
Intermittent Exposure to a Single Bottle of Ethanol Modulates Stress Sensitivity: Impact of Age at Exposure Initiation

 , ,
, ,
Abstract
:1. Key Findings
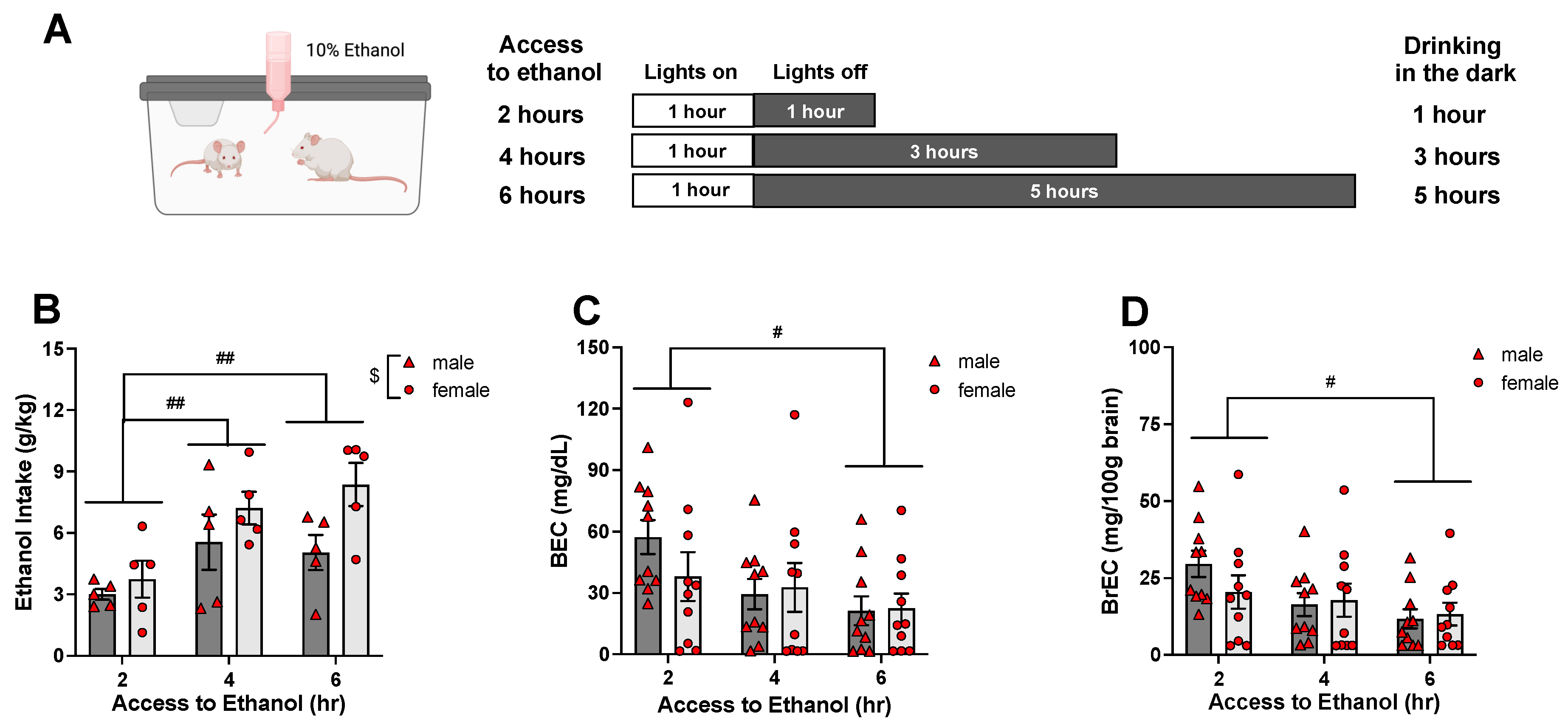
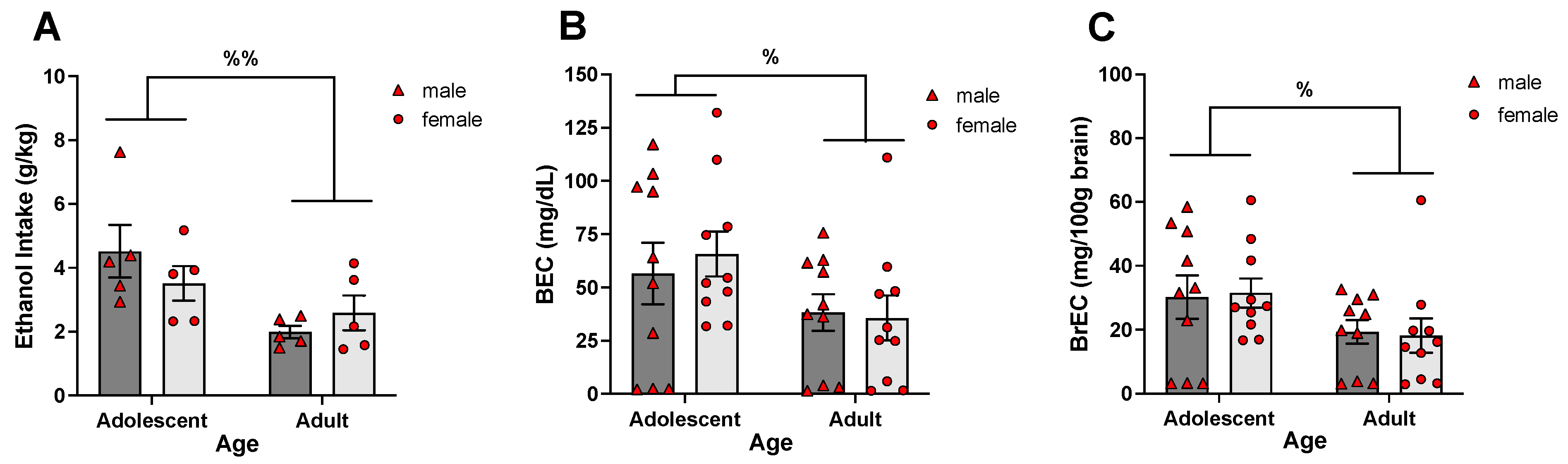
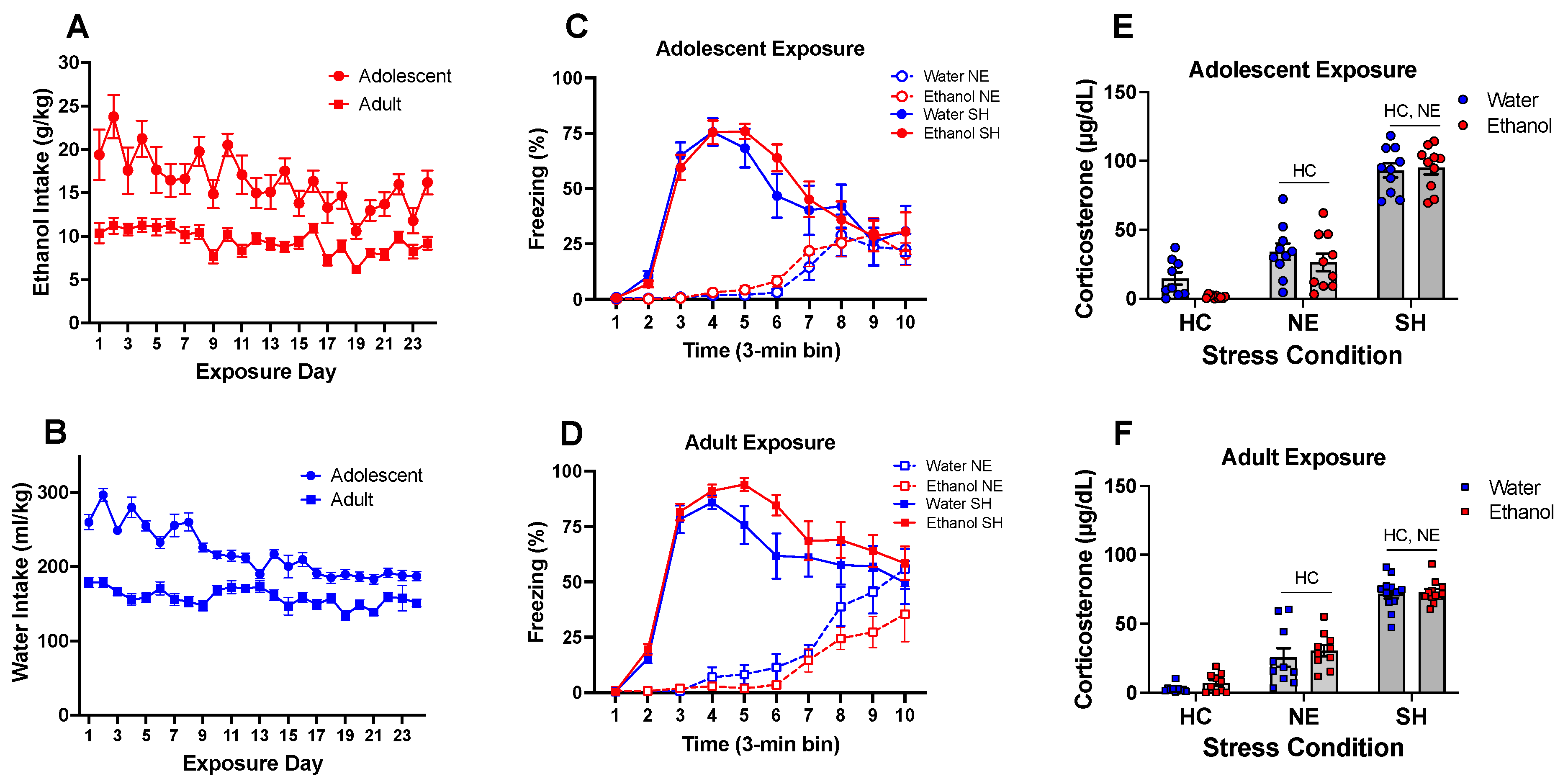
- The present experiments revealed age differences in ethanol intake during exposure to a single bottle of 10% ethanol solution as the only liquid available, with adolescent rats ingesting more ethanol than adults and demonstrating higher blood and brain ethanol concentrations.
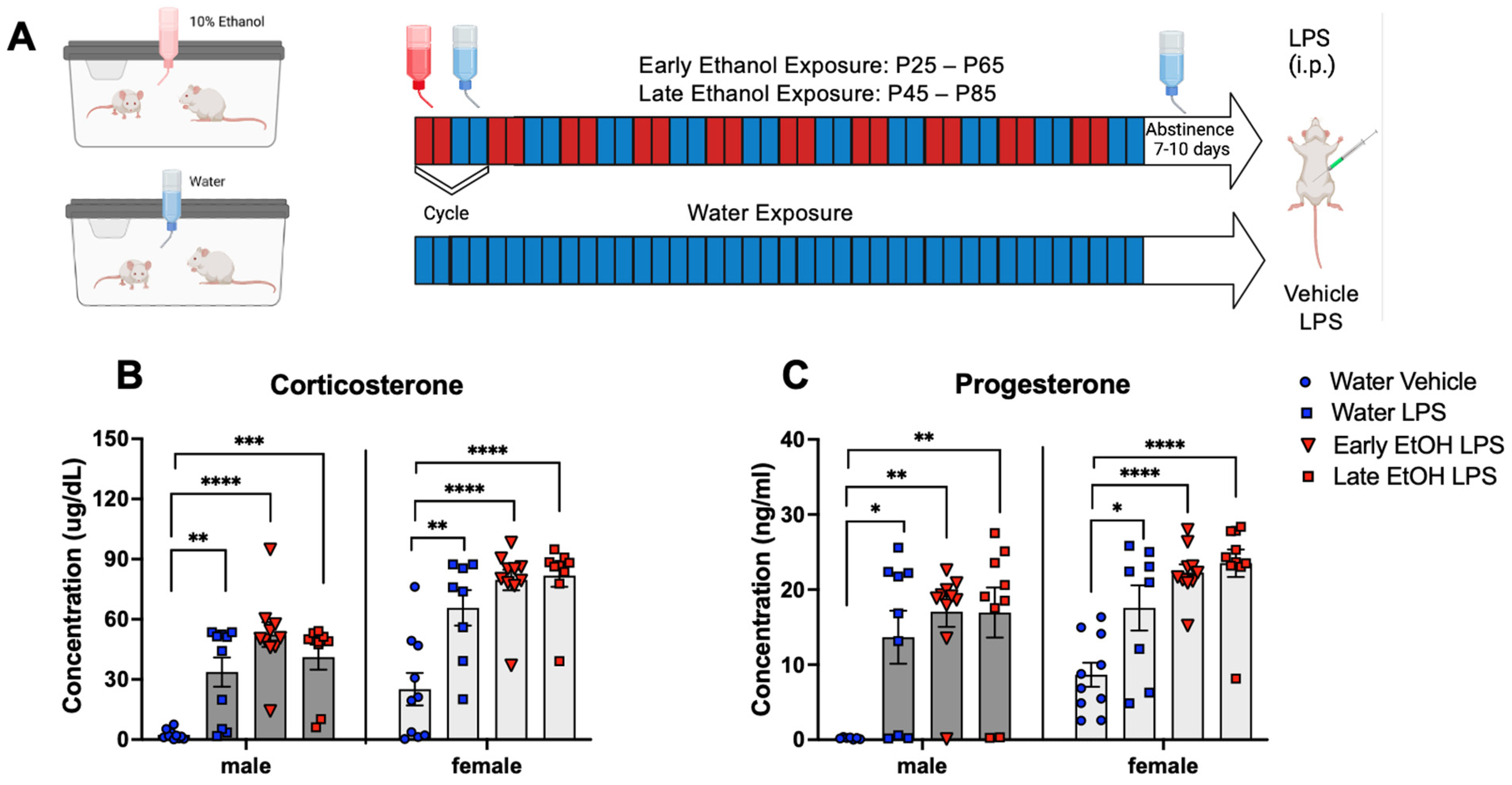
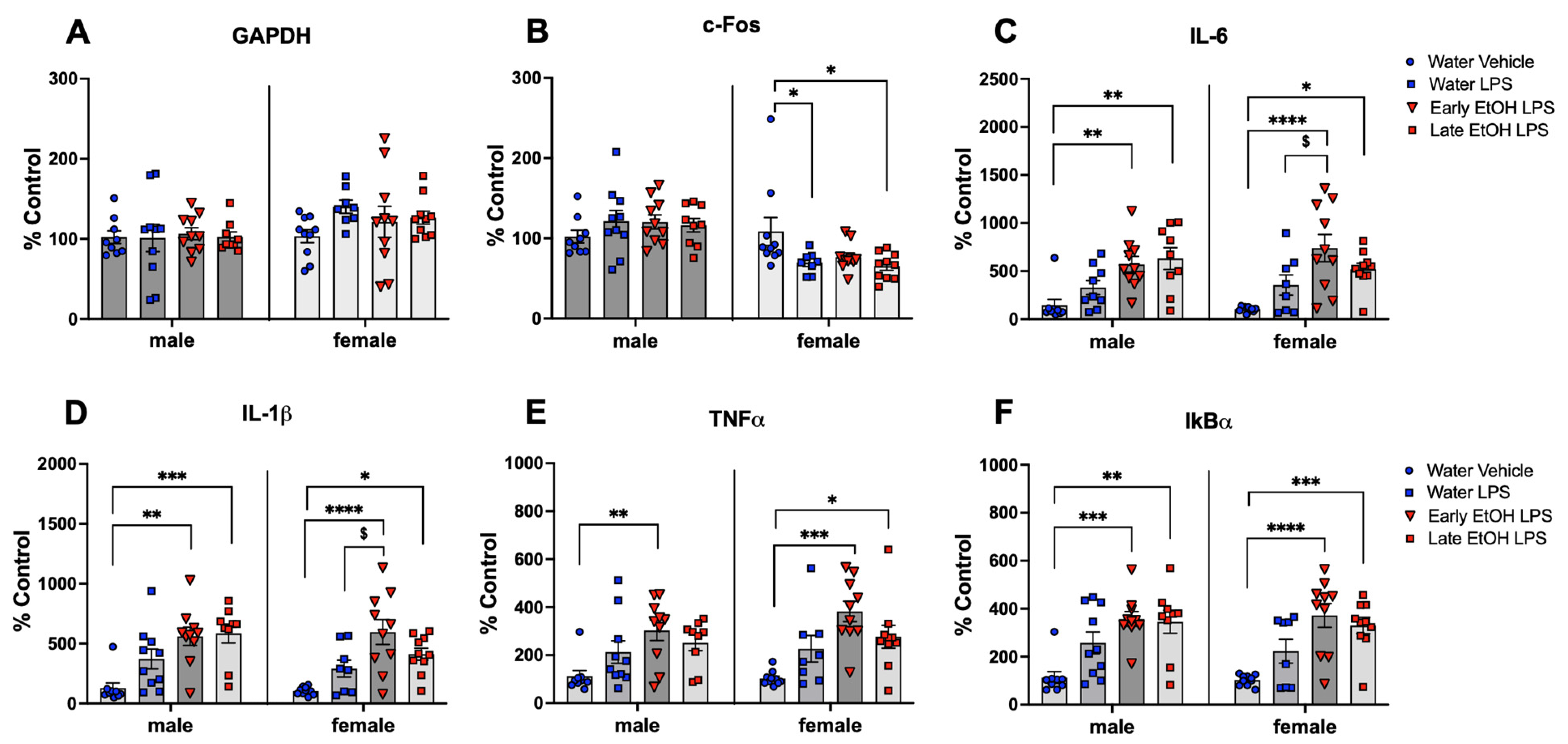
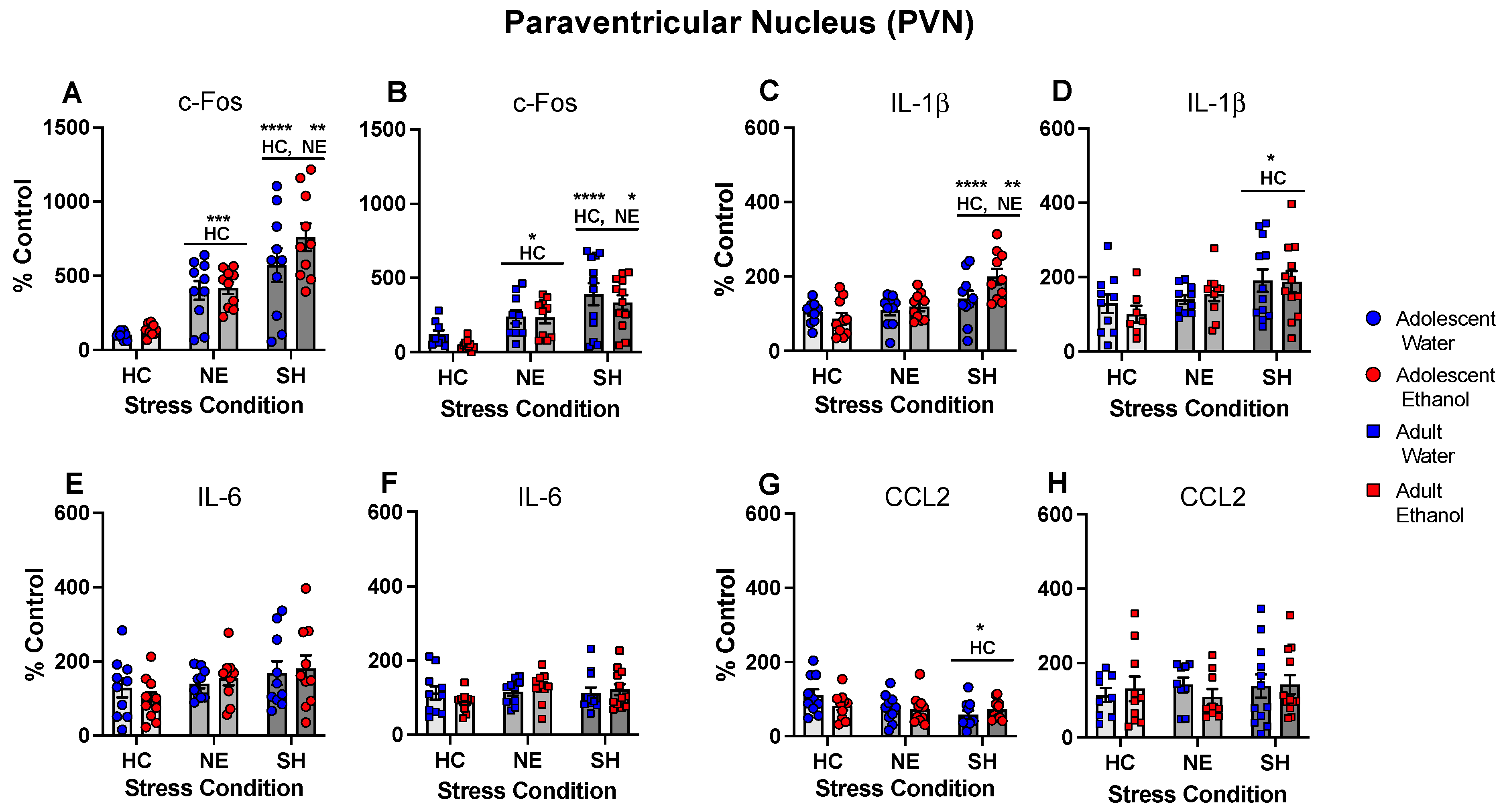
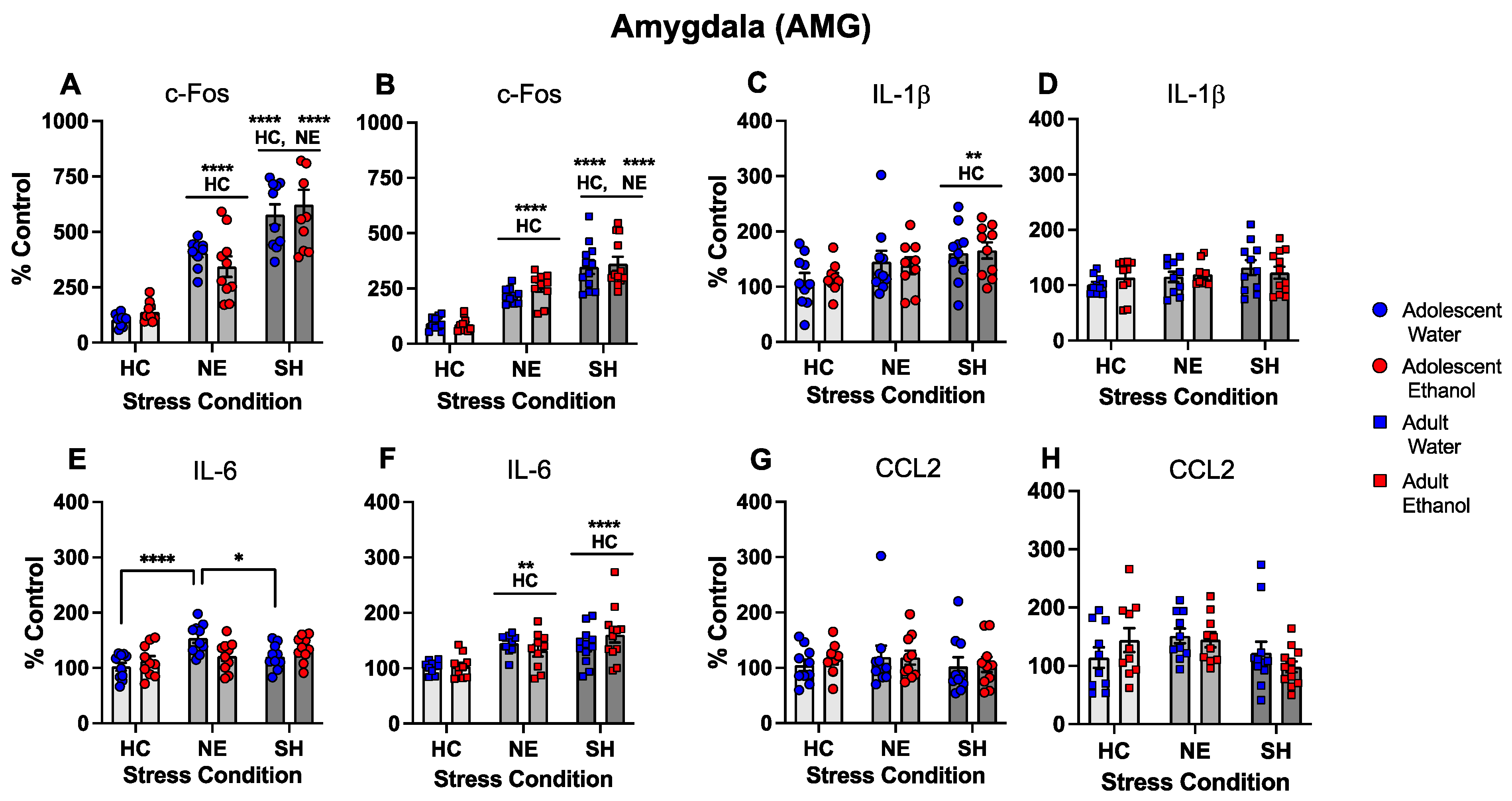
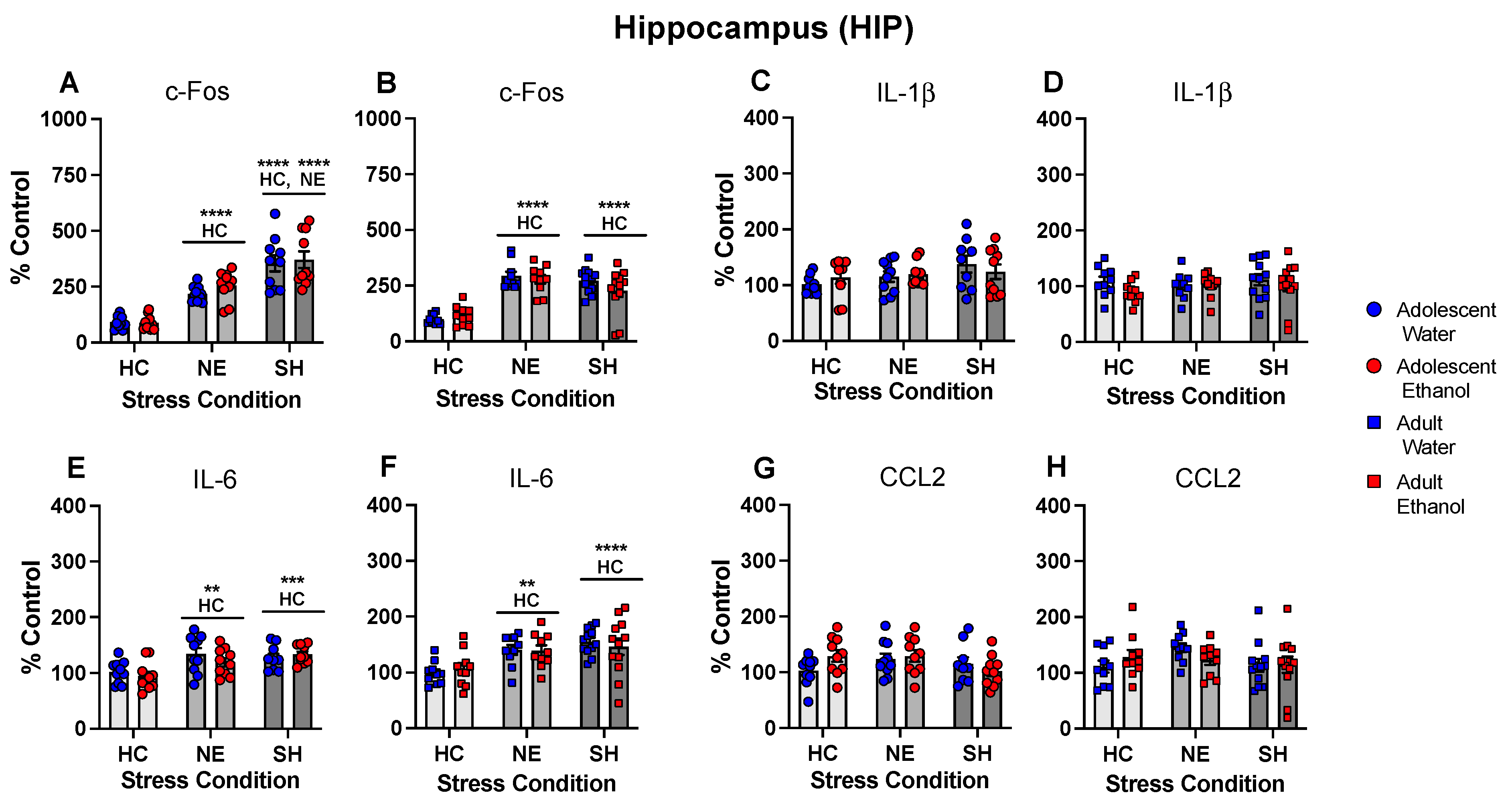
- Chronic intermittent ethanol exposure using a single bottle of 10% ethanol solution as the only liquid available that was initiated either in early or late adolescence enhanced cytokine expression to lipopolysaccharide (LPS) challenge, with this effect being more evident in females following early ethanol exposure.
- Chronic intermittent ethanol exposure of females to a single bottle of 10% ethanol initiated in adulthood exacerbated behavioral signs of distress (freezing), an effect that was not evident after adolescent ethanol consumption.
2. Introduction
3. Materials and Methods
3.1. General Methods
3.1.1. Subjects
3.1.2. A Single Bottle Chronic Intermittent Ethanol Exposure Paradigm
3.2. LPS Challenge
3.3. Stress Challenge
3.4. Tissue Collection
3.5. Corticosterone and Progesterone Analysis
3.6. Reverse-Transcriptase Polymerase Chain Reaction
3.7. Blood and Brain Ethanol Concentrations
3.8. Statistical Analysis
3.9. Experimental Designs and Procedures
4. Results
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Substance Abuse and Mental Health Services Administration. Key Substance Use and Mental Health Indicators in the United States: Results from the 2021 National Survey on Drug Use and Health; HHS Publication No. PEP22-07-01-005, NSDUH Series H-57; Center for Behavioral Health Statistics and Quality, Substance Abuse and Mental Health Services Administration: North Bethesda, MD, USA, 2022. Available online: https://www.samhsa.gov/data/report/2021-nsduh-annual-national-report (accessed on 5 May 2023).
- Miech, R.A.; Johnston, L.D.; Patrick, M.E.; O’Malley, P.M.; Bachman, J.G.; Schulenberg, J.E. Monitoring the Future National Survey Results on Drug Use, 1975–2022: Secondary School Students; Institute for Social Research, The University of Michigan: Ann Arbor, MI, USA, 2023; Available online: http://monitoringthefuture.org/results/publications/monographs/ (accessed on 5 May 2023).
