
Megalin Orchestrates FcRn Endocytosis and Trafficking


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Transient Transfection and Crispr/ Cas9-Mediated Knockout of Megalin
2.2. CRISPR/Cas9-Mediated Knockout of Megalin
2.3. Endocytosis Assay
2.4. Immunocytochemistry
2.5. Super-Resolution Microscopy and Confocal Microscopy
2.6. Co-Immunoprecipitation (Co-IP) and Western Blot
2.7. Statistical Data Analysis
3. Results
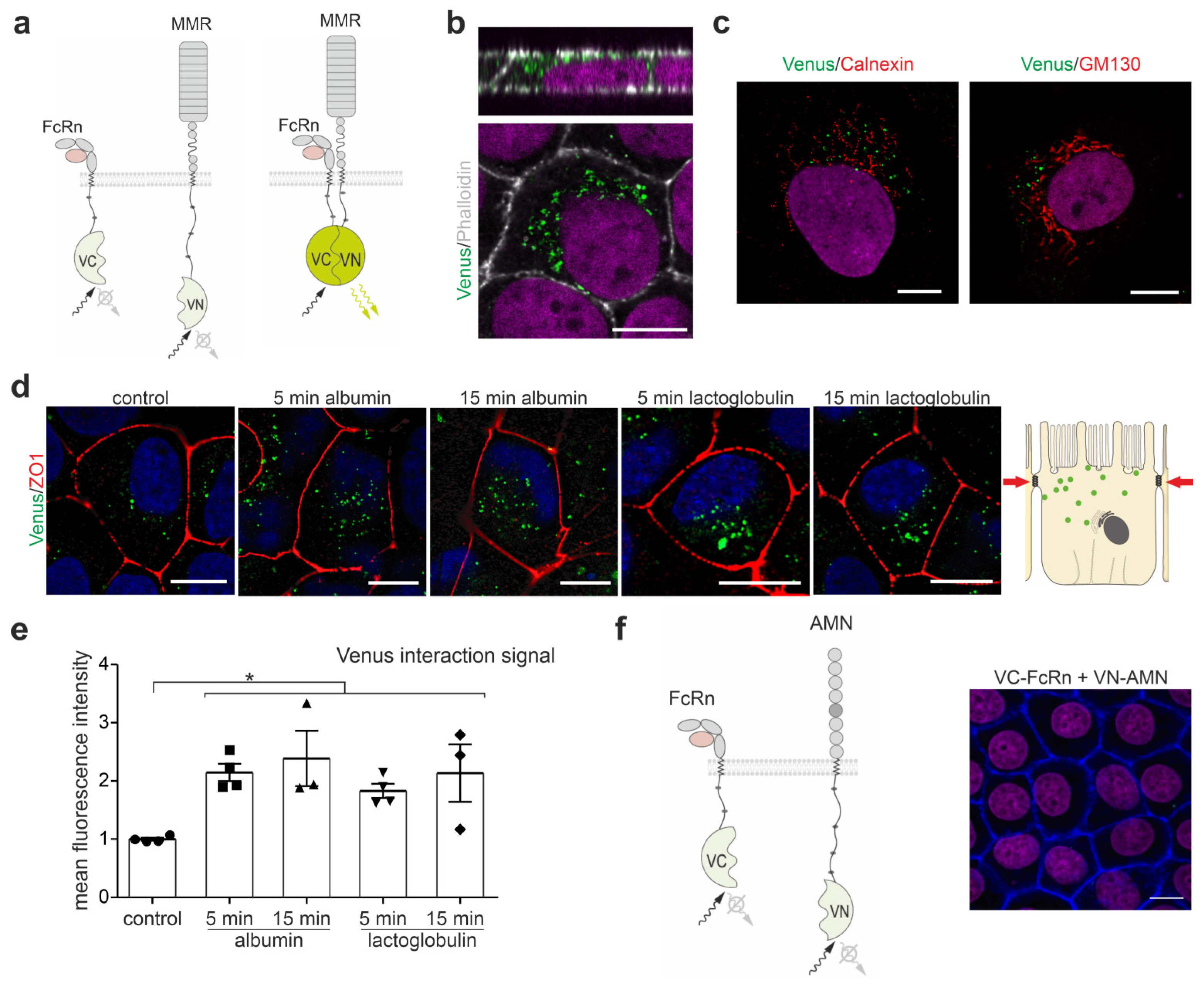
3.1. FcRn Interacts with Megalin in the Endosomal Compartment and Increases after Induction of Endocytosis
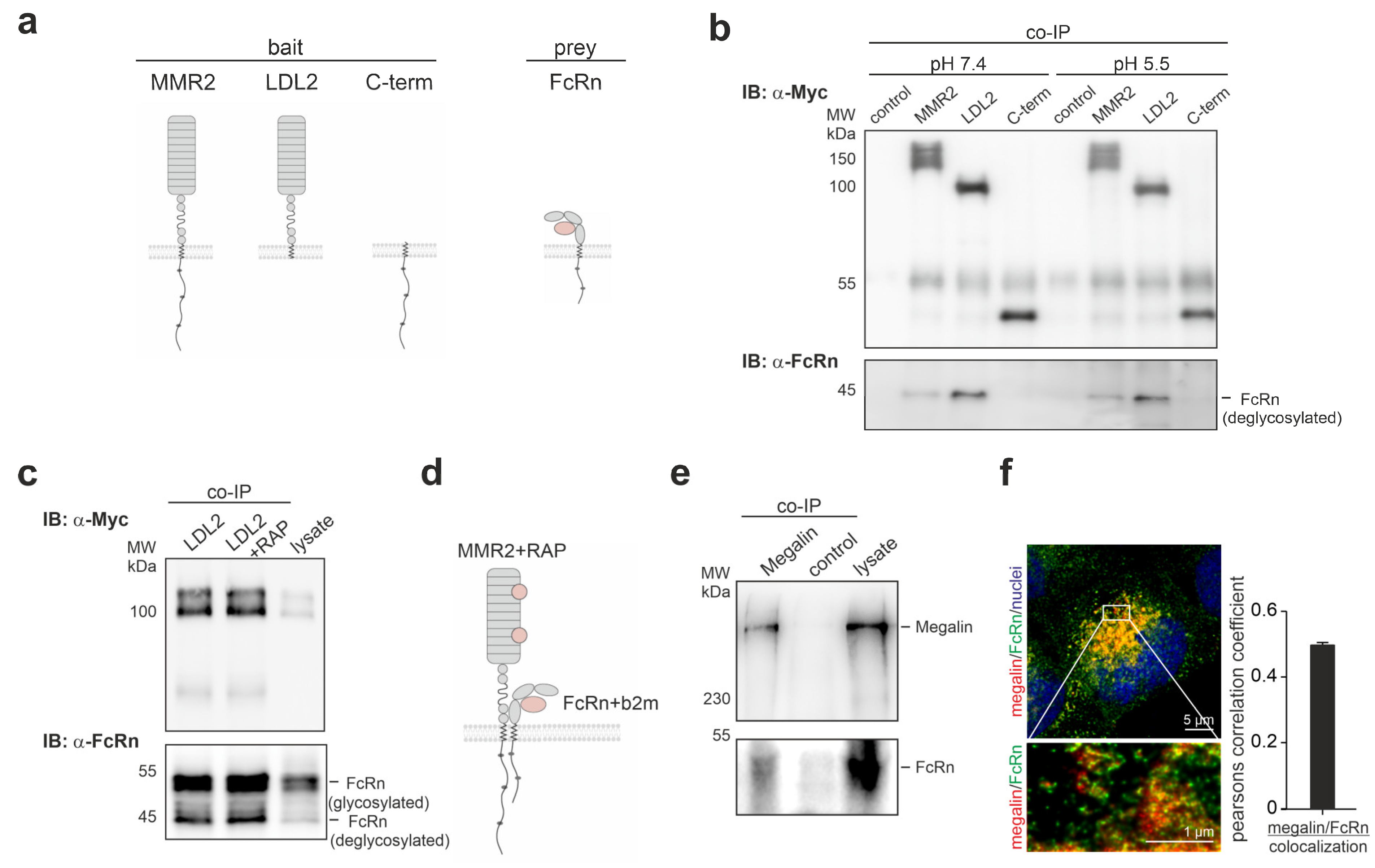
3.2. Megalin and FcRn Interact within Their Extracellular Domains
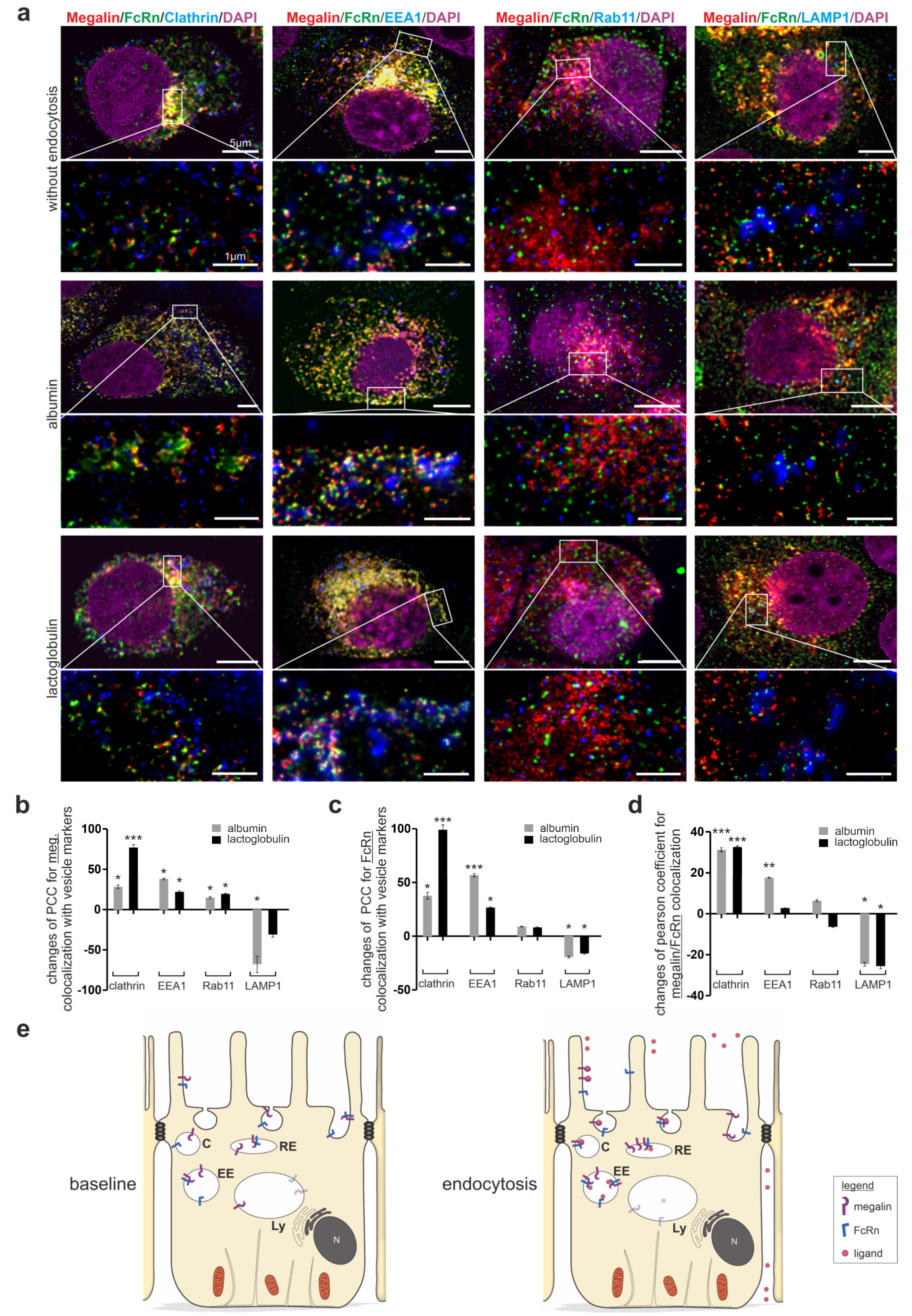
3.3. Megalin and FcRn Interact in Clathrin Vesicles upon Endocytosis Induction
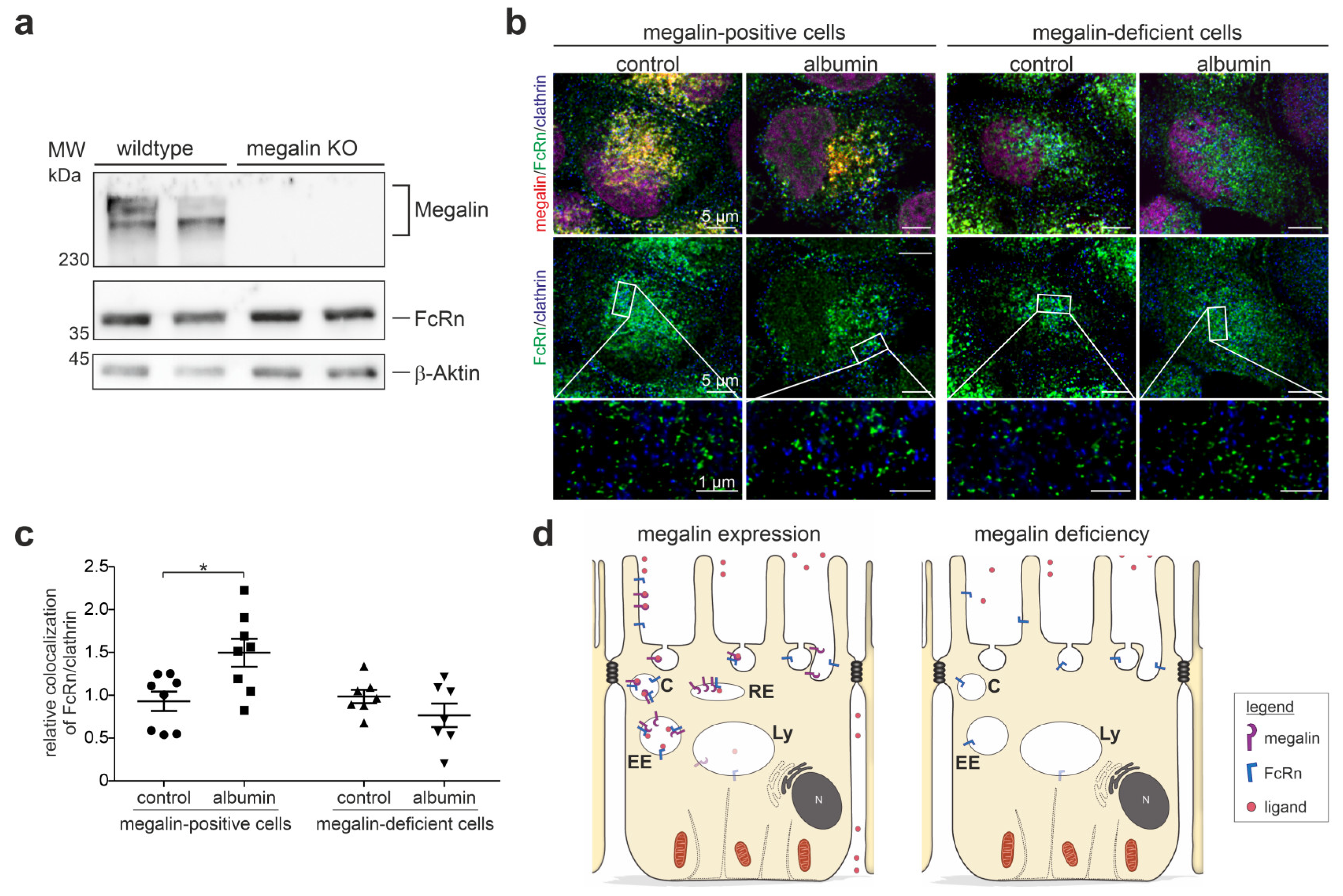
3.4. Megalin Knockout Impairs FcRn Trafficking and Function
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pyzik, M.; Sand, K.M.K.; Hubbard, J.J.; Andersen, J.T.; Sandlie, I.; Blumberg, R.S. The Neonatal Fc Receptor (FcRn): A Misnomer? Front. Immunol. 2019, 10, 1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molitoris, B.A.; Sandoval, R.M.; Yadav, S.P.S.; Wagner, M.C. Albumin uptake and processing by the proximal tubule: Physiological, pathological, and therapeutic implications. Physiol. Rev. 2022, 102, 1625–1667. [Google Scholar] [CrossRef] [PubMed]
