1. Introduction
Multiple sclerosis (MS) is a chronic and degenerative disease of the central nervous system (CNS) with a 30% prevalence [
1]. This neuroinflammatory disease is characterized by peripheral immune cell infiltration into the CNS, oligodendrocyte loss, the formation of demyelinating lesions, astrocyte and microglial activation, that jointly cause neuronal damage [
1,
2,
3]. Neuroimmune interactions between glial, neuronal, and immune cells play important roles in MS pathology. In MS, the infiltration of autoreactive immune cells promotes and sustains neuroinflammation, a dynamic response of glial cells to the disruption of the CNS integrity. An important part of this process is the bidirectional interaction between immune cells and glial cell astrocytes. This interaction contributes to the further infiltration of the immune cells into the CNS parenchyma and to the shaping of the astrocytic inflammatory response [
4,
5,
6,
7,
8,
9,
10,
11,
12]. In addition to the communication via the release of inflammatory cues, astrocytes directly interact with the CNS-infiltrated immune cells (CNS-IICs) on a rapid time scale, indicating that such cell–cell interaction could also be a component of the neuroinflammatory response in MS [
13,
14,
15]. However, much less is known about this direct interaction, even though this abundant type of glial cells should continuously encounter CNS-IICs in their proximity over the course of the disease. Notably, the most effective therapies for MS treatment target integrins [
16], i.e., the ubiquitous transmembrane receptors that mediate cell-to-cell interaction [
17], but the integrin engagement in astroglial direct interactions with CNS-IICs in MS has not been examined.
Astrocytes create an extensive meshwork throughout the entire CNS parenchyma. These glial cells express connexins [
18] that can form gap junctions which interconnect neighboring astrocytes or connexin hemichannels which provide passage for ions and small molecules such as Ca
2+ ions, ATP, and glucose to the extracellular space [
19,
20,
21]. Astrocytes also express a myriad of transmitter receptors and ion channels that enable communication with neighboring cells and allow them to perform maintenance functions in the CNS [
22]. Receptor activation is followed by the transient increase in intracellular Ca
2+ concentration in astrocytes [
22]. Intracellular Ca
2+ increase is caused by the entry of extracellular Ca
2+, and/or its release from the intracellular stores such as endoplasmic reticulum and mitochondria [
23]. In addition, the Ca
2+ transporting machinery of mitochondria provides an efficient route for the regulation of cytosolic Ca
2+ concentration [
24,
25]. Previously, we showed that astrocytes release ATP through connexin hemichannels upon encountering CNS-IICs, ATP then activates the P2X7 receptor (P2X7R) and triggers a Ca
2+ increase in these glial cells [
14]. However, the characteristics and the context of the astroglial P2X7R engagement that occurs in close proximity to the CNS-IICs have not been completely defined, limiting our understanding on the direct interaction between these cells in MS.
To determine the mechanisms responsible for establishing the cell–cell interactions between astrocytes and CNS-IICs, we used the experimental autoimmune encephalomyelitis (EAE) model of MS, and we combined Western immunoblotting, immunohistochemistry, immunoprecipitation, ATP release measurement, and Ca2+ imaging. We show that the astrocyte response to the nearby CNS-IICs involves the coupling of astrocytic P2X7R, connexin-43 (Cx-43), and β3-integrin; in addition, this response is dependent on the mitochondrial activity. These data point toward the astrocyte cell–cell interactions with infiltrating immune cells as a target for the treatment of neuroinflammation in MS.
2. Materials and Methods
2.1. Experimental Animals
The Dark Agouti (DA) rat strain was used in all experiments. Animals were housed under standard laboratory conditions in a 12 h light/dark cycle at 22–24 °C. Food and water were provided ad libitum. EAE was induced in the 2- to 3-month-old rats of both sexes, as was previously described in [
14]. Briefly, spinal cord homogenate (SCH) in phosphate buffer saline (PBS; 50%
w/
v) was mixed with an equal volume of complete Freund’s adjuvant (CFA; Sigma-Aldrich, Darmstadt, Germany), which was supplemented with 5 mg/mL of
Mycobacterium tuberculosis (BD Medical, Franklin Lakes, NJ, USA). Animals were intradermally injected in a hind paw pad with 100 µL of SCH and CFA emulsion. Animals were monitored daily, and the disease score was estimated based on the following scale: 0—no clinical signs, 1—flaccid tail, 2—hind limb paresis, 3—severe hind limb paralysis, and 4—limp tail, complete hind leg, and partial foreleg paralysis. Animals were sacrificed when reaching a 2–4 EAE disease score (
Supplementary Figure S1) together with the age-matched healthy control rats. DA rat pups (1- to 3-day-old) were used for the preparation of the spinal cord astrocyte culture. Animal procedures were carried out in accordance with the protocols of the Ethics committee for the use of Laboratory animals of the Republic of Serbia, which is in compliance with the EU directive (2010/63/EU) on the protection of animals that are used for scientific purposes.
2.2. Protein Isolation
Spinal cords from EAE rats (N = 8,
Supplementary Figure S1) and age-matched controls (N = 8) were isolated. Lumbar spinal cords were homogenized in a cold lysis buffer containing (mM) the following: 50 Tris, 150 NaCl, 10 EDTA, 10 EGTA, 0.1% SDS, and 0.5% Triton X-100. These were supplemented with protease and phosphatase inhibitors (ThermoFisher Scientific, Waltham, MA, USA), and were sonicated for 30 s. Homogenates were incubated in the lysis buffer for an additional 15 min on the ice and centrifuged at 14,000 rpm for 20 min at +4 °C. The supernatants were collected and protein concentration was determined using the Pierce Assay Kit (ThermoFisher Scientific, USA). A Laemmli sample buffer containing 1% β-mercaptoethanol was added to the samples before denaturation at 95 °C for 5 min. Samples were kept at −80 °C until needed for further use, i.e., Western blot.
