
Inversely Regulated Inflammation-Related Processes Mediate Anxiety–Obesity Links in Zebrafish Larvae and Adults

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Larvae and Adult Fish
2.3. Larval Obesity Model
2.4. Caffeine-Induced Anxiety
2.5. Larval Behavior Tests
2.6. Adult Fish Behavior Tests
2.7. Larval Whole-Body Cortisol Assay
2.8. Adipocyte Assessment Assay
2.9. Caffeine Pretreatments
2.10. RNA Extraction
2.11. RNA-Seq
2.12. cDNA Synthesis
2.13. qRT-PCR
2.14. Datasets
2.14.1. Brains of Anxious vs. Less Anxious Adult Zebrafish
2.14.2. Telencephalons of HFD-Fed vs. SD-Fed Adult Zebrafish
2.15. Statistical Analysis
3. Results
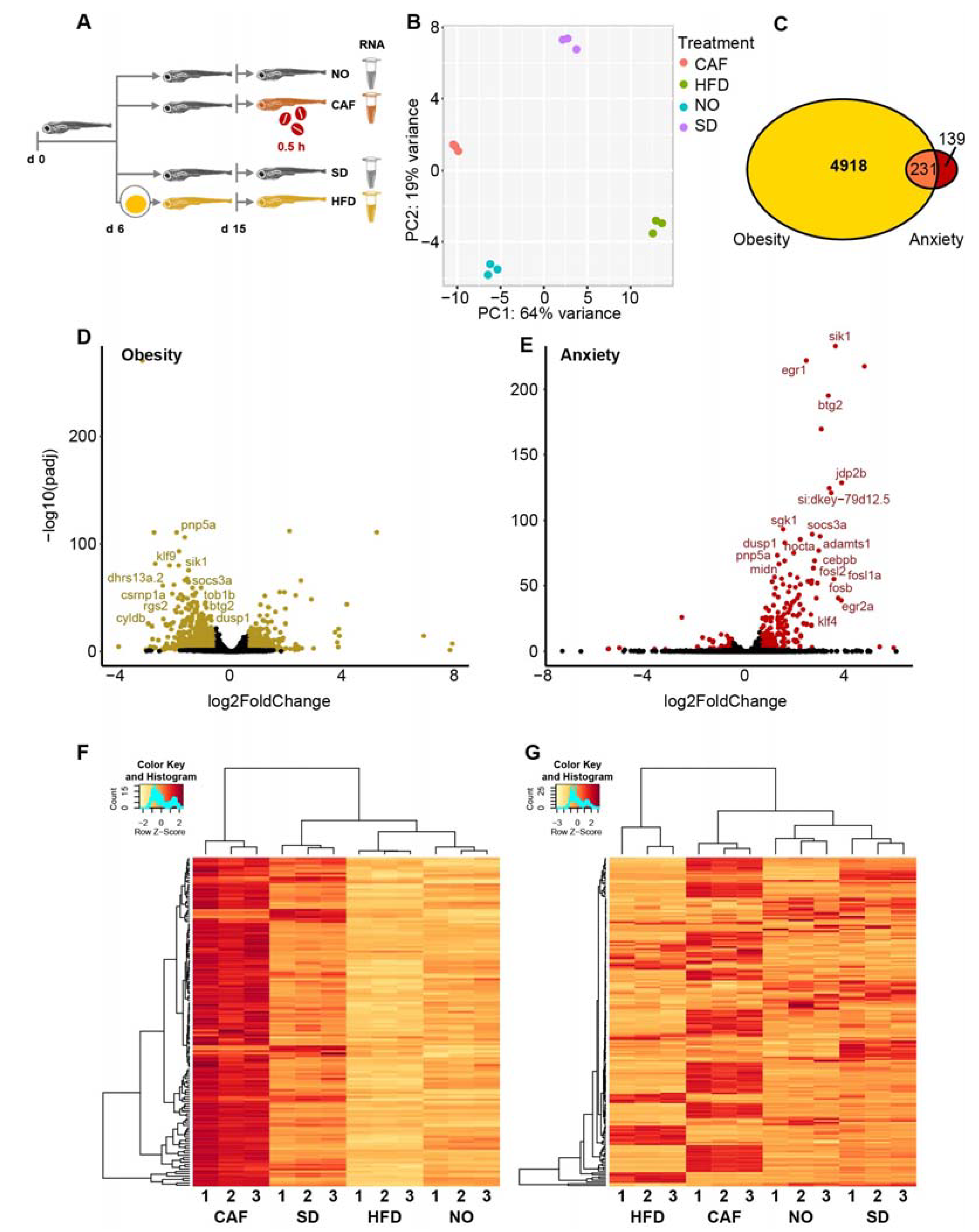
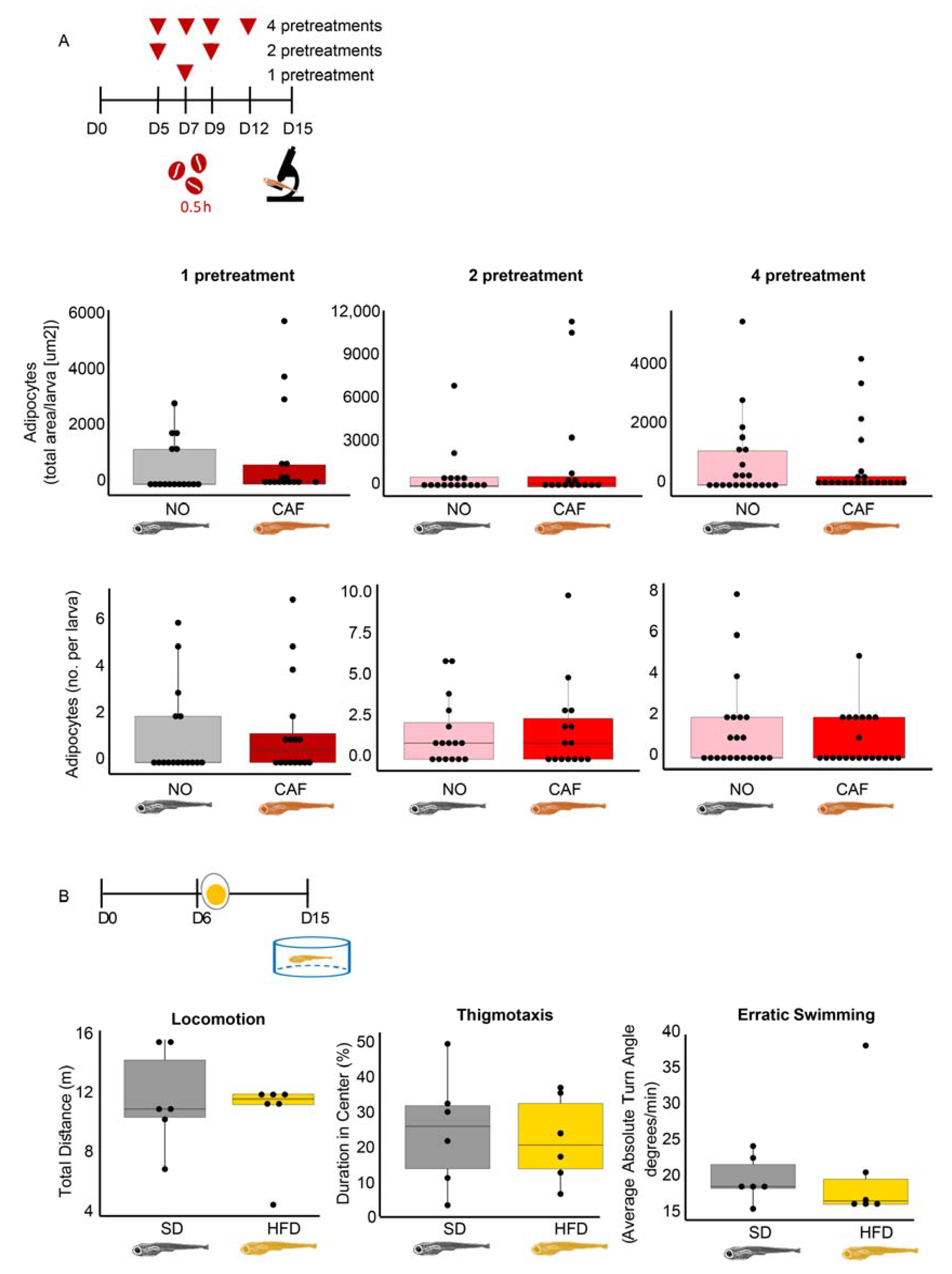
3.1. Experimental Validation of the Anxiety and Obesity Models
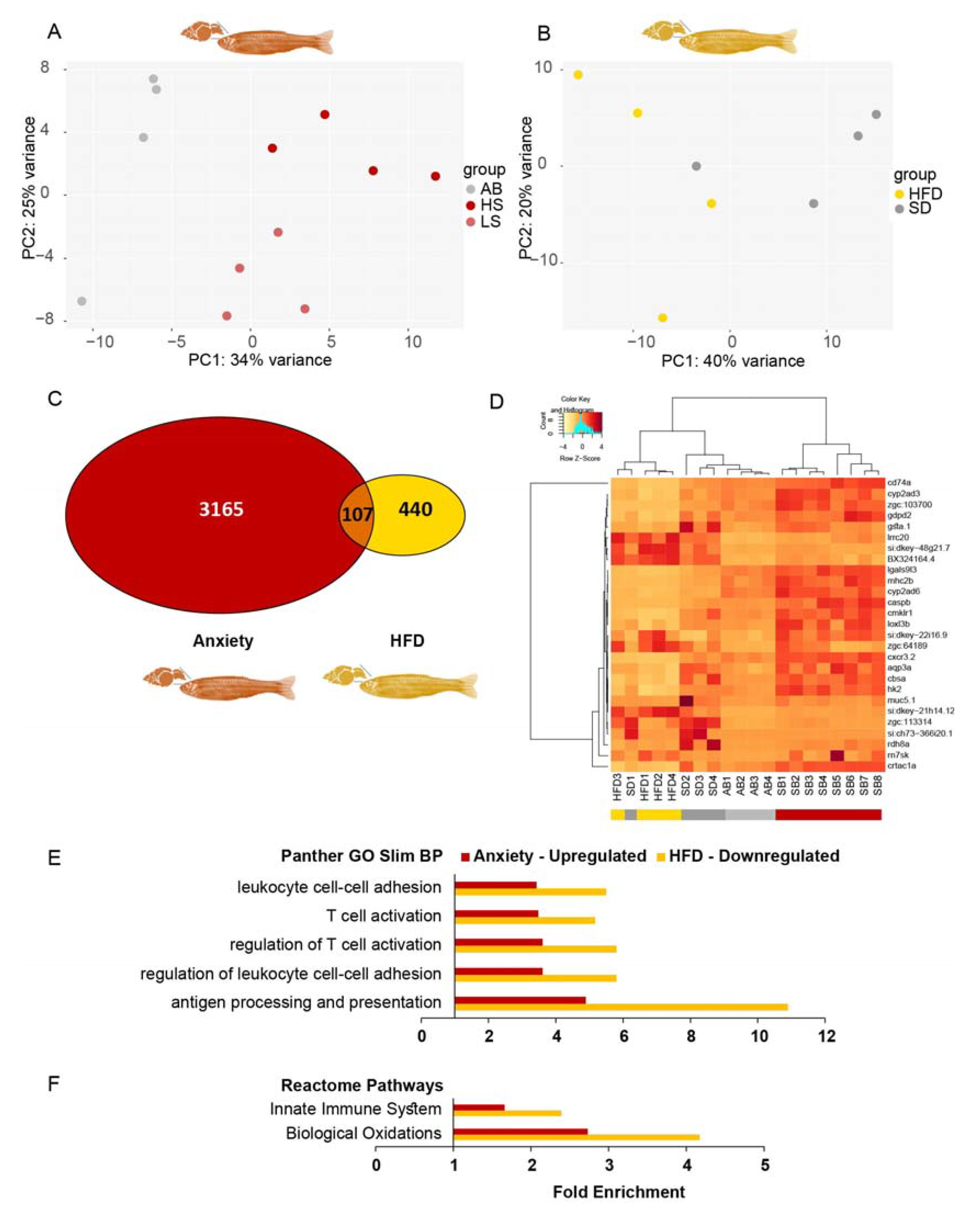
3.2. Most Transcripts That Were Upregulated in Anxious Larvae Were Downregulated in Obese Larvae
3.3. DE Isoform Results Were Similar to DE Gene Results
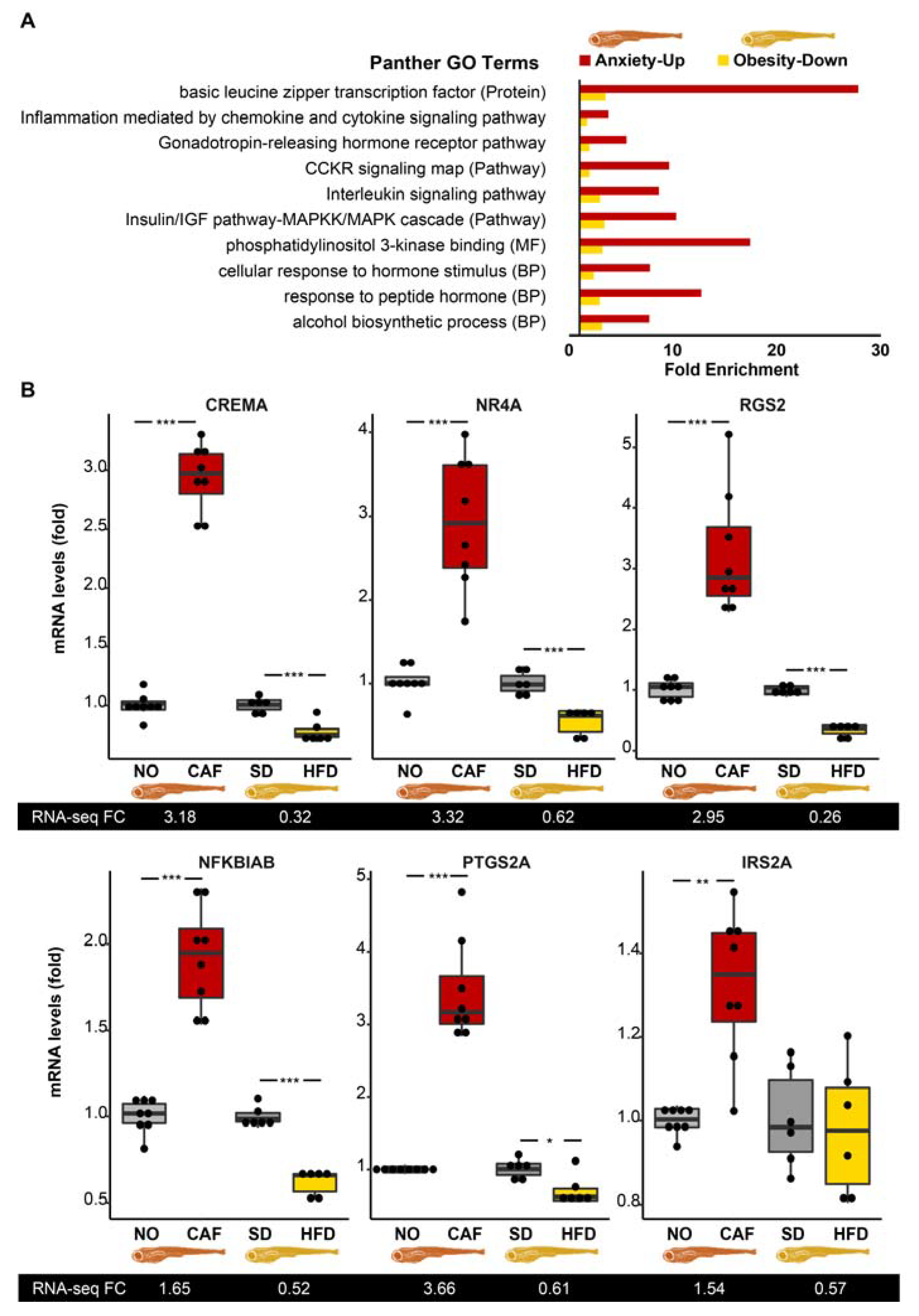
3.4. Inflammation/Immune System Protein-Coding Transcripts Are Upregulated in Anxious Larvae and Downregulated in Obese Larvae
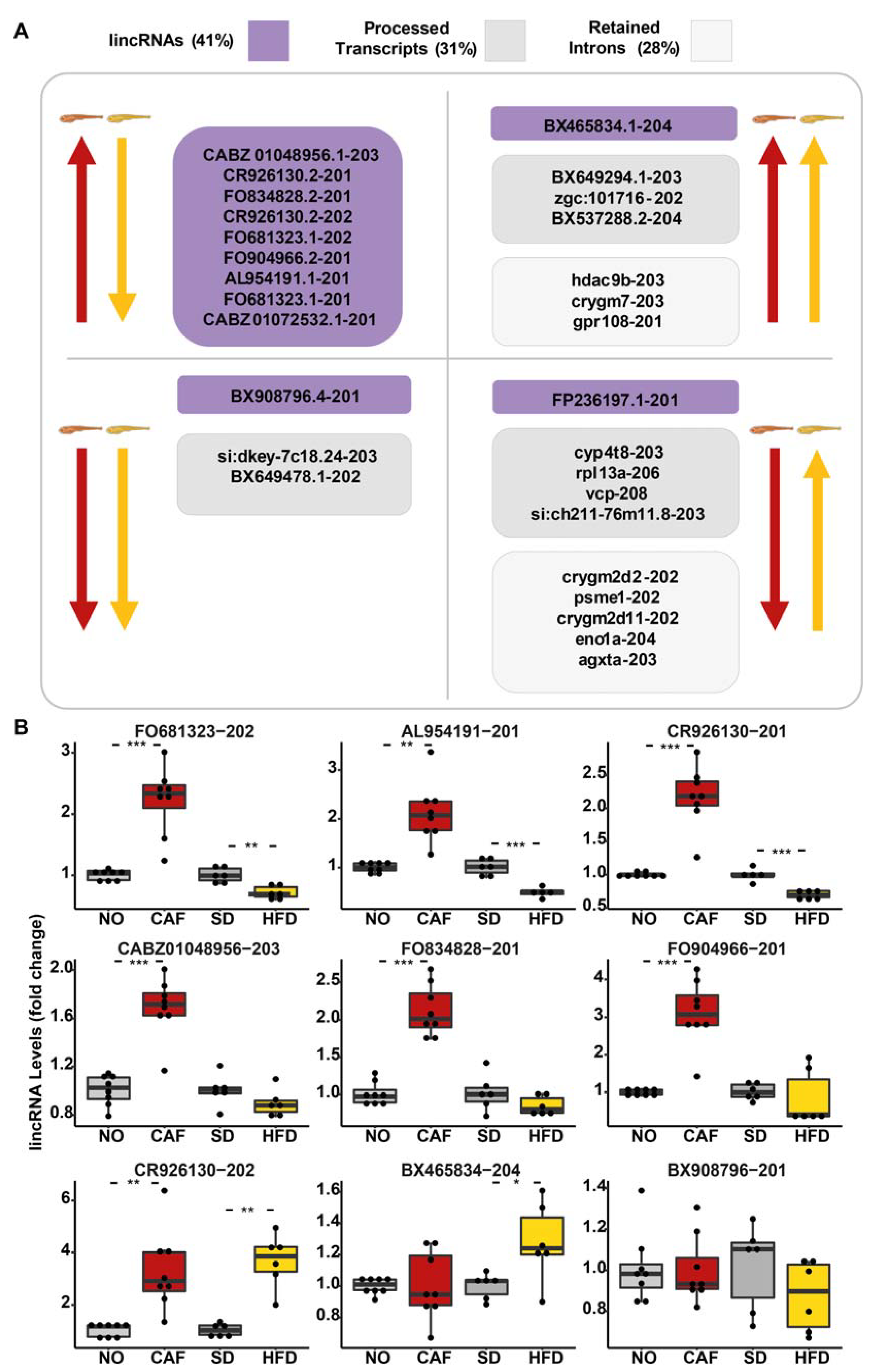
3.5. lincRNAs Upregulated in the Anxiety Model Were Downregulated in the Obesity Model
3.6. The Processed Transcript, si:dkey-7c18.24-203, Was Downregulated in Both the Anxiety and the Obesity Models According to RNA-Seq and qRT-PCR
3.7. Neighboring mRNAs of DE lincRNA Did Not Imply Functions for lincRNA
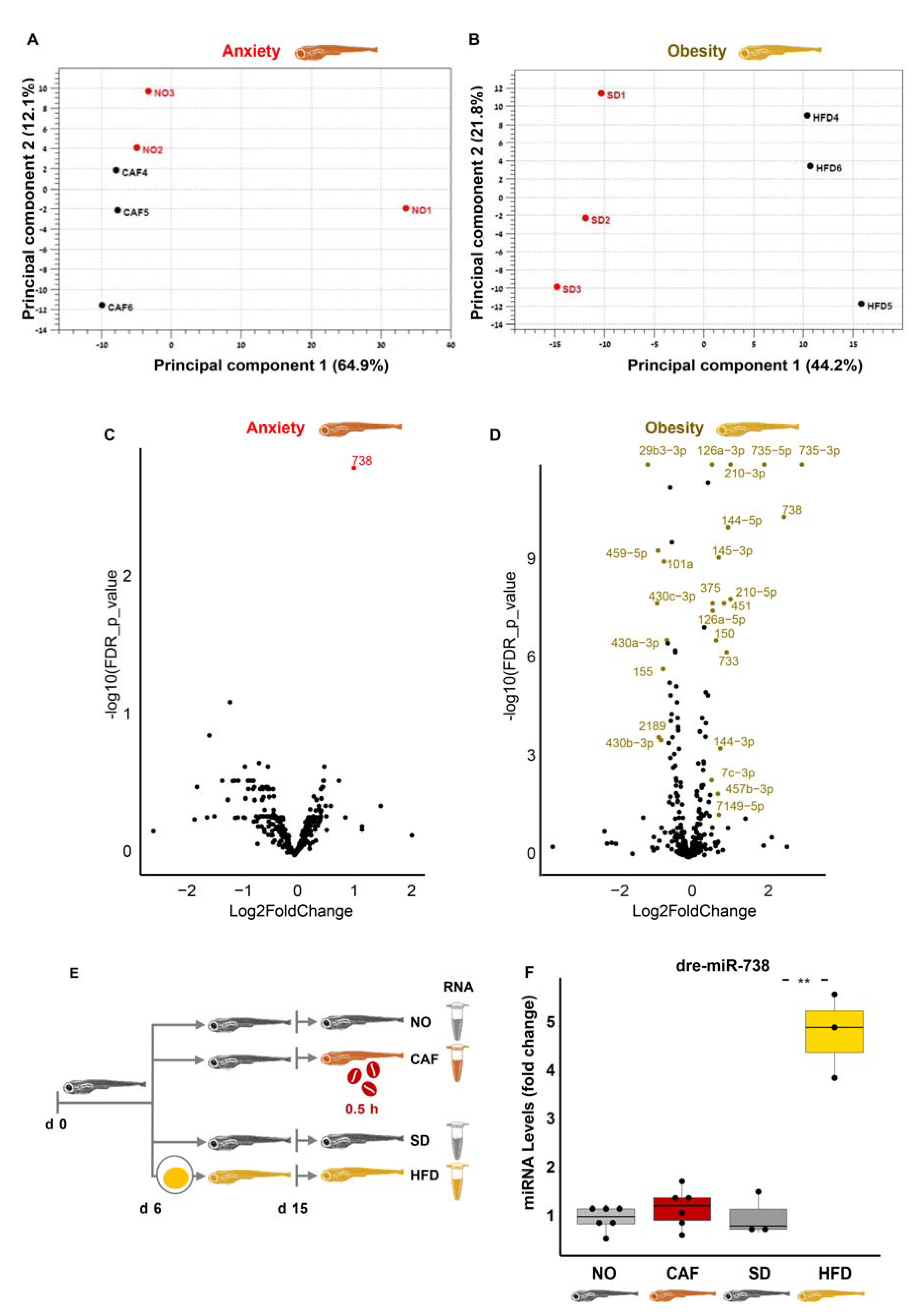
3.8. miRNAs Do Not Seem to Link Anxiety and Obesity in Zebrafish Larvae
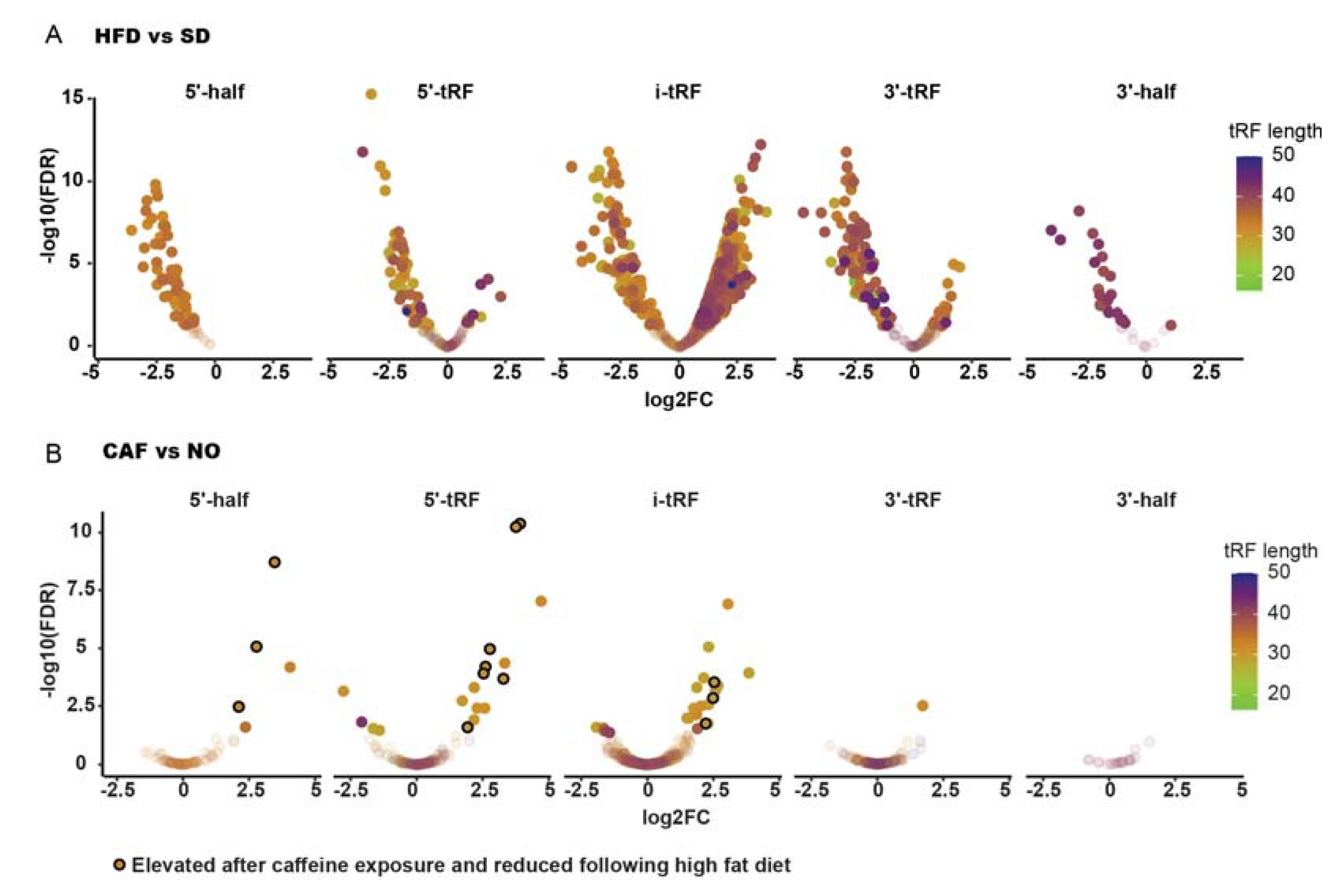
3.9. Larval tRFs Were Upregulated in Anxiety and Downregulated in Obesity
3.10. In Larvae, Obesity Fails to Accompany Anxiety, and Vice Versa
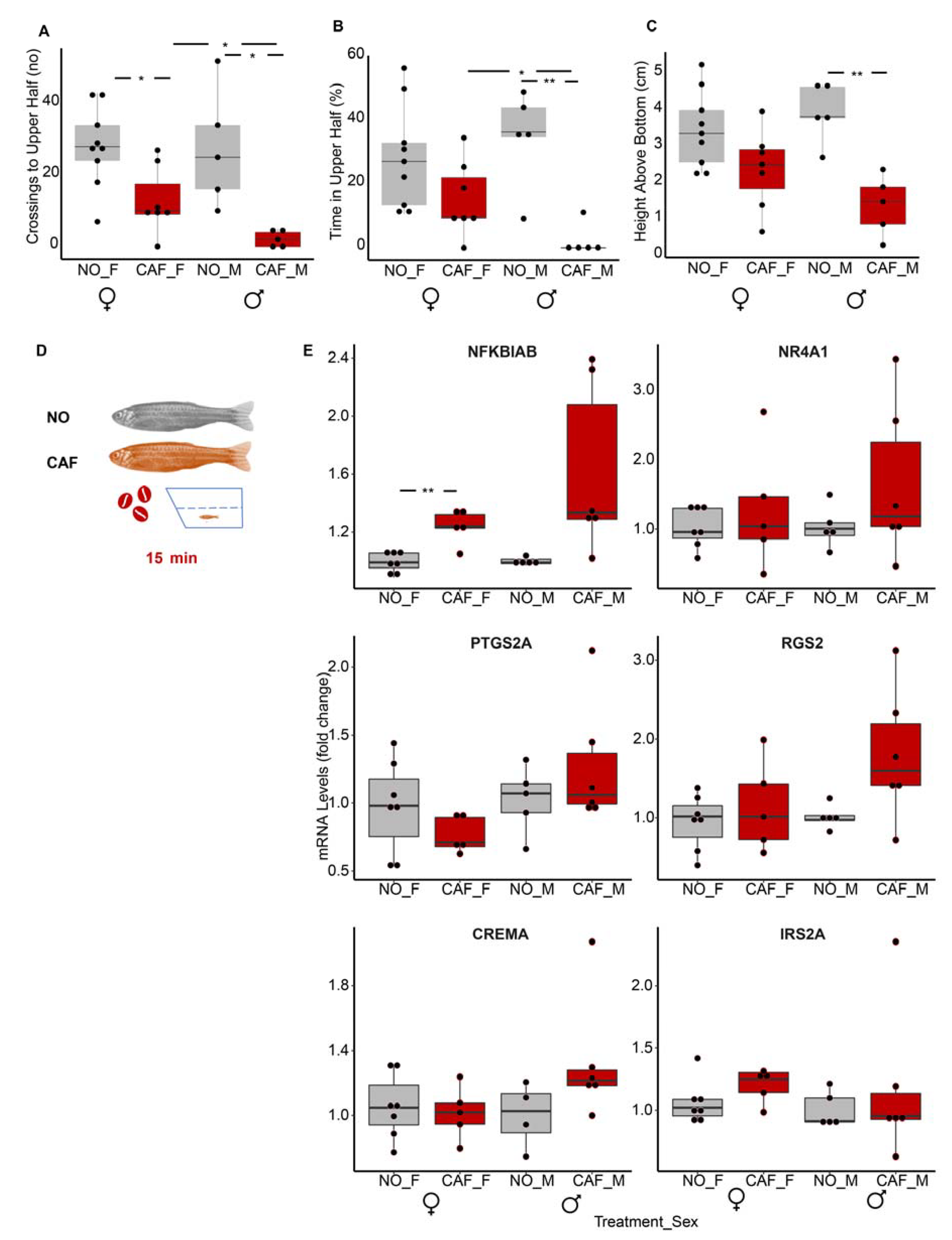
3.11. Age and Sex Both Seem to Affect Immune System-Linked mRNA Transcripts
3.12. LncRNA Profiles Changed with Age and Sex
3.13. Inversely Regulated Inflammation-Related DE Transcripts Differed between Zebrafish Adults and Larvae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robinson, O.J.; Pike, A.C.; Cornwell, B.; Grillon, C. The Translational Neural Circuitry of Anxiety. J. Neurol. Neurosurg. Psychiatry 2019, 90, 1353–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craske, M.G.; Stein, M.B.; Eley, T.C.; Milad, M.R.; Holmes, A.; Rapee, R.M.; Wittchen, H.-U. Anxiety Disorders. Nat. Rev. Dis. Prim. 2017, 3, 17024. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.M.; Deckert, J. Genetics of Anxiety Disorders. Curr. Psychiatry Rep. 2019, 21, 16. [Google Scholar] [CrossRef] [PubMed]
- Walsh, K.; Furey, W.J.; Malhi, N. Narrative Review: COVID-19 and Pediatric Anxiety. J. Psychiatr. Res. 2021, 144, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Sudre, C.H.; Murray, B.; Varsavsky, T.; Graham, M.S.; Penfold, R.S.; Bowyer, R.C.; Pujol, J.C.; Klaser, K.; Antonelli, M.; Canas, L.S.; et al. Attributes and Predictors of Long COVID. Nat. Med. 2021, 27, 626–631. [Google Scholar] [CrossRef]
- Huang, P.L. A Comprehensive Definition for Metabolic Syndrome. Dis. Model. Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Van Reedt Dortland, A.K.B.; Giltay, E.J.; Van Veen, T.; Zitman, F.G.; Penninx, B.W.J.H. Longitudinal Relationship of Depressive and Anxiety Symptoms with Dyslipidemia and Abdominal Obesity. Psychosom. Med. 2013, 75, 83–89. [Google Scholar] [CrossRef]
- Tang, F.; Wang, G.; Lian, Y. Association between Anxiety and Metabolic Syndrome: A Systematic Review and Meta-Analysis of Epidemiological Studies. Psychoneuroendocrinology 2017, 77, 112–121. [Google Scholar] [CrossRef]
- Dutheil, S.; Ota, K.T.; Wohleb, E.S.; Rasmussen, K.; Duman, R.S. High-Fat Diet Induced Anxiety and Anhedonia: Impact on Brain Homeostasis and Inflammation. Neuropsychopharmacology 2016, 41, 1874–1887. [Google Scholar] [CrossRef] [Green Version]
- Xia, G.; Han, Y.; Meng, F.; He, Y.; Srisai, D.; Farias, M.; Dang, M.; Palmiter, R.D.; Xu, Y.; Wu, Q. Reciprocal Control of Obesity and Anxiety–Depressive Disorder via a GABA and Serotonin Neural Circuit. Mol. Psychiatry 2021, 26, 2837–2853. [Google Scholar] [CrossRef]
- Almeida-Suhett, C.P.; Graham, A.; Chen, Y.; Deuster, P. Behavioral Changes in Male Mice Fed a High-Fat Diet Are Associated with IL-1β Expression in Specific Brain Regions. Physiol. Behav. 2017, 169, 130–140. [Google Scholar] [CrossRef]
- Meydan, C.; Shenhar-Tsarfaty, S.; Soreq, H. MicroRNA Regulators of Anxiety and Metabolic Disorders. Trends Mol. Med. 2016, 22, 798–812. [Google Scholar] [CrossRef] [PubMed]