- Sarav, M.; Wang, Y.; Hack, B.K.; Chang, A.; Jensen, M.; Bao, L.; Quigg, R.J. Renal FcRn reclaims albumin but facilitates elimination of IgG. J. Am. Soc. Nephrol. 2009, 20, 1941–1952. [Google Scholar] [CrossRef] [Green Version]
- Chaudhury, C.; Mehnaz, S.; Robinson, J.M.; Hayton, W.L.; Pearl, D.K.; Roopenian, D.C.; Anderson, C.L. The major histocompatibility complex-related Fc receptor for IgG (FcRn) binds albumin and prolongs its lifespan. J. Exp. Med. 2003, 197, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Roopenian, D.C.; Akilesh, S. FcRn: The neonatal Fc receptor comes of age. Nat. Rev. Immunol. 2007, 7, 715–725. [Google Scholar] [CrossRef]
- Jarad, G.; Miner, J.H. Update on the glomerular filtration barrier. Curr. Opin. Nephrol. Hypertens. 2009, 18, 226–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, E.I.; Birn, H.; Rippe, B.; Maunsbach, A.B. Controversies in nephrology: Renal albumin handling, facts, and artifacts! Kidney Int. 2007, 72, 1192–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, L.M.; Sandoval, R.M.; McKee, M.; Osicka, T.M.; Collins, A.B.; Brown, D.; Molitoris, B.A.; Comper, W.D. The normal kidney filters nephrotic levels of albumin retrieved by proximal tubule cells: Retrieval is disrupted in nephrotic states. Kidney Int. 2007, 71, 504–513. [Google Scholar] [CrossRef] [Green Version]