- Dawson, D.A.; Goldstein, R.B.; Patricia Chou, S.; June Ruan, W.; Grant, B.F. Age at First Drink and the First Incidence of Adult-Onset DSM-IV Alcohol Use Disorders. Alcohol. Clin. Exp. Res. 2008, 32, 2149–2160. [Google Scholar] [CrossRef] [PubMed]
- Warner, L.A.; White, H.R.; Johnson, V. Alcohol initiation experiences and family history of alcoholism as predictors of problem-drinking trajectories. J. Stud. Alcohol Drugs 2007, 68, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Chung, T.; Creswell, K.G.; Bachrach, R.; Clark, D.B.; Martin, C.S. Adolescent Binge Drinking. Alcohol Res. 2018, 39, 5–15. [Google Scholar] [PubMed]
- Mehus, C.J.; Patrick, M.E. Alcohol use among 10th-graders: Distinguishing between high-intensity drinking and other levels of use. J. Adolesc. 2020, 83, 27–30. [Google Scholar] [CrossRef]
- Lees, B.; Meredith, L.R.; Kirkland, A.E.; Bryant, B.E.; Squeglia, L.M. Effect of alcohol use on the adolescent brain and behavior. Pharmacol. Biochem. Behav. 2020, 192, 172906. [Google Scholar] [CrossRef]
- Tetteh-Quarshie, S.; Risher, M.L. Adolescent brain maturation and the neuropathological effects of binge drinking: A critical review. Front. Neurosci. 2023, 16, 1040049. [Google Scholar] [CrossRef]
- Spear, L.P. Effects of adolescent alcohol consumption on the brain and behavior. Nat. Rev. Neurosci. 2018, 19, 197–214. [Google Scholar] [CrossRef]
- Squeglia, L.M.; Jacobus, J.; Tapert, S.F. The effect of alcohol use on human adolescent brain structures and systems. Handb. Clin. Neurol. 2014, 125, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Ewing, S.W.; Sakhardande, A.; Blakemore, S.J. The effect of alcohol consumption on the adolescent brain: A systematic review of MRI and fMRI studies of alcohol-using youth. Neuroimage Clin. 2014, 5, 420–437. [Google Scholar] [CrossRef] [Green Version]
- Scaife, J.C.; Duka, T. Behavioural measures of frontal lobe function in a population of young social drinkers with binge drinking pattern. Pharmacol. Biochem. Behav. 2009, 93, 354–362. [Google Scholar] [CrossRef]
- Erickson, E.K.; Grantham, E.K.; Warden, A.S.; Harris, R.A. Neuroimmune signaling in alcohol use disorder. Pharmacol. Biochem. Behav. 2019, 177, 34–60. [Google Scholar] [CrossRef] [PubMed]
- Deak, T.; Kelliher, K.T.; Wojcik, H.J.; Gano, A. Prenatal and adolescent alcohol exposure programs immunity across the lifespan: CNS-mediated regulation. Pharmacol. Biochem. Behav. 2022, 216, 173390. [Google Scholar] [CrossRef] [PubMed]
- Crews, F.T.; Robinson, D.L.; Chandler, L.J.; Ehlers, C.L.; Mulholland, P.J.; Pandey, S.C.; Rodd, Z.A.; Spear, L.P.; Swartzwelder, H.S.; Vetreno, R.P. Mechanisms of Persistent Neurobiological Changes Following Adolescent Alcohol Exposure: NADIA Consortium Findings. Alcohol. Clin. Exp. Res. 2019, 43, 1806–1822. [Google Scholar] [CrossRef] [Green Version]
