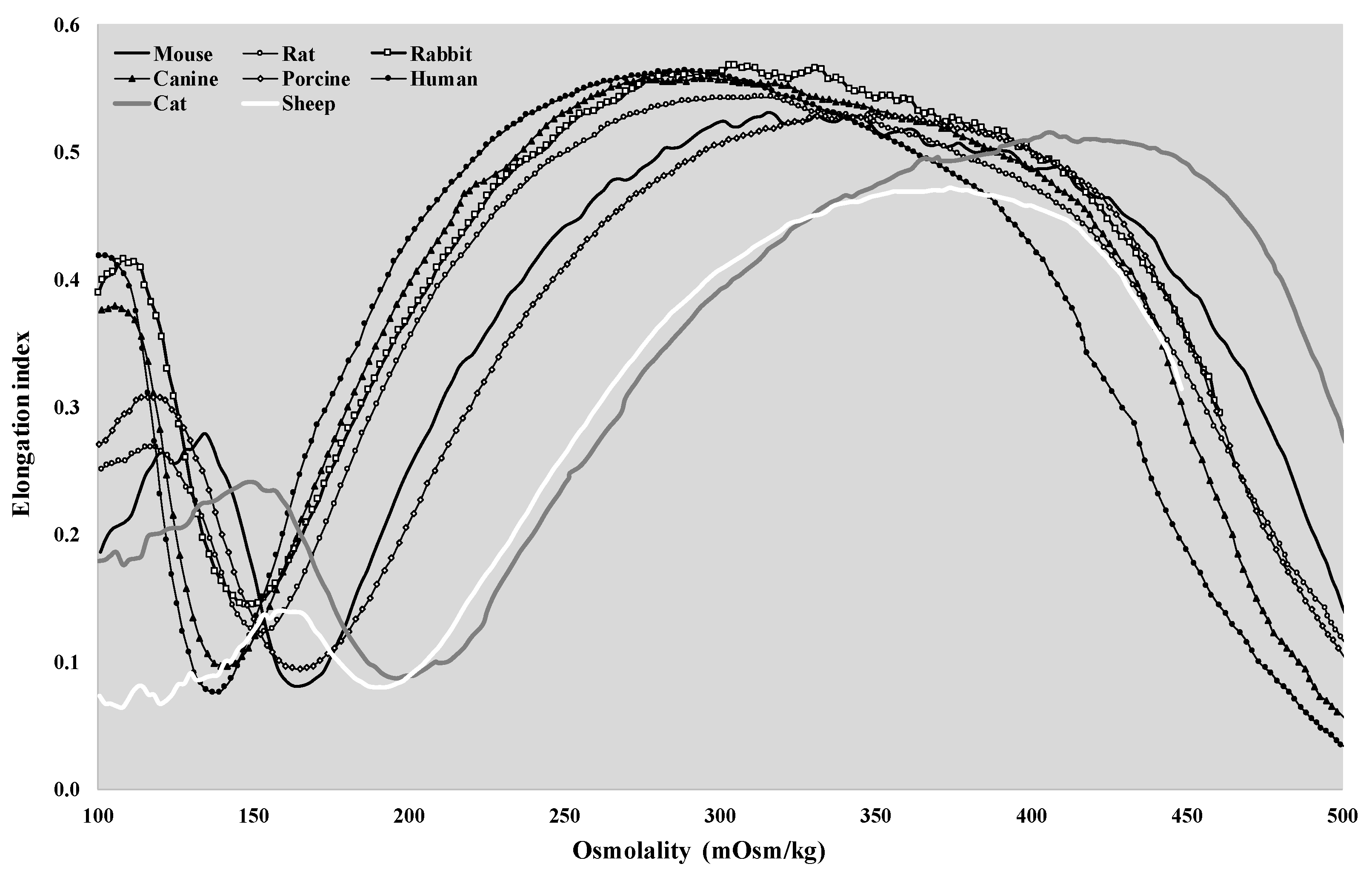
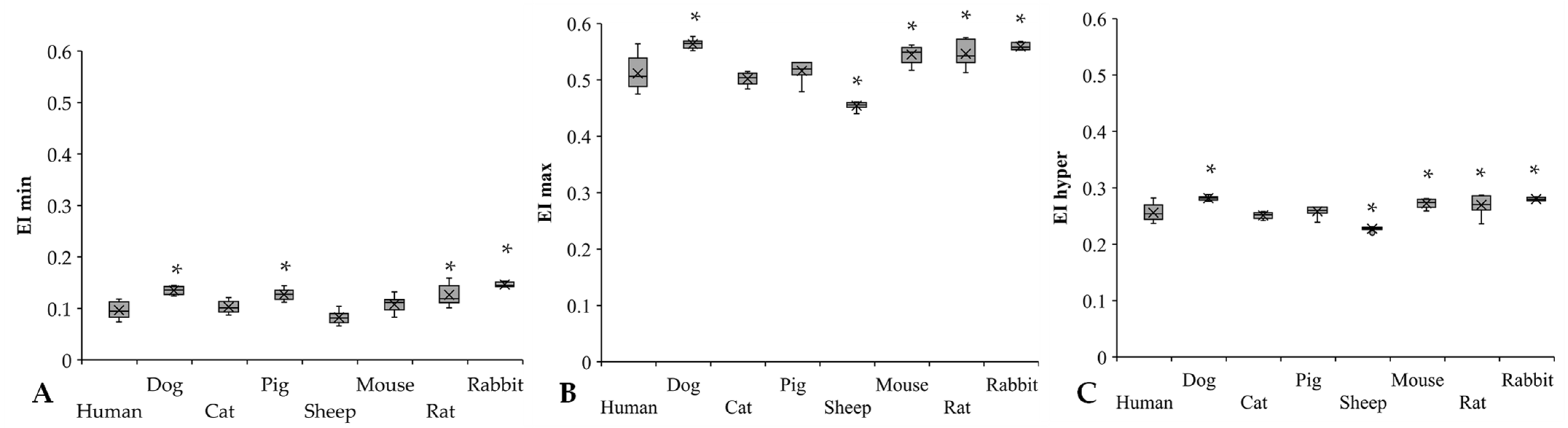
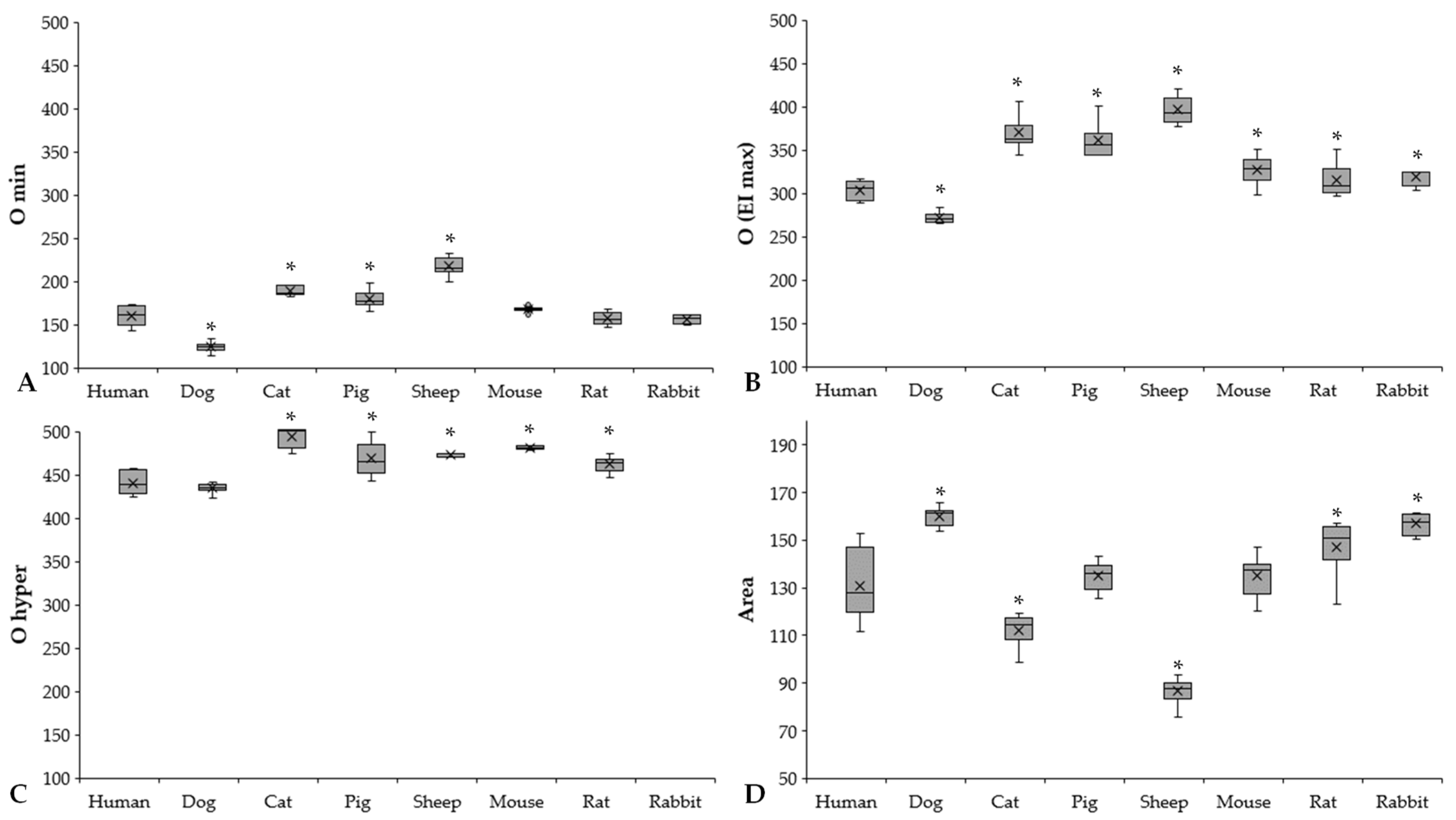
3.3. Osmotic Gradient Deformability
3.4. Correlation of Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baskurt, O.K.; Meiselman, H.J. Blood rheology and hemodynamics. Semin. Thromb. Hemost. 2003, 29, 435–450. [Google Scholar] [PubMed] [Green Version]
- Baskurt, O.K. In vivo correlates of altered blood rheology. Biorheology 2008, 45, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Baskurt, O.K. Mechanisms of blood rheology alterations. In Handbook of Hemorheology and Hemodynamics; Baskurt, O.K., Hardeman, M.R., Rampling, M.W., Meiselman, H.J., Eds.; IOS Press: Amsterdam, The Netherlands, 2007; pp. 170–190. [Google Scholar]
- Piagnerelli, M.; Boudjeltia, K.Z.; Vanhaeverbeek, M.; Vincent, J.L. Red blood cell rheology in sepsis. Intensive Care Med. 2003, 29, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Vayá, A.; Collado, S.; Dasí, M.A.; Pérez, M.L.; Hernandez, J.L.; Barragán, E. Erythrocyte deformability and aggregation in homozygous sickle cell disease. Clin. Hemorheol. Microcirc. 2014, 58, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Vayá, A.; Suescun, M.; Pardo, A.; Fuster, O. Erythrocyte deformability and hereditary elliptocytosis. Clin. Hemorheol. Microcirc. 2014, 58, 471–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemeth, N.; Deak, A.; Szentkereszty, Z.; Peto, K. Effects and influencing factors on hemorheological variables taken into consideration in surgical pathophysiology research. Clin. Hemorheol. Microcirc. 2018, 69, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.K.; Lim, Y.T.; Suh, J.S.; Hah, J.O.; Lee, J.M. Erythrocyte deformability reduction in various pediatric hematologic diseases. Clin. Hemorheol. Microcirc. 2020, 75, 361–367. [Google Scholar] [CrossRef]
- Nemeth, N.; Peto, K.; Magyar, Z.; Klarik, Z.; Varga, G.; Oltean, M.; Mantas, A.; Czigany, Z.; Tolba, R.H. Hemorheological and microcirculatory factors in liver ischemia-reperfusion injury-an update on pathophysiology, molecular mechanisms and protective strategies. Int. J. Mol. Sci. 2021, 22, 1864. [Google Scholar] [CrossRef]