- Christensen, E.I.; Kristoffersen, I.B.; Grann, B.; Thomsen, J.S.; Andreasen, A.; Nielsen, R. A well-developed endolysosomal system reflects protein reabsorption in segment 1 and 2 of rat proximal tubules. Kidney Int. 2021, 99, 841–853. [Google Scholar] [CrossRef]
- Nielsen, R.; Christensen, E.I.; Birn, H. Megalin and cubilin in proximal tubule protein reabsorption: From experimental models to human disease. Kidney Int. 2016, 89, 58–67. [Google Scholar] [CrossRef]
- Willnow, T.E.; Christ, A. Endocytic receptor LRP2/megalin-of holoprosencephaly and renal Fanconi syndrome. Pflugers Arch. 2017, 469, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Gekle, M.; Mildenberger, S.; Freudinger, R.; Silbernagl, S. Functional characterization of albumin binding to the apical membrane of OK cells. Am. J. Physiol. 1996, 271, F286–F291. [Google Scholar] [CrossRef] [PubMed]
- Birn, H.; Christensen, E.I. Renal albumin absorption in physiology and pathology. Kidney Int. 2006, 69, 440–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenten, V.; Menzel, S.; Kunter, U.; Sicking, E.M.; van Roeyen, C.R.; Sanden, S.K.; Kaldenbach, M.; Boor, P.; Fuss, A.; Uhlig, S.; et al. Albumin is recycled from the primary urine by tubular transcytosis. J. Am. Soc. Nephrol. 2013, 24, 1966–1980. [Google Scholar] [CrossRef] [Green Version]
- Pohl, M.; Kaminski, H.; Castrop, H.; Bader, M.; Himmerkus, N.; Bleich, M.; Bachmann, S.; Theilig, F. Intrarenal renin angiotensin system revisited: Role of megalin-dependent endocytosis along the proximal nephron. J. Biol. Chem. 2010, 285, 41935–41946. [Google Scholar] [CrossRef] [Green Version]