- Türkoğlu, M.; Baran, A.; Sulukan, E.; Ghosigharehagaji, A.; Yildirim, S.; Ceyhun, H.A.; Bolat, İ.; Arslan, M.; Ceyhun, S.B. The Potential Effect Mechanism of High-Fat and High-Carbohydrate Diet-Induced Obesity on Anxiety and Offspring of Zebrafish. Eat. Weight Disord. Stud. Anorexia Bulim. Obes. 2022, 27, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Picolo, V.L.; Quadros, V.A.; Canzian, J.; Grisolia, C.K.; Goulart, J.T.; Pantoja, C.; de Bem, A.F.; Rosemberg, D.B. Short-Term High-Fat Diet Induces Cognitive Decline, Aggression, and Anxiety-like Behavior in Adult Zebrafish. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 110, 110288. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, L.; Hagman, E.; Danielsson, P.; Marcus, C.; Persson, M. Anxiety and Depression in Children and Adolescents with Obesity: A Nationwide Study in Sweden. BMC Med. 2020, 18, 30. [Google Scholar] [CrossRef] [Green Version]
- Shimada-Sugimoto, M.; Otowa, T.; Hettema, J.M. Genetics of Anxiety Disorders: Genetic Epidemiological and Molecular Studies in Humans. Psychiatry Clin. Neurosci. 2015, 69, 388–401. [Google Scholar] [CrossRef]
- Ke, W.; Reed, J.N.; Yang, C.; Higgason, N.; Rayyan, L.; Wahlby, C.; Carpenter, A.E.; Civelek, M.; O’Rourke, E.J. Genes in Human Obesity Loci Are Causal Obesity Genes in C. elegans. PLoS Genet. 2021, 17, e1009736. [Google Scholar] [CrossRef]
- Baker, K.D.; Loughman, A.; Spencer, S.J.; Reichelt, A.C. The Impact of Obesity and Hypercaloric Diet Consumption on Anxiety and Emotional Behavior across the Lifespan. Neurosci. Biobehav. Rev. 2017, 83, 173–182. [Google Scholar] [CrossRef]
- Nousen, E.K.; Franco, J.G.; Sullivan, E.L. Unraveling the Mechanisms Responsible for the Comorbidity between Metabolic Syndrome and Mental Health Disorders. Neuroendocrinology 2014, 98, 254–266. [Google Scholar] [CrossRef] [Green Version]
- Hiles, S.A.; Révész, D.; Lamers, F.; Giltay, E.; Penninx, B.W.J.H. Bidirectional Prospective Associations of Metabolic Syndrome Components with Depression, Anxiety, and Antidepressant Use. Depress. Anxiety 2016, 33, 754–764. [Google Scholar] [CrossRef] [Green Version]
- Gainey, S.J.; Kwakwa, K.A.; Bray, J.K.; Pillote, M.M.; Tir, V.L.; Towers, A.E.; Freund, G.G. Short-Term High-Fat Diet (HFD) Induced Anxiety-like Behaviors and Cognitive Impairment Are Improved with Treatment by Glyburide. Front. Behav. Neurosci. 2016, 10, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogrodnik, M.; Zhu, Y.; Langhi, L.G.P.; Tchkonia, T.; Krüger, P.; Fielder, E.; Victorelli, S.; Ruswhandi, R.A.; Giorgadze, N.; Pirtskhalava, T.; et al. Obesity-Induced Cellular Senescence Drives Anxiety and Impairs Neurogenesis. Cell Metab. 2019, 29, 1061–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Caraballo, Y.; Hodgson, K.J.; Morgan, S.A.; Ferrario, C.R.; Vollbrecht, P.J. Enhanced Anxiety-like Behavior Emerges with Weight Gain in Male and Female Obesity-Susceptible Rats. Behav. Brain Res. 2019, 360, 81–93. [Google Scholar] [CrossRef]
- Singleman, C.; Holtzman, N.G. Growth and Maturation in the Zebrafish, Danio rerio: A Staging Tool for Teaching and Research. Zebrafish 2014, 11, 396–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, M.; Yang, E.; Neelkantan, N.; Mikhaylova, A.; Arnold, R.; Poudel, M.K.; Stewart, A.M.; Kalueff, A.V. Developing “integrative” Zebrafish Models of Behavioral and Metabolic Disorders. Behav. Brain Res. 2013, 256, 172–187. [Google Scholar] [CrossRef] [PubMed]
- Lieschke, G.J.; Currie, P.D. Animal Models of Human Disease: Zebrafish Swim into View. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The Zebrafish Reference Genome Sequence and Its Relationship to the Human Genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Zang, L.; Maddison, L.A.; Chen, W. Zebrafish as a Model for Obesity and Diabetes. Front. Cell Dev. Biol. 2018, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- López Nadal, A.; Ikeda-Ohtsubo, W.; Sipkema, D.; Peggs, D.; McGurk, C.; Forlenza, M.; Wiegertjes, G.F.; Brugman, S. Feed, Microbiota, and Gut Immunity: Using the Zebrafish Model to Understand Fish Health. Front. Immunol. 2020, 11, 114. [Google Scholar] [CrossRef]
- Fontana, B.D.; Mezzomo, N.J.; Kalueff, A.V.; Rosemberg, D.B. The Developing Utility of Zebrafish Models of Neurological and Neuropsychiatric Disorders: A Critical Review. Exp. Neurol. 2018, 299, 157–171. [Google Scholar] [CrossRef]
- Novoa, B.; Figueras, A. Zebrafish: Model for the Study of Inflammation and the Innate Immune Response to Infectious Diseases. In Current Topics in Innate Immunity II; Springer: Berlin/Heidelberg, Germany, 2012; Volume 946, pp. 253–275. ISBN 978-1-4614-0105-6. [Google Scholar]