- Katyukhin, L.N.; Kazennov, A.M.; Maslova, M.N.; Matskevich, Y.A. Rheologic properties of mammalian erythrocytes: Relationship to transport ATPases. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1998, 120, 493–498. [Google Scholar] [CrossRef]
- Plasenzotti, R.; Stoiber, B.; Posch, M.; Windberger, U. Red blood cell deformability and aggregation behaviour in different animal species. Clin. Hemorheol. Microcirc. 2004, 31, 105–111. [Google Scholar]
- Windberger, U.; Baskurt, O.K. Comparative Hemorheology. In Handbook of Hemorheology and Hemodynamics; Baskurt, O.K., Hardeman, M.R., Rampling, M.W., Meiselman, H.J., Eds.; IOS Press: Amsterdam, The Netherlands, 2007; pp. 267–285. [Google Scholar]
- Baskurt, O.K.; Meiselman, H.J. Lessons from comparative hemorheology studies. Clin. Hemorheol. Microcirc. 2010, 45, 101–108. [Google Scholar] [CrossRef]
- Nemeth, N.; Sogor, V.; Kiss, F.; Ulker, P. Interspecies diversity of erythrocyte mechanical stability at various combinations in magnitude and duration of shear stress, and osmolality. Clin. Hemorheol. Microcirc. 2016, 63, 381–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barath, B.; Somogyi, V.; Tanczos, B.; Varga, A.; Bereczky, Z.; Nemeth, N.; Deak, A. Examination of the relation between red blood cell aggregation and hematocrit in human and various experimental animals. Clin. Hemorheol. Microcirc. 2021, 78, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Matrai, A.A.; Varga, G.; Tanczos, B.; Barath, B.; Varga, A.; Horvath, L.; Bereczky, Z.; Deak, A.; Nemeth, N. In Vitro effects of temperature on red blood cell deformability and membrane stability in human and various vertebrate species. Clin. Hemorheol. Microcirc. 2021, 78, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Mohandas, N.; Clark, M.R.; Jacobs, M.S.; Shohet, S.B. Analysis of factors regulating erythrocyte deformability. J. Clin. Investig. 1980, 66, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Meiselman, H.J. Morphological determinants of red cell deformability. Scand. J. Clin. Lab. Investig. 1981, 41 (Suppl. 156), 27–34. [Google Scholar] [CrossRef] [PubMed]
- Huisjes, R.; Bogdanova, A.; van Solinge, W.W.; Schiffelers, R.M.; Kaestner, L.; van Wijk, R. Squeezing for life-properties of red blood cell deformability. Front. Physiol. 2018, 9, 656. [Google Scholar] [CrossRef]