- Theilig, F.; Kriz, W.; Jerichow, T.; Schrade, P.; Hahnel, B.; Willnow, T.; Le Hir, M.; Bachmann, S. Abrogation of protein uptake through megalin-deficient proximal tubules does not safeguard against tubulointerstitial injury. J. Am. Soc. Nephrol. 2007, 18, 1824–1834. [Google Scholar] [CrossRef] [Green Version]
- Rinschen, M.M.; Harder, J.L.; Carter-Timofte, M.E.; Zanon Rodriguez, L.; Mirabelli, C.; Demir, F.; Kurmasheva, N.; Ramakrishnan, S.K.; Kunke, M.; Tan, Y.; et al. VPS34-dependent control of apical membrane function of proximal tubule cells and nutrient recovery by the kidney. Sci. Signal. 2022, 15, eabo7940. [Google Scholar] [CrossRef]
- Zhai, X.Y.; Nielsen, R.; Birn, H.; Drumm, K.; Mildenberger, S.; Freudinger, R.; Moestrup, S.K.; Verroust, P.J.; Christensen, E.I.; Gekle, M. Cubilin- and megalin-mediated uptake of albumin in cultured proximal tubule cells of opossum kidney. Kidney Int. 2000, 58, 1523–1533. [Google Scholar] [CrossRef] [Green Version]
- Nagai, J.; Christensen, E.I.; Morris, S.M.; Willnow, T.E.; Cooper, J.A.; Nielsen, R. Mutually dependent localization of megalin and Dab2 in the renal proximal tubule. Am. J. Physiol. Ren. Physiol. 2005, 289, F569–F576. [Google Scholar] [CrossRef]
- Kim, J.; Mohanty, S.; Ganesan, L.P.; Hua, K.; Jarjoura, D.; Hayton, W.L.; Robinson, J.M.; Anderson, C.L. FcRn in the yolk sac endoderm of mouse is required for IgG transport to fetus. J. Immunol. 2009, 182, 2583–2589. [Google Scholar] [CrossRef]