- Doremus-Fitzwater, T.L.; Deak, T. Adolescent neuroimmune function and its interaction with alcohol. Int. Rev. Neurobiol. 2021, 161, 167–208. [Google Scholar]
- Hiller-Sturmhöfel, S.; Spear, L.P. Binge Drinking’s Effects on the Developing Brain-Animal Models. Alcohol Res. 2018, 39, 77–86. [Google Scholar]
- Spear, L.P. The adolescent brain and age-related behavioral manifestations. Neurosci. Biobehav. Rev. 2000, 24, 417–463. [Google Scholar] [CrossRef]
- Spear, L.P. Rewards, aversions and affect in adolescence: Emerging convergences across laboratory animal and human data. Dev. Cogn. Neurosci. 2011, 1, 392–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doremus-Fitzwater, T.L.; Varlinskaya, E.I.; Spear, L.P. Motivational systems in adolescence: Possible implications for age differences in substance abuse and other risk-taking behaviors. Brain Cogn. 2010, 72, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spear, L.P. Adolescent alcohol exposure: Are there separable vulnerable periods within adolescence? Physiol. Behav. 2015, 148, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Vetter-O’Hagen, C.S.; Spear, L.P. Hormonal and physical markers of puberty and their relationship to adolescent-typical novelty-directed behavior. Dev. Psychobiol. 2012, 54, 523–535. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, G.M.; Lew, B.J.; Vedder, L.C.; Savage, L.M. Chronic intermittent ethanol exposure leads to alterations in brain-derived neurotrophic factor within the frontal cortex and impaired behavioral flexibility in both adolescent and adult rats. Neuroscience 2017, 348, 324–334. [Google Scholar] [CrossRef] [Green Version]
- Gass, J.T.; Glen, W.B., Jr.; McGonigal, J.T.; Trantham-Davidson, H.; Lopez, M.F.; Randall, P.K.; Yaxley, R.; Floresco, S.B.; Chandler, L.J. Adolescent alcohol exposure reduces behavioral flexibility, promotes disinhibition, and increases resistance to extinction of ethanol self-administration in adulthood. Neuropsychopharmacology 2014, 39, 2570–2583. [Google Scholar] [CrossRef] [PubMed]
- Sey, N.Y.; Gómez-A, A.; Madayag, A.C.; Boettiger, C.A.; Robinson, D.L. Adolescent intermittent ethanol impairs behavioral flexibility in a rat foraging task in adulthood. Behav. Brain Res. 2019, 373, 112085. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.M.; Oliveri, A.N.; Zhang, C.; Frazier, J.M.; Mackinnon, S.; Cole, G.J.; Levin, E.D. Long-term behavioral impairment following acute embryonic ethanol exposure in zebrafish. Neurotoxicol. Teratol. 2015, 48, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokare, D.M.; Kyzar, E.J.; Zhang, H.; Sakharkar, A.J.; Pandey, S.C. Adolescent alcohol exposure-induced changes in alpha-melanocyte stimulating hormone and neuropeptide Y pathways via histone acetylation in the brain during adulthood. Int. J. Neuropsychopharmacol. 