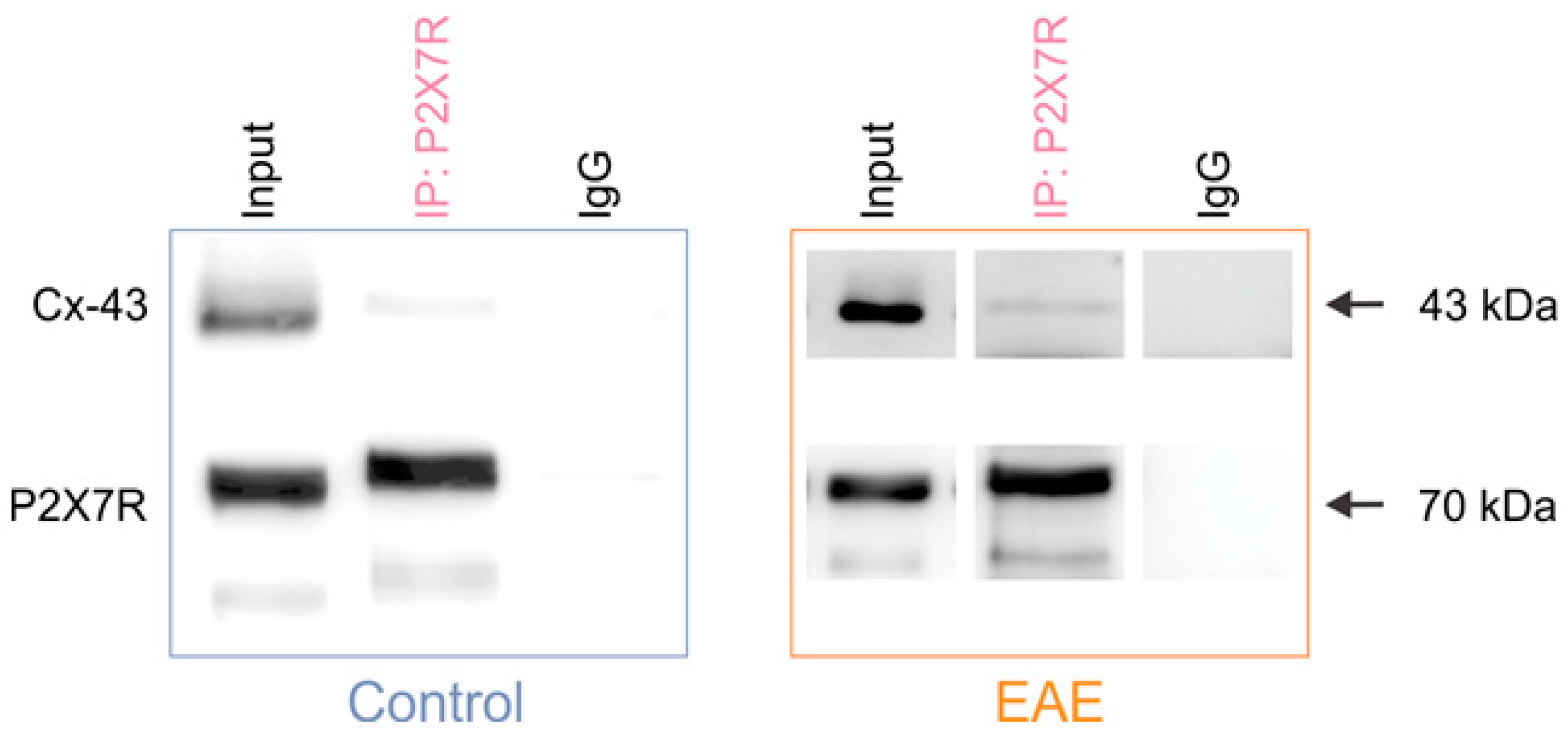
2.3. Coimmunoprecipitation
Lumbar spinal cords were isolated from the 2- to 3-month-old DA control rats (N = 3) and rats with EAE (N = 1) (
Supplementary Figure S1) homogenized in an ice-cold immunoprecipitation (IP) lysis buffer composed of: 10 mM HEPES, 200 mM NaCl, 30 mM EDTA, 0.5% Triton X-100, protease and phosphatase inhibitors, and had pH 7.4. Next, the samples were clarified by centrifugation at 14,000 rpm for 20 min at +4 °C, and protein concentration was determined with a Pierce Assay Kit (ThermoFisher Scientific, USA). Samples were incubated with magnetically labeled Dynabeads
TM (ThermoFisher Scientific, USA) for 1.5 h, and the precleared supernatants were then used as the input for the immunoprecipitation. A total of 15 µL of magnetic beads per sample tube were incubated with a blocking solution containing 5% bovine serum albumin (BSA; Merck, Darmstadt, Germany) in an IP buffer for 1 h and 15 min on a rotator. Next, the beads were incubated with 4 µg of the rabbit anti-P2X7R primary antibody (Alomone Labs, Jerusalem, Israel) or 1 µg of the control rabbit IgG (Millipore, Burlington, MA, USA) in an IP buffer at +4 °C for 2 h on a rotator. After several washes with an IP buffer, 300 µg of the precleared input sample was added to the beads bound to the antibody and incubated at +4 °C on the rotator overnight. Bound proteins were eluted using a Laemmli sample buffer during a 15 min incubation at room temperature (RT). After the addition of 1% β-mercapthoethanol, the samples were denaturated at 95 °C for 5 min and kept at −80 °C until needed for further use, i.e., Western blot.
