
Expression of pY397-FAK and Its miR Regulators Drive Dedifferentiation in the Thyroid Neoplasia Spectrum

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Samples and Clinical Data
2.2. Total RNA Extraction, Reverse Transcription (RT), and Quantitative PCR (qPCR)
2.3. Protein Extraction, Western Blot (WB), and Band Quantification
2.4. Immunohistochemistry and Semiquantitative Scoring Method
2.5. Statistical Analysis
3. Results
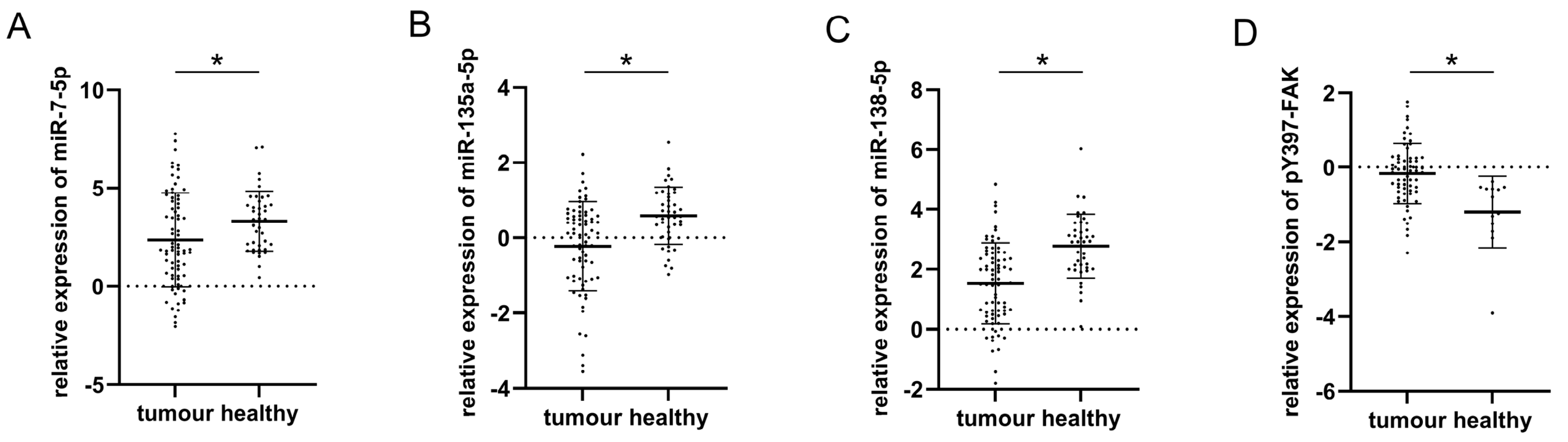
3.1. Relative Expression of miR-7-5p, miR-135a-5p, miR-138-5p, and pY397-FAK in Healthy and Malignant Thyroid Tissue
3.2. Correlation between pY397-FAK IHC Scores (Membranous, Cytoplasmic, and Nuclear); pY397-FAK Western Blot Analysis, miRs (-7-5p, -135a-5p, -138-5p) qPCR Expression, and Diagnosis Graded According to Dedifferentiation Level and Aggressiveness
3.3. Correlation between miRs (-7-5p, -135a-5p, -138-5p) Expressions and Clinicopathological Factors of PTC
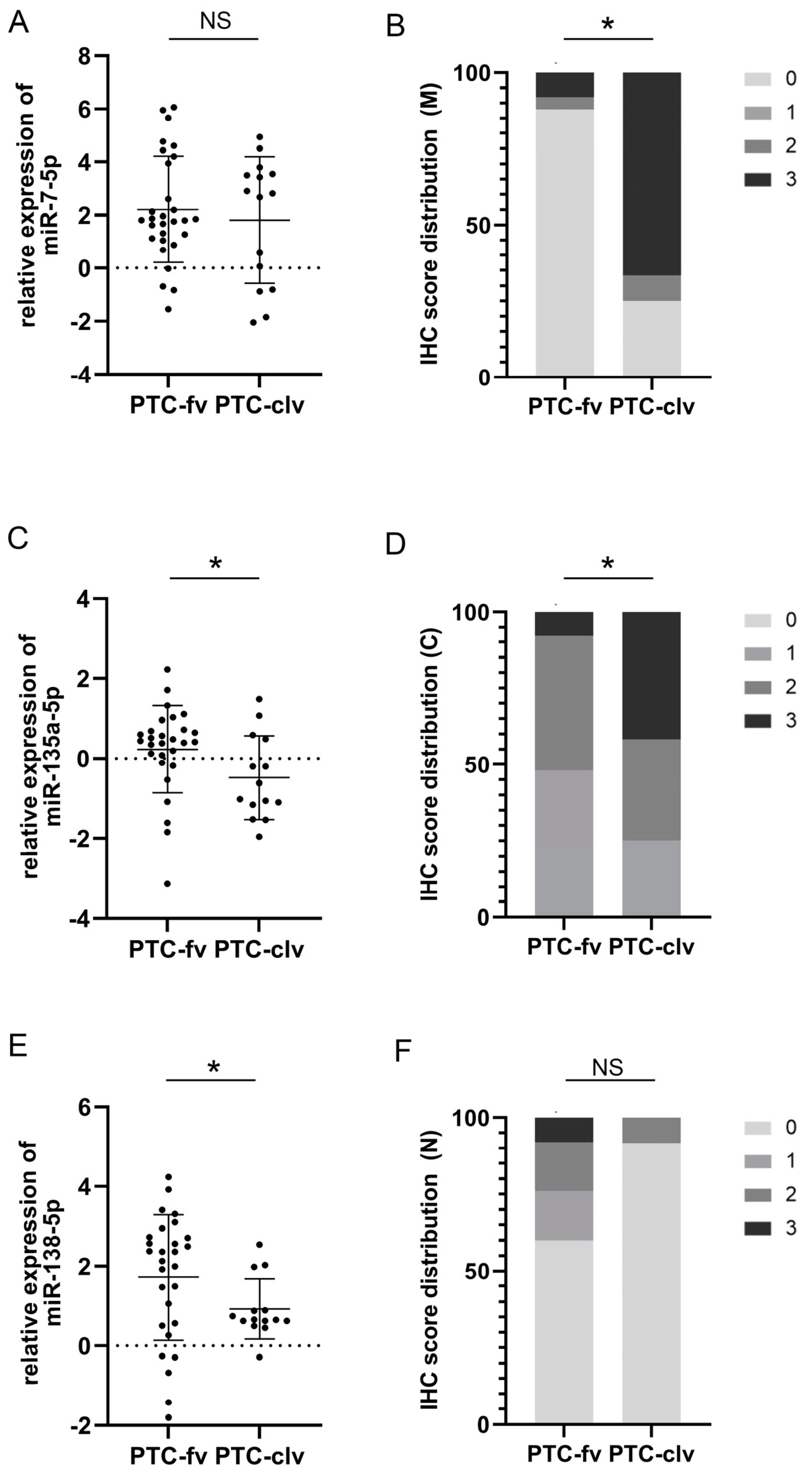
3.4. The Performances of miRs (-7-5p, -135a-5p, -138-5p) Expressions and pY397-FAK IHC Scores (Membranous, Cytoplasmic, and Nuclear) as Discriminators between Follicular and Classical Variants of PTC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cramer, J.D.; Fu, P.; Harth, K.C.; Margevicius, S.; Wilhelm, S.M. Analysis of the rising incidence of thyroid cancer using the Surveillance, Epidemiology and End Results national cancer data registry. Surgery 2010, 148, 1147–1152. [Google Scholar] [CrossRef]
- Qiu, B.; Li, H.; Chang, Q.; Yin, D. Nomograms Predict Survival in Patients with Anaplastic Thyroid Carcinoma. Med. Sci. Monit. 2019, 25, 8447–8456. [Google Scholar] [CrossRef]
- Lang, M.; Longerich, T.; Anamaterou, C. Targeted therapy with vemurafenib in BRAF(V600E)-mutated anaplastic thyroid cancer. Thyroid Res. 2023, 16, 5. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yi, Q.; Tang, L. The roles of nuclear focal adhesion kinase (FAK) on Cancer: A focused review. J. Exp. Clin. Cancer Res. 2019, 38, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provenzano, P.P.; Keely, P.J. The role of focal adhesion kinase in tumor initiation and progression. Cell Adhes. Migr. 2009, 3, 347–350. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.T.; Chen, X.L.; Lim, Y.; Hanson, D.A.; Vo, T.T.; Howerton, K.; Larocque, N.; Fisher, S.J.; Schlaepfer, D.D.; Ilic, D. Nuclear FAK promotes cell proliferation and survival through FERM-enhanced p53 degradation. Mol. Cell 2008, 29, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Golubovskaya, V.M.; Cance, W. Focal adhesion kinase and p53 signal transduction pathways in cancer. Front. Biosci. (Landmark Ed.) 2010, 15, 901–912. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Park, J.W.; Yoon, J.S.; Mok, J.O.; Kim, Y.J.; Park, H.K.; Kim, C.H.; Byun, D.W.; Lee, Y.J.; Jin, S.Y.; et al. Increased expression of focal adhesion kinase in thyroid cancer: Immunohistochemical study. J. Korean Med. Sci. 2004, 19, 710–715. [Google Scholar] [CrossRef] [Green Version]
- Owens, L.V.; Xu, L.; Dent, G.A.; Yang, X.; Sturge, G.C.; Craven, R.J.; Cance, W.G. Focal adhesion kinase as a marker of invasive potential in differentiated human thyroid cancer. Ann. Surg. Oncol. 1996, 3, 100–105. [Google Scholar] [CrossRef]
- Golubovskaya, V.M. Targeting FAK in human cancer: From finding to first clinical trials. Front. Biosci. 2014, 19, 687–706. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Guan, J.-L. Signal transduction by focal adhesion kinase in cancer. Cancer Metastasis Rev. 2009, 28, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Schlaepfer, D.D.; Mitra, S.K.; Ilic, D. Control of motile and invasive cell phenotypes by focal adhesion kinase. Biochim. Biophys. 2004, 1692, 77–102. [Google Scholar] [CrossRef] [PubMed]
- Ignjatović, V.; Janković Miljuš, J.; Rončević, J.; Tatić, S.; Išić Denčić, T.; Đorić, I.; Šelemetjev, S. Focal adhesion kinase splicing and protein activation in papillary thyroid carcinoma progression. Histochem. Cell Biol. 2022, 157, 183–194. [Google Scholar] [CrossRef]
- Shu, J.; Silva, B.V.R.E.; Gao, T.; Xu, Z.; Cui, J. Dynamic and modularized microRNA regulation and its implication in human cancers. Sci. Rep. 2017, 7, 13356. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; Li, G.; Yuan, Y.; He, Y.; Wu, X.; Zhang, W.; Wu, Z.; Chen, T.; Wu, W.; Lobie, P.E.; et al. MicroRNA-7 inhibits epithelial-to-mesenchymal transition and metastasis of breast cancer cells via targeting FAK expression. PLoS ONE 2012, 7, e41523. [Google Scholar] [CrossRef]