- de Abreu, M.S.; Giacomini, A.C.V.V.; Zanandrea, R.; dos Santos, B.E.; Genario, R.; de Oliveira, G.G.; Friend, A.J.; Amstislavskaya, T.G.; Kalueff, A.V. Psychoneuroimmunology and Immunopsychiatry of Zebrafish. Psychoneuroendocrinology 2018, 92, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, R.W.; Jacobson, G.A.; Zhu, P. Circuit Neuroscience in Zebrafish. Curr. Biol. 2010, 20, R371–R381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, R.J.; Bergner, C.L.; Hart, P.C.; Cachat, J.M.; Canavello, P.R.; Elegante, M.F.; Elkhayat, S.I.; Bartels, B.K.; Tien, A.K.; Tien, D.H.; et al. Understanding Behavioral and Physiological Phenotypes of Stress and Anxiety in Zebrafish. Behav. Brain Res. 2009, 205, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenze, E.J.; Mantella, R.C.; Shi, P.; Goate, A.M.; Nowotny, P.; Butters, M.A.; Andreescu, C.; Thompson, P.A.; Rollman, B.L. Elevated Cortisol in Older Adults with Generalized Anxiety Disorder Is Reduced by Treatment: A Placebo-Controlled Evaluation of Escitalopram. Am. J. Geriatr. Psychiatry 2011, 19, 482–490. [Google Scholar] [CrossRef] [Green Version]
- Blay, S.L.; Marinho, V. Anxiety Disorders in Old Age. Curr. Opin. Psychiatry 2012, 25, 462–467. [Google Scholar] [CrossRef]
- Aprile, M.; Katopodi, V.; Leucci, E.; Costa, V. LncRNAs in Cancer: From Garbage to Junk. Cancers 2020, 12, 3220. [Google Scholar] [CrossRef]
- Issler, O.; Chen, A. Determining the Role of microRNAs in Psychiatric Disorders. Nat. Rev. Neurosci. 2015, 16, 201–212. [Google Scholar] [CrossRef]
- Kolshus, E.; Dalton, V.S.; Ryan, K.M.; McLoughlin, D.M. When less is more—microRNAs and psychiatric disorders. Acta Psychiatr. Scand. 2014, 129, 241–256. [Google Scholar] [CrossRef]
- Murphy, C.P.; Singewald, N. Role of microRNAs in Anxiety and Anxiety-Related Disorders. In Brain Imaging in Behavioral Neuroscience; Springer: Berlin/Heidelberg, Germany, 2019; pp. 289–320. ISBN 9783642209246. [Google Scholar]
- Cui, X.; Niu, W.; Kong, L.; He, M.; Jiang, K.; Chen, S.; Zhong, A.; Zhang, Q.; Li, W.; Lu, J.; et al. Long Noncoding RNA as an Indicator Differentiating Schizophrenia from Major Depressive Disorder and Generalized Anxiety Disorder in Nonpsychiatric Hospital. Biomark. Med. 2017, 11, 221–228. [Google Scholar] [CrossRef]
- Spadaro, P.A.; Flavell, C.R.; Widagdo, J.; Ratnu, V.S.; Troup, M.; Ragan, C.; Mattick, J.S.; Bredy, T.W. Long Noncoding RNA-Directed Epigenetic Regulation of Gene Expression Is Associated with Anxiety-like Behavior in Mice. Biol. Psychiatry 2015, 78, 848–859. [Google Scholar] [CrossRef] [Green Version]
- Iacomino, G.; Siani, A. Role of microRNAs in Obesity and Obesity-Related Diseases. Genes Nutr. 2017, 12, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arner, P.; Kulyté, A. microRNA Regulatory Networks in Human Adipose Tissue and Obesity. Nat. Rev. Endocrinol. 2015, 11, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Aryal, B.; Singh, A.K.; Rotllan, N.; Price, N.; Fernández-Hernando, C. microRNAs and Lipid Metabolism. Curr. Opin. Lipidol. 2017, 28, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Lo, K.A.; Huang, S.; Walet, C.; Zhang, Z.; Leow, M.; Meihui, L.; Sun, L. Adipocyte Long Noncoding RNA Transcriptome Analysis of Obese Mice Identified Lnc-Leptin Which Regulates Leptin. Diabetes 2018, 67, 1045–1056. [Google Scholar] [CrossRef] [Green Version]
- Cai, R.; Sun, Y.; Qimuge, N.; Wang, G.; Wang, Y.; Chu, G.; Yu, T.; Yang, G.; Pang, W. Adiponectin AS lncRNA Inhibits Adipogenesis by Transferring from Nucleus to Cytoplasm and Attenuating Adiponectin mRNA Translation. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2018, 1863, 420–432. [Google Scholar] [CrossRef]
- Zorbaz, T.; Madrer, N.; Soreq, H. Cholinergic Blockade of Neuroinflammation: From Tissue to RNA Regulators. Neuronal Signal. 2022, 6, NS20210035. [Google Scholar] [CrossRef]
- Winek, K.; Lobentanzer, S.; Nadorp, B.; Dubnov, S.; Dames, C.; Jagdmann, S.; Moshitzky, G.; Hotter, B.; Meisel, C.; Greenberg, D.S.; et al. Transfer RNA Fragments Replace microRNA Regulators of the Cholinergic Poststroke Immune Blockade. Proc. Natl. Acad. Sci. USA 2020, 117, 32606–32616. [Google Scholar] [CrossRef]
- Mathew, B.A.; Katta, M.; Ludhiadch, A.; Singh, P.; Munshi, A. Role of TRNA-Derived Fragments in Neurological Disorders: A Review. Mol. Neurobiol. 2022, 60, 655–671. [Google Scholar] [CrossRef]