- McMahon, T.J. Red blood cell deformability, vasoactive mediators, and adhesion. Front. Physiol. 2019, 10, 1417. [Google Scholar] [CrossRef]
- Llaudet-Planas, E.; Vives-Corrons, J.L.; Rizzuto, V.; Gómez-Ramírez, P.; Sevilla Navarro, J.; Coll Sibina, M.T.; García-Bernal, M.; Ruiz Llobet, A.; Badell, I.; Velasco-Puyó, P.; et al. Osmotic gradient ektacytometry: A valuable screening test for hereditary spherocytosis and other red blood cell membrane disorders. Int. J. Lab. Hematol. 2018, 40, 94–102. [Google Scholar] [CrossRef]
- Brun, J.F.; Varlet-Marie, E.; Myzia, J.; Raynaud de Mauverger, E.; Pretorius, E. Metabolic influences modulating erythrocyte deformability and eryptosis. Metabolites 2021, 12, 4. [Google Scholar] [CrossRef]
- Reinhart, W.H.; Piety, N.Z.; Goede, J.S.; Shevkoplyas, S.S. Effect of osmolality on erythrocyte rheology and perfusion of an artificial microvascular network. Microvasc. Res. 2015, 98, 102–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Costa, L.; Suner, L.; Galimand, J.; Bonnel, A.; Pascreau, T.; Couque, N.; Fenneteau, O.; Mohandas, N.; Society of Hematology and Pediatric Immunology (SHIP) Group; French Society of Hematology (SFH). Diagnostic tool for red blood cell membrane disorders: Assessment of a new generation ektacytometer. Blood Cells Mol. Dis. 2016, 56, 9–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demanche, R. The Osmotic Fragility of Red Blood Cells of Marine Animals: A Comparative Study. Master’s Thesis, The College of William and Mary in Virginia, Williamsburg, VA, USA, 1980. [Google Scholar]
- Oyewale, J.O. Effects of temperature and pH on osmotic fragility of erythrocytes of the domestic fowl (Gallus domesticus) and guinea-fowl (Numida meleagris). Res. Vet. Sci. 1992, 52, 1–4. [Google Scholar] [CrossRef]
- Aldrich, K.J.; Saunders, D.K.; Sievert, L.M.; Sievert, G. Comparison of erythrocyte osmotic fragility among amphibians, reptiles, birds and mammals. Trans. Kans. Acad. Sci. 2006, 109, 149–158. [Google Scholar]
- Singh, S.; Ponnappan, N.; Verma, A.; Mittal, A. Osmotic tolerance of avian erythrocytes to complete hemolysis in solute free water. Sci. Rep. 2019, 9, 7976. [Google Scholar] [CrossRef] [Green Version]
- Bessis, M.; Mohandas, N.; Feo, C. Automated ektacytometry: A new method of measuring red cell deformability and red cell indices. Blood Cells 1980, 6, 315–327. [Google Scholar]