- Zeng, C.Y.; Zhan, Y.S.; Huang, J.; Chen, Y.X. MicroRNA 7 suppresses human colon cancer invasion and proliferation by targeting the expression of focal adhesion kinase. Mol. Med. Rep. 2016, 13, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golubovskaya, V.M.; Sumbler, B.; Ho, B.; Yemma, M.; Cance, W.G. MiR-138 and MiR-135 directly target focal adhesion kinase, inhibit cell invasion, and increase sensitivity to chemotherapy in cancer cells. Anticancer Agents Med. Chem. 2014, 14, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Yeh, M.; Oh, C.S.; Yoo, J.Y.; Kaur, B.; Lee, T.J. Pivotal role of microRNA-138 in human cancers. Am. J. Cancer Res. 2019, 9, 1118–1126. [Google Scholar]
- Augenlicht, A.; Saiselet, M.; Decaussin-Petrucci, M.; Andry, G.; Dumont, J.E.; Maenhaut, C. MiR-7-5p inhibits thyroid cell proliferation by targeting the EGFR/MAPK and IRS2/PI3K signaling pathways. Oncotarget 2021, 12, 1587–1599. [Google Scholar] [CrossRef]
- Chen, N.; Yin, D.; Lun, B.; Guo, X. LncRNA GAS8-AS1 suppresses papillary thyroid carcinoma cell growth through the miR-135b-5p/CCND2 axis. Biosci. Rep. 2019, 39, BSR20181440. [Google Scholar] [CrossRef] [Green Version]
- Mitomo, S.; Maesawa, C.; Ogasawara, S.; Iwaya, T.; Shibazaki, M.; Yashima-Abo, A.; Kotani, K.; Oikawa, H.; Sakurai, E.; Izutsu, N.; et al. Downregulation of miR-138 is associated with overexpression of human telomerase reverse transcriptase protein in human anaplastic thyroid carcinoma cell lines. Cancer Sci. 2008, 99, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Kadkhoda, S.; Eslami, S.; Mahmud Hussen, B.; Ghafouri-Fard, S. A review on the importance of miRNA-135 in human diseases. Front. Genet. 2022, 13, 973585. [Google Scholar] [CrossRef] [PubMed]
- DeLellis, R.A.; Lloyd, R.V.; Heitz, P.U.; Eng, C.; Eng, C.; Machado De Avila, R.A. Pathology and Genetics of Tumours of Endocrine Organs. In World Health Organization Classification of Tumours; IARC Press: Lyon, France, 2004. [Google Scholar]
- Edge, S.B.; Byrd, D.R.; Compton, C.C.; Fritz, A.G.; Greene, F.L.; Trotti, A. AJCC Cancer Staging Manual, 7th ed.; Springer: New York, NY, USA, 2010; pp. 87–96. [Google Scholar]