- Kim, H.K.; Yeom, J.H.; Kay, M.A. Transfer RNA-Derived Small RNAs: Another Layer of Gene Regulation and Novel Targets for Disease Therapeutics. Mol. Ther. 2020, 28, 2340–2357. [Google Scholar] [CrossRef]
- Rosace, D.; López, J.; Blanco, S. Emerging Roles of Novel Small Non-Coding Regulatory RNAs in Immunity and Cancer. RNA Biol. 2020, 17, 1196–1213. [Google Scholar] [CrossRef] [Green Version]
- Paldor, I.; Madrer, N.; Vaknine Treidel, S.; Shulman, D.; Greenberg, D.S.; Soreq, H. Cerebrospinal Fluid and Blood Profiles of Transfer RNA Fragments Show Age, Sex, and Parkinson’s Disease-Related Changes. J. Neurochem. 2022, 164, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Zang, L.; Shimada, Y.; Nakayama, H.; Kim, Y.; Chu, D.C.; Juneja, L.R.; Kuroyanagi, J.; Nishimura, N. RNA-Seq Based Transcriptome Analysis of the Anti-Obesity Effect of Green Tea Extract Using Zebrafish Obesity Models. Molecules 2019, 24, 3256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serikuly, N.; Alpyshov, E.T.; Wang, D.; Wang, J.; Yang, L.; Hu, G.; Yan, D.; Demin, K.A.; Kolesnikova, T.O.; Galstyan, D.; et al. Effects of Acute and Chronic Arecoline in Adult Zebrafish: Anxiolytic-like Activity, Elevated Brain Monoamines and the Potential Role of Microglia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 104, 109977. [Google Scholar] [CrossRef] [PubMed]
- den Broeder, M.; Moester, M.; Kamstra, J.; Cenijn, P.; Davidoiu, V.; Kamminga, L.; Ariese, F.; de Boer, J.; Legler, J. Altered Adipogenesis in Zebrafish Larvae Following High Fat Diet and Chemical Exposure Is Visualised by Stimulated Raman Scattering Microscopy. Int. J. Mol. Sci. 2017, 18, 894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlegel, A.; Gut, P. Metabolic Insights from Zebrafish Genetics, Physiology, and Chemical Biology. Cell. Mol. Life Sci. 2015, 72, 2249–2260. [Google Scholar] [CrossRef] [Green Version]
- Van Den Bos, R.; Mes, W.; Galligani, P.; Heil, A.; Zethof, J.; Flik, G.; Gorissen, M. Further Characterisation of Differences between TL and AB Zebrafish (Danio rerio): Gene Expression, Physiology and Behaviour at Day 5 of the Larval Stage. PLoS ONE 2017, 12, e0175420. [Google Scholar] [CrossRef] [Green Version]
- Richendrfer, H.; Pelkowski, S.D.; Colwill, R.M.; Creton, R. On the Edge: Pharmacological Evidence for Anxiety-Related Behavior in Zebrafish Larvae. Behav. Brain Res. 2012, 228, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Rosa, L.V.; Ardais, A.P.; Costa, F.V.; Fontana, B.D.; Quadros, V.A.; Porciúncula, L.O.; Rosemberg, D.B. Different Effects of Caffeine on Behavioral Neurophenotypes of Two Zebrafish Populations. Pharmacol. Biochem. Behav. 2018, 165, 1–8. [Google Scholar] [CrossRef]
- Tran, S.; Fulcher, N.; Nowicki, M.; Desai, P.; Tsang, B.; Facciol, A.; Chow, H.; Gerlai, R. Time-Dependent Interacting Effects of Caffeine, Diazepam, and Ethanol on Zebrafish Behaviour. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 75, 16–27. [Google Scholar] [CrossRef]
- Aleström, P.; D’Angelo, L.; Midtlyng, P.J.; Schorderet, D.F.; Schulte-Merker, S.; Sohm, F.; Warner, S. Zebrafish: Housing and Husbandry Recommendations. Lab. Anim. 2019, 54, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Yeh, C.-M.; Glöck, M.; Ryu, S. An Optimized Whole-Body Cortisol Quantification Method for Assessing Stress Levels in Larval Zebrafish. PLoS ONE 2013, 8, e79406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minchin, J.E.N.; Rawls, J.F. In Vivo Analysis of White Adipose Tissue in Zebrafish. Methods Cell Biol. 2011, 105, 63–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio rerio), 5th ed.; University of Oregon Press: Eugene, OR, USA, 2007. [Google Scholar]
- Loher, P.; Telonis, A.G.; Rigoutsos, I. MINTmap: Fast and Exhaustive Profiling of Nuclear and Mitochondrial TRNA Fragments from Short RNA-Seq Data. Sci. Rep. 2017, 7, srep41184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Balcells, I.; Cirera, S.; Busk, P.K. Specific and Sensitive Quantitative RT-PCR of miRNAs with DNA Primers. BMC Biotechnol. 2011, 11, 70. [Google Scholar] [CrossRef] [Green Version]
- Busk, P.K. A Tool for Design of Primers for microRNA-Specific Quantitative RT-QPCR. BMC Bioinform. 2014, 15, 29. [Google Scholar] [CrossRef] [Green Version]
- Tang, R.; Dodd, A.; Lai, D.; McNabb, W.C.; Love, D.R. Validation of Zebrafish (Danio rerio) Reference Genes for Quantitative Real-Time RT-PCR Normalization. Acta Biochim. Biophys. Sin. 2007, 39, 384–390. [Google Scholar] [CrossRef] [Green Version]
- Wong, R.Y.; Godwin, J. Neurotranscriptome Profiles of Multiple Zebrafish Strains. Genom. Data 2015, 5, 206–209. [Google Scholar] [CrossRef] [Green Version]