- Hardeman, M.; Goedhart, P.; Shin, S. Methods in hemorheology. In Handbook of Hemorheology and Hemodynamics; Baskurt, O.K., Hardeman, M.R., Rampling, M.W., Meiselman, H.J., Eds.; IOS Press: Amsterdam, The Netherlands, 2007; pp. 242–266. [Google Scholar]
- Clark, M.R.; Mohandas, N.; Shohet, S.B. Osmotic gradient ektacytometry: Comprehensive characterization of red cell volume and surface maintenance. Blood 1983, 61, 899–910. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, N.; Kiss, F.; Miszti-Blasius, K. Interpretation of osmotic gradient ektacytometry (osmoscan) data: A comparative study for methodological standards. Scand. J. Clin. Lab. Investig. 2015, 75, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Son, M.; Lee, Y.S.; Lee, M.J.; Park, Y.; Bae, H.-R.; Lee, S.Y.; Shin, M.-G.; Yang, S. Effects of osmolality and solutes on the morphology of red blood cells according to three-dimensional refractive index tomography. PLoS ONE 2021, 16, e026210. [Google Scholar] [CrossRef]
- Tanczos, B.; Somogyi, V.; Bombicz, M.; Juhasz, B.; Nemeth, N.; Deak, A. Changes of hematological and hemorheological parameters in rabbits with hypercholesterolemia. Metabolites 2021, 11, 249. [Google Scholar] [CrossRef]
- Nemeth, N.; Kiss, F.; Klarik, Z.; Miko, I. Comparative osmotic gradient ektacytometry data on inter-species differences of experimental animals. Clin. Hemorheol. Microcirc. 2014, 57, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskurt, O.K.; Boynard, M.; Cokelet, G.C.; Connes, P.; Cooke, B.M.; Forconi, S.; Liao, F.; Hardeman, M.R.; Jung, F.; Meiselman, H.J.; et al. International Expert Panel for Standardization of Hemorheological Methods. New Guidelines for Hemorheological Laboratory Techniques. Clin. Hemorheol. Microcirc. 2009, 42, 75–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskurt, O.K.; Hardeman, M.R.; Uyuklu, M.; Ulker, P.; Cengiz, M.; Nemeth, N.; Shin, S.; Alexy, T.; Meiselman, H.J. Parameterization of red blood cell elongation index–shear stress curves obtained by ektacytometry. Scand. J. Clin. Lab. Investig. 2009, 69, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, M. Basic concepts and practical equations on osmolality: Biochemical approach. Clin. Biochem. 2016, 49, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Parrow, N.L.; Violet, P.C.; Tu, H.; Nichols, J.; Pittman, C.A.; Fitzhugh, C.; Fleming, R.E.; Mohandas, N.; Tisdale, J.F.; Levine, M. Measuring deformability and red cell heterogeneity in blood by ektacytometry. J. Vis. Exp. 2018, 131, e56910. [Google Scholar] [CrossRef] [PubMed]