2.4. Immunoblot
The total proteins isolated from the lumbar spinal cords of the EAE (
Supplementary Figure S1) and control rats, the precleared input and IP samples, or the cultured astrocytes were used for Western immunoblotting. Equal amounts of proteins (20 µg for tissue preparations and 10 µg for culture preparations) were separated by electrophoresis on 8% or 10% polyacrylamide gel. Next, the proteins were transferred to PVDF membranes (Immobilon, Merck, Germany) via a semi-dry transfer using the Trans-Blot Turbo Transfer System (Bio-Rad Laboratories, Hercules, CA, USA). PVDF membranes were next saturated in a 5% fat-free dried milk (SERVA, Heidelberg, Germany) in a Tris buffer saline with 0.1% Tween20 (TBST) (Fisher Bioreagents, Pittsburgh, PA, USA). These were then incubated overnight at +4 °C with the following primary antibodies in a 5% milk in TBST: rabbit anti-P2X7R (1:1000; Alomone Labs, Israel); rabbit β3-integrin (1:1000; Synaptic Systems, Göttingen, Germany); guinea pig anti-Cx-43 (1:1000; Alomone Labs, Israel); and rabbit anti-GAPDH (1:2000; DAKO Agilent, Carpinteria, CA, USA). The next day, the membranes were washed 3 times for 10 min in TBST and then incubated for 2 h at RT with a following horse radish peroxidase conjugated secondary antibodies: goat anti-rabbit (1:5000; Abcam, Cambridge, UK) and goat anti-guinea pig (1:5000; Santa Cruz Biotechnology, Dallas, TX, USA). After several washes, the membranes were revealed using a chemiluminescence detection kit (Western-ECL substrate, Bio-Rad Systems, Hercules, CA, USA), and were visualized using a ChemiDoc-It imager (UVP Systems, Upland, CA, USA). The experiments were performed in duplicate, and the analysis was conducted using the Gel Analyzer in Fiji ImageJ Software (NIH, Bethesda, MD, USA).
2.5. Spinal Cord Astrocyte Culture Preparation
1-to-3-days old DA rat pups, were used for cell culture preparation. The primary astrocyte cultures were prepared as was previously described in [
14]. Briefly, the animals were decapitated, and their spinal cords were isolated. The meninges were carefully removed and the spinal cord was dissociated in PBS. The tissue was mechanically dissociated, and the homogenate was centrifuged at 500×
g for 5 min at RT. The supernatant was discarded and the pellet was resuspended in a full Dulbecco’s modified Eagle medium (DMEM, Sigma Aldrich, Germany), which contained 10% fetal bovine serum (FBS, Gibco, Billings, MT, USA), 100 µM of sodium-pyruvate (Sigma Aldrich, Germany), and 1% penicillin-streptomycin (ThermoFisher Scientific, USA). The centrifugation–resuspension cycle was repeated, and the cell suspension was plated in a Petri dish. The cells were maintained in a humidified atmosphere of 5% CO
2/95% air at 37 °C. The culture media was changed every 2–3 days until reaching confluence. The cells were washed with PBS, detached by trypsinization (0.025% trypsin and 0.02% EDTA in PBS) for 2 min, transferred to a tube with full DMEM, and then centrifuged at 500×
g for 5 min at the RT. The supernatant was discarded, the pellet was resuspended in a full DMEM, and the cells were plated in a Petri dish at a density of 10
4 cells/cm
2. After reaching confluence, the cells were trypsinized, centrifuged at 500×
g for 5 min at the RT, resuspended in a full DMEM, and seeded on poly-L-lysine (PLL)-coated circular glass coverslips at a density of 10