- Wong, R.Y.; Perrin, F.; Oxendine, S.E.; Kezios, Z.D.; Sawyer, S.; Zhou, L.; Dereje, S.; Godwin, J. Comparing Behavioral Responses across Multiple Assays of Stress and Anxiety in Zebrafish (Danio rerio). Behaviour 2012, 149, 1205–1240. [Google Scholar] [CrossRef] [Green Version]
- Meguro, S.; Hosoi, S.; Hasumura, T. High-Fat Diet Impairs Cognitive Function of Zebrafish. Sci. Rep. 2019, 9, 17063. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2; Use R!; Springer International Publishing: Cham, Switzerland, 2016; ISBN 978-3-319-24275-0. [Google Scholar]
- Yin, J.; Zhang, J.; Lu, Q. The Role of Basic Leucine Zipper Transcription Factor E4BP4 in the Immune System and Immune-Mediated Diseases. Clin. Immunol. 2017, 180, 5–10. [Google Scholar] [CrossRef]
- Madrer, N.; Soreq, H. Cholino-ncRNAs Modulate Sex-Specific- and Age-Related Acetylcholine Signals. FEBS Lett. 2020, 594, 2185–2198. [Google Scholar] [CrossRef]
- Welty, F.K.; Alfaddagh, A.; Elajami, T.K. Targeting Inflammation in Metabolic Syndrome. Transl. Res. 2016, 167, 257–280. [Google Scholar] [CrossRef]
- Francés, D.E.; Motiño, O.; Agrá, N.; González-Rodríguez, Á.; Fernández-Álvarez, A.; Cucarella, C.; Mayoral, R.; Castro-Sánchez, L.; García-Casarrubios, E.; Boscá, L.; et al. Hepatic Cyclooxygenase-2 Expression Protects against Diet-Induced Steatosis, Obesity, and Insulin Resistance. Diabetes 2015, 64, 1522–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Calvo, R.; Tajes, M.; Vázquez-Carrera, M. The NR4A Subfamily of Nuclear Receptors: Potential New Therapeutic Targets for the Treatment of Inflammatory Diseases. Expert Opin. Ther. Targets 2017, 21, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Catrysse, L.; van Loo, G. Inflammation and the Metabolic Syndrome: The Tissue-Specific Functions of NF-ΚB. Trends Cell Biol. 2017, 27, 417–429. [Google Scholar] [CrossRef]
- Cai, D.; Liu, T. INTRODUCTION: Brain Inflammation in Metabolic Syndrome. Aging 2012, 4, 98–115. [Google Scholar] [CrossRef] [PubMed]
- McNabb, H.J.; Zhang, Q.; Sjögren, B. Emerging Roles for Regulator of G Protein Signaling 2 in (Patho)physiology. Mol. Pharmacol. 2020, 98, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Hui, L.; Zheng, K.; Liu, L.; Liu, J.; Lv, W. Silencing of RGS2 Enhances Hippocampal Neuron Regeneration and Rescues Depression-like Behavioral Impairments through Activation of CAMP Pathway. Brain Res. 2020, 1746, 147018. [Google Scholar] [CrossRef]
- Deniz, E.; Erman, B. Long Noncoding RNA (lincRNA), a New Paradigm in Gene Expression Control. Funct. Integr. Genom. 2017, 17, 135–143. [Google Scholar] [CrossRef]
- Khyzha, N.; Khor, M.; DiStefano, P.V.; Wang, L.; Matic, L.; Hedin, U.; Wilson, M.D.; Maegdefessel, L.; Fish, J.E. Regulation of CCL2 Expression in Human Vascular Endothelial Cells by a Neighboring Divergently Transcribed Long Noncoding RNA. Proc. Natl. Acad. Sci. USA 2019, 116, 16410–16419. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.; Elegante, M.; Bartels, B.; Elkhayat, S.; Tien, D.; Roy, S.; Goodspeed, J.; Suciu, C.; Tan, J.; Grimes, C.; et al. Analyzing Habituation Responses to Novelty in Zebrafish (Danio rerio). Behav. Brain Res. 2010, 208, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Fontana, B.D.; Cleal, M.; Parker, M.O. Female Adult Zebrafish (Danio rerio) Show Higher Levels of Anxiety-like Behavior than Males, but Do Not Differ in Learning and Memory Capacity. Eur. J. Neurosci. 2020, 52, 2604–2613. [Google Scholar] [CrossRef] [PubMed]
- Mendes, N.F.; Castro, G.; Guadagnini, D.; Tobar, N.; Cognuck, S.Q.; Elias, L.L.K.; Boer, P.A.; Prada, P.O. Knocking down Amygdalar PTP1B in Diet-Induced Obese Rats Improves Insulin Signaling/Action, Decreases Adiposity and May Alter Anxiety Behavior. Metabolism 2017, 70, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto, M.; Herzog, C.; Pacheco, J.A.; Fujisaka, S.; Bullock, K.; Clish, C.B.; Kahn, C.R. Gut Microbiota Modulate Neurobehavior through Changes in Brain Insulin Sensitivity and Metabolism. Mol. Psychiatry 2018, 23, 2287–2301. [Google Scholar] [CrossRef]
- Fourrier, C.; Bosch-Bouju, C.; Boursereau, R.; Sauvant, J.; Aubert, A.; Capuron, L.; Ferreira, G.; Layé, S.; Castanon, N. Brain Tumor Necrosis Factor-α Mediates Anxiety-like Behavior in a Mouse Model of Severe Obesity. Brain. Behav. Immun. 2019, 77, 25–36. [Google Scholar] [CrossRef]
- Pierce, G.L.; Kalil, G.Z.; Ajibewa, T.; Holwerda, S.W.; Persons, J.; Moser, D.J.; Fiedorowicz, J.G. Anxiety Independently Contributes to Elevated Inflammation in Humans with Obesity. Obesity 2017, 25, 286–289. [Google Scholar] [CrossRef] [Green Version]