2017, 20, 758–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varlinskaya, E.I.; Johnson, J.M.; Przybysz, K.R.; Deak, T.; Diaz, M.R. Adolescent forced swim stress increases social anxiety-like behaviors and alters kappa opioid receptor function in the basolateral amygdala of male rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 98, 109812. [Google Scholar] [CrossRef]
- Vetreno, R.P.; Lawrimore, C.J.; Rowsey, P.J.; Crews, F.T. Persistent Adult Neuroimmune Activation and Loss of Hippocampal Neurogenesis Following Adolescent Ethanol Exposure: Blockade by Exercise and the Anti-inflammatory Drug Indomethacin. Front. Neurosci. 2018, 12, 200. [Google Scholar] [CrossRef] [Green Version]
- Coleman, L.G., Jr.; Liu, W.; Oguz, I.; Styner, M.; Crews, F.T. Adolescent binge ethanol treatment alters adult brain regional volumes, cortical extracellular matrix protein and behavioral flexibility. Pharmacol. Biochem. Behav. 2014, 116, 142–151. [Google Scholar] [CrossRef] [Green Version]
- Coleman, L.G., Jr.; Crews, F.T.; Vetreno, R.P. The persistent impact of adolescent binge alcohol on adult brain structural, cellular, and behavioral pathology: A role for the neuroimmune system and epigenetics. Int. Rev. Neurobiol. 2021, 160, 1–44. [Google Scholar] [CrossRef]
- Guerri, C.; Pascual, M. Impact of neuroimmune activation induced by alcohol or drug abuse on adolescent brain development. Int. J. Dev. Neurosci. 2019, 77, 89–98. [Google Scholar] [CrossRef]
- Guo, Y.; Yan, M.; Li, L.; Zhao, L.; Li, Y. Treadmill Exercise Prevents Cognitive Impairments in Adolescent Intermittent Ethanol Rats by Reducing the Excessive Activation of Microglia Cell in the Hippocampus. Int. J. Mol. Sci. 2022, 23, 14701. [Google Scholar] [CrossRef]
- Vetreno, R.P.; Patel, Y.; Patel, U.; Walter, T.J.; Crews, F.T. Adolescent intermittent ethanol reduces serotonin expression in the adult raphe nucleus and upregulates innate immune expression that is prevented by exercise. Brain Behav. Immun. 2017, 60, 333–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vore, A.S.; Barney, T.M.; Gano, A.; Varlinskaya, E.I.; Deak, T. Adolescent intermittent ethanol (AIE) produces sex specific alterations in adult neuroimmune gene expression and ethanol sensitivity that are independent of ethanol metabolism. Neuropharmacology 2021, 195, 108635. [Google Scholar] [CrossRef] [PubMed]