- Basolo, F.; Torregrossa, L.; Giannini, R.; Miccoli, M.; Lupi, C.; Sensi, E.; Berti, P.; Elisei, R.; Vitti, P.; Baggiani, A.; et al. Correlation between BRAF V600E mutation and tumour invasiveness in papillary thyroid carcinomas smaller than 20 milimeteres: Analysis of 1060 cases. J. Clin. Endocrinol. Metab. 2010, 95, 4197–4205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šelemetjev, S.; Bartolome, A.; Išić Denčić, T.; Đorić, I.; Paunović, I.; Tatić, S.; Cvejić, D. Overexpression of epidermal growth factor receptor and its downstream effector, focal adhesion kinase, correlates with papillary thyroid carcinoma progression. Int. J. Exp. Pathol. 2018, 99, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Andisha, N.M.; McMillan, D.C.; Gujam, F.J.A.; Roseweir, A.; Edwards, J. The relationship between phosphorylation status of focal adhesion kinases, molecular subtypes, tumour microenvironment and survival in patients with primary operable ductal breast cancer. Cell Signal. 2019, 60, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Guan, J.L. Focal adhesion kinase and it’s signaling pathways in cell migration and angiogenesis. Adv. Drug Deliv. Rev. 2011, 63, 610–615. [Google Scholar] [CrossRef] [Green Version]
- Cary, L.A.; Guan, J.L. Focal adhesion kinase in integrin-mediated signaling. Front. Biosci. 1999, 4, D102–D113. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, R.V.; Buehler, D.; Khanafshar, E. Papillary thyroid carcinoma variants. Head Neck Pathol. 2011, 5, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Tong, J.; Ruan, M.; Jin, Y.; Fu, H.; Cheng, L.; Luo, Q.; Liu, Z.; Lv, Z.; Chen, L. Poorly differentiated thyroid carcinoma: A clinician’s perspective. Eur. Thyroid J. 2022, 11, e220021. [Google Scholar] [CrossRef]
- Smallridge, R.C.; Marlow, L.A.; Copland, J.A. Anaplastic thyroid cancer: Molecular pathogenesis and emerging therapies. Endocr. Relat. Cancer 2009, 16, 17–44. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, M.; Gaweł, D.; Godlewska, M. Novel Inhibitor-Based Therapies for Thyroid Cancer—An Update. Int. J. Mol. Sci. 2021, 22, 11829. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L. Ninety-four cases of encapsulated follicular variant of papillary thyroid carcinoma: A name change to Noninvasive Follicular Thyroid Neoplasm with Papillary-like Nuclear Features would help prevent overtreatment. Mod. Pathol. 