- Marín-Juez, R.; Jong-Raadsen, S.; Yang, S.; Spaink, H.P. Hyperinsulinemia Induces Insulin Resistance and Immune Suppression via Ptpn6/Shp1 in Zebrafish. J. Endocrinol. 2014, 222, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, C.C.; Cheng, H.H.; Tewari, M. MicroRNA Profiling: Approaches and Considerations. Nat. Rev. Genet. 2012, 13, 358–369. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Y.; Tan, X.; Mao, X.; Wei, D.; Yao, Y.; Jiang, P.; Mo, D.; Wang, T.; Yan, F. Identification of TRNA-Derived Fragments Expression Profile in Breast Cancer Tissues. Curr. Genom. 2019, 20, 199–213. [Google Scholar] [CrossRef]
- Filetti, V.; La Ferlita, A.; Di Maria, A.; Cardile, V.; Graziano, A.C.E.; Rapisarda, V.; Ledda, C.; Pulvirenti, A.; Loreto, C. Dysregulation of microRNAs and tRNA-Derived ncRNAs in Mesothelial and Mesothelioma Cell Lines after Asbestiform Fiber Exposure. Sci. Rep. 2022, 12, 9181. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.-Q.; Li, Y.-Y.; Wang, H.-L.; Mao, X.-T.; Guo, J.-X.; Wang, F.; Huang, L.-J.; Ma, X.-Y.; Gao, Z.-J.; Chen, W.; et al. Stress-Induced Metabolic Disorder in Peripheral CD4+ T Cells Leads to Anxiety-like Behavior. Cell 2019, 179, 864–879.e19. [Google Scholar] [CrossRef] [PubMed]
- Filiano, A.J.; Xu, Y.; Tustison, N.J.; Marsh, R.L.; Baker, W.; Smirnov, I.; Overall, C.C.; Gadani, S.P.; Turner, S.D.; Weng, Z.; et al. Unexpected Role of Interferon-γ in Regulating Neuronal Connectivity and Social Behaviour. Nature 2016, 535, 425–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, G.; Cho, K.A.; Song, J.; Kim, Y.K. Transcriptomic Analysis of High Fat Diet Fed Mouse Brain Cortex. Front. Genet. 2019, 10, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeilly, A.D.; Stewart, C.A.; Sutherland, C.; Balfour, D.J.K. High Fat Feeding Is Associated with Stimulation of the Hypothalamic-Pituitary-Adrenal Axis and Reduced Anxiety in the Rat. Psychoneuroendocrinology 2015, 52, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Poitou, C.; Perret, C.; Mathieu, F.; Truong, V.; Blum, Y.; Durand, H.; Alili, R.; Chelghoum, N.; Pelloux, V.; Aron-Wisnewsky, J.; et al. Bariatric Surgery Induces Disruption in Inflammatory Signaling Pathways Mediated by Immune Cells in Adipose Tissue: A RNA-seq Study. PLoS ONE 2015, 10, e0125718. [Google Scholar] [CrossRef]
- Fu, S.P.; Hong, H.; Lu, S.F.; Hu, C.J.; Xu, H.X.; Li, Q.; Yu, M.L.; Ou, C.; Meng, J.Z.; Wang, T.L.; et al. Genome-Wide Regulation of Electro-Acupuncture on the Neural Stat5-Loss-Induced Obese Mice. PLoS ONE 2017, 12, e0181948. [Google Scholar] [CrossRef] [Green Version]
- Rohm, T.V.; Meier, D.T.; Olefsky, J.M.; Donath, M.Y. Inflammation in Obesity, Diabetes, and Related Disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef]
- Mraz, M.; Haluzik, M. The Role of Adipose Tissue Immune Cells in Obesity and Low-Grade Inflammation. J. Endocrinol. 2014, 222, 113–127. [Google Scholar] [CrossRef] [Green Version]
- Michopoulos, V.; Powers, A.; Gillespie, C.F.; Ressler, K.J.; Jovanovic, T. Inflammation in Fear- and Anxiety-Based Disorders: PTSD, GAD, and Beyond. Neuropsychopharmacology 2017, 42, 254–270. [Google Scholar] [CrossRef] [Green Version]
- Felger, J.C. Imaging the Role of Inflammation in Mood and Anxiety-Related Disorders. Curr. Neuropharmacol. 2017, 15, 533–558. [Google Scholar] [CrossRef]
- Soreq, H. Checks and Balances on Cholinergic Signaling in Brain and Body Function. Trends Neurosci. 2015, 38, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus Nerve Stimulation Attenuates the Systemic Inflammatory Response to Endotoxin. Nature 2000, 405, 458–462. [Google Scholar] [CrossRef]
- Shaked, I.; Meerson, A.; Wolf, Y.; Avni, R.; Greenberg, D.; Gilboa-Geffen, A.; Soreq, H. MicroRNA-132 Potentiates Cholinergic Anti-Inflammatory Signaling by Targeting Acetylcholinesterase. Immunity 2009, 31, 965–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aten, S.; Page, C.E.; Kalidindi, A.; Wheaton, K.; Niraula, A.; Godbout, J.P.; Hoyt, K.R.; Obrietan, K. miR-132/212 Is Induced by Stress and Its Dysregulation Triggers Anxiety-Related Behavior. Neuropharmacology 2019, 144, 256–270. [Google Scholar] [CrossRef] [PubMed]
- Hanin, G.; Yayon, N.; Tzur, Y.; Haviv, R.; Bennett, E.R.; Udi, S.; Krishnamoorthy, Y.R.; Kotsiliti, E.; Zangen, R.; Efron, B.; et al. miRNA-132 Induces Hepatic Steatosis and Hyperlipidaemia by Synergistic Multitarget Suppression. Gut 2018, 67, 1124–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provenzano, F.; Deleidi, M. Reassessing Neurodegenerative Disease: Immune Protection Pathways and Antagonistic Pleiotropy. Trends Neurosci. 2021, 44, 771–780. [Google Scholar] [CrossRef]