- Zaninoni, A.; Fermo, E.; Vercellati, C.; Consonni, D.; Marcello, A.P.; Zanella, A.; Cortelezzi, A.; Barcellini, W.; Bianchi, P. Use of laser assisted optical rotational cell analyzer (LoRRca MaxSis) in the diagnosis of RBC membrane disorders, enzyme defects, and congenital dyserythropoietic anemias: A monocentric study on 202 patients. Front. Physiol. 2018, 9, 451. [Google Scholar] [CrossRef]
- Zaidi, A.U.; Buck, S.; Gadgeel, M.; Herrera-Martinez, M.; Mohan, A.; Johnson, K.; Bagla, S.; Johnson, R.M.; Ravindranath, Y. Clinical diagnosis of red cell membrane disorders: Comparison of osmotic gradient ektacytometry and eosin maleimide (EMA) fluorescence test for red cell band 3 (AE1, SLC4A1) content for clinical diagnosis. Front. Physiol. 2020, 11, 636. [Google Scholar] [CrossRef]
- Cheuvront, S.N.; Kenefick, R.W.; Heavens, K.R.; Spitz, M.G. A comparison of whole blood and plasma osmolality and osmolarity. J. Clin. Lab. Anal. 2014, 28, 368–373. [Google Scholar] [CrossRef]
- Heavens, K.R.; Kenefick, R.W.; Caruso, E.M.; Spitz, M.G.; Cheuvront, S.N. Validation of equations used to predict plasma osmolality in a healthy adult cohort. Am. J. Clin. Nutr. 2014, 100, 1252–1256. [Google Scholar] [CrossRef] [Green Version]
- Waymouth, C. Osmolality of mammalian blood and of media for culture of mammalian cells. In Vitro 1970, 6, 109–127. [Google Scholar] [CrossRef]
- Tennant, G.B.; Bailey-Wood, R.; Dallimore, C.M. Effect of medium osmolality on the growth of murine continuous marrow cultures. Cell Tissue Kinet. 1985, 18, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.D.; Warner, D.S.; Traynelis, V.C.; Todd, M.M. Plasma osmolality and brain water content in a rat glioma model. Neurosurg. 1994, 34, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Guillaumin, J.; DiBartola, S.P. Disorders of sodium and water homeostasis. Vet. Clin. N. Am. Small Anim. Pract. 2017, 47, 293–312. [Google Scholar] [CrossRef] [PubMed]