4 cells/cm
2. Cells were used within the following 3 days for experiments.
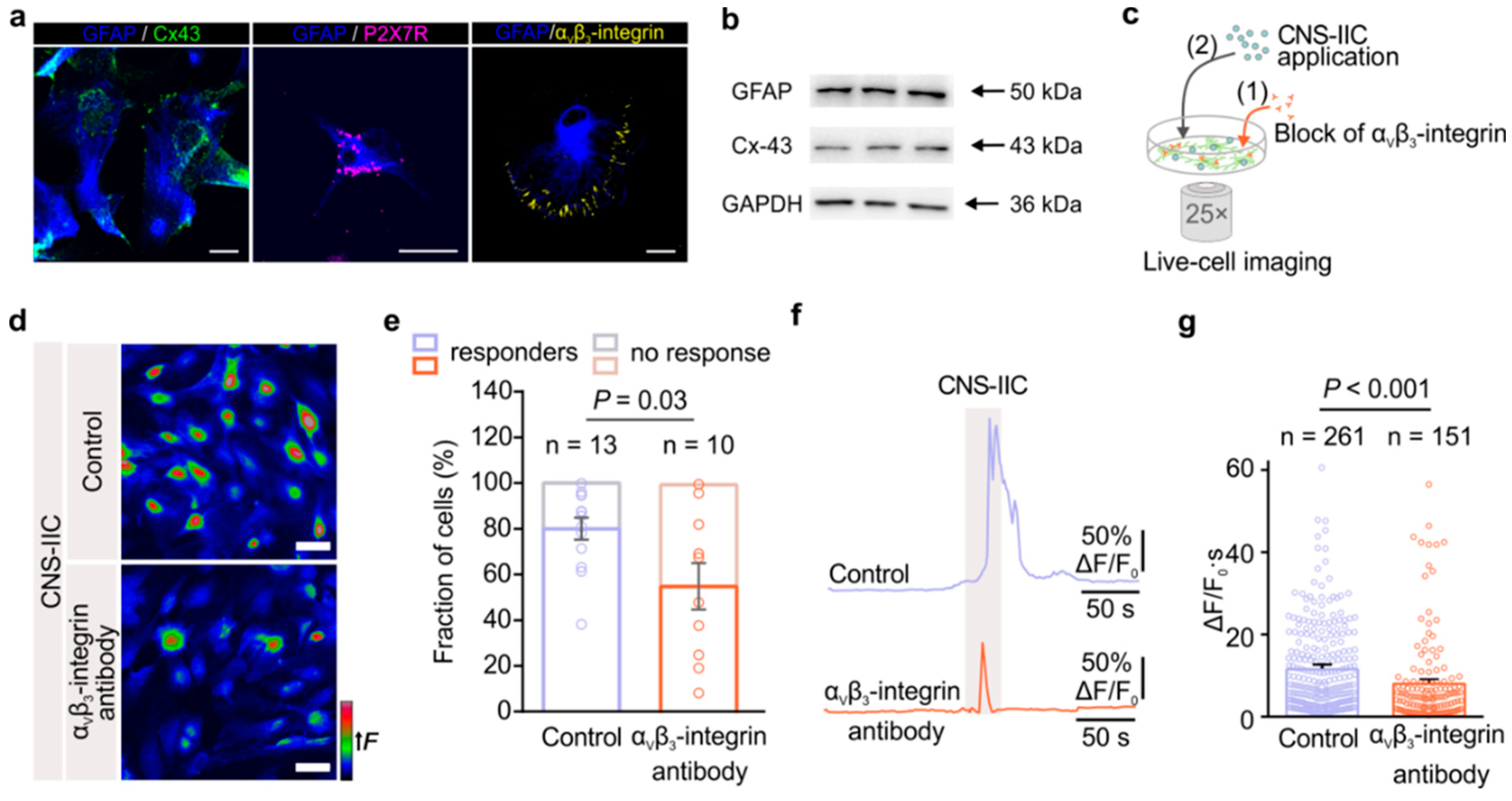
2.6. Immunofluorescence
The control rats and rats with EAE (
Supplementary Figure S1) were anesthetized by an intraperitoneal injection of ketamine (55 mg/kg) and xylazine (5 mg/kg), and were transcardially perfused with 0.9% physiological saline followed by 4% paraformaldehyde (PFA). The isolated spinal cords were postfixed in a 4% PFA for an additional 24 h and then dehydrated in rising sucrose concentrations of 10%, 20%, and 30% in 0.2 M phosphate buffer (PB). The tissues were frozen and cut at a 25 µm thickness using a cryostat (Leica Biosystems, Deer Park, IL, USA), and were then kept at –20 °C until needed for further use. The lumbar spinal cord sections were rehydrated in PBS for 10 min, and were then incubated for 1 h in a blocking solution containing the following: 10% normal donkey serum (NDS, Sigma-Aldrich, Germany), 2% BSA (Merck, Germany), and 0.1% Triton-X in PBS. The sections were incubated with primary antibodies in a PBS with 2% NDS at 4 °C overnight. The following primary antibodies were used: guinea pig anti-Cx-43 (1:200; Alomone Labs, Israel); mouse anti-CD4 (1:100; Abcam, UK); rabbit anti-glutamate transporter-1 (GLT-1; 1:100, Frontier Institute, Sendai, Japan); mouse anti-glutamine synthetase (GS; 1:100, BD Biosciences, Franklin Lakes, NJ, USA); and rabbit anti-P2X7R (1:200; Alomone Labs, Israel). Next, the sections were washed in PBS 3 times for 5 min and incubated with secondary antibodies in PBS with a 2% NDS at RT for 2 h. The following secondary antibodies were used: donkey anti-guinea pig Alexa 488 or Alexa 647; donkey anti-mouse Alexa 555; goat anti-mouse Alexa 488; goat anti-rabbit Alexa 555; and donkey anti-rabbit Alexa 647 (1:200 for all secondary antibodies used; Invitrogen, Waltham, MA, USA). The sections were washed in PBS 3 times for 5 min and mounted using a MOWIOL medium (Sigma-Aldrich, Germany).
For labeling the astrocytes in culture, the cells were plated on a circular glass coverslips at a density of 104 cells/cm2. Then, they were fixed 48 h later in a 4% PFA for 30 min at RT. The cells were rinsed in PBS 3 times for 5 min, as well as blocked in a 10% NGS, 2% BSA, and 0.05% Triton-X in PBS for 45 min; they were then incubated with primary antibodies that were diluted in PBS overnight at 4 °C. The following primary antibodies were used: guinea pig anti-Cx-43 (1:200; Alomone Labs, Israel); rabbit anti-P2X7R (1:200; Alomone Labs, Israel); mouse anti-αvβ3-integrin (1:100; Abcam, UK); and chicken anti-GFAP (1:500; Abcam, UK). The cells were rinsed 3 times for 10 min and then incubated for 2 h at RT with the following secondary antibodies diluted in PBS: donkey anti-guinea pig Alexa 488; goat anti-rabbit Alexa 555; goat anti-mouse AlexaFluor 555 (1:200; Invitrogen, USA); and goat anti-chicken AlexaFluor 647 (1:200; Invitrogen, USA). The cells were mounted with a MOWIOL medium after rinsing 3 times in PBS for 10 min.