2016, 29, 698–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henke, L.E.; Pfeifer, J.D.; Baranski, T.J.; DeWees, T.; Grigsby, P.W. Long-term outcomes of follicular variant vs classic papillary thyroid carcinoma. Endocr. Connect. 2018, 7, 1226–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Correlation Parameters | IHC(M) pY397-FAK | IHC(C) pY397-FAK | IHC(N) pY397-FAK | qPCR miR-7-5p | qPCR miR-135a-5p | qPCR miR-138-5p | WB pY397-FAK | Histotype Agg. | |
|---|---|---|---|---|---|---|---|---|---|
| IHC(M) pY397-FAK | r | 0.755 | −0.374 | −0.031 | −0.108 | −0.155 | 0.495 | 0.367 | |
| p | 0.000 a | 0.002 a | 0.810 a | 0.416 a | 0.237 a | 0.000 a | 0.002 a | ||
| IHC(C) pY397-FAK | r | 0.755 | −0.214 | 0.052 | −0.033 | 0.001 | 0.470 | 0.258 | |
| p | 0.000 a | 0.080 a | 0.690 a | 0.804 a | 0.993 a | 0.000 a | 0.034 a | ||
| IHC(N) pY397-FAK | r | 0.374 | −0.214 | 0.030 | 0.037 | 0.046 | −0.112 | −0.180 | |
| p | 0.002 a | 0.080 a | 0.817 a | 0.782 a | 0.730 a | 0.415 a | 0.142 a | ||
| qPCR miR-7-5p | r | −0.031 | 0.052 | 0.030 | 0.317 | 0.597 | −0.100 | 0.042 | |
| p | 0.810 a | 0.690 a | 0.817 a | 0.008 b | 0.000 b | 0.460 b | 0.723 a | ||
| qPCR miR-135a-5p | r | −0.108 | −0.033 | 0.037 | 0.317 | 0.451 | −0.097 | −0.308 | |
| p | 0.416 a | 0.804 a | 0.782 a | 0.008 b | 0.000 b | 0.480 b | 0.008 a | ||
| qPCR miR-138-5p | r | −0.155 | 0.001 | 0.046 | 0.597 | 0.451 | −0.116 | −0.097 | |
| p | 0.237 a | 0.993 a | 0.730 a | 0.000 b | 0.000 b | 0.394 b | 0.416 a | ||
| WB pY397-FAK | r | 0.495 | 0.470 | −0.112 | −0.100 | −0.097 | −0.116 | 0.255 | |
| p | 0.000 a | 0.000 a | 0.415 a | 0.460 b | 0.480 b | 0.394 b | 0.043 a | ||
| Histotype agg. | r | 0.367 | 0.258 | −0.180 | 0.042 | −0.308 | −0.097 | 0.255 | |
| p | 0.002 a | 0.034 a | 0.142 a | 0.723 a | 0.008 a | 0.416 a | 0.043 a |
| N (75) | miR-7-5p | p | miR-135a-5p | p | miR-138-5p | p | ||
|---|---|---|---|---|---|---|---|---|
| Age in Years (Range) | 14–89 | r = −0.078 | 0.505 | r = −0.122 | 0.618 a | r = 0.059 | 0.618 a | |
| Gender | Male | 16 | 3.385 ± 2.13 | 0.059 b | −0.103 ± 1.333 | 0.68 b | 1.867 ± 1.11 | 0.287 b |
| Female | 60 | 2.117 ± 2.403 | −0.25 ± 1.16 | 1.446 ± 1.406 | ||||
| Tumor size (mm) | range | 2–120 | r = −0.130 | 0.265 a | r = −0.067 | 0.578 a | r = −0.025 | 0.831 a |