- Shpakova, N.; Orlova, N. About the mechanism of mammalian erythrocytes osmotic stability. Probl. Cryobiol. Cryomed. 2020, 30, 331–342. [Google Scholar] [CrossRef]
- Clark, M.R.; Guatelli, J.C.; Mohandas, N.; Shohet, S.B. Influence of red cell water content on the morphology of sickling. Blood 1980, 55, 823–830. [Google Scholar] [CrossRef] [Green Version]
- Benga, G. Comparative studies of water permeability of red blood cells from humans and over 30 animal species: An overview of 20 years of collaboration with Philip Kuchel. Eur. Biophys. J. 2013, 42, 33–46. [Google Scholar] [CrossRef]
- Renoux, C.; Faivre, M.; Bessaa, A.; Da Costa, L.; Joly, P.; Gauthier, A.; Connes, P. Impact of surface-area-to-volume ratio, internal viscosity and membrane viscoelasticity on red blood cell deformability measured in isotonic condition. Sci. Rep. 2019, 9, 6771. [Google Scholar] [CrossRef]
- Heo, Y.; Jung, H.; Shin, S. Osmotic deformability of erythrocytes at various shear stresses. Clin. Hemorheol. Microcirc. 2015, 59, 211–218. [Google Scholar] [CrossRef]
- Somogyi, V.; Peto, K.; Deak, A.; Tanczos, B.; Nemeth, N. Effects of aging and gender on micro-rheology of blood in 3 to 18 months old male and female Wistar (Crl:WI) rats. Biorheology 2018, 54, 127–140. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Human | Dog | Cat | Pig | Sheep | Mouse | Rat | Rabbit |
|---|---|---|---|---|---|---|---|---|
| WBC (G/L) | 6.13 ± 0.85 | 11.76 ± 2.33 * | 10.1 ± 4.34 * | 12.93 ± 3.05 * | 145.93 ± 37.13 * | 4.85 ± 0.99 * | 2.57 ± 0.5* | 5.41 ± 0.28 * |
| RBC (T/L) | 4.57 ± 0.21 | 8.16 ± 0.51 * | 11.52 ± 0.89 * | 6 ± 0.56 * | 10.41 ± 0.98 * | 8.3 ± 0.41 * | 5.95 ± 0.55 * | 6.02 ± 0.27 * |
| Hgb (g/L) | 127.69 ± 10.16 | 185.83 ± 7.73 * | 153.54 ± 14.22 * | 97.44 ± 8.61 * | 115.75 ± 12.23 * | 131.6 ± 6.55 | 108.8 ± 3.61 * | 127.5 ± 4.47 |
| Hct (%) | 42.65 ± 1.62 | 55.65 ± 2.75 * | 56.3 ± 5.89 * | 35.23 ± 3.13 * | 43.48 ± 4.19 | 44.6 ± 2.36 * | 36.48 ± 4.23 * | 40.31 ± 1.48 * |
| MCV (fL) | 93.4 ± 1.65 | 68.2 ± 1.26 * | 48.85 ± 2.9 * | 59.56 ± 3.11 * | 41.77 ± 0.9 * | 53.72 ± 0.44 * | 61.17 ± 1.96 * | 66.84 ± 1.51 * |
| MCH (pg) | 28.04 ± 2.12 | 22.78 ± 0.61 * | 13.33 ± 0.6 * | 16.36 ± 1.75 * | 11.11 ± 0.41 * | 15.87 ± 0.24 * | 18.41 ± 1.68 * | 21.15 ± 0.72 * |