- Gano, A.; Vore, A.S.; Sammakia, M.N.; Deak, T. Assessment of Extracellular Cytokines in the Hippocampus of the Awake Behaving Rat Using Large-Molecule Microdialysis Combined with Multiplex Arrays after Acute and Chronic Ethanol Exposure. Alcohol. Clin. Exp. Res. 2019, 43, 640–654. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.L.; Amodeo, L.R.; Chandler, L.J.; Crews, F.T.; Ehlers, C.L.; Gómez-A, A.; Healey, K.L.; Kuhn, C.M.; Macht, V.A.; Marshall, S.A.; et al. The role of sex in the persistent effects of adolescent alcohol exposure on behavior and neurobiology in rodents. Int. Rev. Neurobiol. 2021, 160, 305–340. [Google Scholar] [CrossRef] [PubMed]
- Pascual, M.; Montesinos, J.; Marcos, M.; Torres, J.-L.; Costa-Alba, P.; García-García, F.; Laso, F.-J.; Guerri, C. Gender differences in the inflammatory cytokine and chemokine profiles induced by binge ethanol drinking in adolescence. Addict. Biol. 2017, 22, 1829–1841. [Google Scholar] [CrossRef] [PubMed]
- Spear, L.P. Consequences of adolescent use of alcohol and other drugs: Studies using rodent models. Neurosci. Biobehav. Rev. 2016, 70, 228–243. [Google Scholar] [CrossRef] [Green Version]
- Shultz, J.; Weiner, H.; Westcott, J. Retardation of ethanol absorption by food in the stomach. J. Stud. Alcohol 1980, 41, 861–870. [Google Scholar] [CrossRef]
- In, H.S.; Kim, D.W.; Park, Y.M.; Kim, B. Experimental intraperitoneal injection of alcohol in rats: Peritoneal findings and histopathology. Toxicol. Rep. 2014, 1, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Glover, E.J.; Khan, F.; Clayton-Stiglbauer, K.; Chandler, L.J. Impact of sex, strain, and age on blood ethanol concentration and behavioral signs of intoxication during ethanol vapor exposure. Neuropharmacology 2021, 184, 108393. [Google Scholar] [CrossRef]
- Jeanblanc, J.; Rolland, B.; Gierski, F.; Martinetti, M.P.; Naassila, M. Animal models of binge drinking, current challenges to improve face validity. Neurosci. Biobehav. Rev. 2019, 106, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, G.M.; Stewart, W.N.; Savage, L.M. Chronic drinking during adolescence predisposes the adult rat for continued heavy drinking: Neurotrophin and behavioral adaptation after long-term, continuous ethanol exposure. PLoS ONE 2016, 11, e0149987. [Google Scholar] [CrossRef] [Green Version]
- Spear, L.P. Timing eclipses amount: The critical importance of intermittency in alcohol exposure effects. Alcohol. Clin. Exp. Res. 2020, 44, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Marsland, P.; Vore, A.S.; DaPrano, E.; Paluch, J.M.; Blackwell, A.A.; Varlinskaya, E.I.; Deak, T. Sex-specific effects of ethanol consumption in older Fischer 344 rats on microglial dynamics and Aβ (1-42) accumulation. Alcohol 2023, 107, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Vore, A.S.; Doremus-Fitzwater, T.; Gano, A.; Deak, T. Adolescent Ethanol Exposure Leads to Stimulus-Specific Changes in Cytokine Reactivity and Hypothalamic-Pituitary-Adrenal Axis Sensitivity in Adulthood. Front. Behav. Neurosci. 2017, 11, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsland, P.; Parrella, A.; Vore, A.S.; Barney, T.M.; Varlinskaya, E.I.; Deak, T. Male, but not female, Sprague Dawley rats display enhanced fear learning following acute ethanol withdrawal (hangover). Pharmacol. Biochem. Behav. 2021, 208, 173229. [Google Scholar] [CrossRef]