| Intraglandular dissemination | Absent | 41 | 2.711 ± 2.456 | 0.2 b | −0.2 ± 1.275 | 0.864 b | 1.426 ± 1.608 | 0.48 b |
| Present | 34 | 1.998 ± 2.287 | −0.248 ± 1.092 | 1.653 ± 0.997 | ||||
| Lymph node metastasis | Absent | 62 | 2.334 ± 2.327 | 0.678 b | −0.148 ± 1.108 | 0.221 b | 1.551 ± 1.363 | 0.78 b |
| Present | 13 | 2.64 ± 2.768 | −0.627 ± 1.56 | 1.431 ± 1.363 | ||||
| Degree of tumor infiltration | 1/2 | 40 | 2.604 ± 2.236 | 0.3 b | −0.021 ± 1.079 | 0.5 b | 1.744 ± 1.501 | 0.134 b |
| 3/4 | 32 | 2.001 ± 2.587 | −0.577 ± 1.246 | 1.255 ± 1.116 | ||||
| Extra-thyroid invasion | Absent | 54 | 2.41 ± 2.464 | 0.902 b | −0.211 ± 1.224 | 0.905 b | 1.54 ± 1.422 | 0.933 b |
| Present | 21 | 2.332 ± 2.253 | −0.249 ± 1.115 | 1.51 ± 1.187 | ||||
| pT | Low (1/2) | 54 | 2.409 ± 2.408 | 0.92 b | −0.224 ± 1.19 | 0.981 b | 1.601 ± 1.461 | 0.579 b |
| High (3/4) | 21 | 2.351 ± 2.408 | −0.217 ± 1.204 | 1.419 ± 1.178 | ||||
| pTNM | Low (1/2) | 52 | 2.422 ± 2.317 | 0.648 b | −0.143 ± 1.168 | 0.323 b | 1.566 ± 1.385 | 0.525 b |
| High (3/4) | 22 | 2.144 ± 2.539 | −0.452 ± 1.244 | 1.347 ± 1.224 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ignjatović Jocić, V.; Janković Miljuš, J.; Išić Denčić, T.; Živaljević, V.; Tatić, S.; Đorić, I.; Šelemetjev, S. Expression of pY397-FAK and Its miR Regulators Drive Dedifferentiation in the Thyroid Neoplasia Spectrum. Cells 2023, 12, 1721. https://doi.org/10.3390/cells12131721
Ignjatović Jocić V, Janković Miljuš J, Išić Denčić T, Živaljević V, Tatić S, Đorić I, Šelemetjev S. Expression of pY397-FAK and Its miR Regulators Drive Dedifferentiation in the Thyroid Neoplasia Spectrum. Cells. 2023; 12(13):1721. https://doi.org/10.3390/cells12131721
Chicago/Turabian StyleIgnjatović Jocić, Valentina, Jelena Janković Miljuš, Tijana Išić Denčić, Vladan Živaljević, Svetislav Tatić, Ilona Đorić, and Sonja Šelemetjev. 2023. "Expression of pY397-FAK and Its miR Regulators Drive Dedifferentiation in the Thyroid Neoplasia Spectrum" Cells 12, no. 13: 1721. https://doi.org/10.3390/cells12131721
APA StyleIgnjatović Jocić, V., Janković Miljuš, J., Išić Denčić, T., Živaljević, V., Tatić, S., Đorić, I., & Šelemetjev, S. (2023). Expression of pY397-FAK and Its miR Regulators Drive Dedifferentiation in the Thyroid Neoplasia Spectrum. Cells, 12(13), 1721. https://doi.org/10.3390/cells12131721