- Kim, Y.; Puhl, H.L., 3rd; Chen, E.; Taumoefolau, G.H.; Nguyen, T.A.; Kliger, D.S.; Blank, P.S.; Vogel, S.S. VenusA206 Dimers Behave Coherently at Room Temperature. Biophys. J. 2019, 116, 1918–1930. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.E.; Kim, Y.; Huh, W.K.; Park, H.O. Bimolecular Fluorescence Complementation (BiFC) Analysis: Advances and Recent Applications for Genome-Wide Interaction Studies. J. Mol. Biol. 2015, 427, 2039–2055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, D.D.; Bussel, J.B. Neonatal Fc receptor in human immunity: Function and role in therapeutic intervention. J. Allergy Clin. Immunol. 2020, 146, 467–478. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, K.M.; Lam, M.; Subramanian, L.; Shakya, R.; Wu, Z.; Newton, E.E.; Simister, N.E. Effects of mutations in potential phosphorylation sites on transcytosis of FcRn. J. Cell Sci. 2001, 114, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Wernick, N.L.; Haucke, V.; Simister, N.E. Recognition of the tryptophan-based endocytosis signal in the neonatal Fc Receptor by the mu subunit of adaptor protein-2. J. Biol. Chem. 2005, 280, 7309–7316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickinson, B.L.; Claypool, S.M.; D’Angelo, J.A.; Aiken, M.L.; Venu, N.; Yen, E.H.; Wagner, J.S.; Borawski, J.A.; Pierce, A.T.; Hershberg, R.; et