- Doremus-Fitzwater, T.L.; Gano, A.; Paniccia, J.E.; Deak, T. Male adolescent rats display blunted cytokine responses in the CNS after acute ethanol or lipopolysaccharide exposure. Physiol. Behav. 2015, 148, 131–144. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Doremus, T.L.; Brunell, S.C.; Rajendran, P.; Spear, L.P. Factors influencing elevated ethanol consumption in adolescent relative to adult rats. Alcohol. Clin. Exp. Res. 2005, 29, 1796–1808. [Google Scholar] [CrossRef]
- Vetter-O’Hagen, C.; Varlinskaya, E.; Spear, L. Sex differences in ethanol intake and sensitivity to aversive effects during adolescence and adulthood. Alcohol Alcohol. 2009, 44, 547–554. [Google Scholar] [CrossRef]
- Lara Aparicio, S.Y.; Laureani Fierro, Á.d.J.; Aranda Abreu, G.E.; Toledo Cárdenas, R.; García Hernández, L.I.; Coria Ávila, G.A.; Rojas Durán, F.; Aguilar, M.E.H.; Manzo Denes, J.; Chi-Castañeda, L.D.; et al. Current opinion on the use of c-Fos in neuroscience. NeuroSci 2022, 3, 687–702. [Google Scholar] [CrossRef]
- Vallés, S.L.; Blanco, A.M.; Pascual, M.; Guerri, C. Chronic ethanol treatment enhances inflammatory mediators and cell death in the brain and in astrocytes. Brain Pathol. 2004, 14, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Marsland, P.; Parrella, A.; Orlofsky, M.; Lovelock, D.F.; Vore, A.S.; Varlinskaya, E.I.; Deak, T. Neuroendocrine and neuroimmune responses in male and female rats: Evidence for functional immaturity of the neuroimmune system during early adolescence. Eur. J. Neurosci. 2022, 55, 2311–2325. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.; Gregor, D.; Peng, Z.; Li, J.; Bekker, A.; Ye, J. Chronic intermittent voluntary alcohol drinking induces hyperalgesia in Sprague-Dawley rats. Int. J. Physiol. Pathophysiol. Pharmacol. 2015, 7, 136. [Google Scholar]
- Holgate, J.Y.; Shariff, M.; Mu, E.W.; Bartlett, S. A rat drinking in the dark model for studying ethanol and sucrose consumption. Front. Behav. Neurosci. 2017, 11, 29. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.W.; Jönsson, K.A. Food-induced lowering of blood-ethanol profiles and increased rate of elimination immediately after a meal. J. Forensic Sci. 1994, 39, 1084–1093. [Google Scholar] [CrossRef]
- Lumeng, L.; Bosron, W.F.; Li, T.K. Quantitative correlation of ethanol elimination rates in vivo with liver alcohol dehydrogenase activities in fed, fasted and food-restricted rats. Biochem Pharmacol. 1979, 28, 1547–1551. [Google Scholar] [CrossRef]