| MCHC (g/L) | 309.5 ± 31.2 | 334.25 ± 4.71 | 273.31 ± 10.06 * | 238.56 ± 45.9 * | 266.31 ± 13.59 * | 295.05 ± 4.45 | 301.85 ± 31.29 | 316.25 ± 3.2 |
| Plt (G/L) | 260.38 ± 28.66 | 411.67 ± 100.43 * | 396.75 ± 38.51 | 641.88 ± 135 * | 413.5 ± 0.71 * | 880.05 ± 114.38 * | 686.45 ± 118.2 * | 481.88 ± 13.56 * |
| Variable | Human | Dog | Cat | Pig | Sheep | Mouse | Rat | Rabbit |
|---|---|---|---|---|---|---|---|---|
| ΔEI | 0.42 ± 0.02 | 0.43 ± 0.01 | 0.41 ± 0.02 | 0.39 ± 0.02 * | 0.37 ± 0.01 * | 0.47 ± 0.06 * | 0.41 ± 0.04 | 0.41 ± 0.01 |
| ΔO | 143.5 ± 4.87 | 147.33 ± 10.29 | 180.2 ± 20.59 * | 181 ± 12.01 * | 179.25 ± 12.24 * | 158.6 ± 13.78 * | 157.8 ± 13.38 * | 163 ± 6.48 * |
| ΔEI/ΔO | 0.003 ± 0.00 | 0.003 ± 0.00 | 0.002 ± 0.00 | 0.002 ± 0.00 | 0.002 ± 0.00 | 0.003 ± 0.00 | 0.003 ± 0.00 | 0.003 ± 0.00 |
| EI max/EI min (rEI) | 5.39 ± 0.67 | 4.186 ± 0.31 * | 4.96 ± 0.67 | 4.09 ± 0.41 * | 5.61 ± 0.78 | 5.11 ± 0.69 | 4.39 ± 0.74 * | 3.82 ± 0.14 * |
| O (EI max)/O min (rO) | 1.9 ± 0.08 | 2.187 ± 0.14 | 1.95 ± 0.09 | 2.01 ± 0.06 | 1.82 ± 0.07 | 1.94 ± 0.09 | 2 ± 0.07 | 2.04 ± 0.03 |
| rEI/rO | 2.84 ± 0.41 | 1.93 ± 0.25 * | 2.54 ± 0.32 | 2.03 ± 0.2 * | 3.01 ± 0.45 | 2.64 ± 0.41 | 2.19 ± 0.34 | 1.87 ± 0.07 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varga, A.; Matrai, A.A.; Barath, B.; Deak, A.; Horvath, L.; Nemeth, N. Interspecies Diversity of Osmotic Gradient Deformability of Red Blood Cells in Human and Seven Vertebrate Animal Species. Cells 2022, 11, 1351. https://doi.org/10.3390/cells11081351
Varga A, Matrai AA, Barath B, Deak A, Horvath L, Nemeth N. Interspecies Diversity of Osmotic Gradient Deformability of Red Blood Cells in Human and Seven Vertebrate Animal Species. Cells. 2022; 11(8):1351. https://doi.org/10.3390/cells11081351
Chicago/Turabian StyleVarga, Adam, Adam Attila Matrai, Barbara Barath, Adam Deak, Laszlo Horvath, and Norbert Nemeth. 2022. "Interspecies Diversity of Osmotic Gradient Deformability of Red Blood Cells in Human and Seven Vertebrate Animal Species" Cells 11, no. 8: 1351. https://doi.org/10.3390/cells11081351
APA StyleVarga, A., Matrai, A. A., Barath, B., Deak, A., Horvath, L., & Nemeth, N. (2022). Interspecies Diversity of Osmotic Gradient Deformability of Red Blood Cells in Human and Seven Vertebrate Animal Species. Cells, 11(8), 1351. https://doi.org/10.3390/cells11081351