al. Ca2+-dependent calmodulin binding to FcRn affects immunoglobulin G transport in the transcytotic pathway. Mol. Biol. Cell 2008, 19, 414–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gburek, J.; Konopska, B.; Golab, K. Renal Handling of Albumin-From Early Findings to Current Concepts. Int. J. Mol. Sci. 2021, 22, 5809. [Google Scholar] [CrossRef]
- Elsakka, E.G.E.; Mokhtar, M.M.; Hegazy, M.; Ismail, A.; Doghish, A.S. Megalin, a multi-ligand endocytic receptor, and its participation in renal function and diseases: A review. Life Sci. 2022, 308, 120923. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dahlke, E.; Anan, Y.; Klie, L.M.; Hartkopf, A.E.; Theilig, F. Megalin Orchestrates FcRn Endocytosis and Trafficking. Cells 2023, 12, 53. https://doi.org/10.3390/cells12010053
Dahlke E, Anan Y, Klie LM, Hartkopf AE, Theilig F. Megalin Orchestrates FcRn Endocytosis and Trafficking. Cells. 2023; 12(1):53. https://doi.org/10.3390/cells12010053
Chicago/Turabian StyleDahlke, Eileen, Yaman Anan, Lea Maximiliane Klie, Ariane Elisabeth Hartkopf, and Franziska Theilig. 2023. "Megalin Orchestrates FcRn Endocytosis and Trafficking" Cells 12, no. 1: 53. https://doi.org/10.3390/cells12010053
APA StyleDahlke, E., Anan, Y., Klie, L. M., Hartkopf, A. E., & Theilig, F. (2023). Megalin Orchestrates FcRn Endocytosis and Trafficking. Cells, 12(1), 53. https://doi.org/10.3390/cells12010053