- Finger, B.C.; Dinan, T.G.; Cryan, J.F. High-Fat Diet Selectively Protects against the Effects of Chronic Social Stress in the Mouse. Neuroscience 2011, 192, 351–360. [Google Scholar] [CrossRef]
- Dornellas, A.P.S.; Boldarine, V.T.; Pedroso, A.P.; Carvalho, L.O.T.; de Andrade, I.S.; Vulcani-Freitas, T.M.; dos Santos, C.C.C.; do Nascimento, C.M.d.P.O.; Oyama, L.M.; Ribeiro, E.B. High-Fat Feeding Improves Anxiety-Type Behavior Induced by Ovariectomy in Rats. Front. Neurosci. 2018, 12, 557. [Google Scholar] [CrossRef] [Green Version]
- Astiz, M.; Pernía, O.; Barrios, V.; Garcia-Segura, L.M.; Diz-Chaves, Y. Short-Term High-Fat Diet Feeding Provides Hypothalamic but Not Hippocampal Protection against Acute Infection in Male Mice. Neuroendocrinology 2016, 104, 40–50. [Google Scholar] [CrossRef]
- Amelianchik, A.; Merkel, J.; Palanisamy, P.; Kaneki, S.; Hyatt, E.; Norris, E.H. The Protective Effect of Early Dietary Fat Consumption on Alzheimer’s Disease–Related Pathology and Cognitive Function in Mice. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2021, 7, e12173. [Google Scholar] [CrossRef] [PubMed]
- Bahr, J.; Klöting, N.; Wilke, B.; Klöting, I.; Follak, N. High-Fat Diet Protects BB/OK Rats from Developing Type 1 Diabetes. Diabetes Metab. Res. Rev. 2011, 27, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Elhaik Goldman, S.; Goez, D.; Last, D.; Naor, S.; Liraz Zaltsman, S.; Sharvit-Ginon, I.; Atrakchi-Baranes, D.; Shemesh, C.; Twitto-Greenberg, R.; Tsach, S.; et al. High-Fat Diet Protects the Blood-Brain Barrier in an Alzheimer’s Disease Mouse Model. Aging Cell 2018, 17, e12818. [Google Scholar] [CrossRef] [PubMed]
- Di Meco, A.; Jelinek, J.; Lauretti, E.; Curtis, M.E.; Issa, J.P.J.; Praticό, D. Gestational High Fat Diet Protects 3xTg Offspring from Memory Impairments, Synaptic Dysfunction, and Brain Pathology. Mol. Psychiatry 2019, 26, 7006–7019. [Google Scholar] [CrossRef]
- Di Meco, A.; Praticò, D. Early-Life Exposure to High-Fat Diet Influences Brain Health in Aging Mice. Aging Cell 2019, 18, e13040. [Google Scholar] [CrossRef] [Green Version]
- Gautier, Y.; Bergeat, D.; Serrand, Y.; Réthoré, N.; Mahérault, M.; Malbert, C.H.; Meurice, P.; Coquery, N.; Moirand, R.; Val-Laillet, D. Western Diet, Obesity and Bariatric Surgery Sequentially Modulated Anxiety, Eating Patterns and Brain Responses to Sucrose in Adult Yucatan Minipigs. Sci. Rep. 2020, 10, 20130. [Google Scholar] [CrossRef]
- Luyer, M.D.; Greve, J.W.M.; Hadfoune, M.; Jacobs, J.A.; Dejong, C.H.; Buurman, W.A. Nutritional Stimulation of Cholecystokinin Receptors Inhibits Inflammation via the Vagus Nerve. J. Exp. Med. 2005, 202, 1023–1029. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.R.; Petrie, J.E.; Shaw, M.W.; Hu, C.; Oakley, C.M.; Woods, S.J.; Patel, B.V.; O’Dea, K.P.; Takata, M. High-Fat Feeding Protects Mice from Ventilator-Induced Lung Injury, via Neutrophil-Independent Mechanisms. Crit. Care Med. 2017, 45, e831–e839. [Google Scholar] [CrossRef] [Green Version]
- Hutagalung, R.; Marques, J.; Kobylka, K.; Zeidan, M.; Kabisch, B.; Brunkhorst, F.; Reinhart, K.; Sakr, Y. The Obesity Paradox in Surgical Intensive Care Unit Patients. Intensive Care Med. 2011, 37, 1793–1799. [Google Scholar] [CrossRef]
- Hogue, C.W.; Stearns, J.D.; Colantuoni, E.; Robinson, K.A.; Stierer, T.; Mitter, N.; Pronovost, P.J.; Needham, D.M. The Impact of Obesity on Outcomes after Critical Illness: A Meta-Analysis. Intensive Care Med. 2009, 35, 1152–1170. [Google Scholar] [CrossRef]
- Wang, Y.; Li, S.; Liu, W.; Wang, F.; Hu, L.F.; Zhong, Z.M.; Wang, H.; Liu, C.F. Vesicular Monoamine Transporter 2 (Vmat2) Knockdown Elicits Anxiety-like Behavior in Zebrafish. Biochem. Biophys. Res. Commun. 2016, 470, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; Tao, S.; Jing Goh, Y.; Chaganty, V.; See, K.; Purushothaman, K.; Orbán, L.; Mathuru, A.S.; Wohland, T.; Winkler, C. A Neurexin2aa Deficiency Results in Axon Pathfinding Defects and Increased Anxiety in Zebrafish. Hum. Mol. Genet. 2020, 29, 3765–3780. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Fei, F.; Sun, S.; Zhang, D.; Dong, Q.; Wang, X.; Wang, L. Increased Anxiety Was Found in Serpini1 Knockout Zebrafish Larval. Biochem. Biophys. Res. Commun. 2021, 534, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Lucon-Xiccato, T.; Tomain, M.; D’Aniello, S.; Bertolucci, C. Bdnf Loss Affects Activity, Sociability, and Anxiety-like Behaviour in Zebrafish. Behav. Brain Res. 2023, 436, 114115. [Google Scholar] [CrossRef] [PubMed]