- Spiteri, N.J. Circadian patterning of feeding, drinking and activity during diurnal food access in rats. Physiol. Behav. 1982, 28, 139–147. [Google Scholar] [CrossRef]
- Criado, J.R.; Wills, D.N.; Walker, B.M.; Ehlers, C.L. Effects of adolescent ethanol exposure on sleep in adult rats. Alcohol 2008, 42, 631–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerma-Cabrera, J.M.; Carvajal, F.; Alcaraz-Iborra, M.; de la Fuente, L.; Navarro, M.; Thiele, T.E.; Cubero, I. Adolescent binge-like ethanol exposure reduces basal α-MSH expression in the hypothalamus and the amygdala of adult rats. Pharmacol. Biochem. Behav. 2013, 110, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Broadwater, M.A.; Liu, W.; Crews, F.T.; Spear, L.P. Persistent loss of hippocampal neurogenesis and increased cell death following adolescent, but not adult, chronic ethanol exposure. Dev. Neurosci. 2014, 36, 297–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing protein abundance and mRNA expression levels on a genomic scale. Genome Biol. 2003, 4, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Bonilla, A.; Richardson, H.N. Sex Differences in the Neurobiology of Alcohol Use Disorder. Alcohol Res. 2020, 40, 4. [Google Scholar] [CrossRef] [PubMed]
- Sawchenko, P.E.; Li, H.Y.; Ericsson, A. Circuits and mechanisms governing hypothalamic responses to stress: A tale of two paradigms. Prog. Brain Res. 2000, 122, 61–78. [Google Scholar] [PubMed]
- Buck, H.M.; Hueston, C.M.; Bishop, C.; Deak, T. Enhancement of the hypothalamic–pituitary–adrenal axis but not cytokine responses to stress challenges imposed during withdrawal from acute alcohol exposure in Sprague–Dawley rats. Psychopharmacology 2011, 218, 203–215. [Google Scholar] [CrossRef] [Green Version]
- Blair, R.S.; Acca, G.M.; Tsao, B.; Stevens, N.; Maren, S.; Nagaya, N. Estrous cycle contributes to state-dependent contextual fear in female rats. Psychoneuroendocrinology 2022, 141, 105776. [Google Scholar] [CrossRef]
- Criado, J.R.; Ehlers, C.L. Effects of adolescent onset voluntary drinking followed by ethanol vapor exposure on subsequent ethanol consumption during protracted withdrawal in adult Wistar rats. Pharmacol. Biochem. Behav. 2013, 103, 622–630. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.U.; Varlinskaya, E.I.; Dannenhoffer, C.A.; Spear, L.P. Adolescent intermittent ethanol exposure: Effects on pubertal development, novelty seeking, and social interaction in adulthood. Alcohol 2019, 75, 19–29. [Google Scholar] [CrossRef]