2.7. Confocal Image Acquisition and Analysis
Images of the immunolabeled frozen spinal cord sections were acquired on a confocal laser-scanning microscope (LSM 510, Carl Zeiss GmbH, Jena, Germany), which was equipped with Ar multi-line (488 nm) and HeNe (543 and 633 nm) lasers. The 63× (N.A. 1.4) oil immersion objective was used to acquire images with a pixel size of 143 nm in a 1 µm z-step interval. Assessment of Cx-43 and P2X7R expression and colocalization was done on 3–4 slices of the ventral lumbar spinal cord per animal. For each slice, 4 images of gray and 4 images of white matter were acquired and analyzed. Image acquisition parameters were kept constant for the control and EAE samples.
Image analysis was performed using Fiji ImageJ software (NIH, USA). The signal intensity of P2X7R and Cx-43 was measured as the integrated density. The colocalization of the Cx-43/P2X7R, Cx-43/GLT-1, and Cx-43/GS signals was assessed via Pearson’s correlation coefficient (PCC) by using the Just Another Colocalization Plugin (JACoP) [
26]. The PCC was calculated from a 7–10 µm z-stack, which represented the average of the PCC values that were determined for each z-plane. The P2X7R and Cx-43 signal intensities and the distribution of their colocalization were further analyzed in manually set regions of interest (ROI): the one that covers the proximity of the infiltrated CD4
+ T cells (tProximity of the CD4
+ T cell), and the one that covers a random area without immune cells (Random). The analysis was performed within a 3–20 µm radius from the center of the established ROIs. The positions of the random ROIs were established within a 50 µm distance from the center of Proximity to CD4
+ T cell’s ROIs. The distribution of the Cx-43 and P2X7R signal intensities in the established ROIs in the maximum z-projection of a 7 µm z-stack were obtained with the Radial Profile plugin. For the Sholl analysis of the Cx-43/P2X7R signal colocalizations, PCC measurements were performed in the established ROIs in a 0.5 µm radius increment of the circles. The analysis of the established ROIs excluded the signals from the labeled CD4
+ T cells that were identified as small circles of a 5 µm radius. This enabled a specific assessment of the astrocytic P2X7R and Cx-43 signal intensities and their colocalization in the immediate proximity of the CD4
+ T cell boundary. The number of colocalized Cx43/P2X7R signals intersecting a circle at each step size was divided with perimeter of the corresponding circle; we named this value the “density”. This analysis was also performed in a healthy animal (the control ROIs). The density of the P2X7R/Cx-43 colocalization was calculated as an average of the 7 µm z-stack, where each z-plane was analyzed separately.
2.8. Immune Cell Isolation
CNS-IICs were isolated form the EAE spinal cords (
Supplementary Figure S1) as was previously described in [
27]. Briefly, the animals with EAE were anesthetized by an intraperitoneal injection of ketamine (55 mg/kg) and xylazine (5 mg/kg), and were transcardially perfused with an ice cold PBS (50–100 mL per animal). The spinal cords were isolated and mechanically dissociated in an ice-cold PBS that was supplemented with a 3% FBS (Gibco, ThermoFisher Scientific, USA). Next, the homogenate was passed through a 40 µm cell strainer to obtain the single cell suspension, which was then centrifuged at 100×
g for 10 min at 4 °C. The supernatant was discarded and the pellet was resuspended in 3 mL of 30% isotonic Percoll (Sigma Aldrich, Germany). Next, it was slowly placed over 3 mL of 70% isotonic Percoll and centrifuged at 850×
g for 40 min at RT. The cell layer containing CNS-IICs that were located between the 30% and 70% Percoll layers was carefully extracted and transferred to a tube with RPMI (Gibco, ThermoFisher Scientific, USA), which was supplemented with 5% FBS, centrifuged for 5 min at 500×
g at RT, and then resuspended in an RPMI with 5% FBS. The centrifugation–resuspension step was repeated twice. The isolated CNS-IICs were resuspended in an RPMI with 5% FBS and kept at 4 °C. The CNS-IICs were centrifuged at 500×
g for 5 min at RT, counted with a hemocytometer, and then resuspended in ECS at concentrations of 50 × or 25 × 10
3 cells/mL prior to application.
2.9. ATP Release Measurement
The ATP release from astrocytes was measured using, as described in the manufacturer’s instructions, a luciferin-luciferase assay ATPLite kit (PerkinElmer, Waltham, MA, USA). The astrocytes were seeded on the bottom of the 96-well plate at a 104 cells/cm2 density. Three experimental conditions were established: the control astrocytes in ECS, astrocytes treated with a 50 × 103 CNS-IIC/well, and astrocytes that were preincubated with 20 µM CGP37157 (Tocris Bioscience, Bristol, UK) and treated with a 50 × 103 CNS-IIC/well. Bioluminescence was measured using a Chameleon plate reader (Cole-Parmer, Vernon Hills, IL, USA).