- Speedie, N.; Gerlai, R. Alarm Substance Induced Behavioral Responses in Zebrafish (Danio rerio). Behav. Brain Res. 2008, 188, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Yan, B.; Bao, M.; Shen, J.; Zheng, P.; Wu, D.; Wang, J.; Li, Z.; Jiang, K. Early Life Exposure to Chronic Unpredictable Stress Induces Anxiety-like Behaviors and Increases the Excitability of Cerebellar Neurons in Zebrafish. Behav. Brain Res. 2023, 437, 114160. [Google Scholar] [CrossRef]
- Doepker, C.; Lieberman, H.R.; Smith, A.P.; Peck, J.D.; El-Sohemy, A.; Welsh, B.T. Caffeine: Friend or Foe? Annu. Rev. Food Sci. Technol. 2016, 7, 117–137. [Google Scholar] [CrossRef]
- El Yacoubi, M.; Ledent, C.; Parmentier, M.; Costentin, J.; Vaugeois, J.M. The Anxiogenic-like Effect of Caffeine in Two Experimental Procedures Measuring Anxiety in the Mouse Is Not Shared by Selective A2A Adenosine Receptor Antagonists. Psychopharmacology 2000, 148, 153–163. [Google Scholar] [CrossRef]
- Acheson, K.J.; Zahorska-Markiewicz, B.; Pittet, P.; Anantharaman, K.; Jéquier, E. Caffeine and Coffee: Their Influence on Metabolic Rate and Substrate Utilization in Normal Weight and Obese Individuals. Am. J. Clin. Nutr. 1980, 33, 989–997. [Google Scholar] [CrossRef] [Green Version]
- Dulloo, A.G. The Search for Compounds That Stimulate Thermogenesis in Obesity Management: From Pharmaceuticals to Functional Food Ingredients. Obes. Rev. 2011, 12, 866–883. [Google Scholar] [CrossRef]
- Zheng, X.; Dai, W.; Chen, X.; Wang, K.; Zhang, W.; Liu, L.; Hou, J. Caffeine Reduces Hepatic Lipid Accumulation through Regulation of Lipogenesis and ER Stress in Zebrafish Larvae. J. Biomed. Sci. 2015, 22, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Oliveira, J.; Silva, P.F.; Luchiari, A.C. Coffee Time: Low Caffeine Dose Promotes Attention and Focus in Zebrafish. Learn. Behav. 2019, 47, 227–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, G.C. Pleiotropy, Natural Selection, and the Evolution of Senescence. Evolution 1957, 11, 398–411. [Google Scholar] [CrossRef]
- Pontzer, H.; Yamada, Y.; Sagayama, H.; Ainslie, P.N.; Andersen, L.F.; Anderson, L.J.; Arab, L.; Baddou, I.; Bedu-Addo, K.; Blaak, E.E.; et al. Daily Energy Expenditure through the Human Life Course. Science 2021, 373, 808–812. [Google Scholar] [CrossRef]
- Misri, S.; Kendrick, K. Treatment of Perinatal Mood and Anxiety Disorders: A Review. Can. J. Psychiatry 2007, 52, 489–498. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| tRF | Sequence | Type | tRNA |
|---|---|---|---|
| tRF-29-JQR7F8YKIRJZ | CATATGGTCTAGCGGTTAGGATTCCTGGT | i-tRF | Glu |
| tRF-30-FP18LPMBQ4NK | AGCAGAGTGGCGCAGCGGAAGCGTGCTGGG | 5′-tRF | Met |
| tRF-30-KQI34JRX6N38 | CCCATATGGTCTAGCGGTTAGGATTCCTGG | i-tRF | Glu |
| tRF-30-KY7343RX6NMH | CCCTGGTGGTCTAGTGGTTAGGATTCGGCG | i-tRF | Glu |
| tRF-30-PIR8YP9LON4V | GCACTGGTGGTTCAGTGGTAGAATTCTCGC | 5′-tRF | Val |
| tRF-31-86V8WPMN1E8Y0 | TCCCATATGGTCTAGCGGTTAGGATTCCTGG | 5′-tRF | Glu |
| tRF-31-87R8WP9I1EWJ0 | TCCCTGGTGGTCTAGTGGCTAGGATTCGGCG | 5′-tRF | Glu |
| tRF-31-87R8WP9N1EWJ0 | TCCCTGGTGGTCTAGTGGTTAGGATTCGGCG | 5′-tRF | Glu |
| tRF-31-PSQP4PW3FJI0B | GCCCGGCTAGCTCAGTCGGTAGAGCATGAGA | 5′-tRF | Lys |
| tRF-31-PW5SVP9N15WV0 | GCCGTGATCGTATAGTGGTTAGTACTCTGCG | 5′-tRF | His |
| tRF-32-86V8WPMN1E8YN | TCCCATATGGTCTAGCGGTTAGGATTCCTGGT | 5′-half | Glu |
| tRF-32-87R8WP9N1EWJM | TCCCTGGTGGTCTAGTGGTTAGGATTCGGCGC | 5′-half | Glu |
| tRF-32-PSQP4PW3FJI01 | GCCCGGCTAGCTCAGTCGGTAGAGCATGAGAC | 5′-half | Lys |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yehuda, H.; Madrer, N.; Goldberg, D.; Soreq, H.; Meerson, A. Inversely Regulated Inflammation-Related Processes Mediate Anxiety–Obesity Links in Zebrafish Larvae and Adults. Cells 2023, 12, 1794. https://doi.org/10.3390/cells12131794
Yehuda H, Madrer N, Goldberg D, Soreq H, Meerson A. Inversely Regulated Inflammation-Related Processes Mediate Anxiety–Obesity Links in Zebrafish Larvae and Adults. Cells. 2023; 12(13):1794. https://doi.org/10.3390/cells12131794
Chicago/Turabian StyleYehuda, Hila, Nimrod Madrer, Doron Goldberg, Hermona Soreq, and Ari Meerson. 2023. "Inversely Regulated Inflammation-Related Processes Mediate Anxiety–Obesity Links in Zebrafish Larvae and Adults" Cells 12, no. 13: 1794. https://doi.org/10.3390/cells12131794
APA StyleYehuda, H., Madrer, N., Goldberg, D., Soreq, H., & Meerson, A. (2023). Inversely Regulated Inflammation-Related Processes Mediate Anxiety–Obesity Links in Zebrafish Larvae and Adults. Cells, 12(13), 1794. https://doi.org/10.3390/cells12131794