- Kasten, C.R.; Carzoli, K.L.; Sharfman, N.M.; Henderson, T.; Holmgren, E.B.; Lerner, M.R.; Miller, M.C.; Wills, T.A. Adolescent alcohol exposure produces sex differences in negative affect-like behavior and group I mGluR BNST plasticity. Neuropsychopharmacology 2020, 45, 1306–1315. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Experimental Condition | Ethanol Intake (g/kg) | |
|---|---|---|---|
| Male | Female | ||
| Experiment 1 | 2 h access on Day 2 | 6.41 ± 2.52 | 11.54 ± 2.66 |
| 4 h access on Day 2 | 10.07 ± 2.06 | 13.15 ± 2.69 | |
| 6 h access on Day 2 | 6.09 ± 1.01 | 16.87 ± 4.88 | |
| Experiment 2 | Adolescent | 13.34 ± 1.50 | 14.66 ± 2.64 |
| Adult | 5.22 ± 0.53 | 7.99 ± 0.70 | |
| Cycle | Adolescent Exposure Initiation | Adult Exposure Initiation | ||
|---|---|---|---|---|
| Water Mean ± SEM (g) | Ethanol Mean ± SEM (g) | Water Mean ± SEM (g) | Ethanol Mean ± SEM (g) | |
| 1 | 109.07 ± 1.78 | 106.37 ± 1.84 | 244.91 ± 3.39 | 238.63 ± 2.87 |
| 2 | 132.27 ± 1.73 | 127.87 ± 1.92 | 252.53 ± 3.59 | 245.50 ± 3.00 |
| 3 | 151.03 ± 1.78 | 146.80 ± 1.90 | 257.47 ± 4.47 | 249.53 ± 3.45 |
| 4 | 165.37 ± 1.74 | 161.00 ± 1.94 | 257.69 ± 3.46 | 249.25 ± 3.37 |
| 5 | 179.00 ± 1.87 | 172.67 ± 2.00 | 258.66 ± 3.81 | 251.78 ± 3.02 |
| 6 | 190.37 ± 2.27 | 184.60 ± 1.99 | 262.72 ± 4.06 | 258.72 ± 3.09 |
| 7 | 201.03 ± 2.27 | 195.90 ± 1.94 | 264.03 ± 4.07 | 261.19 ± 3.75 |
| 8 | 209.33 ± 2.53 | 207.03 ± 2.11 | 266.78 ± 3.55 | 259.67 ± 2.91 |
| 9 | 217.67 ± 2.45 | 213.20 ± 1.90 | 267.38 ± 3.52 | 259.98 ± 3.12 |
| 10 | 225.20 ± 2.81 | 221.53 ± 1.94 | 269.78 ± 3.58 | 262.87 ± 2.92 |
| 11 | 230.80 ± 3.24 | 225.9 ± 2.01 | 271.25 ± 3.60 | 263.53 ± 3.03 |
| 12 | 235.30 ± 30.5 | 231.23 ± 2.19 | 271.50 ± 3.56 | 263.59 ± 3.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marsland, P.; Trapp, S.; Vore, A.; Lutzke, A.; Varlinskaya, E.I.; Deak, T. Intermittent Exposure to a Single Bottle of Ethanol Modulates Stress Sensitivity: Impact of Age at Exposure Initiation. Cells 2023, 12, 1991. https://doi.org/10.3390/cells12151991
Marsland P, Trapp S, Vore A, Lutzke A, Varlinskaya EI, Deak T. Intermittent Exposure to a Single Bottle of Ethanol Modulates Stress Sensitivity: Impact of Age at Exposure Initiation. Cells. 2023; 12(15):1991. https://doi.org/10.3390/cells12151991
Chicago/Turabian StyleMarsland, Paige, Sarah Trapp, Andrew Vore, Ashley Lutzke, Elena I. Varlinskaya, and Terrence Deak. 2023. "Intermittent Exposure to a Single Bottle of Ethanol Modulates Stress Sensitivity: Impact of Age at Exposure Initiation" Cells 12, no. 15: 1991. https://doi.org/10.3390/cells12151991
APA StyleMarsland, P., Trapp, S., Vore, A., Lutzke, A., Varlinskaya, E. I., & Deak, T. (2023). Intermittent Exposure to a Single Bottle of Ethanol Modulates Stress Sensitivity: Impact of Age at Exposure Initiation. Cells, 12(15), 1991. https://doi.org/10.3390/cells12151991