2.10. Time-Lapse Fluorescence Imaging
In order to monitor the Ca2+ dynamics in the cultured astrocytes, we used cell-permeable Fluo4-AM. The cultured astrocytes previously seeded on coverslips were loaded with 5 µM Fluo4-AM (Invitrogen, USA) for 30 min at RT in ECS. ECS contained the following (in mM): 140 NaCl, 5 KCl, 2 CaCl2, 2 MgCl2, 10 D-glucose (all from Sigma-Aldrich, Germany), and 10 HEPES (Biowest, Nuaillé, France), and had pH 7.4 and 300 mOsm osmolality. After washing 2 times for 10 min, the coverslip was placed in the recording chamber. Time-lapse imaging was carried out by using an AxioObserver A1 microscope with an LD LCI Plan-Apochromat 25× (N.A. 0.8) water immersion objective lens (Carl Zeiss), an “evolve”-EM 512 Digital Camera System (Photometrics, Tucson, AZ, USA), and VisiView® high-performance software (Visi-Chrome, Visitron Systems GmbH, Puchheim, Germany). The Fluo4-AM was excited at 480 nm with a Xenon Short Arc lamp (Ushio, Tokyo, Japan), which was coupled to the VisiChrome Polychromatic Illumination System (Visitron Systems). The excitation and emission lights were passed through a FITC filter set (Chroma Technology Inc., Bellows Falls, VT, USA), and the frame scanning was performed at a frequency of 1 Hz. The cell perfusion system consisted of a three-barrel pipette connected to a Valve-Controlled Gravity Perfusion System (ALA Scientific Instruments Inc, Farmingdale, NY, USA). Each of the barrels was made of glass capillaries with an 800 µm inner diameter. The three-barrel pipette was attached to a micromanipulator to allow for the precise positioning of the pipette tip in the close vicinity of the astrocytes—specifically at the edge of the field of view, 1 mm above the coverslip surface, and at an angle of 45° relative to the bottom of the recording chamber. The accurate exchange between bathing the ECS, CNS-IICs, and ATP barrel solutions was performed using a valve-controlled system. The perfusion through the barrels was held constant at the flow rate of 4 mL/min, thus allowing for the exclusive perfusion of astrocytes via a solution from a single barrel. After 100 s of baseline recording, the bath application of ECS was switched to a solution containing CNS-IICs. The immune cells were applied for 20–30 s and then the perfusion was stopped for additional 70–80 s to allow for the establishment of a direct interaction between the CNS-IICs and astrocytes. The ECS was re-applied to a bath for 100 s to remove the CNS-IICs. Finally, the bathing solution was switched for 5 s to the one containing ATP. The CNS-IICs were applied at a concentrations of 25 or 50 × 103 cells/mL. ATP was applied at 200 µM concentration (Sigma-Aldrich, Germany). ATP was applied to confirm the viability of the astrocytes.
To block the mitochondrial Na+/Ca2+ (NCLX) and H+/Ca2+ (HCX) exchangers, the astrocytes were incubated with 20 µM CGP-37157 (7-chloro-5-(2-chlorophenyl)-1,5-dihydro-4,1-benzothiazepin-2(3H)-one; Tocris Bioscience, UK). To block the astrocytic αvβ3-integrin, the astrocytes were preincubated for 2 h with the primary mouse anti-αvβ3-integrin antibody (1 µg/mL; Abcam, UK). The data in the presence of the drug or the antibody were compared with randomly interleaved control data that was obtained without the drug or the preincubation period.
Analysis was performed using Fiji ImageJ software. The ROIs of the individual astrocytes were outlined based on the Ca
2+ response evoked by ATP. The fluorescent signal intensity (F) was measured in each ROI and normalized to the baseline fluorescence (F
0), which was conducted according to the formula of ΔF/F
0 = (F − F
0)/F
0, to express the Ca
2+ change. The astrocytes were considered responders when the CNS-IICs-evoked Ca
2+ change was greater than 3 × SD of the baseline signal for at least 5 s. The Ca
2+ signals detected by Fluo4-AM were quantified by measuring the area under the trace of the fluorescence change during the period of CNS-IIC presence in the proximity of the astrocytes (100 s). We integrated the consecutive ΔF/F
0 signals as follows: 100 s before and 100 s from the start of CNS-IICs application. The resulting values are expressed as ΔF/F
0·s in all graphs. The rate of cytoplasmic Ca
2+ efflux was obtained by measuring the slope of the linear fit of the decay phase of the evoked Ca
2+ response [
28,
29]. The area, peak amplitude, rise time, decay time, and decay slope of the Ca
2+ change were determined in Clampfit version 10 (version 10.6.2.2. for Windows, Molecular Devices, San Jose, CA, USA).
2.11. Statistical Analysis
The data were analyzed and plotted using GraphPad Prism Software version 6 (version 6.01. for Windows, GraphPad Software, La Jolla, CA, USA) and SigmaPlot Software version 12 (version 12.2.0.45. for Windows, Systat Sofrware, Palo Alto, CA, USA). To assess the difference between the two groups when the data sets showed a normal distribution, a two-tailed unpaired Student’s t-test was used, while a non-parametric Mann–Whitney rank sum test was applied when the normality test failed. Multiple groups were compared using a one-way ANOVA analysis on the ranks (Kruskal–Wallis test), which was followed by the Student Newman–Keuls post hoc test. A two-way ANOVA analysis on the ranks was used for multiple comparisons between the group data sets. Statistical tests were used to measure the significance, along with the corresponding significance levels (p value) and sample size (these are shown in the figures and figure legends). The p values were considered significant if they were less than 0.05. All data are presented as the mean ± SEM. The dots represent individual measurements. N indicates the number of animals examined. For ATP measurement, n denotes the number of cultures from different animals. For the analysis of the time-lapse imaging data, n denotes the number of astrocytes that were obtained from at least three cultures from different animals.
4. Discussion
A mesh-like distribution of astrocytes provides coverage of almost the entire CNS and positions them to establish multiple interactions with the CNS resident cells. These interactions occur in a healthy brain and involve the astrocytic crosstalk with neurons, microglia, oligodendrocytes, other astrocytes, and blood vessels, thus allowing astrocytes to fulfill metabolic and homeostatic maintenance functions [
39,
40,
41]. In neuroinflammatory conditions, however, these interactions change and may affect the pathological processes that depend on the stimuli present in the inflamed milieu [
42,
43]. Moreover, as demonstrated in our previous study, astrocytes also establish direct cell-to-cell interactions with the CNS non-resident autoreactive immune cells that infiltrate into the CNS parenchyma [
14]; extending previous reports showing that astrocyte communication with the components of the immune system is mediated by inflammatory signals. In great part, all of these cell-to-cell interactions occur through astroglial ATP-based purinergic signaling [
44,
45]. The signaling mediated by purines operates on a second-based time scale, enabling fast communications between astrocytes and other cells in their proximity. In a healthy brain astroglial ATP acts as a gliotransmitter, while in pathological conditions it is considered to be a danger signal. In both cases, ATP activates purinergic receptors that induce the downstream Ca
2+ signals in astrocytes. Astroglial purinergic signaling is a hallmark of many CNS diseases, including MS [
32,
46]. This highlights the importance of understanding the context in which purinergic receptors are engaged and to define the mechanisms that control their signaling. By using an EAE rat model, we discovered that P2X7R interacts with astroglial Cx-43 and concentrates in the immediate proximity of CNS-IICs. In addition, we found that P2X7R interacts with β
3-integrin and that P2X7R-dependent direct astrocyte–autoreactive immune cell interactions are dependent on astroglial mitochondrial activity and α
vβ
3-integrins.
Purinergic P2X7Rs contribute to the pathological events of CNS diseases such as epilepsy, Alzheimer’s disease, and MS [
43,
47,
48]. These receptors have been connected with the profound long-lasting changes in astrocyte properties, which—in these diseases—typically induce reactive astroglial remodeling (e.g., astrogliosis) [
45]. In the EAE animal model of MS, a blockade of P2X7Rs by the administration of an antagonist Brilliant blue G decreases astrogliosis in the rat forebrain and alleviates the neurological symptoms of the disease [
30]. Another study using P2X7R null mice showed that P2X7R deficiency reduces EAE incidence, as well as reduces astroglial activation and axonal damage, but does not influence immune cell infiltration into the CNS [
31]. MS is considered to be a white matter disease; however, lesions are also prominent in the gray matter of the brain. Accordingly, previous research demonstrated that astrocytes near the chronic active and inactive brain lesions express P2X7Rs in both the white and gray matter of the frontal cortex of patients with MS [
32]. The expression of P2X7Rs was also demonstrated in the hypertrophic astrocytes surrounding the perivascular infiltrates in the brain of MS patients [
46], suggesting that this purinergic receptor type is a critical component of astrocyte interaction with CNS immune cell infiltrates. By performing studies in the spinal cord of the rats with EAE, we discovered that astroglial P2X7Rs concentrate in the immediate proximity of the infiltrated CD4
+ T cells in gray matter. We did not detect a prominent expression of P2X7Rs in astrocytes, neither in the control nor in EAE rat spinal cord tissue. This is in agreement with previous research, where it has been shown that the P2X7Rs in the CNS are localized on astrocytes at a low density [
49]. In addition, aside from the analysis performed in the proximity of the CNS-infiltrated CD4
+ T cells, Cx-43 and P2X7R showed a decrease in the colocalization in EAE. Together, these results suggest that P2X7R expression is increased and persists in the astrocytes that are in the immediate proximity of the CD4
+ T cell in EAE, and that this receptor can contribute to the cell–cell interactions between astrocytes and autoreactive immune cells in the inflamed CNS.
P2X7R expression has been studied in MS and EAE. Augmented expression has been observed in the frontal cortex of patients with secondary progressive MS [
32]. In the present study, at the peak of EAE, we observed a decrease in the P2X7R expression in the spinal cord for both females and males, while it was previously shown that the expression of this receptor increases in the forebrain [
30]. Alterations in the P2X7R expression support the premise that these purinergic receptors are involved in the pathological processes of MS, however, the activity and mechanisms that control their activation remain largely unknown. Following the activation of P2X7Rs in astrocytes, many signaling pathways are activated that stimulate inflammation and also release of gliotransmitters that can modulate neuronal activity [
45]. In our previous studies, we revealed that astroglial P2X7Rs are activated upon the direct astrocytic interaction with CNS-IICs [
14], which could provide an explanation for the persisting expression of these purinergic receptors in the astrocytes that are in the immediate proximity of the CNS immune cell infiltrates. In the present study, we further reveal that mitochondrial activity contributes to this cell–cell interaction. The block of mNCLX and HCX mitochondrial transporters augmented the ATP release from astrocytes, and markedly increased the CNS-IICs-evoked astrocytic Ca
2+ response by slowing the efflux of cytosolic Ca
2+. As the activity of mNCLX is linked to oxidative phosphorylation [
50], our data provide an indication of the contribution of this transporter to ATP production by astrocytes when they encounter CNS-IICs. Direct assessment, however, would help to define how mitochondrial activity is modulated during the astrocyte interaction with immune cells. In addition to producing ATP, mitochondria serve as Ca
2+ storage organelles that efficiently regulate cytosolic Ca
2+ [
25]. This has been observed in a variety of cells including astrocytes, cardiomyocytes, and pancreatic β cells [
24,
51], suggesting that the mitochondrial Ca
2+ regulation function is critically important for cellular metabolism. In acute brain slices and in vivo, mitochondria display spontaneous Ca
2+ activity [
52,
53], which is thought to reflect an increase in metabolic demand and cellular stress [
54]. Thus, the frequency of mitochondria-regulated Ca
2+ transients in astrocytes increases in the animal model of amyotrophic lateral sclerosis in which the mutant enzyme superoxide dismutase is overexpressed [
53]. This suggests that astrocyte “cell-intrinsic” mitochondria-regulated Ca
2+ signals are perturbed in the CNS disease. Although further studies are necessary to assess the parallel mitochondrial and cytosolic Ca
2+ levels in astrocytes during their interaction with CNS-IICs, our results here indicate that mitochondrial activity facilitates the termination of the CNS-IIC-evoked astroglial Ca
2+ signals. The effects we describe appear to be mediated primarily within astrocytes since we excluded the influence of other CNS-resident cells by studying the mitochondrial regulation of the astrocyte Ca
2+ response in pure astrocyte cultures. Notably, we observed that ATP application also evokes a larger astroglial Ca
2+ response after blocking the mNCLX and HCX-dependent mitochondrial Ca
2+ transports, suggesting that the regulation of astrocytic cytosolic Ca
2+ levels by mitochondria is modified during different pathological conditions in which the level of this purine is elevated. Together, our previous [
14] and herein described results suggest that mitochondria are involved in the regulation of the P2X7R activity in astrocytes. The frequent CNS-IICs-evoked P2X7R-dependent Ca
2+ signals within astrocytes may contribute to the release of gliotransmitters and to the activation of inflammatory pathways in MS. Indeed, the block of P2X7Rs (see above) results in the reduction in astrogliosis and in the improvement of EAE symptoms.
Essential aspects of the intracellular Ca
2+ signals described in cultured astrocytes have been substantiated in acute brain slices, as well as in vivo [
55,
56,
57,
58]. However, the data obtained on cultured control astrocytes in this study require further confirmation through the use of brain slices and in vivo approach. This includes using animals with EAE in which mitochondria are specifically labeled, and/or the P2X7Rs are specifically removed from the astrocytes to assess the contribution of the astrocytic Ca
2+ signaling in this disease. Notably, we also consider that the astrocyte–immune cell interaction revealed in our studies may be shaped by the inputs from other cells in the neuroinflammatory CNS environment, predominantly that derived by microglia. Indeed, on one side, microglia engage in cross-talk with CNS-infiltrated T lymphocytes and modulate immune cell function in the CNS [
59,
60]; while, on the other side, microglia can also promote neurotoxic astroglial activity in MS [
61]. Furthermore, microglia show dynamic changes over the course of EAE, and CD4 expression in these glial cells has been correlated with the improvement of clinical symptoms for this disease [
62]. Consequently, the net effect of microglial signals on the astrocyte–immune cell interaction in EAE is difficult to predict and requires detailed studies involving manipulation of different types of individual glial cell activities in different stages of disease.
Astrocytes release ATP, which then activates the purinergic receptors through different mechanisms—including the Cx-43 hemichannels [
63]. This has been shown for astrocytic P2Y1Rs in Alzheimer’s disease [
64], and for astroglial P2X7Rs in our previous research on astrocyte interaction with CNS-IICs that were isolated from EAE rats [
14]. Now, we reveal that Cx-43 interacts with the P2X7Rs in EAE, which provides further evidence that purine release through the hemichannels could directly activate this purinergic receptor in astrocytes. Our data also demonstrate that the association of these proteins is important for the cell–cell interactions between astrocytes and the CNS-infiltrated immune cells in EAE. Indeed, we found that Cx-43 colocalization with P2X7Rs is higher in the immediate proximity of CD4
+ T cells. Moreover, we demonstrate that P2X7R also interacts with the β
3-integrin in EAE, and that the block of astrocytic α
vβ
3-integrin reduces their direct interaction with the CNS-IICs isolated from EAE rats. Previous studies showed that the functional coupling between α
vβ
3-integrin, Cx-43, and P2X7Rs regulates astrocyte migration [
38], and our data shown here further demonstrate that this coupling constitutes a signaling cascade of the events of the cell–cell interactions between astrocytes and the immune cells that invade the CNS in EAE. Moreover, our data point toward the role of mitochondria in the regulation of this coupling in astrocytes. Additional studies, however, will help elucidate the link between astroglial mitochondrial activity and α
vβ
3- integrin in EAE. Integrins are important mediators of cell-to-cell interactions, contributing to the inflammatory processes in MS [
65], and our data suggest that they could be involved in establishing a contact between astrocytes and CNS-IICs. Indeed, many integrins are upregulated in the CNS resident cells and immune cells in MS [
65], and we observed an increase in β
3-integrin expression in the spinal cord of rats with EAE. Our data on the interaction of P2X7Rs with α
vβ
3-integrin and Cx-43, and on the expression of astrocytic P2X7Rs at the proximity to the infiltrated CD4
+ T cells, suggest that, at a certain level, this increase in integrin expression occurs in astrocytes at the peak of EAE. Notably, integrin targeting is applied in the treatment of many pathological conditions including MS [
16]. In MS, the α
4β
7 and α
4β
1-integrin blockers that are currently applied in therapeutic treatment target immune cells and control their infiltration into the CNS. Yet, immune cell-mediated mechanisms alone cannot explain the neuroinflammation and neuronal degeneration in MS without the CNS-resident cells component. Our findings thus open possibility to target astrocytes in development of innovative non-immune cell-based therapeutic strategies for treatment of MS. Such a strategy could be effective in saving neurons from damage as virtually every aspect of brain function involves a neuron–astroglial partnership.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





